- Обмен липидов. Внутритканевые превращения липидов

Содержание
- 3. Источники триацилглицеролов в жировой ткани: 1) Хиломикроны (осуществляют транспорт экзогенных жиров) 2) ЛПОНП (осуществляют транспорт собственных
- 4. МОБИЛИЗАЦИЯ ЖИРОВ Гидролиз внутриклеточного жира осуществляется под действием гормончувствительной липазы - ТАГ-липазы. Образовавшийся диацилглицерол другие тканевые
- 5. Регуляция активности ТАГ-липазы Глюкагон и адреналин через аденилатциклазную систему активируют протеинкиназу А, которая фосфорилирует и активируют
- 6. ОКИСЛЕНИЕ ГЛИЦЕРОЛА В ТКАНЯХ Глицерол может использоваться в качестве: 1) субстрата окисления 2) субстрата глюконеогенеза 3)
- 7. Обозначение атомов углерода в жирной кислоте β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ β-Окисление - специфический путь катаболизма жирных
- 8. АКТИВАЦИЯ ЖИРНОЙ КИСЛОТЫ
- 9. ТРАНСПОРТ АКТИВИРОВАННОЙ ЖИРНОЙ КИСЛОТЫ В МИТОХОНДРИИ Карнитинацилтрансфераза I- регуляторный фермент β-окисления. + АДФ, АМФ, ацил-КоА -
- 10. РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ В МАТРИКСЕ МИТОХОНДРИЙ
- 11. РЕЗУЛЬТАТ 1 ОБОРОТА ЦИКЛА β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ: - молекула ацетил-КоА ФАДН2 НАДН2 укороченный на 2
- 12. ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ 1 виток β-окисления жирных кислот дает: 1 НАДН2→3 АТФ 1
- 13. ОСОБЕННОСТИ β- ОКИСЛЕНИЯ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
- 14. РЕГУЛЯЦИЯ СКОРОСТИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ - АТФ/АДФ - НАД+/НАДН2 наличие субстрата- жирных кислот, поступающих в
- 15. КЕТОНОВЫЕ ТЕЛА Содержание кетоновых тел в сыворотке крови человека в норме 0,03- 0,6 мМ/л Концентрация кетоновых
- 16. БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (в митохондриях печени)
- 17. синтез ГМГ-КоА-синтазы увеличивается при повышении концентрации жирных кислот в крови (голодание, физическая работа) - ГМГ-КоА-синтаза ингибируется
- 18. КЕТОНОВЫЕ ТЕЛА-СУБСТРАТЫ ОКИСЛЕНИЯ (миокард, почки, скелетные мышцы)
- 19. ПРИЧИНЫ АКТИВАЦИИ КЕТОГЕНЕЗА -сахарный диабет, -длительное голодание, -несбалансированное питание (нарушение соотношения углеводов и липидов пищи), -продолжительная
- 20. У новорожденных в крови наблюдается нарастание содержания кетоновых тел. Особенно велика их концентрация в крови в
- 21. Роль инсулина в депонировании жира Инсулин стимулирует синтез ТАГ, по следующим причинам в его пребывании увеличивается
- 23. Скачать презентацию
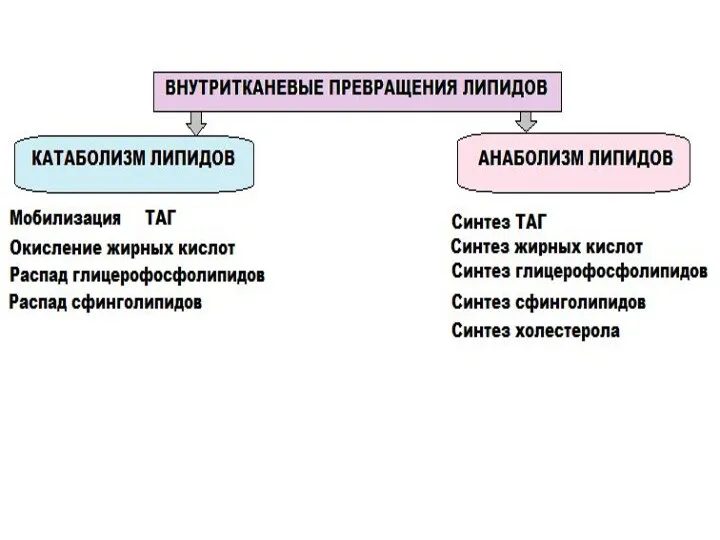
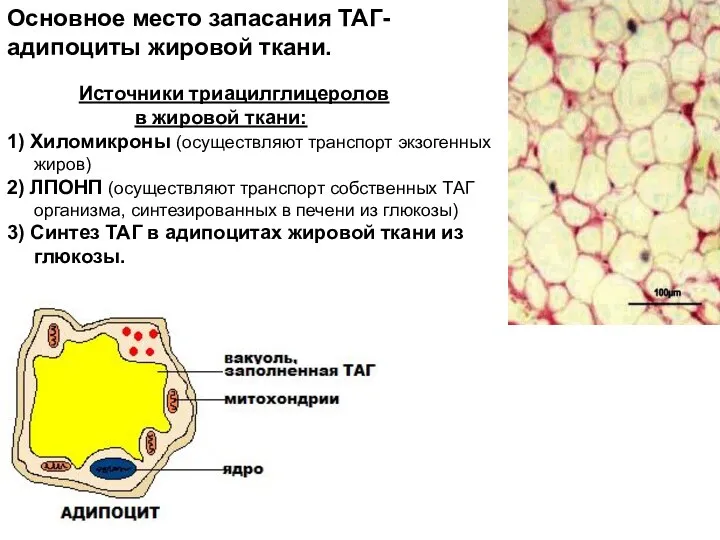
Источники триацилглицеролов
в жировой ткани:
1) Хиломикроны (осуществляют транспорт экзогенных
Источники триацилглицеролов
в жировой ткани:
1) Хиломикроны (осуществляют транспорт экзогенных
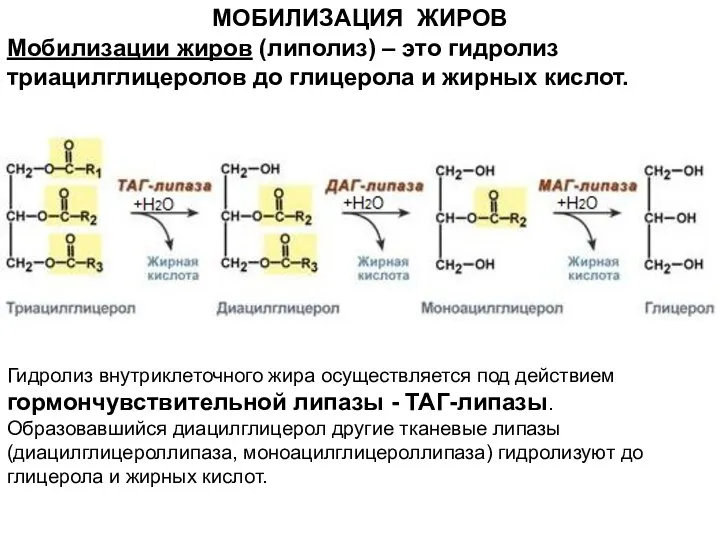
МОБИЛИЗАЦИЯ ЖИРОВ
Гидролиз внутриклеточного жира осуществляется под действием гормончувствительной липазы - ТАГ-липазы.
МОБИЛИЗАЦИЯ ЖИРОВ
Гидролиз внутриклеточного жира осуществляется под действием гормончувствительной липазы - ТАГ-липазы.
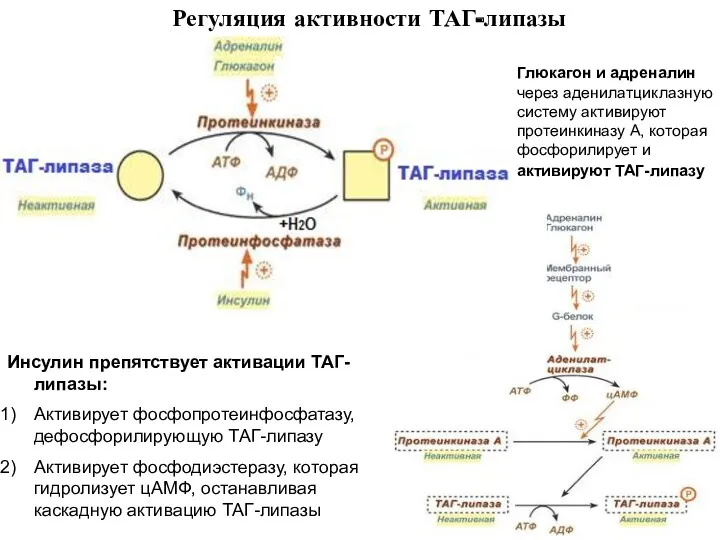
Регуляция активности ТАГ-липазы
Глюкагон и адреналин через аденилатциклазную систему активируют протеинкиназу А,
Регуляция активности ТАГ-липазы
Глюкагон и адреналин через аденилатциклазную систему активируют протеинкиназу А,
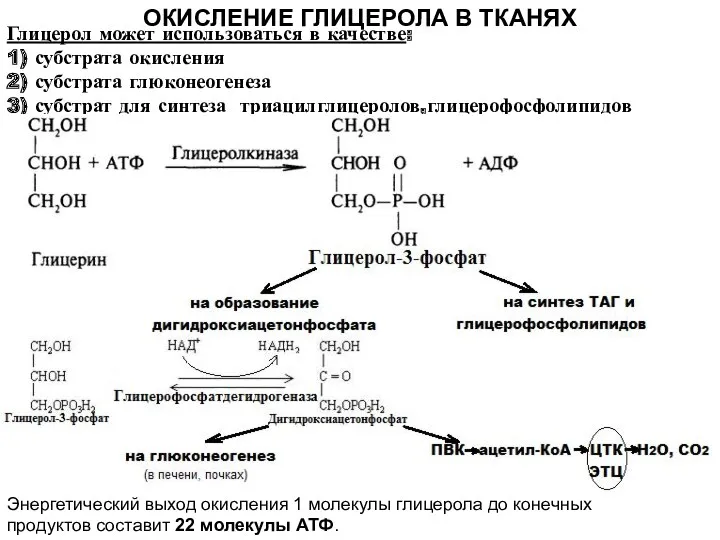
ОКИСЛЕНИЕ ГЛИЦЕРОЛА В ТКАНЯХ
Глицерол может использоваться в качестве:
1) субстрата окисления
2)
ОКИСЛЕНИЕ ГЛИЦЕРОЛА В ТКАНЯХ
Глицерол может использоваться в качестве:
1) субстрата окисления
2)

Обозначение атомов углерода в жирной кислоте
β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
β-Окисление - специфический
Обозначение атомов углерода в жирной кислоте
β- ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
β-Окисление - специфический
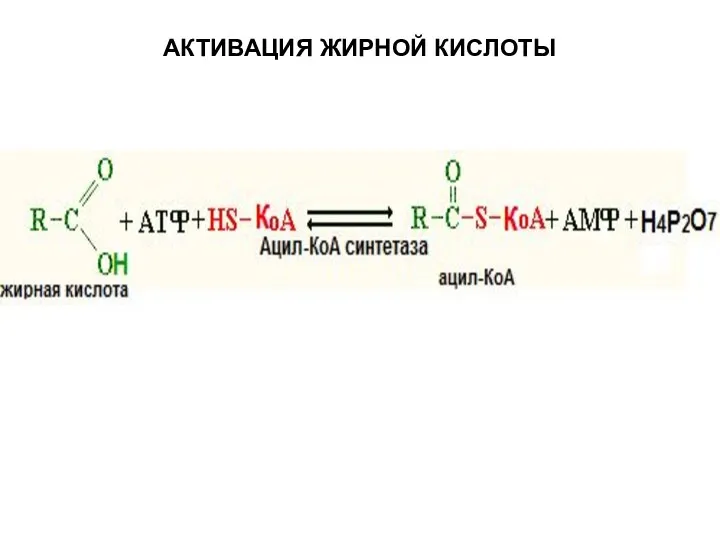
АКТИВАЦИЯ ЖИРНОЙ КИСЛОТЫ
АКТИВАЦИЯ ЖИРНОЙ КИСЛОТЫ
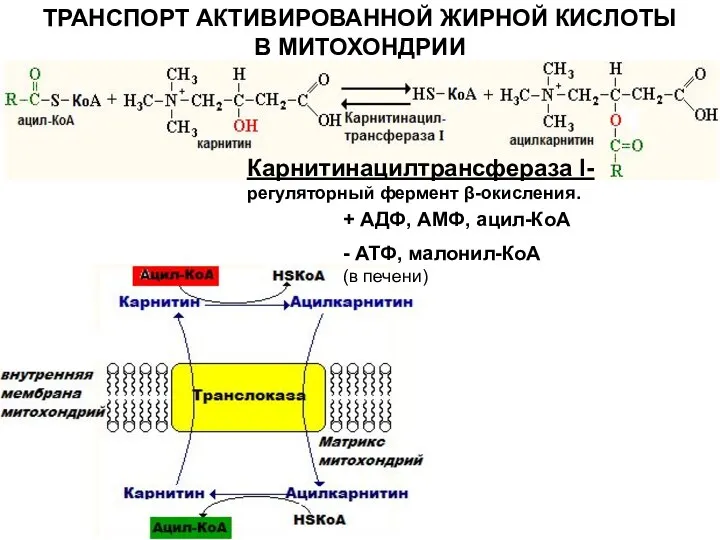
ТРАНСПОРТ АКТИВИРОВАННОЙ ЖИРНОЙ КИСЛОТЫ В МИТОХОНДРИИ
Карнитинацилтрансфераза I- регуляторный фермент β-окисления.
+
ТРАНСПОРТ АКТИВИРОВАННОЙ ЖИРНОЙ КИСЛОТЫ В МИТОХОНДРИИ
Карнитинацилтрансфераза I- регуляторный фермент β-окисления.
+
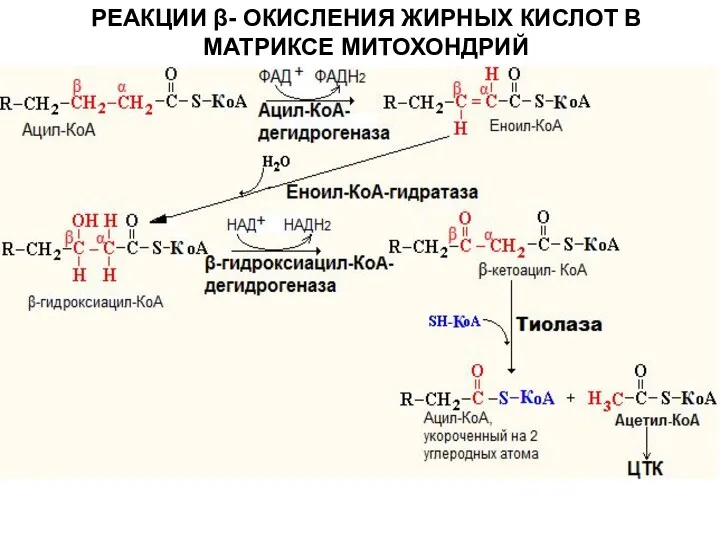
РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ В МАТРИКСЕ МИТОХОНДРИЙ
РЕАКЦИИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ В МАТРИКСЕ МИТОХОНДРИЙ
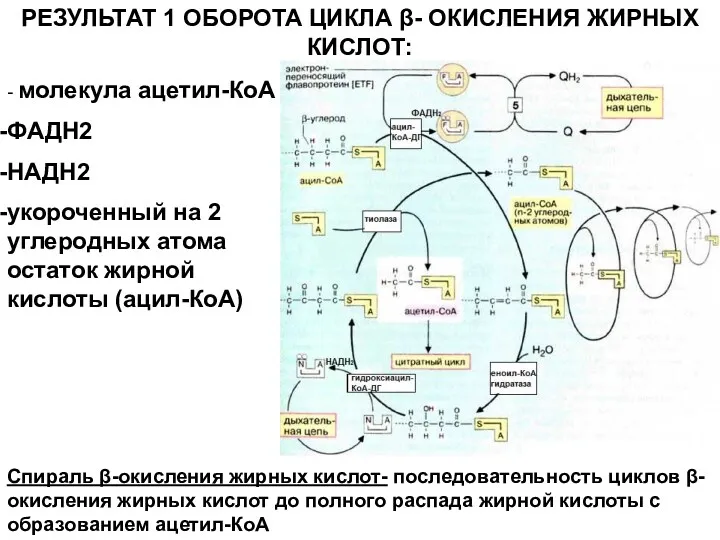
РЕЗУЛЬТАТ 1 ОБОРОТА ЦИКЛА β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ:
- молекула ацетил-КоА
ФАДН2
НАДН2
укороченный на
РЕЗУЛЬТАТ 1 ОБОРОТА ЦИКЛА β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ:
- молекула ацетил-КоА
ФАДН2
НАДН2
укороченный на
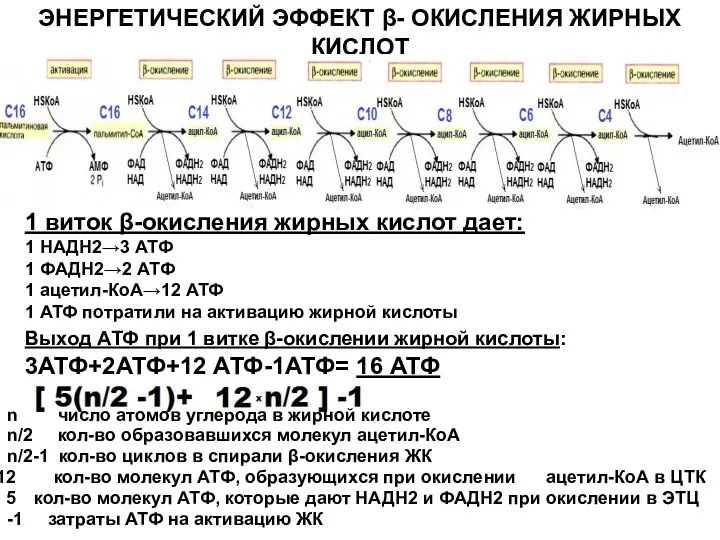
ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
1 виток β-окисления жирных кислот дает:
1
ЭНЕРГЕТИЧЕСКИЙ ЭФФЕКТ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
1 виток β-окисления жирных кислот дает:
1
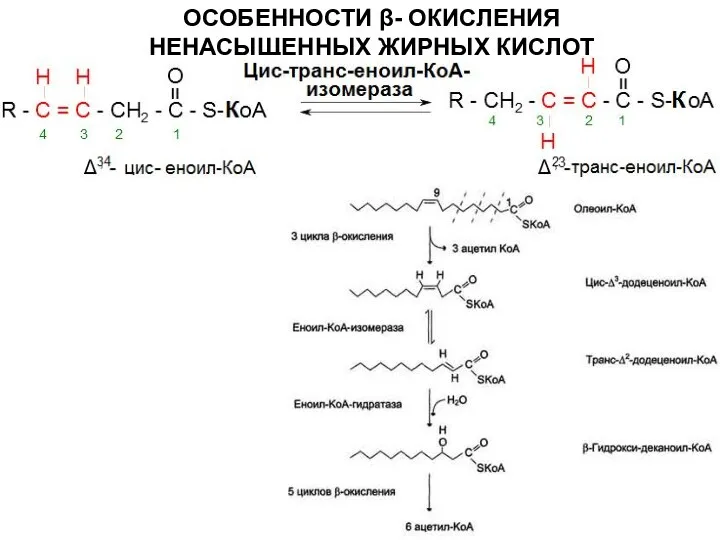
ОСОБЕННОСТИ β- ОКИСЛЕНИЯ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ
ОСОБЕННОСТИ β- ОКИСЛЕНИЯ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ

РЕГУЛЯЦИЯ СКОРОСТИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
- АТФ/АДФ
- НАД+/НАДН2
наличие субстрата- жирных кислот,
РЕГУЛЯЦИЯ СКОРОСТИ β- ОКИСЛЕНИЯ ЖИРНЫХ КИСЛОТ
- АТФ/АДФ
- НАД+/НАДН2
наличие субстрата- жирных кислот,

КЕТОНОВЫЕ ТЕЛА
Содержание кетоновых тел в сыворотке крови человека в норме 0,03-
КЕТОНОВЫЕ ТЕЛА
Содержание кетоновых тел в сыворотке крови человека в норме 0,03-
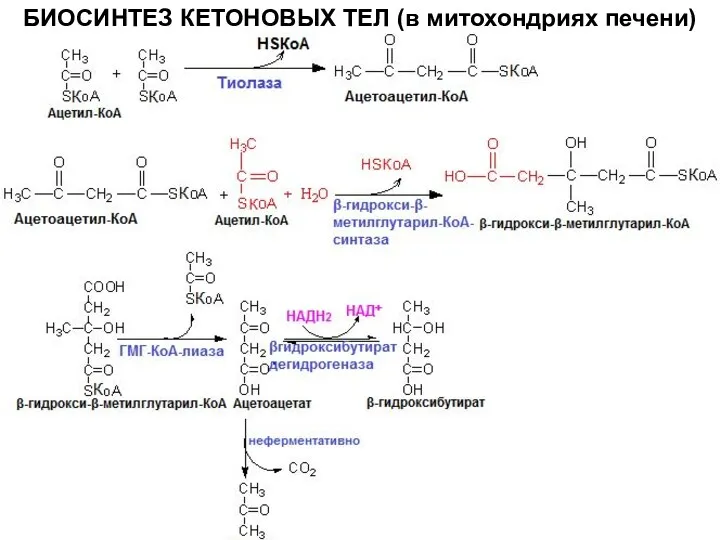
БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (в митохондриях печени)
БИОСИНТЕЗ КЕТОНОВЫХ ТЕЛ (в митохондриях печени)

синтез ГМГ-КоА-синтазы увеличивается при повышении концентрации жирных кислот в крови (голодание,
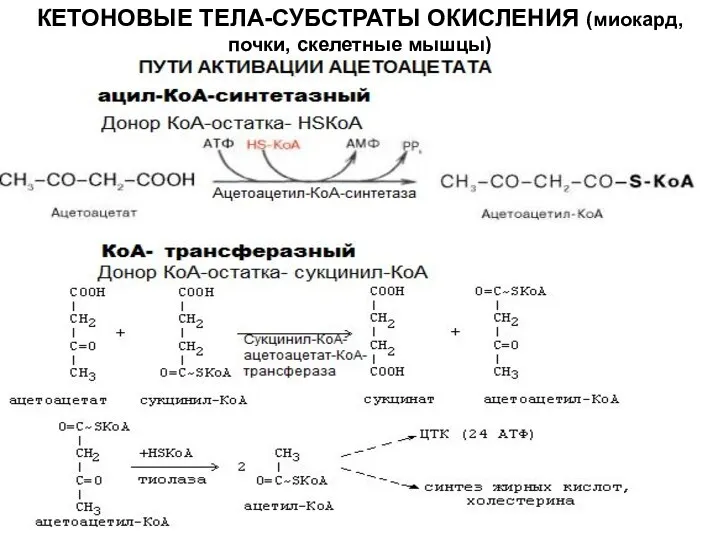
КЕТОНОВЫЕ ТЕЛА-СУБСТРАТЫ ОКИСЛЕНИЯ (миокард, почки, скелетные мышцы)
КЕТОНОВЫЕ ТЕЛА-СУБСТРАТЫ ОКИСЛЕНИЯ (миокард, почки, скелетные мышцы)

ПРИЧИНЫ АКТИВАЦИИ КЕТОГЕНЕЗА
-сахарный диабет,
-длительное голодание,
-несбалансированное питание (нарушение соотношения углеводов и липидов
ПРИЧИНЫ АКТИВАЦИИ КЕТОГЕНЕЗА
-сахарный диабет,
-длительное голодание,
-несбалансированное питание (нарушение соотношения углеводов и липидов

У новорожденных в крови наблюдается нарастание содержания кетоновых тел. Особенно велика
У новорожденных в крови наблюдается нарастание содержания кетоновых тел. Особенно велика
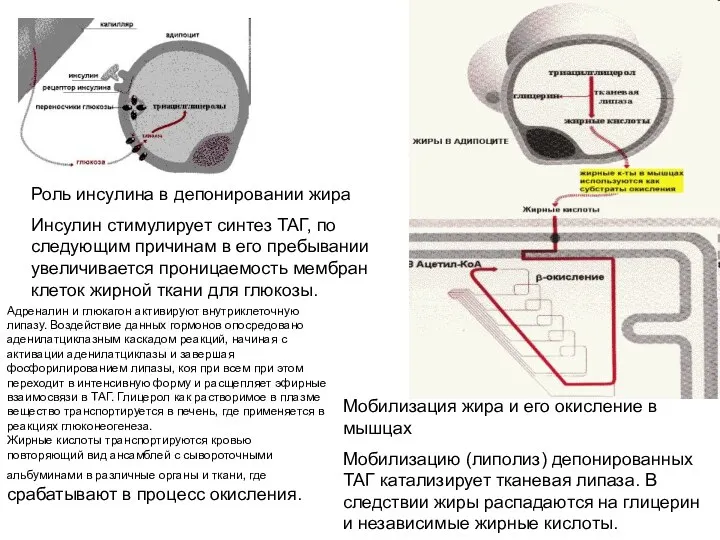
Роль инсулина в депонировании жира
Инсулин стимулирует синтез ТАГ, по следующим причинам
Роль инсулина в депонировании жира
Инсулин стимулирует синтез ТАГ, по следующим причинам
 Лягушка. Чудесное превращение
Лягушка. Чудесное превращение Разработка интеллектуальной системы освещения растений
Разработка интеллектуальной системы освещения растений Молекулярные и клеточные механизмы старения: современные концепции
Молекулярные и клеточные механизмы старения: современные концепции Разнообразие живой природы. Царства живых организмов (5 класс)
Разнообразие живой природы. Царства живых организмов (5 класс) 3c57d4db65194731b010a474653bcc81
3c57d4db65194731b010a474653bcc81 Терморегуляция организма. Урок биологии в 8 классе
Терморегуляция организма. Урок биологии в 8 классе Селекція тварин
Селекція тварин Ссавці. Особливості зовнішньої будови ссавців
Ссавці. Особливості зовнішньої будови ссавців Химический состав бактериальной клетки
Химический состав бактериальной клетки Эволюционное учение. Микро- и макроэволюция. ССТЭ. Генетические процессы в популяциях человека
Эволюционное учение. Микро- и макроэволюция. ССТЭ. Генетические процессы в популяциях человека Процесс кровообращения. Сердце, анатомия и физиология
Процесс кровообращения. Сердце, анатомия и физиология Морфологические признаки семян овощных и декоративных культур
Морфологические признаки семян овощных и декоративных культур Тестовый контроль знаний учащихся на уроках биологии
Тестовый контроль знаний учащихся на уроках биологии Глотка. Пищевод. Желудок
Глотка. Пищевод. Желудок Введение в курс гистологии, эмбриологии, цитологии. Задачи и методы изучения дисциплины
Введение в курс гистологии, эмбриологии, цитологии. Задачи и методы изучения дисциплины Химический состав растений
Химический состав растений Особенности новейших технологий производства ферментных препаратов: амилаз, протеаз, липаз, лактаз, глюкоксидаз 2 часть
Особенности новейших технологий производства ферментных препаратов: амилаз, протеаз, липаз, лактаз, глюкоксидаз 2 часть Дуб
Дуб Майстерність маскування
Майстерність маскування Технологии выращивания декоративных растений в защищенном грунте
Технологии выращивания декоративных растений в защищенном грунте Биохимия печени. (Лекция 31)
Биохимия печени. (Лекция 31) Биотические связи в природе. 9 класс
Биотические связи в природе. 9 класс Wild life in Britain
Wild life in Britain Анатомия и физиология органов речи
Анатомия и физиология органов речи Бүйрек физиологиясы. Несеп түзілуінің механизмі
Бүйрек физиологиясы. Несеп түзілуінің механизмі Дихання рослин
Дихання рослин Отряды Млекопитающих
Отряды Млекопитающих Прокариоты. Особенности внешнего строения прокариотической клетки
Прокариоты. Особенности внешнего строения прокариотической клетки