- Обмен липидов

Содержание
- 2. Содержание темы: Обмен липидов: особенности преваривания и всасывания. β-Окисление жирных кислот в митохондриях и его связь
- 3. Переваривание жиров Переваривание экзогенного жира обязательно требует предварительного эмульгирования. Эмульгаторы – вещества амфифильной природы. Они снижают
- 4. Липаза Язычная липаза. Вырабатывается клетками слизистой оболочки задней части языка. Действие этого фермента проявляется только в
- 5. Действие липазы Образовавшиеся жирные кислоты и моноацилглицерины могут всасываться в кишечную стенку.
- 6. Таким образом, продуктами переваривания жира являются глицерин, жирные кислоты и моноацилглицерины. Всасываются продукты переваривания путем предварительного
- 7. Катаболизм жиров Жиры хранятся до момента их использования. Катаболизм жира идет в три этапа: 1. Гидролиз
- 8. Пути метаболизма глицерина 1. Глицерин может быть повторно использован для синтеза жира или других липидов. 2.
- 9. Распад глицерина по пути к углеводам Если глицерин распадается по пути к углеводам, то происходит дегидрирование:
- 10. Пути использования жирных кислот Жирные кислоты могут вступать в реакции только после активации. Активация жирных кислот
- 12. Судьба жирных кислот Для активной жирной кислоты, как и для глицерина, возможны два пути метаболических превращений:
- 15. β-окисление Если жирная кислота попадает в митохондрию, то она обязательно подвергнется катаболизму до ацетил-КоА.
- 16. Механизм β-окисления ЖК
- 20. Цикличность процесса Укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает в новый цикл реакций
- 21. Энергетический выход окисления ЖК Для того, чтобы рассчитывать энергетический выход при распаде жирных кислот можно воспользоваться
- 22. Особенности окисления ненасыщенных ЖК
- 26. Скорость окисления ЖК В результате β-окисления ЖК образуется Ацетил-КоА. Скорость β-окисления определяется скоростью процессов липолиза. Ускорение
- 27. Синтез кетоновых тел
- 28. Синтез кетоновых тел (продолжение)
- 29. Кетоновые тела и сахарный диабет Ацетон, который образуется при спонтанном (неферментативном) декарбоксилировании ацетоацетата, в организме не
- 30. Синтез жирных кислот β-окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах
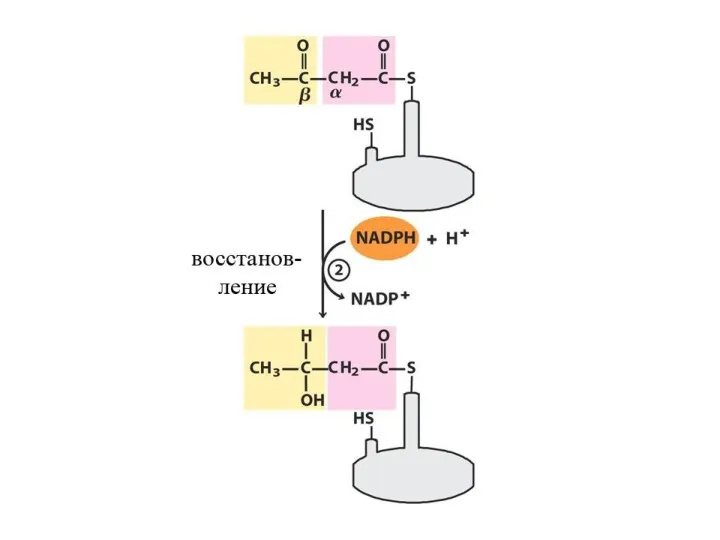
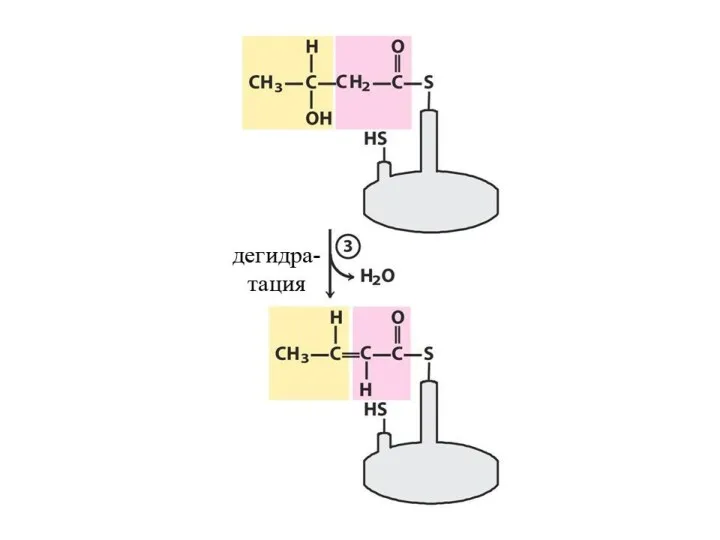
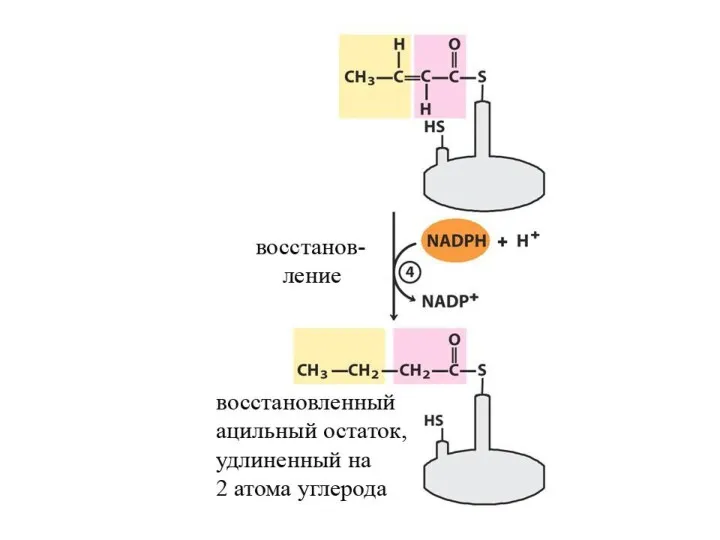
- 31. Далее происходит поэтапное удлинение молекулы Ацетил-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА.
- 39. Скачать презентацию

Содержание темы:
Обмен липидов: особенности преваривания и всасывания.
β-Окисление жирных кислот в
Содержание темы:
Обмен липидов: особенности преваривания и всасывания.
β-Окисление жирных кислот в

Переваривание жиров
Переваривание экзогенного жира обязательно требует предварительного эмульгирования. Эмульгаторы – вещества
Переваривание жиров
Переваривание экзогенного жира обязательно требует предварительного эмульгирования. Эмульгаторы – вещества

Липаза
Язычная липаза. Вырабатывается клетками слизистой оболочки задней части языка. Действие этого
Липаза
Язычная липаза. Вырабатывается клетками слизистой оболочки задней части языка. Действие этого
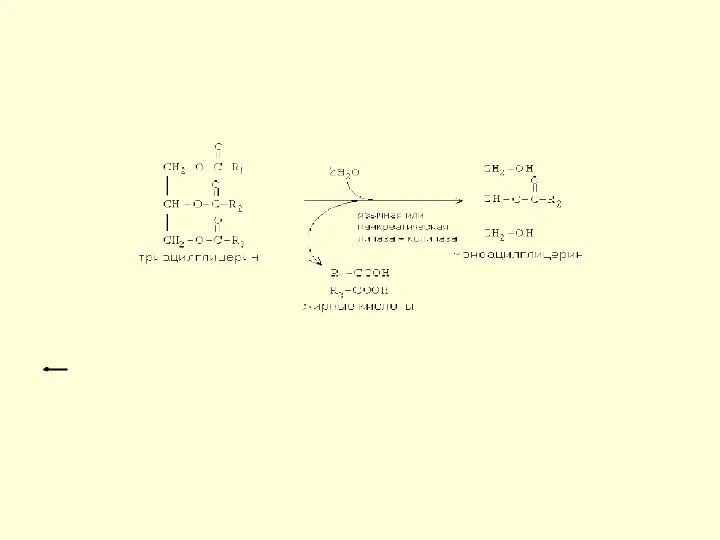
Действие липазы
Образовавшиеся жирные кислоты и моноацилглицерины могут всасываться в кишечную стенку.
Действие липазы
Образовавшиеся жирные кислоты и моноацилглицерины могут всасываться в кишечную стенку.

Таким образом, продуктами переваривания жира являются глицерин, жирные кислоты и моноацилглицерины.
Таким образом, продуктами переваривания жира являются глицерин, жирные кислоты и моноацилглицерины.

Катаболизм жиров
Жиры хранятся до момента их использования. Катаболизм жира идет в
Катаболизм жиров
Жиры хранятся до момента их использования. Катаболизм жира идет в
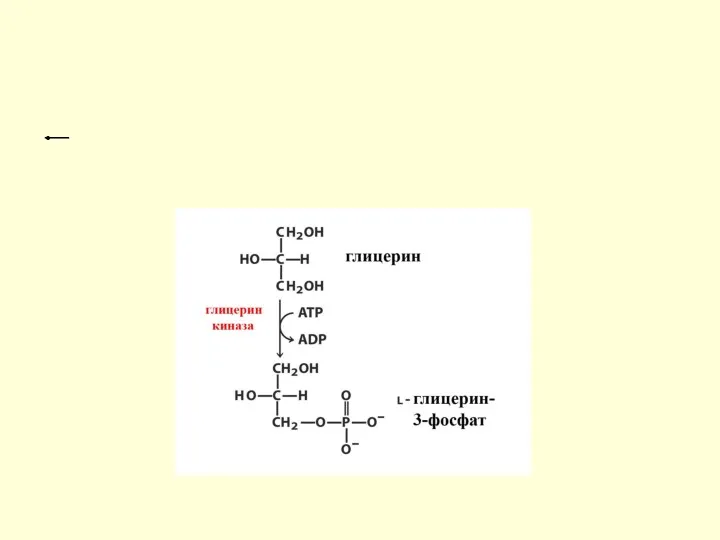
Пути метаболизма глицерина
1. Глицерин может быть повторно использован для синтеза жира
Пути метаболизма глицерина
1. Глицерин может быть повторно использован для синтеза жира
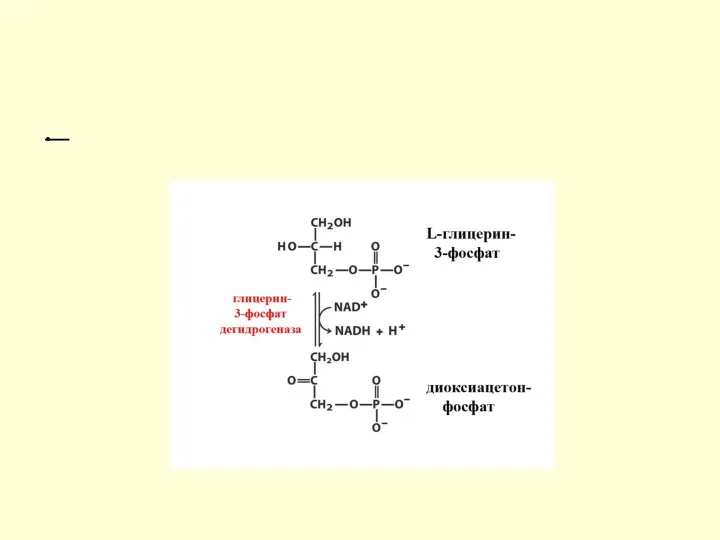
Распад глицерина по пути к углеводам
Если глицерин распадается по пути к
Распад глицерина по пути к углеводам
Если глицерин распадается по пути к
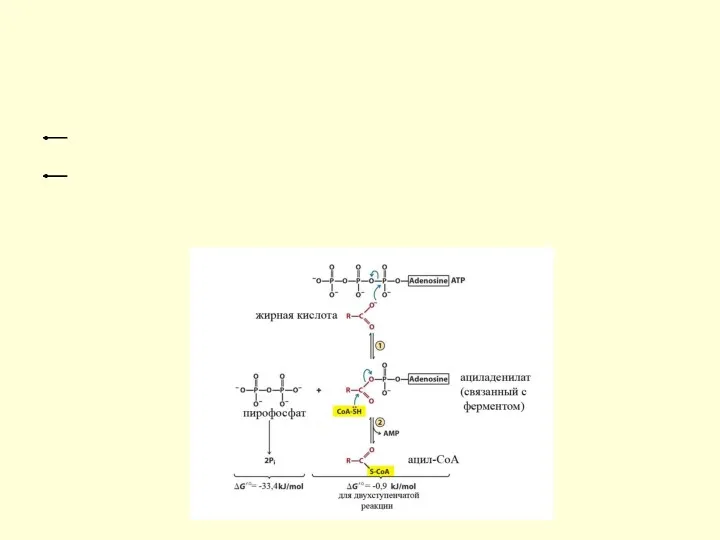
Пути использования жирных кислот
Жирные кислоты могут вступать в реакции только после
Пути использования жирных кислот
Жирные кислоты могут вступать в реакции только после
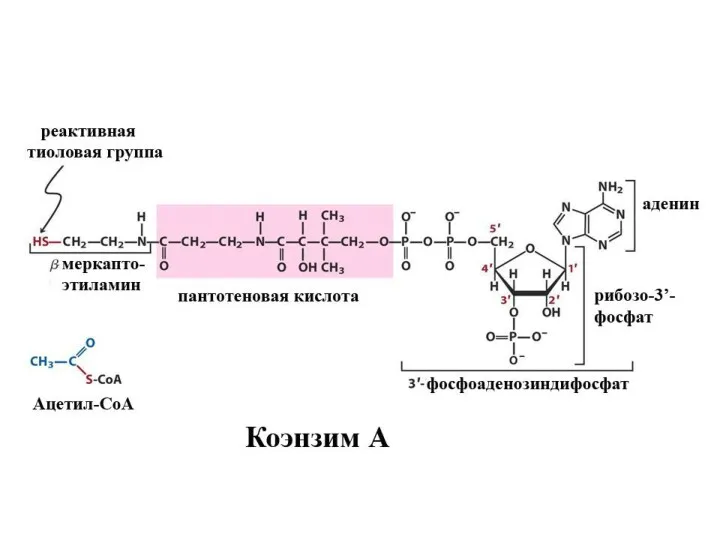

Судьба жирных кислот
Для активной жирной кислоты, как и для глицерина, возможны
Судьба жирных кислот
Для активной жирной кислоты, как и для глицерина, возможны
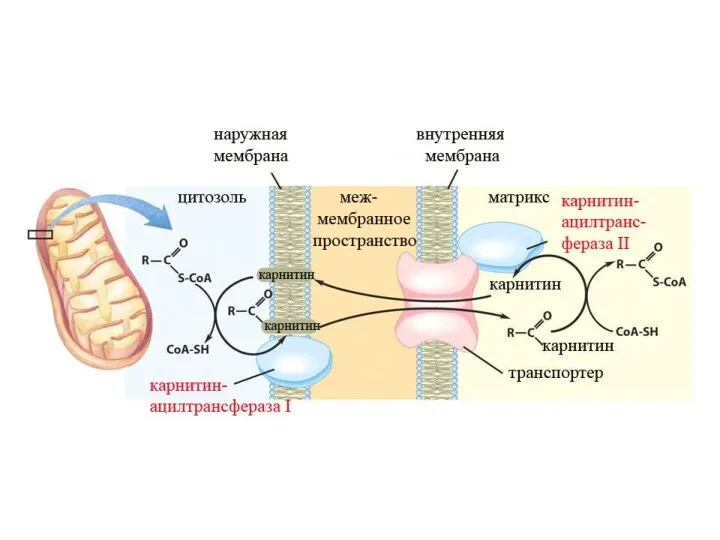
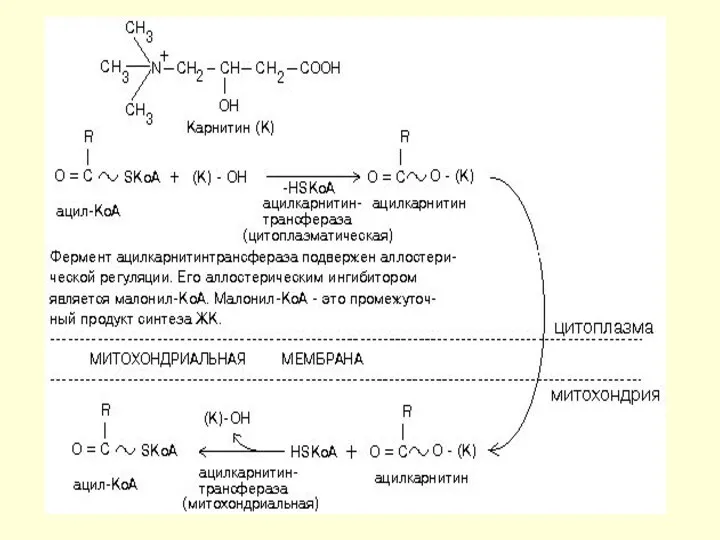
β-окисление
Если жирная кислота попадает в митохондрию, то она обязательно подвергнется
β-окисление
Если жирная кислота попадает в митохондрию, то она обязательно подвергнется
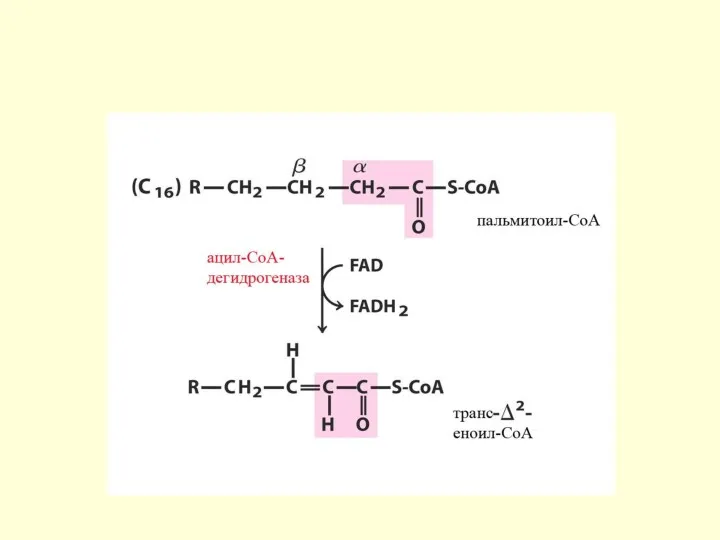
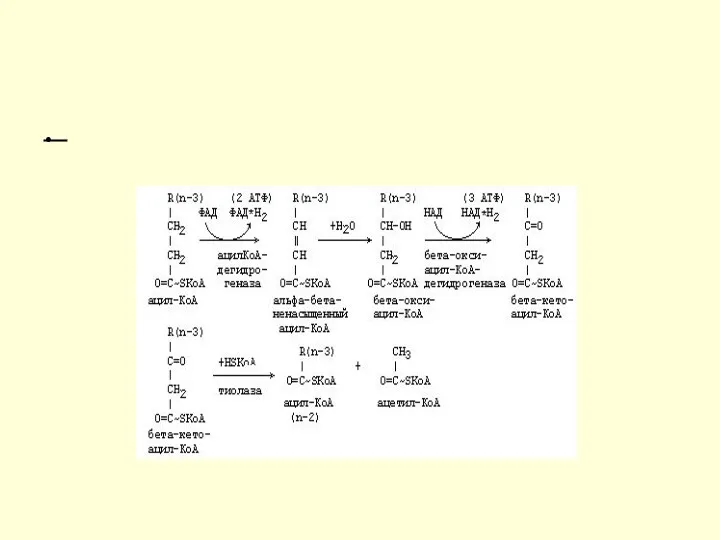
Механизм β-окисления ЖК
Механизм β-окисления ЖК
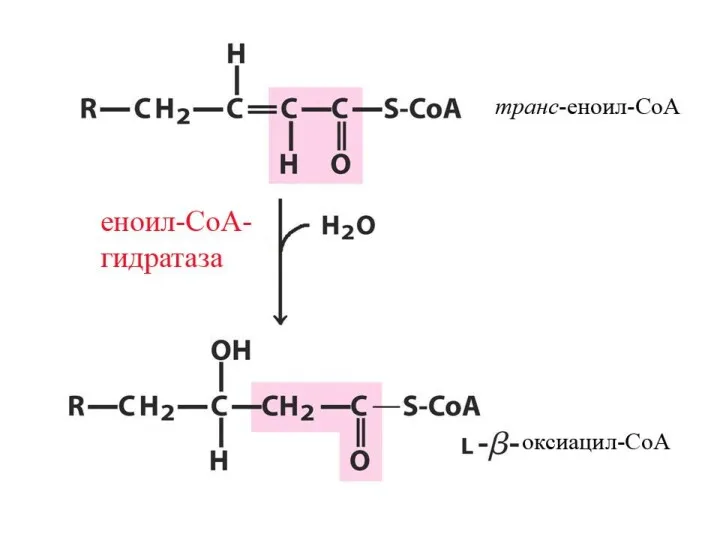
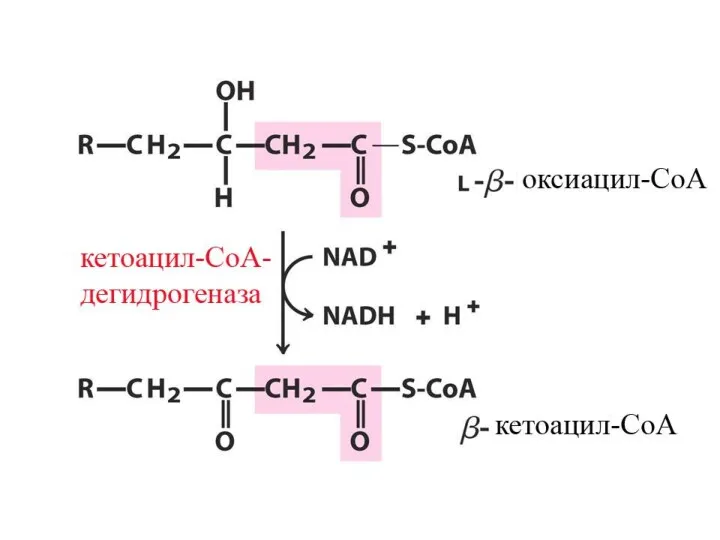
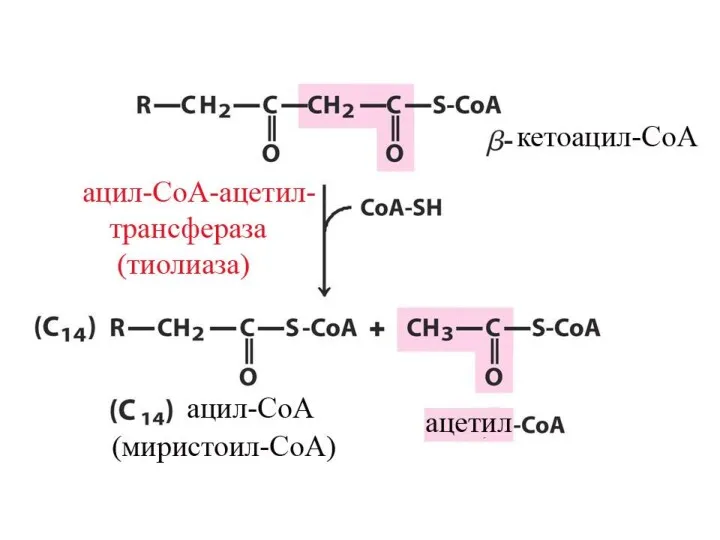
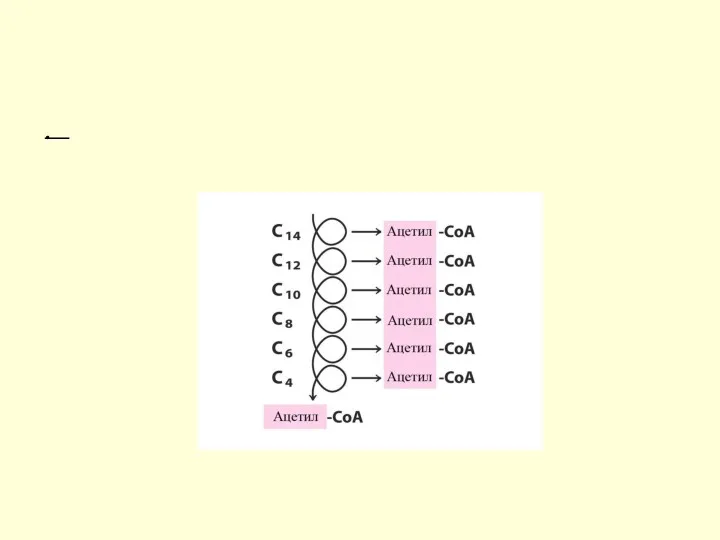
Цикличность процесса
Укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает
Цикличность процесса
Укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает
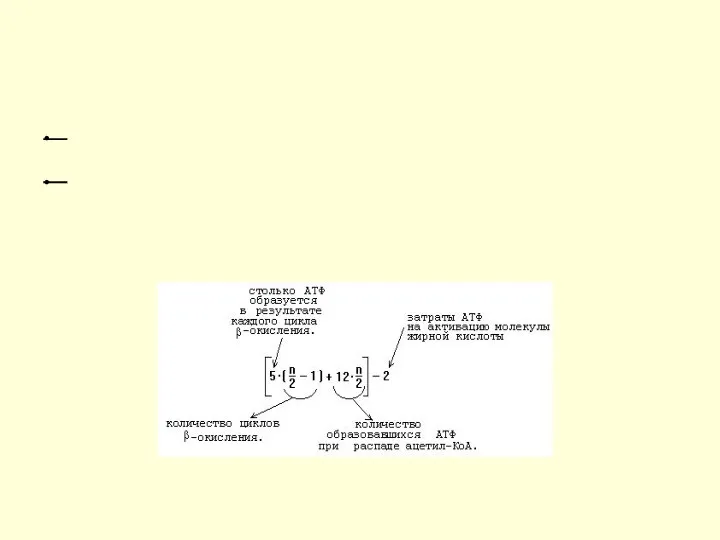
Энергетический выход окисления ЖК
Для того, чтобы рассчитывать энергетический выход при распаде
Энергетический выход окисления ЖК
Для того, чтобы рассчитывать энергетический выход при распаде
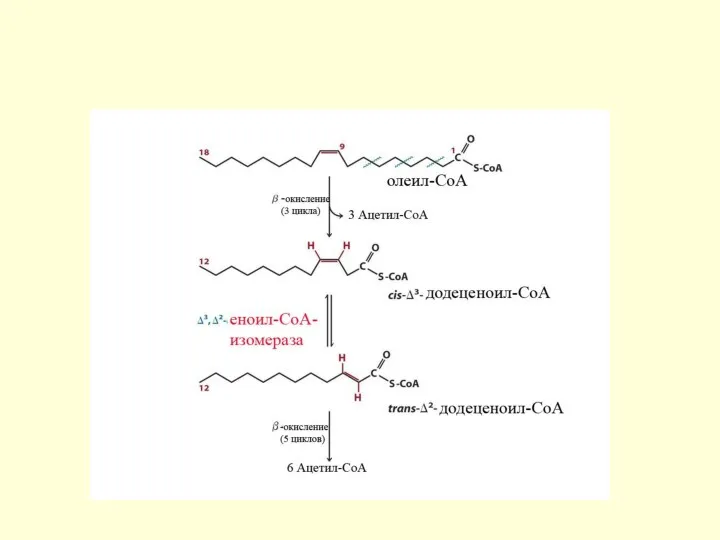
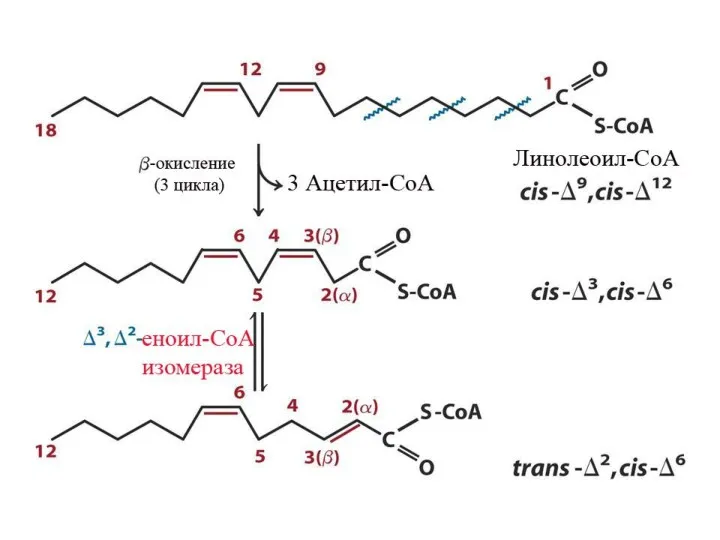
Особенности окисления ненасыщенных ЖК
Особенности окисления ненасыщенных ЖК
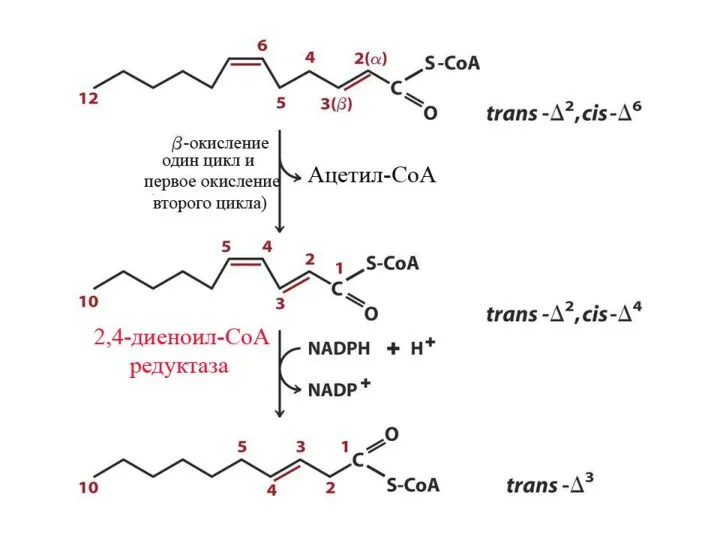
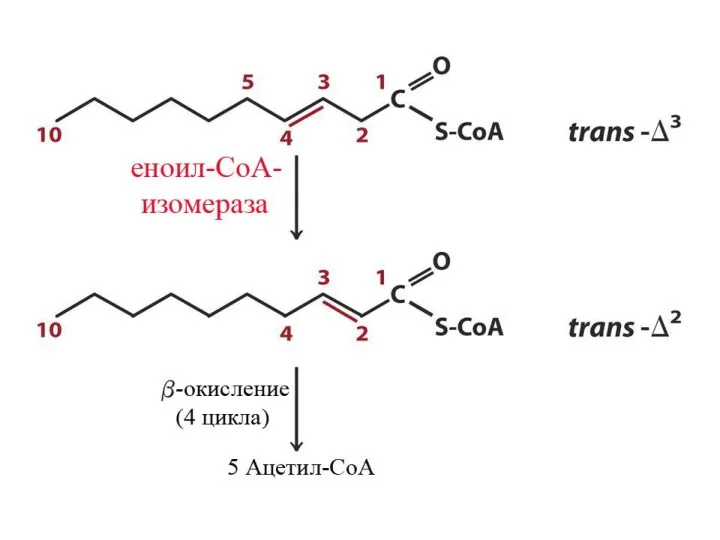

Скорость окисления ЖК
В результате β-окисления ЖК образуется Ацетил-КоА. Скорость β-окисления определяется
Скорость окисления ЖК
В результате β-окисления ЖК образуется Ацетил-КоА. Скорость β-окисления определяется
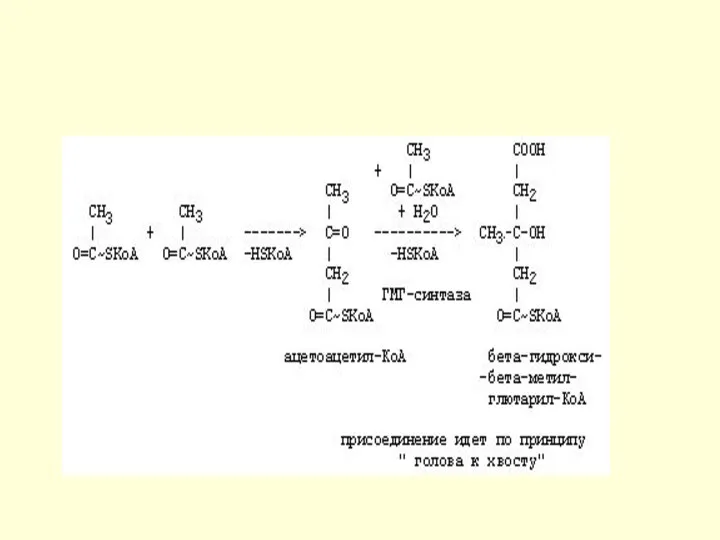
Синтез кетоновых тел
Синтез кетоновых тел
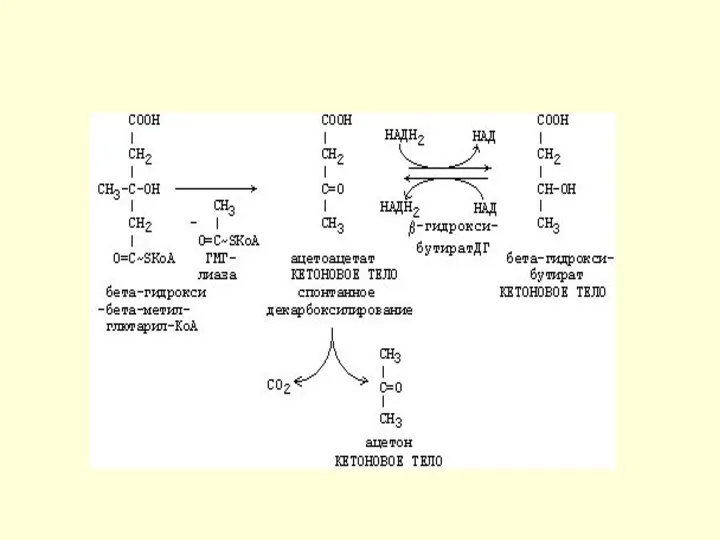
Синтез кетоновых тел (продолжение)
Синтез кетоновых тел (продолжение)

Кетоновые тела и сахарный диабет
Ацетон, который образуется при спонтанном (неферментативном) декарбоксилировании
Кетоновые тела и сахарный диабет
Ацетон, который образуется при спонтанном (неферментативном) декарбоксилировании
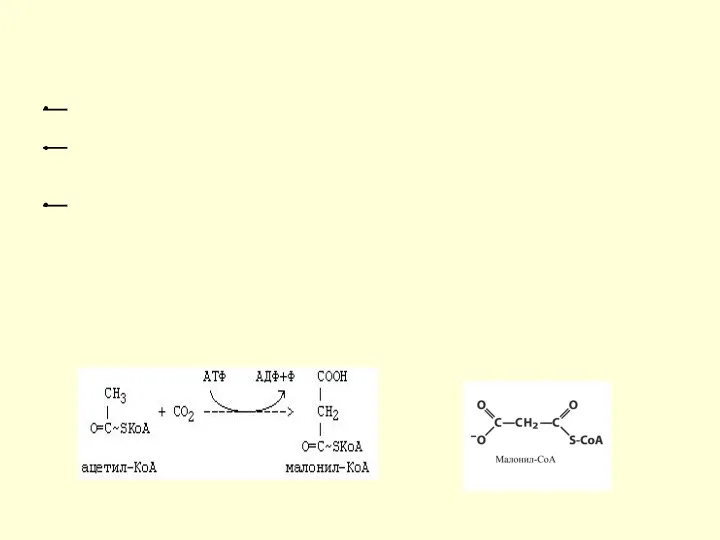
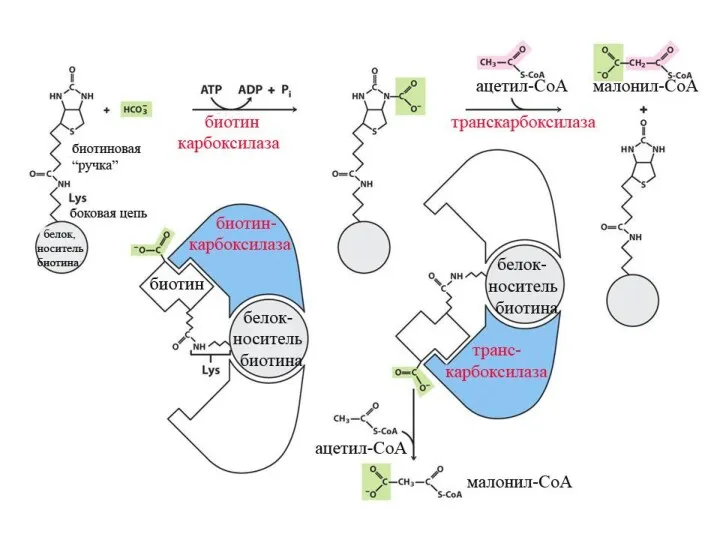
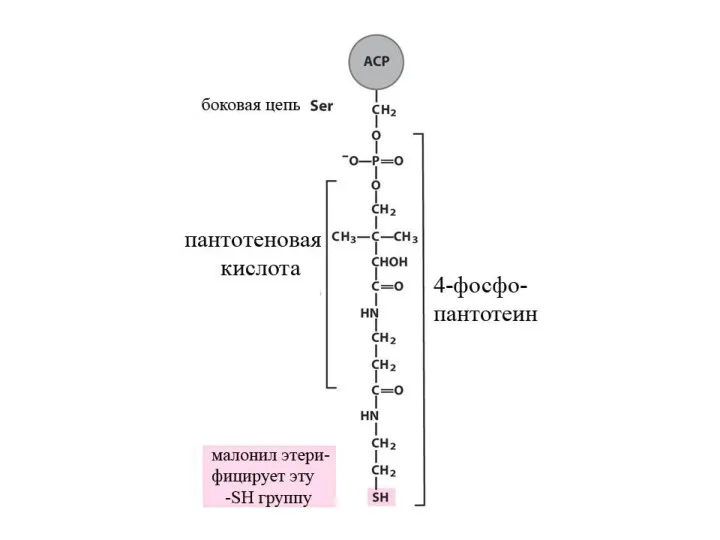
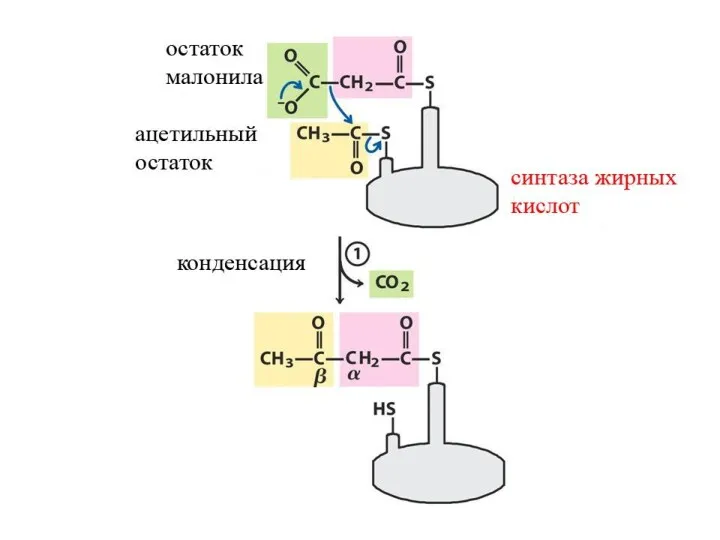
Синтез жирных кислот
β-окисление протекает в митохондриях, а синтез жирных кислот протекает
Синтез жирных кислот
β-окисление протекает в митохондриях, а синтез жирных кислот протекает

Далее происходит поэтапное удлинение молекулы Ацетил-КоА на 2 углеродных атома за
Далее происходит поэтапное удлинение молекулы Ацетил-КоА на 2 углеродных атома за
 Строение сердца. Сердечный цикл
Строение сердца. Сердечный цикл Основы кролиководства
Основы кролиководства Адаптации организмов к условиям обитания
Адаптации организмов к условиям обитания О котах. Породы котов
О котах. Породы котов Человек. Природа человека
Человек. Природа человека Взаимоотношения человека и природы, как фактор развития биосферы
Взаимоотношения человека и природы, как фактор развития биосферы Взаимоотношения микроорганизмов. Факторы внешней среды и микроорганизмы. Экология микроорганизмов
Взаимоотношения микроорганизмов. Факторы внешней среды и микроорганизмы. Экология микроорганизмов Катаболизм. Цикл трикарбоновых кислот
Катаболизм. Цикл трикарбоновых кислот Развитие эволюционной теории. (Лекция 3)
Развитие эволюционной теории. (Лекция 3) Копротрофы. Паразиты высших растений
Копротрофы. Паразиты высших растений Кислотно – щелочное равновесие и основные буферные системы организма
Кислотно – щелочное равновесие и основные буферные системы организма Метаболизмнің гормондық реттелуі. Қоршаған және өндірістік орта факторларының әсер ету бағасының биохимиялық критерийлері
Метаболизмнің гормондық реттелуі. Қоршаған және өндірістік орта факторларының әсер ету бағасының биохимиялық критерийлері Видообразование - микроэволюция
Видообразование - микроэволюция Испарение воды растениями. Листопад
Испарение воды растениями. Листопад Строение клетки. 5 кл
Строение клетки. 5 кл Северный олень
Северный олень Искусственные природные сообщества
Искусственные природные сообщества Науки об организме человека
Науки об организме человека Кинетика и регуляция ферментативного катализа
Кинетика и регуляция ферментативного катализа Азот в пище, воде и организме человека
Азот в пище, воде и организме человека “Професія” дощового черв’яка
“Професія” дощового черв’яка Отчёт-презентация по проектному уроку (биология, 6 класс, тема Шляпочные грибы)
Отчёт-презентация по проектному уроку (биология, 6 класс, тема Шляпочные грибы) Общий обзор головного мозга; его части как производные мозговых пузырей
Общий обзор головного мозга; его части как производные мозговых пузырей Увеличение антропогенного пресса водоёмы
Увеличение антропогенного пресса водоёмы Глобальная роль Fungi в биосфере Земли. Отрицательная роль грибов для человека
Глобальная роль Fungi в биосфере Земли. Отрицательная роль грибов для человека Теория стационарного состояния
Теория стационарного состояния Физминутка
Физминутка презентация Кастовая система семьи медоносной пчелы
презентация Кастовая система семьи медоносной пчелы