- Нуклеосомный уровень

Содержание
- 2. Нуклеосомный уровень Наименьшей структурно-функциональной единицей хромосом является – нуклеосома. Спираль ДНК соединяется с группами из восьми
- 3. Нуклеосомный кор
- 5. Между нуклеосомами имеются соединяющие их участки ДНК, содержащие около 60 пар нуклеотидов. Их называют спейсерными или
- 6. Нуклеосомный уровень
- 7. нуклеосомная нить
- 8. Нуклеосомная нить (электронная микроскопия)
- 9. При развертывании нуклеосомы весь хроматин становится активным. Дисковидные нуклеосомы имеют диаметр 10 нм, высоту 5 нм.
- 10. Второй: нуклеомерный уровень укладки хроматина этот уровень укладки ДНК. осуществляется за счёт гистоновых белков и образуется
- 11. Нуклесомный уровень дает формирование «цепочки из бусинок». Далее 6 нуклеосом сближаются и соединяются посредством гистонового белка
- 12. Хромомерный уровень. Дальнейшая упаковка хроматина дает формирование гетерохроматина. Все остальные уровни компактизации связаны с укладкой хроматиновых
- 14. Дальнейшая упаковка хромосом-хромонемная происходит сближение хромомер и образуются толстые нити, которые становятся видны в световом микроскопе.
- 15. И последний уровень структурной организации хроматина – пятый - хроматидный . Хромонемы укладываются спирально или петлеобразно,
- 16. ХРОМОСОМНЫЙ Метафазная хромосома состоит из двух хроматид, соединенных первичной перетяжкой – центромерой. Таким образом, в результате
- 17. ХРОМОСОМА К началу XX в. углубленное изучение поведения этих структур в ходе самовоспроизведения клеток, при созревании
- 18. Строение метафазной хромосомы Термин хромосома был предложен в 1888 г. немецким морфологом В. Вальдейером, который применил
- 19. Хромосомы всех эукариотических клеток построены по одному плану. Они включают в себя три основных компонента: короткое
- 20. Строение хромосомы
- 21. Центромера делит хромосому на два плеча: Хромосомы с равными плечами называют равноплечими или метацентрическими, с плечами
- 22. Кинетохоры Кинетохоры– это сложные комплексы, состоящие из многих белков. Морфологически они очень сходны, имеют одинаковое строение,
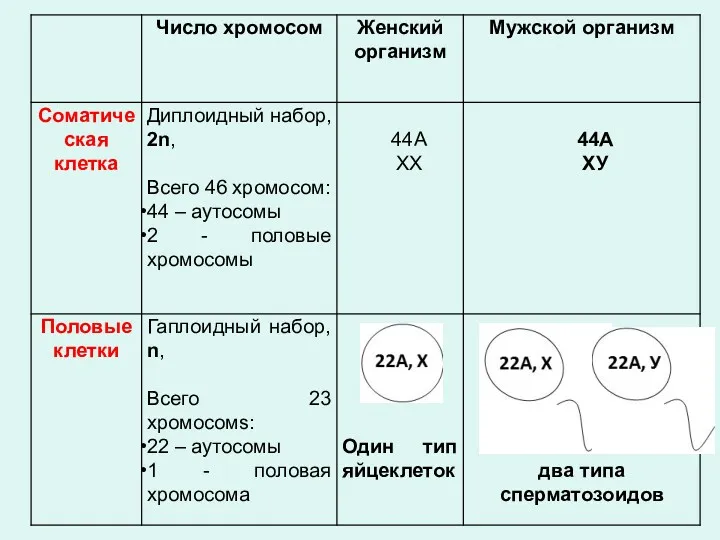
- 23. Хромосомы подразделяются на аутосомы (одинаковые у обоих полов). и гетеросомы, или половые хромосомы (разные для мужских
- 24. кариотип Кариотип — диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецифическим признаком и
- 25. Совокупность числа и морфологии хромосом данного вида называется - КАРИОТИП
- 27. Классификация хромосом
- 28. Парижская классификация В основе Парижской классификации хромосом человека (1971 г.) лежат методы специальной дифференциальной их окраски,
- 29. Денверская классификация
- 30. К группе А относят 1—3 пары хромосом. Это cамые крупные, метацентрические и субметацентрические хромосомы, их центромерный
- 31. Группа D (13—15 пары). Хромосомы акроцентрические, ЦИ 15. Размеры хромосом около 4,2 мкм. Группа Е (16—18
- 32. 11-я хромосома человека HBB — β-субъединица гемоглобина 16-я хромосома человека 2-я хромосома человека
- 33. Геном человека Вся масса ДНК гаплоидной клетки – называется геномом. Структуру и функции генома изучает, специальная
- 35. Скачать презентацию

Нуклеосомный уровень
Наименьшей структурно-функциональной единицей хромосом является – нуклеосома.
Спираль ДНК соединяется с
Нуклеосомный уровень
Наименьшей структурно-функциональной единицей хромосом является – нуклеосома.
Спираль ДНК соединяется с
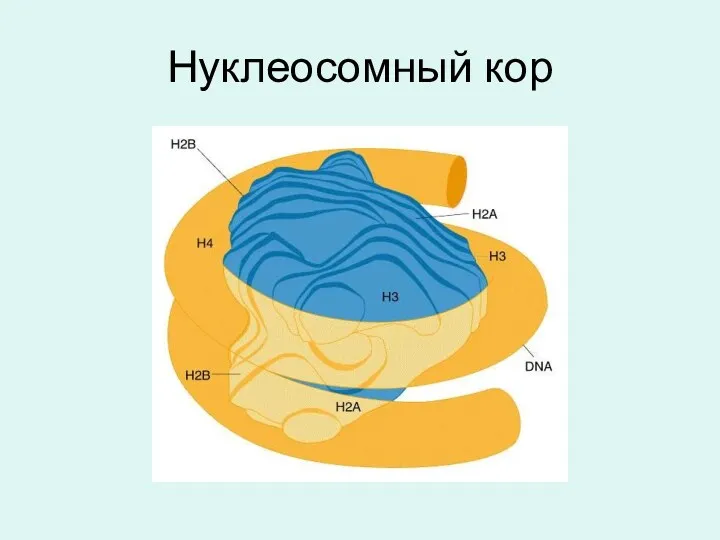
Нуклеосомный кор
Нуклеосомный кор
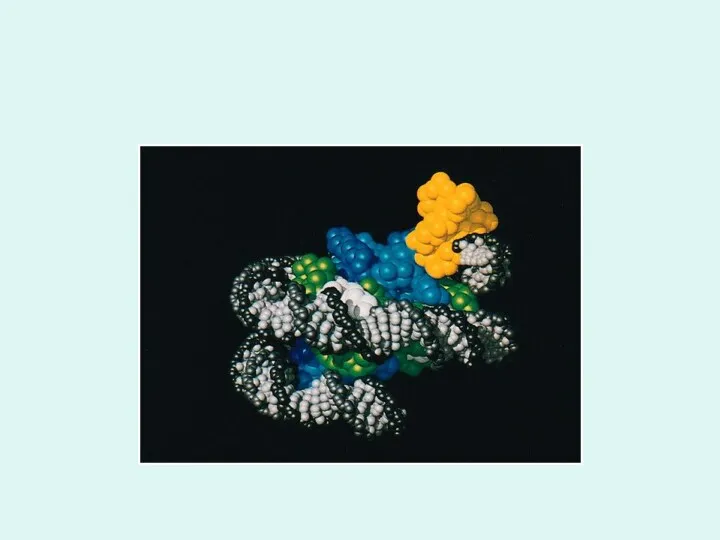

Между нуклеосомами имеются соединяющие их участки ДНК, содержащие около 60 пар
Между нуклеосомами имеются соединяющие их участки ДНК, содержащие около 60 пар
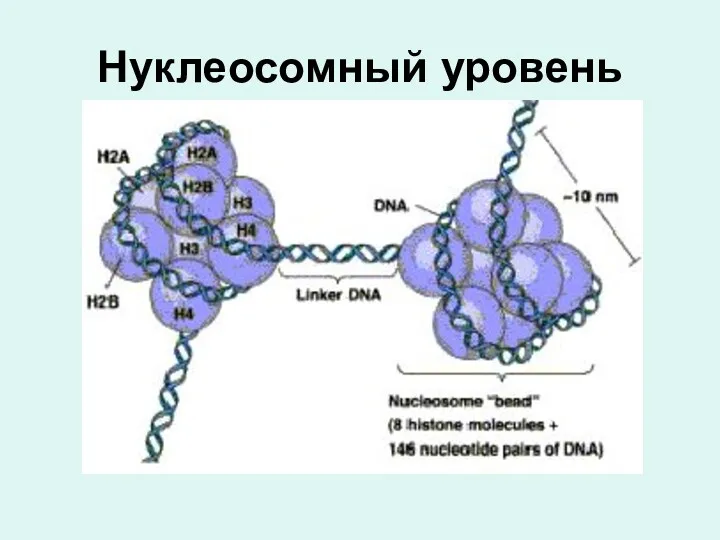
Нуклеосомный уровень
Нуклеосомный уровень
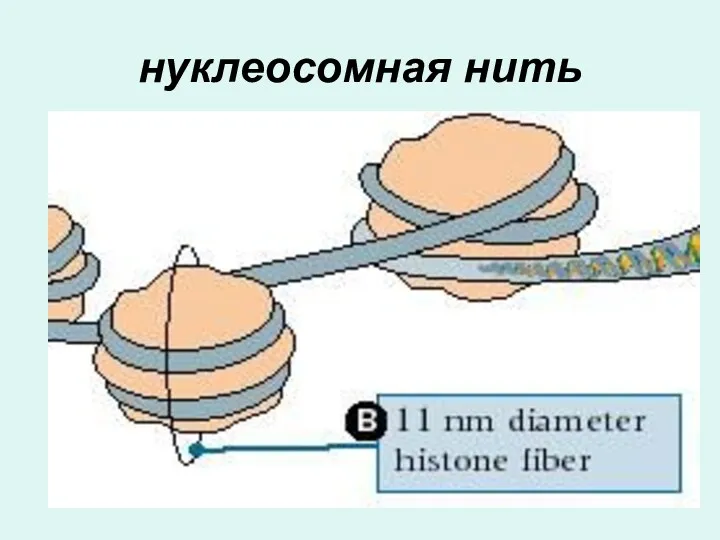
нуклеосомная нить
нуклеосомная нить
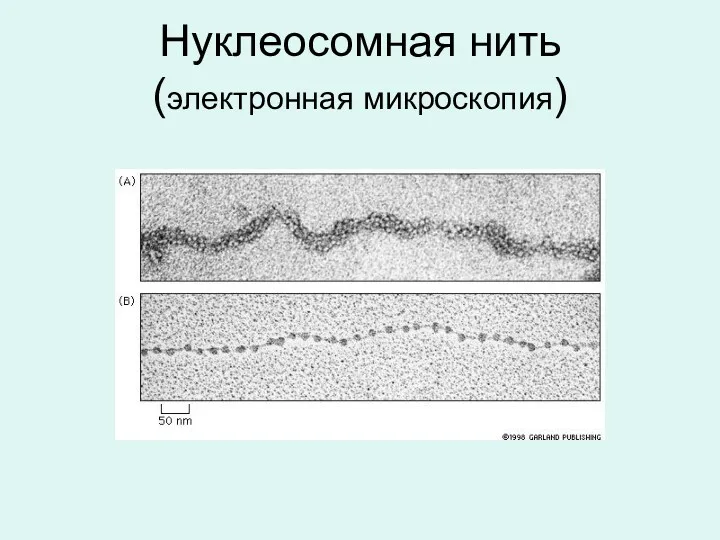
Нуклеосомная нить
(электронная микроскопия)
Нуклеосомная нить
(электронная микроскопия)

При развертывании нуклеосомы весь хроматин становится активным.
Дисковидные нуклеосомы имеют диаметр
При развертывании нуклеосомы весь хроматин становится активным.
Дисковидные нуклеосомы имеют диаметр

Второй: нуклеомерный уровень укладки хроматина
этот уровень укладки ДНК. осуществляется за счёт
Второй: нуклеомерный уровень укладки хроматина
этот уровень укладки ДНК. осуществляется за счёт
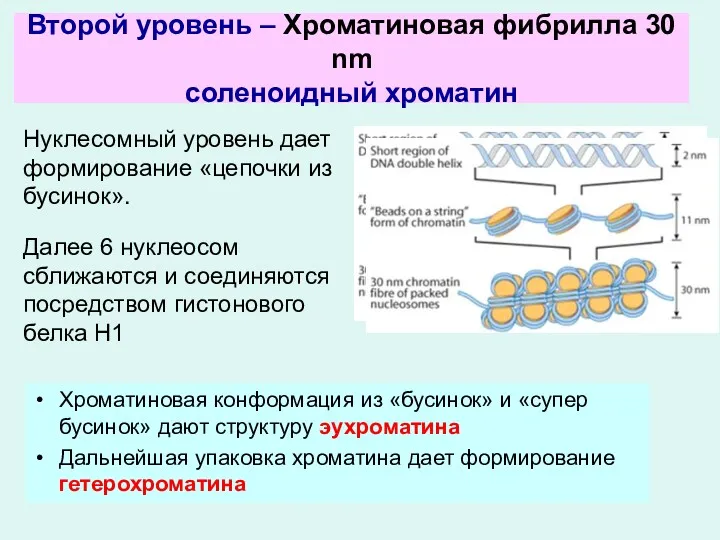
Нуклесомный уровень дает формирование «цепочки из бусинок».
Далее 6 нуклеосом сближаются и
Нуклесомный уровень дает формирование «цепочки из бусинок».
Далее 6 нуклеосом сближаются и

Хромомерный уровень. Дальнейшая упаковка хроматина дает формирование гетерохроматина.
Все остальные уровни
Хромомерный уровень. Дальнейшая упаковка хроматина дает формирование гетерохроматина.
Все остальные уровни
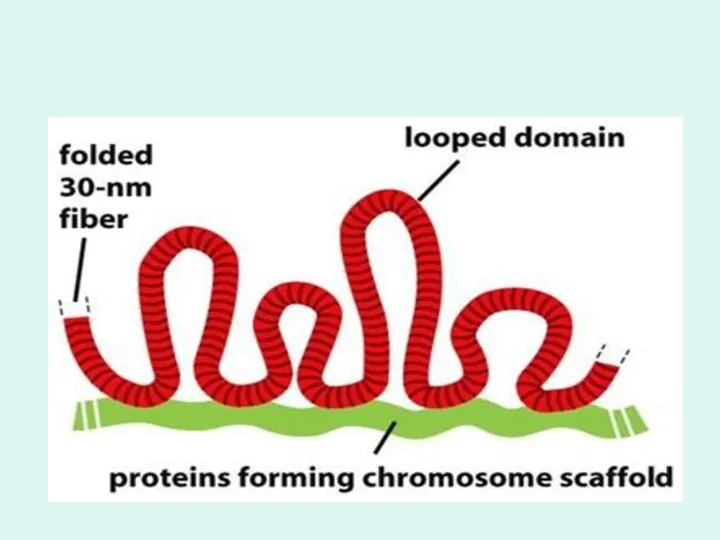

Дальнейшая упаковка хромосом-хромонемная
происходит сближение хромомер и образуются толстые нити, которые становятся
Дальнейшая упаковка хромосом-хромонемная
происходит сближение хромомер и образуются толстые нити, которые становятся

И последний уровень структурной организации хроматина – пятый - хроматидный .
И последний уровень структурной организации хроматина – пятый - хроматидный .

ХРОМОСОМНЫЙ
Метафазная хромосома состоит из двух хроматид, соединенных первичной перетяжкой – центромерой.
ХРОМОСОМНЫЙ
Метафазная хромосома состоит из двух хроматид, соединенных первичной перетяжкой – центромерой.

ХРОМОСОМА
К началу XX в. углубленное изучение поведения этих структур в
ХРОМОСОМА
К началу XX в. углубленное изучение поведения этих структур в

Строение метафазной хромосомы
Термин хромосома был предложен в 1888 г. немецким морфологом
Строение метафазной хромосомы
Термин хромосома был предложен в 1888 г. немецким морфологом
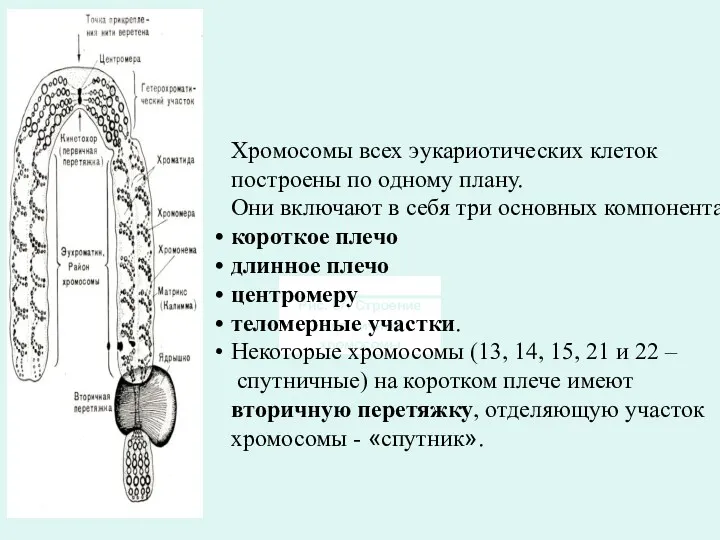
Хромосомы всех эукариотических клеток
построены по одному плану.
Они включают в
Хромосомы всех эукариотических клеток
построены по одному плану.
Они включают в
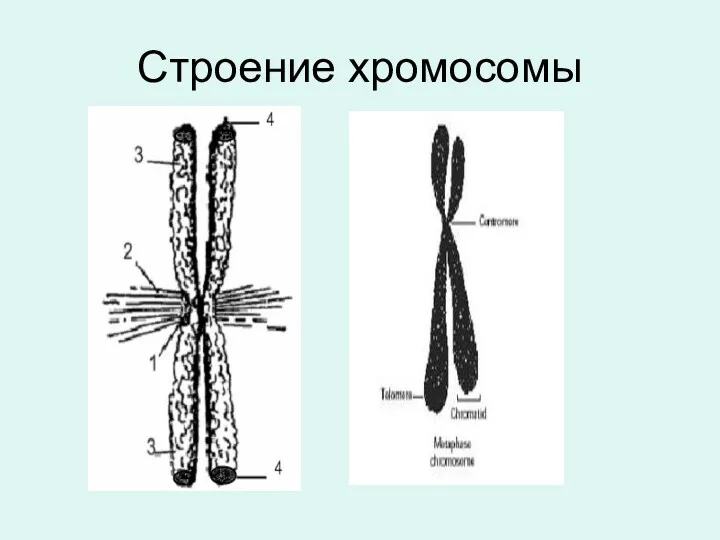
Строение хромосомы
Строение хромосомы

Центромера делит хромосому на два плеча:
Хромосомы с равными плечами называют
Центромера делит хромосому на два плеча:
Хромосомы с равными плечами называют

Кинетохоры
Кинетохоры– это сложные комплексы, состоящие из многих белков. Морфологически они очень
Кинетохоры
Кинетохоры– это сложные комплексы, состоящие из многих белков. Морфологически они очень
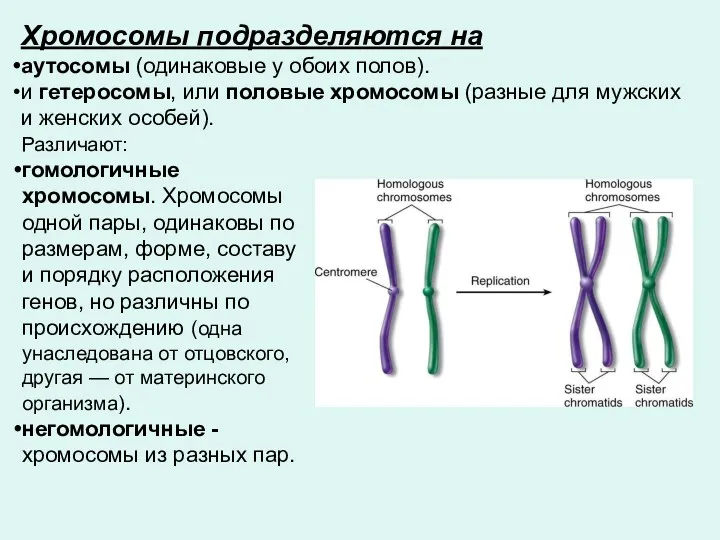
Хромосомы подразделяются на
аутосомы (одинаковые у обоих полов).
и гетеросомы, или
Хромосомы подразделяются на
аутосомы (одинаковые у обоих полов).
и гетеросомы, или

кариотип
Кариотип — диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида,
кариотип
Кариотип — диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида,
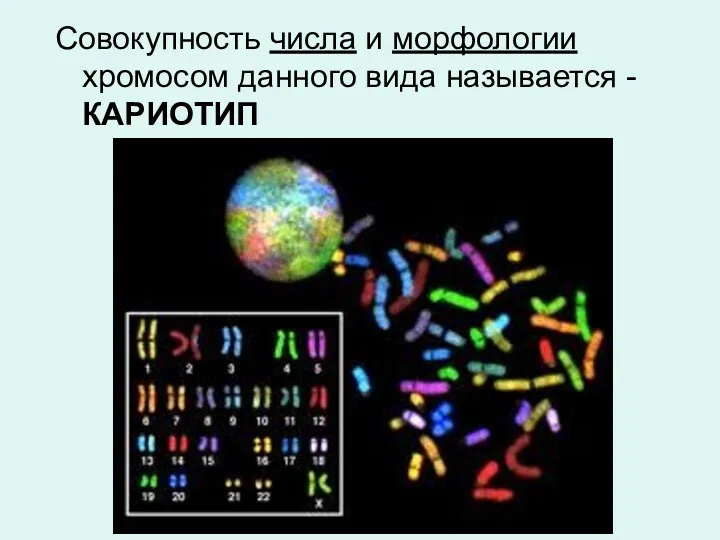
Совокупность числа и морфологии хромосом данного вида называется - КАРИОТИП
Совокупность числа и морфологии хромосом данного вида называется - КАРИОТИП
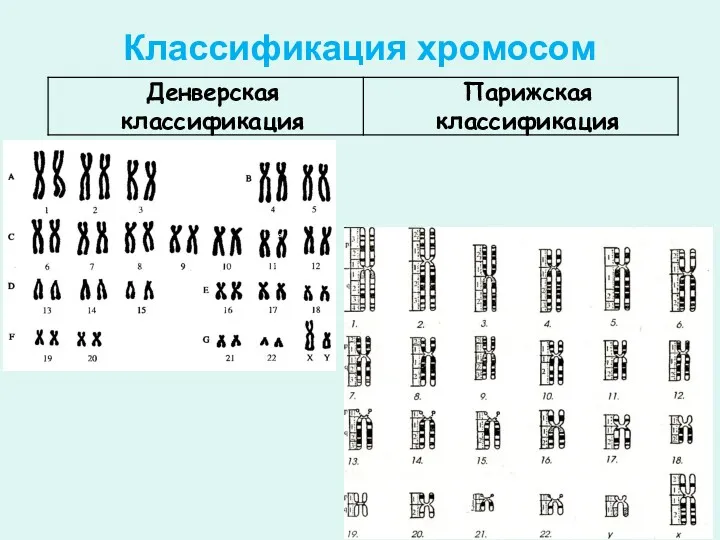
Классификация хромосом
Классификация хромосом

Парижская классификация
В основе Парижской классификации хромосом человека (1971 г.) лежат методы
Парижская классификация
В основе Парижской классификации хромосом человека (1971 г.) лежат методы

Денверская классификация
Денверская классификация

К группе А относят 1—3 пары хромосом. Это cамые крупные, метацентрические

Группа D (13—15 пары). Хромосомы акроцентрические, ЦИ 15. Размеры хромосом около
Группа D (13—15 пары). Хромосомы акроцентрические, ЦИ 15. Размеры хромосом около
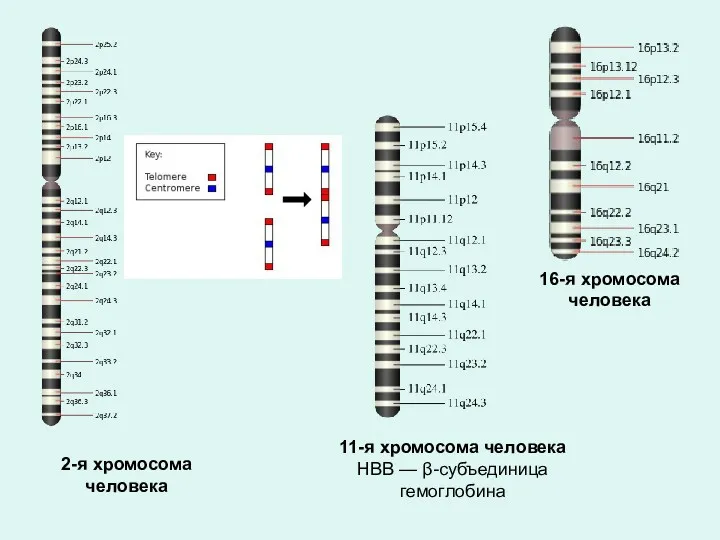
11-я хромосома человека
HBB — β-субъединица гемоглобина
16-я хромосома
человека
2-я хромосома
человека
11-я хромосома человека
HBB — β-субъединица гемоглобина
16-я хромосома
человека
2-я хромосома
человека
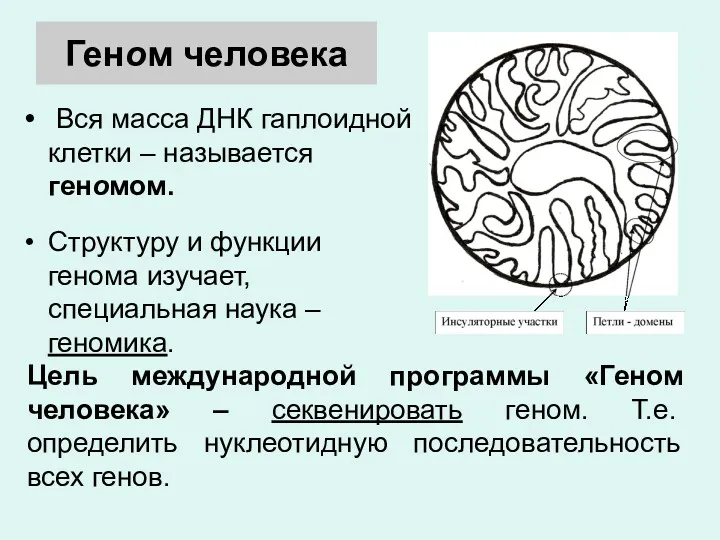
Геном человека
Вся масса ДНК гаплоидной клетки – называется геномом.
Структуру и
Геном человека
Вся масса ДНК гаплоидной клетки – называется геномом.
Структуру и
 Размножение и оплодотворение растений
Размножение и оплодотворение растений Деление клетки. Митоз и мейоз
Деление клетки. Митоз и мейоз Возрастные особенности слуховой сенсорной системы
Возрастные особенности слуховой сенсорной системы Демэкология. Определения популяции
Демэкология. Определения популяции Общая характеристика Покрытосеменных и их классификация
Общая характеристика Покрытосеменных и их классификация Питанием животных: растительноядных, хищников и паразитов
Питанием животных: растительноядных, хищников и паразитов Сапоніни
Сапоніни Начальные этапы развития жизни на Земле
Начальные этапы развития жизни на Земле РИ 1
РИ 1 Отряд Перепончатокрылые
Отряд Перепончатокрылые Кто в каком домике живёт. (Окружающий мир, 1 класс)
Кто в каком домике живёт. (Окружающий мир, 1 класс) Основы экспериментальной биологии. Лекция 7. Горизонты науки. Ч.3 Синтетическая биология
Основы экспериментальной биологии. Лекция 7. Горизонты науки. Ч.3 Синтетическая биология Семейство злаков
Семейство злаков Chemical mutagenes
Chemical mutagenes Потоки вещества и энергии в экосистемах
Потоки вещества и энергии в экосистемах Строение семян однодольных и двудольных растений
Строение семян однодольных и двудольных растений Общая и медицинская генетика
Общая и медицинская генетика Искусственные природные сообщества
Искусственные природные сообщества Доказательство эволюции животных. Сравнительно-анатомические доказательства
Доказательство эволюции животных. Сравнительно-анатомические доказательства Закономерности работы головного мозга
Закономерности работы головного мозга История изучения клетки. Клеточная теория
История изучения клетки. Клеточная теория Введение в биохимию. Строение и функции белков (часть 1)
Введение в биохимию. Строение и функции белков (часть 1) Физиология питания
Физиология питания Деление царств живой природы на группы
Деление царств живой природы на группы Презентация к уроку биологии в 9 классе Происхождение человека
Презентация к уроку биологии в 9 классе Происхождение человека Анатомия и физиология человека. Предмет и его значение. Методы изучения. История
Анатомия и физиология человека. Предмет и его значение. Методы изучения. История Деление клетки. Митоз
Деление клетки. Митоз Педсовет Современные подходы к преподаванию в условиях ФГОС
Педсовет Современные подходы к преподаванию в условиях ФГОС