Нуклеин қышқылдары мен амин қышқылдарының биосинтезі. Нуклеин қышқылдарының құрылысы мен атқаратын қызметі презентация
- Нуклеин қышқылдары мен амин қышқылдарының биосинтезі. Нуклеин қышқылдарының құрылысы мен атқаратын қызметі

Содержание
- 2. 1868 ж. Швейцария биологы Эридрих Мишер ірің клеткаларының ядросынан құрамында фосфоры бар зат бөліп алған. Ядродан
- 3. ДНҚ геннің материалдық негізі, ол геннің келесі қасиеттеріне ие болады: 1. Өз-өзін дәл қайталау, соның арқасында
- 4. ДНҚ генетикалық ақпаратты негізгі жеткізуші, себебі: Бір ағзаның әртүрлі ұлпасынан алынған ДНҚ нуклеотид құрамы бірдей. Әртүрлі
- 5. Уотсон мен Крик моделінің ерекшілігі: Репликация мәселесін, яғни тұқым қуалаушылықтың белгісін сипаттауы. ДНҚ синтезін көрсететін реакция:
- 6. ДНҚ синтезіне қатысатын ферменттер: 1. ДНҚ тәуелді ДНҚ полимеразасы (I, II,III). ДНҚ полимеразасы элонгацияға тікелей байланысты.
- 7. ДНҚ лигазасы үзіктерді өзара жалғастырады. Жапон ғалымы Оказаки ДНҚ молекласының қысқа фрагментте синтезделетінін тапты. Фрагмент 3/(ОН)
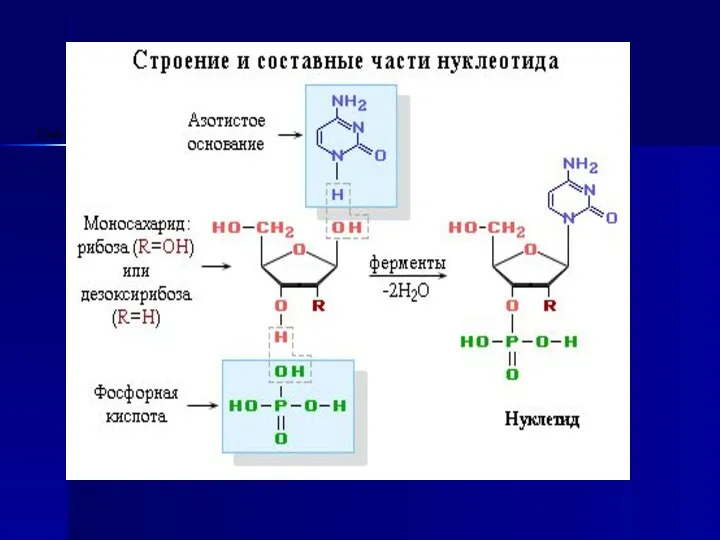
- 8. Нуклеин қышқылдары - нуклеотид полимерлерінен құралған. Нуклеотидтердің химиялық құрамы - пиримидиндік немесе пуриндік негіздер, пентоз, фосфор
- 10. Дезоксирибоза Рибоза
- 13. Нуклеин қышқылдарының бірінші реттік құрылымы: мононуклеотидтер бір-бірімен «3́-5́́ байланыс» арқылы қосылады, мұнда фосфор қышқылының қалдығы 5′-фосфатты
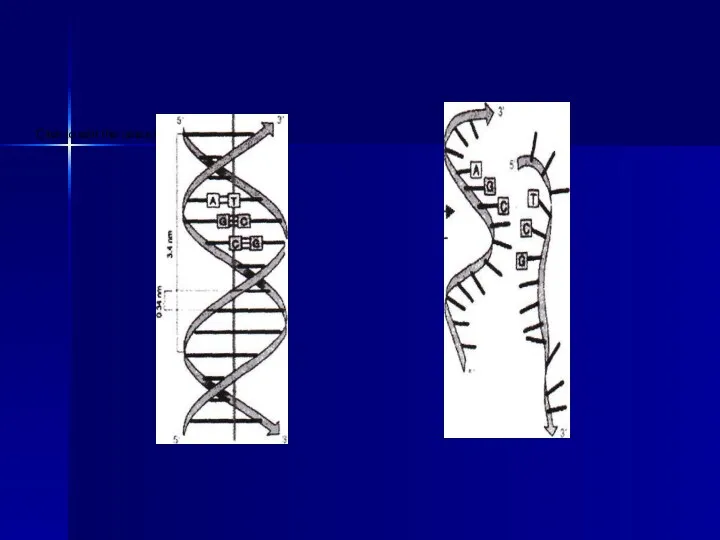
- 14. Америка ғалымдары Дж.Уотсон және ағылшын физигі Фрэнсис Крик (1953 ж.) ұсынған ДНҚ молекуласының екіншілік құрылымы қос
- 15. Бір тізбектегі нуклеотидтің орналасуы өз бетінмен еркінірек болса да, екінші тізбектегі нуклеотидтердің орны бірінші тізбектің нуклеотидтік
- 16. Әрбір тізбек төрт нуклеотидтің: аденин = тимин, цитозин ≡ гуанин байланыс арқылы байланысады. ДНҚ –ның екіншілік
- 17. Генетикалық кодтың құрылысы Генетикалық кодтың касиеттері: триплеттік (әрбір амин қышқылы үш нуклеотидпен кодталады); барлық тірі ағзаларда
- 18. Маңызды қызметі: триплеттілігі, биологиялық кодта код саны - 3. үш нуклеотидтік қалдықтан бір амин қышкылы қолданылады.
- 19. ДНҚ синтезіне қатысатын негізгі фермент - ДНҚ-на тәуелді ДНҚ полимераза (I,II,III). ДНҚ молекуласының синтезіне қажетті негізгі
- 20. ДНҚ синтезіне қатысатын ферменттер: 1. ДНҚ-на тәуелді ДНҚ-полимеразасы (I, П, III). III ДНҚ-полимеразасы элонгацияға тікелей қатысатын
- 21. ДНҚ-ның биосинтезі үш кезеңнен түрады: 1. Инициация. ДНҚ-на тәуелді РНҚ-полимеразасы жұмыс істейді, сннтездің басы. ДНҚ-ны екі
- 22. РНҚ биосинтезі - транскрипция. РНҚ -ның биосинтезі немесе ақуыз синтезінің бірінші кезеңі, яғни ДНҚ- дағы генетикалық
- 24. Ақуыз синтезінің «Фабрикасы»
- 25. Ақуыз биосинтезінің тізбегі
- 28. Ақуыз биосинтезі бірнеше сатыдан тұрады. Амин қышқылдарының белсендірілуі. Гиалоплазмаға жасушаралық сұйықтықтан диффузия және осмос нәтижесінде амин
- 29. Екінші сатысында комплекс түзіледі: активтелген аминқышқылы қосылыстан өзіне сәйкес ферментімен цитоплазманың тРНҚ тасымалданады. Үрдіс аминоацил -
- 30. Ақуыздардың синтезделуі екі кезеңнен тұрады: Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос тізбегінін біреу комплементарлы көшірмесі
- 31. Трансляцияның өзі 5 кезеңнен түрад: Трансляцияның 1-ші кезеңі: амин қышқылдарының белсендендірілуі өтеді, оған 20 амин қышқылы,
- 32. Трансляцияның 2-ші кезеңінде полипептидтік тізбектің инициациясы жалғасады, оған а-РНҚ, ақуыз синтезін бастаушы кодон (АУК) қажет. Трансляцияның
- 33. Трансляцияның 4 -ші кезеңі - Терминация яғни синтездің аяқталу кезеңі, оған: АТФ; ақуыз синтезінің біткенін білдіруші
- 34. Трансляцияның 5-ші кезеңі - кеңістіктегі полипептидтік тізбектің орналасуы және процессинг. Бұл кезеңде полипептид өзінің кеңістіктегі екінші
- 36. Скачать презентацию

1868 ж. Швейцария биологы Эридрих Мишер ірің клеткаларының ядросынан құрамында фосфоры
1868 ж. Швейцария биологы Эридрих Мишер ірің клеткаларының ядросынан құрамында фосфоры

ДНҚ геннің материалдық негізі, ол геннің келесі қасиеттеріне ие болады:
1. Өз-өзін
ДНҚ геннің материалдық негізі, ол геннің келесі қасиеттеріне ие болады:
1. Өз-өзін

ДНҚ генетикалық ақпаратты негізгі жеткізуші, себебі:
Бір ағзаның әртүрлі ұлпасынан алынған ДНҚ
ДНҚ генетикалық ақпаратты негізгі жеткізуші, себебі:
Бір ағзаның әртүрлі ұлпасынан алынған ДНҚ

Уотсон мен Крик моделінің ерекшілігі:
Репликация мәселесін, яғни тұқым қуалаушылықтың белгісін сипаттауы.
Уотсон мен Крик моделінің ерекшілігі:
Репликация мәселесін, яғни тұқым қуалаушылықтың белгісін сипаттауы.

ДНҚ синтезіне қатысатын ферменттер:
1. ДНҚ тәуелді ДНҚ полимеразасы (I, II,III). ДНҚ
ДНҚ синтезіне қатысатын ферменттер:
1. ДНҚ тәуелді ДНҚ полимеразасы (I, II,III). ДНҚ

ДНҚ лигазасы үзіктерді өзара жалғастырады. Жапон ғалымы Оказаки ДНҚ молекласының қысқа
ДНҚ лигазасы үзіктерді өзара жалғастырады. Жапон ғалымы Оказаки ДНҚ молекласының қысқа

Нуклеин қышқылдары - нуклеотид полимерлерінен құралған.
Нуклеотидтердің химиялық құрамы - пиримидиндік
Нуклеин қышқылдары - нуклеотид полимерлерінен құралған.
Нуклеотидтердің химиялық құрамы - пиримидиндік
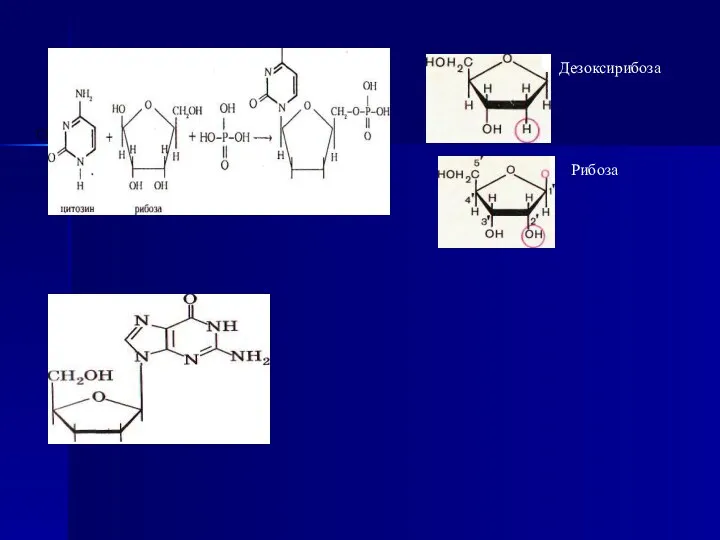
Дезоксирибоза
Рибоза
Дезоксирибоза
Рибоза


Нуклеин қышқылдарының бірінші реттік құрылымы: мононуклеотидтер бір-бірімен «3́-5́́ байланыс» арқылы қосылады,
Нуклеин қышқылдарының бірінші реттік құрылымы: мононуклеотидтер бір-бірімен «3́-5́́ байланыс» арқылы қосылады,

Америка ғалымдары Дж.Уотсон және ағылшын физигі Фрэнсис Крик (1953 ж.) ұсынған
Америка ғалымдары Дж.Уотсон және ағылшын физигі Фрэнсис Крик (1953 ж.) ұсынған

Бір тізбектегі нуклеотидтің орналасуы өз бетінмен еркінірек болса да, екінші тізбектегі
Бір тізбектегі нуклеотидтің орналасуы өз бетінмен еркінірек болса да, екінші тізбектегі
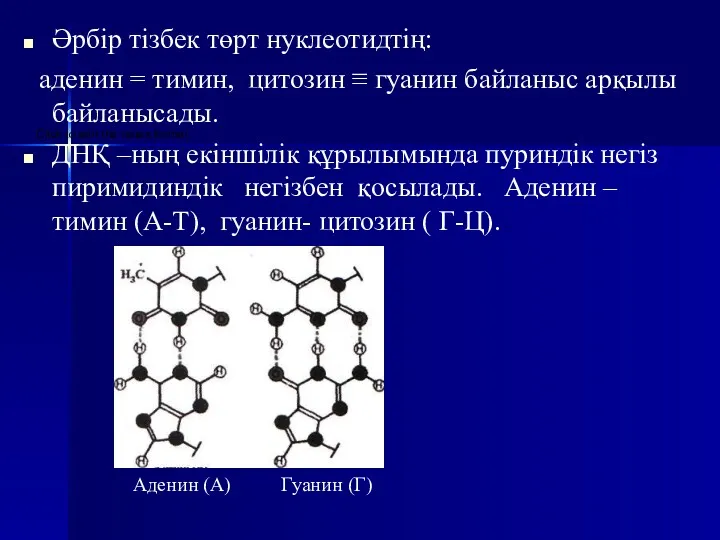
Әрбір тізбек төрт нуклеотидтің:
аденин = тимин, цитозин ≡ гуанин байланыс
Әрбір тізбек төрт нуклеотидтің:
аденин = тимин, цитозин ≡ гуанин байланыс

Генетикалық кодтың құрылысы
Генетикалық кодтың касиеттері: триплеттік (әрбір амин қышқылы үш
Генетикалық кодтың құрылысы
Генетикалық кодтың касиеттері: триплеттік (әрбір амин қышқылы үш

Маңызды қызметі: триплеттілігі, биологиялық кодта код саны - 3.
үш нуклеотидтік
Маңызды қызметі: триплеттілігі, биологиялық кодта код саны - 3.
үш нуклеотидтік

ДНҚ синтезіне қатысатын негізгі фермент - ДНҚ-на тәуелді ДНҚ полимераза (I,II,III).
ДНҚ
ДНҚ синтезіне қатысатын негізгі фермент - ДНҚ-на тәуелді ДНҚ полимераза (I,II,III).
ДНҚ

ДНҚ синтезіне қатысатын ферменттер:
1. ДНҚ-на тәуелді ДНҚ-полимеразасы (I, П, III).
ДНҚ синтезіне қатысатын ферменттер:
1. ДНҚ-на тәуелді ДНҚ-полимеразасы (I, П, III).

ДНҚ-ның биосинтезі үш кезеңнен түрады:
1. Инициация. ДНҚ-на тәуелді РНҚ-полимеразасы жұмыс
ДНҚ-ның биосинтезі үш кезеңнен түрады:
1. Инициация. ДНҚ-на тәуелді РНҚ-полимеразасы жұмыс

РНҚ биосинтезі - транскрипция. РНҚ -ның биосинтезі немесе ақуыз синтезінің бірінші
РНҚ биосинтезі - транскрипция. РНҚ -ның биосинтезі немесе ақуыз синтезінің бірінші

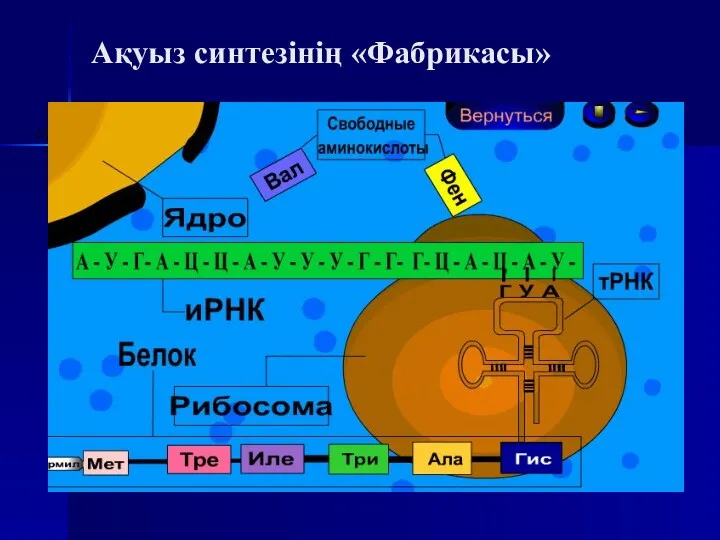
Ақуыз синтезінің «Фабрикасы»
Ақуыз синтезінің «Фабрикасы»
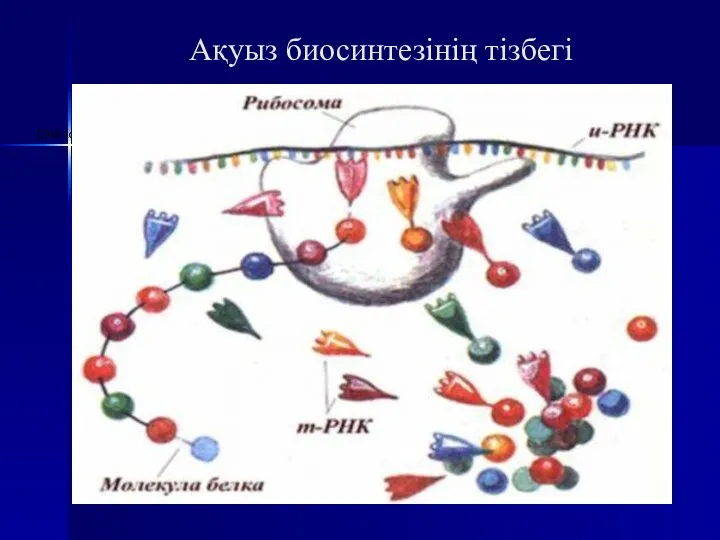
Ақуыз биосинтезінің тізбегі
Ақуыз биосинтезінің тізбегі
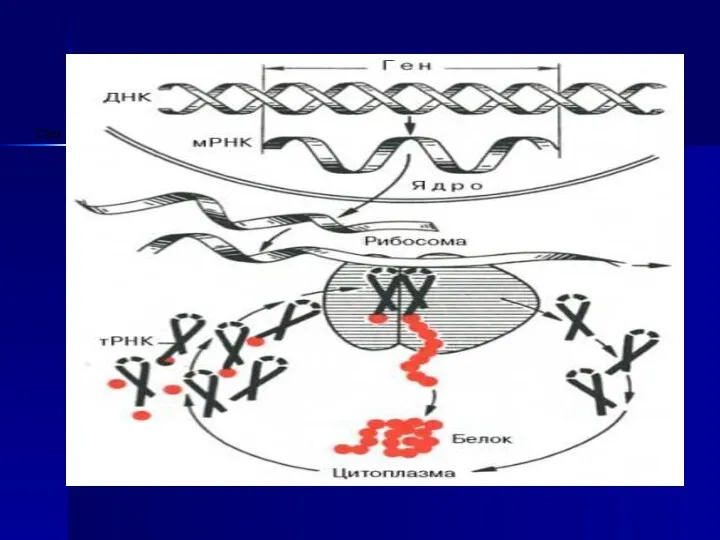
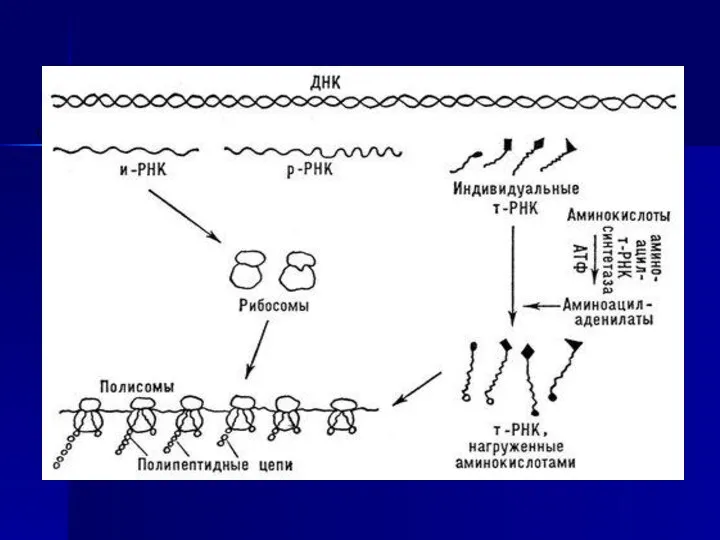

Ақуыз биосинтезі бірнеше сатыдан тұрады.
Амин қышқылдарының белсендірілуі.
Гиалоплазмаға жасушаралық сұйықтықтан
Амин қышқылдарының белсендірілуі.
Гиалоплазмаға жасушаралық сұйықтықтан

Екінші сатысында комплекс түзіледі: активтелген аминқышқылы қосылыстан өзіне сәйкес ферментімен цитоплазманың
Екінші сатысында комплекс түзіледі: активтелген аминқышқылы қосылыстан өзіне сәйкес ферментімен цитоплазманың

Ақуыздардың синтезделуі екі кезеңнен тұрады:
Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос
Ақуыздардың синтезделуі екі кезеңнен тұрады:
Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос

Трансляцияның өзі 5 кезеңнен түрад:
Трансляцияның 1-ші кезеңі: амин қышқылдарының белсендендірілуі өтеді,
Трансляцияның өзі 5 кезеңнен түрад:
Трансляцияның 1-ші кезеңі: амин қышқылдарының белсендендірілуі өтеді,

Трансляцияның 2-ші кезеңінде полипептидтік тізбектің инициациясы жалғасады, оған а-РНҚ, ақуыз синтезін
Трансляцияның 2-ші кезеңінде полипептидтік тізбектің инициациясы жалғасады, оған а-РНҚ, ақуыз синтезін

Трансляцияның 4 -ші кезеңі - Терминация яғни синтездің аяқталу кезеңі, оған:
Трансляцияның 4 -ші кезеңі - Терминация яғни синтездің аяқталу кезеңі, оған:

Трансляцияның 5-ші кезеңі - кеңістіктегі полипептидтік тізбектің орналасуы және процессинг. Бұл
Трансляцияның 5-ші кезеңі - кеңістіктегі полипептидтік тізбектің орналасуы және процессинг. Бұл
 Анализ генной экспрессии на модели Mycoplasma gallisepticum
Анализ генной экспрессии на модели Mycoplasma gallisepticum Обмен веществ и энергии. Терморегуляция
Обмен веществ и энергии. Терморегуляция Растения и животные разных материков
Растения и животные разных материков Факторы эволюции. Приспособленность. Критерии вида
Факторы эволюции. Приспособленность. Критерии вида Функции печени
Функции печени Рост и развитие растительного организма
Рост и развитие растительного организма Анатомия и физиология дыхательной системы
Анатомия и физиология дыхательной системы Белки
Белки Микрофлора полости рта
Микрофлора полости рта конспект урока по биологии
конспект урока по биологии Фотосинтез. Суммарная реакция фотосинтеза
Фотосинтез. Суммарная реакция фотосинтеза Бесполое размножение
Бесполое размножение Отряд насекомоядные
Отряд насекомоядные Скворцы - вестники весны
Скворцы - вестники весны Всероссийская проверочная работа по биологии
Всероссийская проверочная работа по биологии Сады средневековья
Сады средневековья Кіт Лісовий
Кіт Лісовий Буындар биомеханикасы
Буындар биомеханикасы Бесполое и половое размножение
Бесполое и половое размножение Дикие и домашние животные
Дикие и домашние животные Класс Земноводные
Класс Земноводные Факторы эволюции. Приспособленность. Критерии вида
Факторы эволюции. Приспособленность. Критерии вида презентация Вред курения
презентация Вред курения Болезни томатов и меры борьбы с ними
Болезни томатов и меры борьбы с ними Иммуноферментный анализ
Иммуноферментный анализ Биологические основы паразитизма
Биологические основы паразитизма Плесневые грибы
Плесневые грибы Пчелы
Пчелы