Обмен углеводов: значение, переваривание. Гликолиз. Пентозофосфатный путь окисления глюкозы презентация
- Обмен углеводов: значение, переваривание. Гликолиз. Пентозофосфатный путь окисления глюкозы

Содержание
- 2. ПЛАН ЛЕКЦИИ: Обмен углеводов. Гликолиз. Пентозофосфатный путь окисления глюкозы.
- 3. Обмен углеводов в организме человека складываются из следующих процессов: Расщепление в желудочно-кишечном тракте до моносахаридов поступающих
- 4. ОБМЕН УГЛЕВОДОВ Распад (гликогеногенолиз) и синтез (гликогеногенез) гликогена Аэробный гликолиз Окислительное декарбоксилирование пирувата ЦТК Пентозо- фосфатный
- 5. Две основные функции: Углеводы – источник углеродов, который необходим для синтеза ряда соединений (белков, нуклеиновых кислот,
- 6. Другие функции: Резервная (крахмал, гликоген). Структурная (полисахариды образуют прочный остов в комплексе с белками и липидами,
- 7. Переваривание углеводов в организме Источником углеводов для организма служат углеводы пищи - крахмал, сахароза и лактоза.
- 8. Переваривание углеводов Амилаза слюны расщепляет α-1,4-гликозидные связи в крахмале. В ротовой полости происходит лишь частичное переваривание
- 9. Переваривание углеводов Последующее переваривание нерасщепленного или частично расщепленного крахмала происходит в кишечнике. В двенадцатиперстной кишке pH
- 10. Переваривание углеводов Мальтоза и изомальтоза вместе с другими пищевыми дисахаридами - сахарозой и лактозой - гидролизуются
- 11. Переваривание углеводов
- 12. Продукты полного переваривания углеводов – глюкоза, галактоза и фруктоза – через клетки кишечника поступают в кровь.
- 13. Гликолиз Гликолиз (от греч. glykys – сладкий, lysys – распад ) – один центральных путей катаболизма
- 14. Гликолиз включает превращения трех разных типов: Распад углеродного скелета глюкозы с образованием пирувата ( путь атомов
- 15. АЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит расщепление глюкозы с образованием двух
- 16. Стадии гликолиза I. 1 реакция. Фосфорилирование глюкозы: реакция протекает необратимо, катализируется глюкокиназой или гексокиназой и требует
- 17. Ферменты участвующие в фосфорилировании глюкозы. Связывание гексокиназы с гексозой происходит по типу индуцированного соответствия: молекула фермента
- 18. Второй реакцией глюколиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
- 19. Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ: Данная реакция
- 20. Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы: 5%
- 21. Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
- 22. II. На второй стадии гликолиза происходит запасание энергии. Из одной молекулы ГЛЮ образуется две молекулы глицеральдегид-3-фосфата,
- 23. Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении
- 24. Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицерат превращается 2-фосфоглицерат. Фермент – фосфоглицерамутаза. Реакция
- 25. Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицерат в результате отщепления молекулы воды переходит в фосфоенолпируват,
- 26. Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование).
- 27. Конечным продуктом аэробного гликолиза является пируват, а энергетический баланс складывается из 2 молекул АТФ образовавшихся в
- 28. Глицерофосфатный челночный механизм Цитоплазма Митохондрии дигидрокси- ацетонфосфат Глицерол-3-фосфат 2 2 2НАД·Н + Н+ 2НАД + ФАД+
- 29. Малат-аспартатный челночный механизм Цитоплазма Митохондрии ЩУК малат НАД·Н + Н+ НАД + глутамат аспартат глутамат аспартат
- 30. Баланс аэробного гликолиза 1 реакция - 1 АТФ 3 реакция - 1 АТФ 6 реакция +
- 31. АНАЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит расщепление глюкозы с образованием двух
- 32. Дальше процесс идет в зависимости от наличия или отсутствия кислорода в клетке: При анаэробных условиях, например
- 33. В этих условиях образовавшийся при гликолизе НАДН регенерируется за счет пирувата, который восстанавливается до лактата. Электроны,
- 34. Баланс анаэробного гликолиза 1 реакция - 1 АТФ 3 реакция - 1 АТФ 6 реакция -
- 35. Пентозофосфатный путь окисления глюкозы Локализация: молочная железа, эмбриональная и жировая ткань, эритроциты, легкие, щитовидная железа, мозг,
- 36. В пентозофосфатном пути превращения глюкозы можно выделить: Окислительный этап Поставляет клеткам кофермент NADPH (использующийся как донор
- 37. Глюкоза Глюкозо -6-фосфат Фруктозо -6-фосфат Глицеральдегид-3-фосфат Гликолиз 2НАДФ+ 2НАДФН·Н+ Окислительная фаза Синтез жирных кислот Синтез стероидов
- 38. Условия протекания пентозофосфатного пути окисления глюкозы Судьба глюкозо-6-фосфата — вступит ли он в гликолиз или пентозофосфатный
- 39. НАДФ+ НАДФН+Н+ Глюкозо-6-фосфатдегидрогеназа Глюконатлактон-гидратаза Н2О НАДФ+ НАДФН+Н+ 6-Фосфоглюконатдегидрогеназа СО2 Реакции окислительного этапа пентозофосфатного пути
- 41. Скачать презентацию

ПЛАН ЛЕКЦИИ:
Обмен углеводов.
Гликолиз.
Пентозофосфатный путь окисления глюкозы.
ПЛАН ЛЕКЦИИ:
Обмен углеводов.
Гликолиз.
Пентозофосфатный путь окисления глюкозы.

Обмен углеводов в организме человека складываются из следующих процессов:
Расщепление в желудочно-кишечном
Обмен углеводов в организме человека складываются из следующих процессов:
Расщепление в желудочно-кишечном
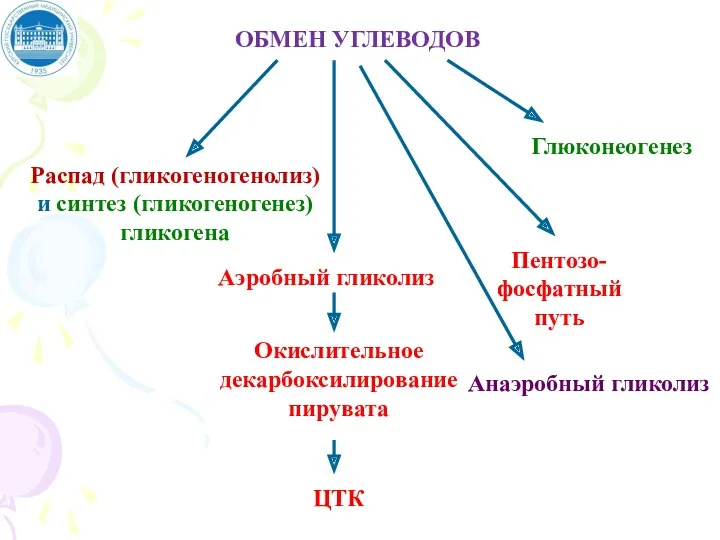
ОБМЕН УГЛЕВОДОВ
Распад (гликогеногенолиз)
и синтез (гликогеногенез)
гликогена
Аэробный гликолиз
Окислительное
декарбоксилирование
пирувата
ЦТК
Пентозо-
фосфатный
путь
Глюконеогенез
Анаэробный гликолиз
ОБМЕН УГЛЕВОДОВ
Распад (гликогеногенолиз)
и синтез (гликогеногенез)
гликогена
Аэробный гликолиз
Окислительное
декарбоксилирование
пирувата
ЦТК
Пентозо-
фосфатный
путь
Глюконеогенез
Анаэробный гликолиз

Две основные функции:
Углеводы – источник углеродов, который необходим для синтеза ряда
Две основные функции:
Углеводы – источник углеродов, который необходим для синтеза ряда

Другие функции:
Резервная (крахмал, гликоген).
Структурная (полисахариды образуют прочный остов в комплексе с
Другие функции:
Резервная (крахмал, гликоген).
Структурная (полисахариды образуют прочный остов в комплексе с

Переваривание
углеводов в организме
Источником углеводов для организма служат углеводы пищи -
Переваривание
углеводов в организме
Источником углеводов для организма служат углеводы пищи -

Переваривание углеводов
Амилаза слюны расщепляет α-1,4-гликозидные связи в крахмале. В ротовой полости
Переваривание углеводов
Амилаза слюны расщепляет α-1,4-гликозидные связи в крахмале. В ротовой полости

Переваривание углеводов
Последующее переваривание нерасщепленного или частично расщепленного крахмала происходит в кишечнике.
Переваривание углеводов
Последующее переваривание нерасщепленного или частично расщепленного крахмала происходит в кишечнике.

Переваривание углеводов
Мальтоза и изомальтоза вместе с другими пищевыми дисахаридами - сахарозой
Переваривание углеводов
Мальтоза и изомальтоза вместе с другими пищевыми дисахаридами - сахарозой
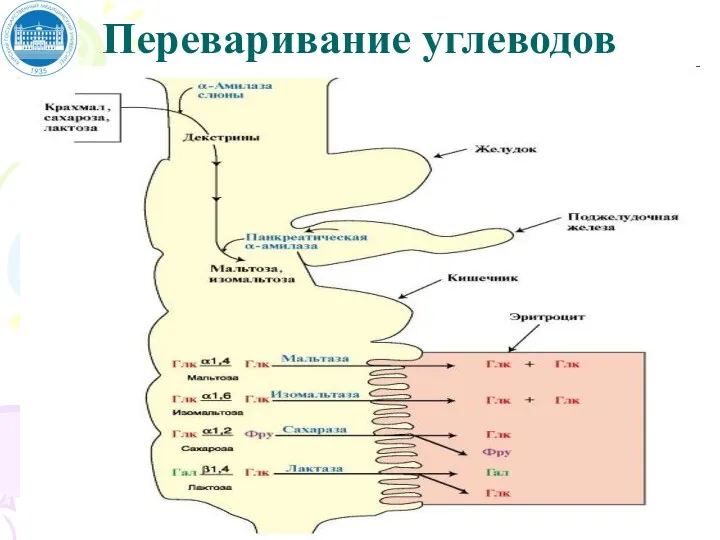
Переваривание углеводов
Переваривание углеводов

Продукты полного переваривания углеводов – глюкоза, галактоза и фруктоза –
Продукты полного переваривания углеводов – глюкоза, галактоза и фруктоза –

Гликолиз
Гликолиз (от греч. glykys – сладкий, lysys – распад )
Гликолиз
Гликолиз (от греч. glykys – сладкий, lysys – распад )

Гликолиз включает превращения трех разных типов:
Распад углеродного скелета глюкозы с
Гликолиз включает превращения трех разных типов:
Распад углеродного скелета глюкозы с
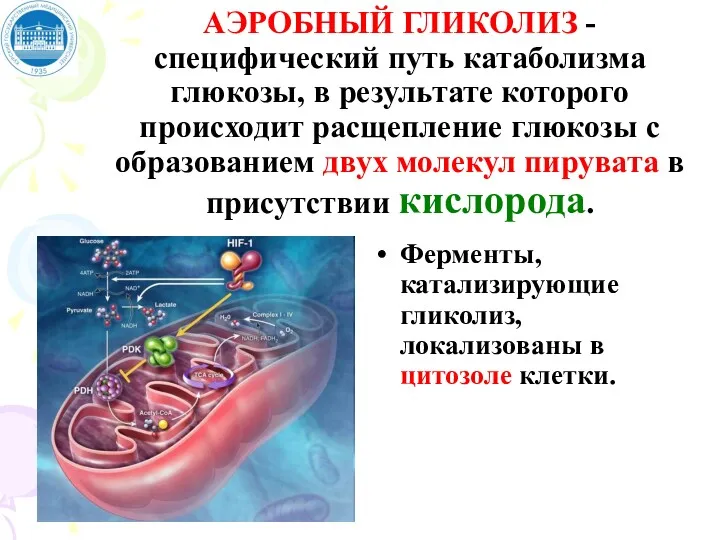
АЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит
АЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит
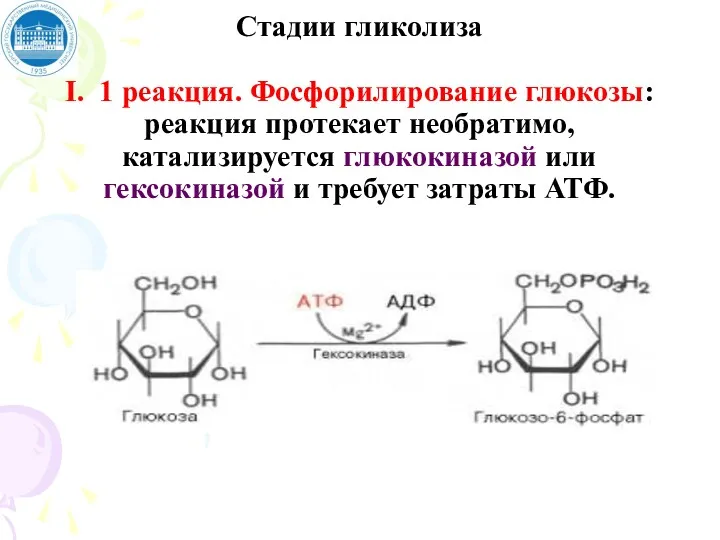
Стадии гликолиза
I. 1 реакция. Фосфорилирование глюкозы: реакция протекает необратимо, катализируется глюкокиназой
Стадии гликолиза I. 1 реакция. Фосфорилирование глюкозы: реакция протекает необратимо, катализируется глюкокиназой

Ферменты участвующие в фосфорилировании глюкозы.
Связывание гексокиназы с гексозой происходит по типу
Ферменты участвующие в фосфорилировании глюкозы.
Связывание гексокиназы с гексозой происходит по типу
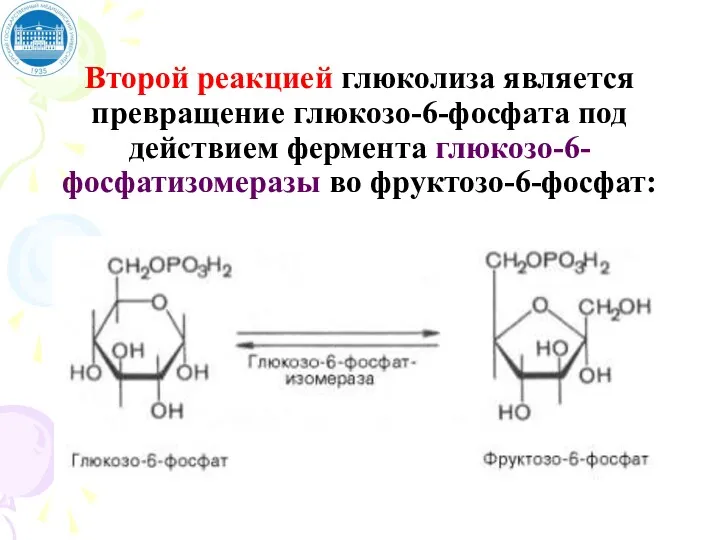
Второй реакцией глюколиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
Второй реакцией глюколиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
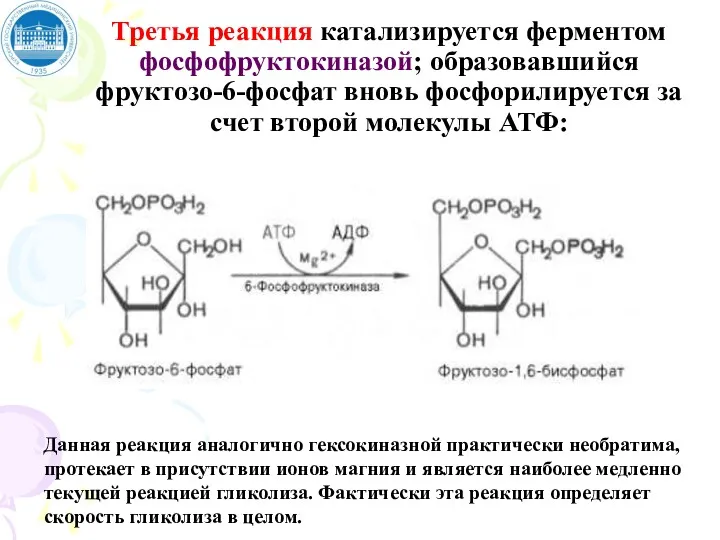
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
Данная реакция
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
Данная реакция
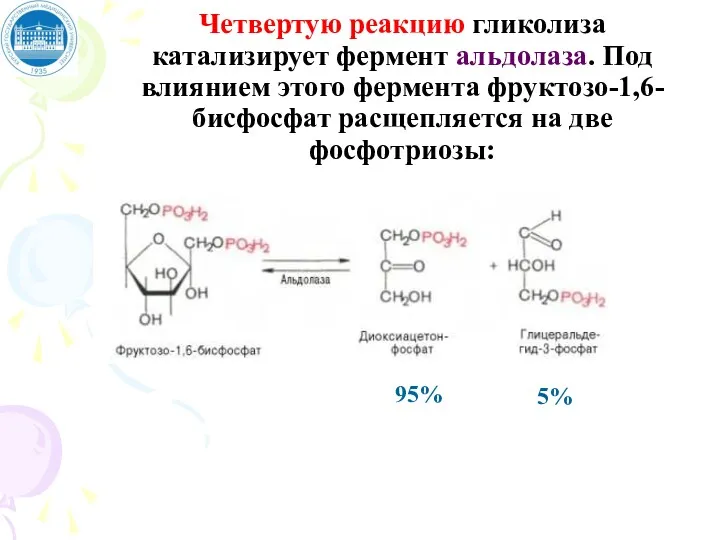
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат
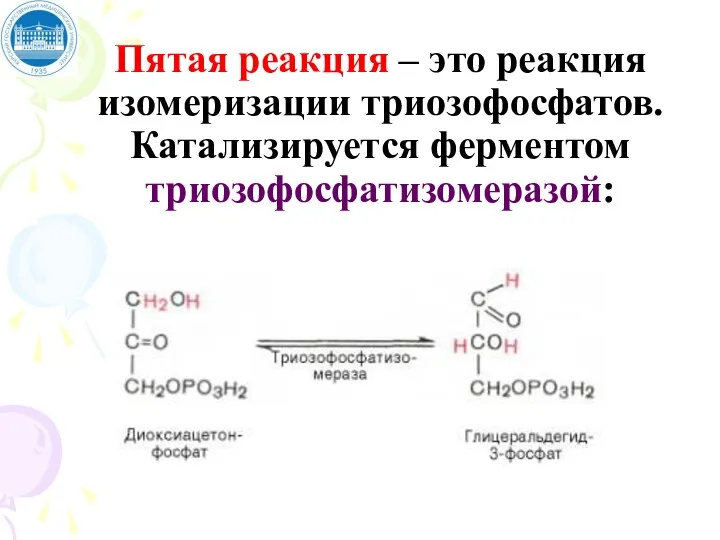
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
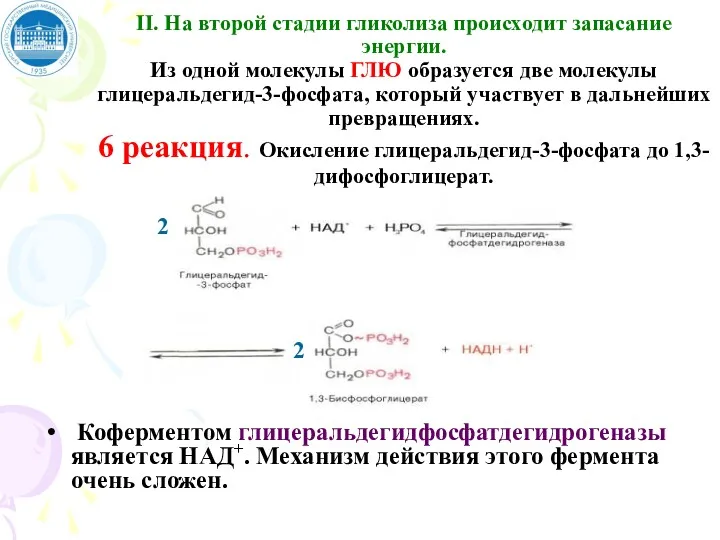
II. На второй стадии гликолиза происходит запасание энергии.
Из одной молекулы ГЛЮ
II. На второй стадии гликолиза происходит запасание энергии. Из одной молекулы ГЛЮ
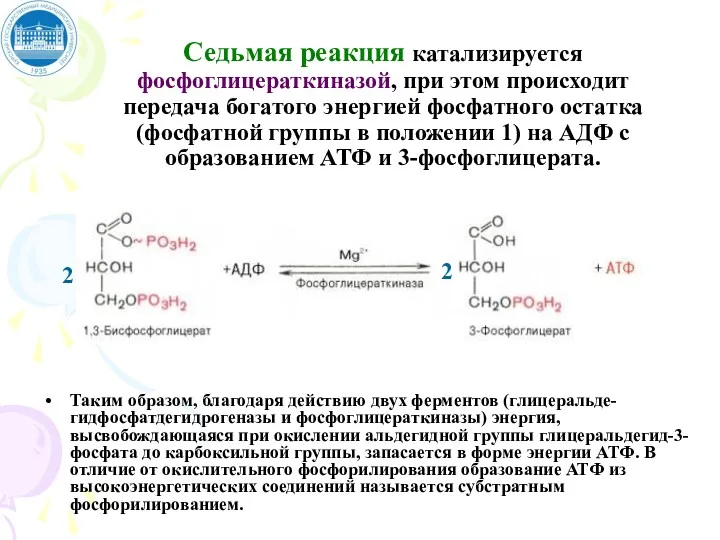
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного
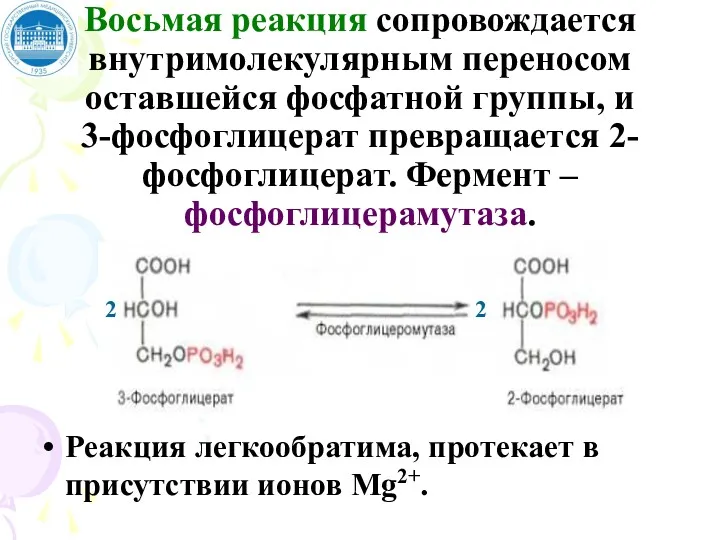
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и
3-фосфоглицерат превращается
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицерат превращается
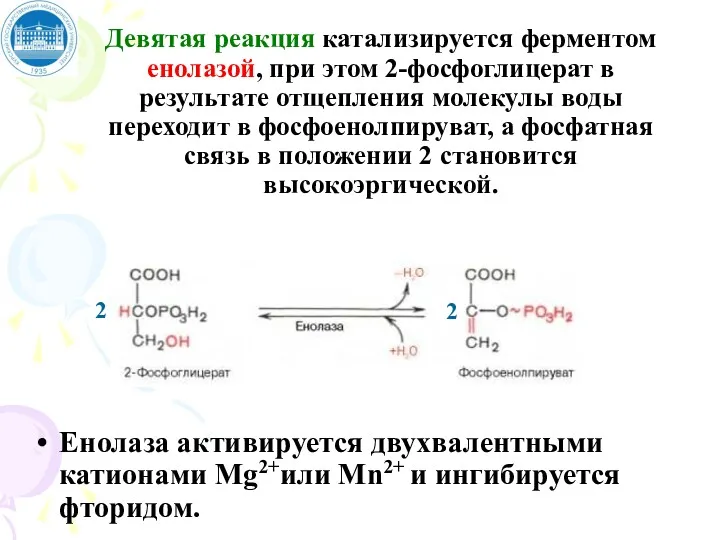
Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицерат в результате отщепления
Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицерат в результате отщепления
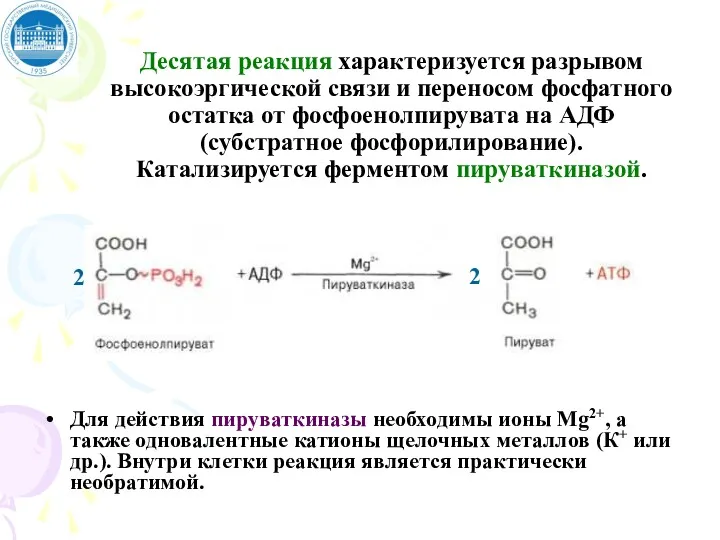
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от

Конечным продуктом аэробного гликолиза является
пируват, а энергетический баланс складывается
Конечным продуктом аэробного гликолиза является
пируват, а энергетический баланс складывается
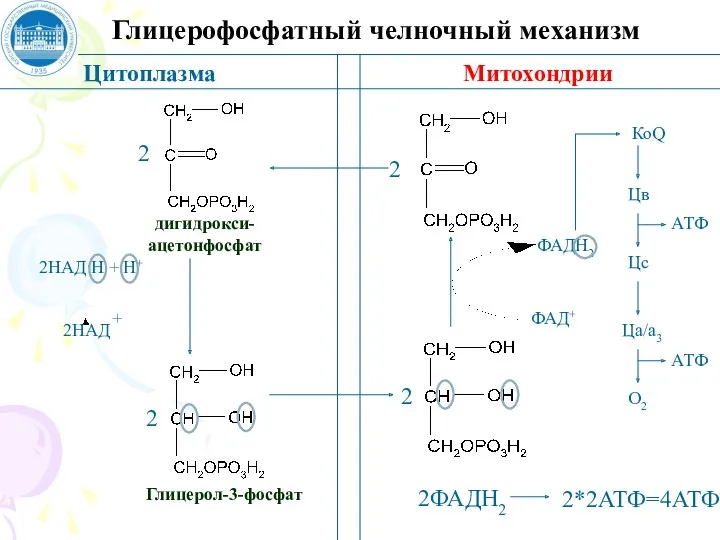
Глицерофосфатный челночный механизм
Цитоплазма
Митохондрии
дигидрокси-
ацетонфосфат
Глицерол-3-фосфат
2
2
2НАД·Н + Н+
2НАД
+
ФАД+
ФАДН2
2
2
КоQ
Цв
Цс
Ца/а3
O2
АТФ
АТФ
2ФАДН2
2*2АТФ=4АТФ
Глицерофосфатный челночный механизм
Цитоплазма
Митохондрии
дигидрокси-
ацетонфосфат
Глицерол-3-фосфат
2
2
2НАД·Н + Н+
2НАД
+
ФАД+
ФАДН2
2
2
КоQ
Цв
Цс
Ца/а3
O2
АТФ
АТФ
2ФАДН2
2*2АТФ=4АТФ
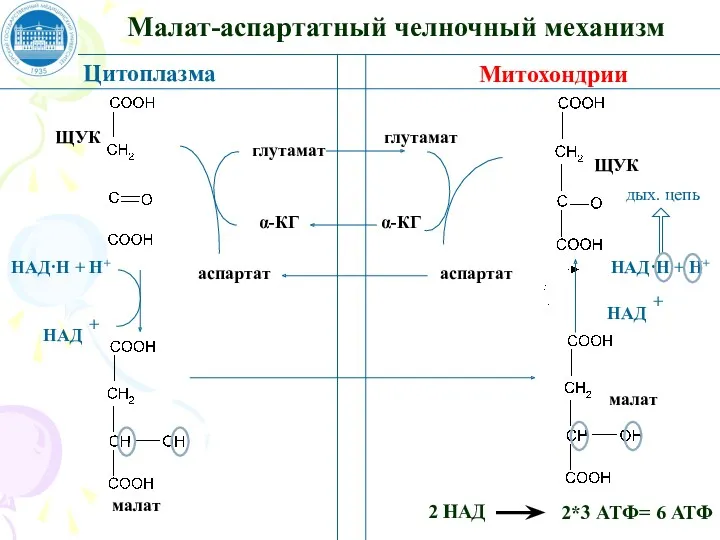
Малат-аспартатный челночный механизм
Цитоплазма
Митохондрии
ЩУК
малат
НАД·Н + Н+
НАД
+
глутамат
аспартат
глутамат
аспартат
ЩУК
НАД·Н + Н+
НАД
+
дых. цепь
малат
α-КГ
α-КГ
2 НАД
2*3
Малат-аспартатный челночный механизм
Цитоплазма
Митохондрии
ЩУК
малат
НАД·Н + Н+
НАД
+
глутамат
аспартат
глутамат
аспартат
ЩУК
НАД·Н + Н+
НАД
+
дых. цепь
малат
α-КГ
α-КГ
2 НАД
2*3

Баланс аэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6
Баланс аэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6
АНАЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит
АНАЭРОБНЫЙ ГЛИКОЛИЗ - специфический путь катаболизма глюкозы, в результате которого происходит

Дальше процесс идет в зависимости от наличия или отсутствия кислорода
Дальше процесс идет в зависимости от наличия или отсутствия кислорода

В этих условиях образовавшийся при гликолизе НАДН регенерируется за счет
В этих условиях образовавшийся при гликолизе НАДН регенерируется за счет

Баланс анаэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6
Баланс анаэробного гликолиза
1 реакция - 1 АТФ
3 реакция - 1 АТФ
6

Пентозофосфатный путь окисления глюкозы
Локализация: молочная железа, эмбриональная и жировая ткань,
Пентозофосфатный путь окисления глюкозы
Локализация: молочная железа, эмбриональная и жировая ткань,

В пентозофосфатном пути превращения глюкозы можно выделить:
Окислительный этап
Поставляет клеткам кофермент NADPH
В пентозофосфатном пути превращения глюкозы можно выделить:
Окислительный этап
Поставляет клеткам кофермент NADPH
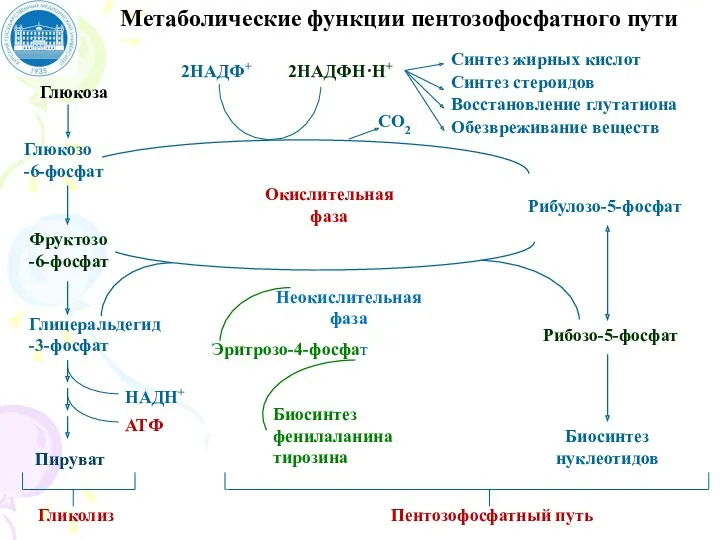
Глюкоза
Глюкозо
-6-фосфат
Фруктозо
-6-фосфат
Глицеральдегид-3-фосфат
Гликолиз
2НАДФ+
2НАДФН·Н+
Окислительная фаза
Синтез жирных кислот
Синтез стероидов
Восстановление глутатиона
Неокислительная фаза
Биосинтез
нуклеотидов
Рибулозо-5-фосфат
Рибозо-5-фосфат
Метаболические функции пентозофосфатного пути
СО2
Обезвреживание веществ
Пируват
НАДН+
АТФ
Пентозофосфатный
Глюкоза
Глюкозо
-6-фосфат
Фруктозо
-6-фосфат
Глицеральдегид-3-фосфат
Гликолиз
2НАДФ+
2НАДФН·Н+
Окислительная фаза
Синтез жирных кислот
Синтез стероидов
Восстановление глутатиона
Неокислительная фаза
Биосинтез
нуклеотидов
Рибулозо-5-фосфат
Рибозо-5-фосфат
Метаболические функции пентозофосфатного пути
СО2
Обезвреживание веществ
Пируват
НАДН+
АТФ
Пентозофосфатный

Условия протекания пентозофосфатного пути окисления глюкозы
Судьба глюкозо-6-фосфата — вступит ли он
Условия протекания пентозофосфатного пути окисления глюкозы
Судьба глюкозо-6-фосфата — вступит ли он
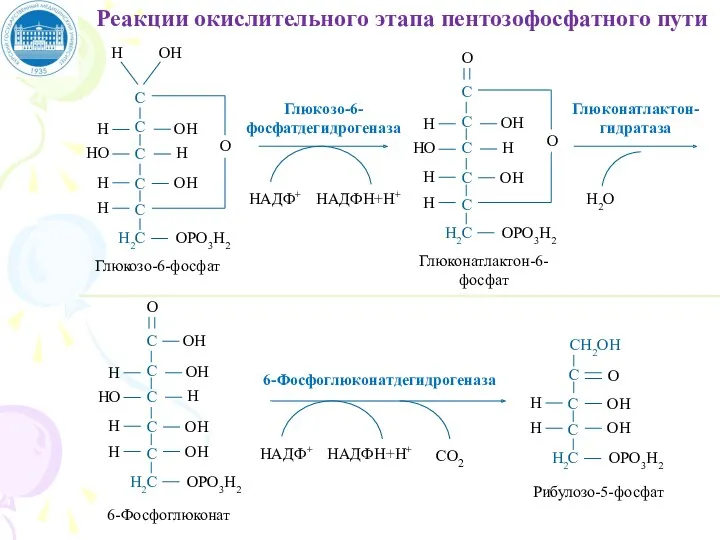
НАДФ+
НАДФН+Н+
Глюкозо-6-фосфатдегидрогеназа
Глюконатлактон-гидратаза
Н2О
НАДФ+
НАДФН+Н+
6-Фосфоглюконатдегидрогеназа
СО2
Реакции окислительного этапа пентозофосфатного пути
НАДФ+
НАДФН+Н+
Глюкозо-6-фосфатдегидрогеназа
Глюконатлактон-гидратаза
Н2О
НАДФ+
НАДФН+Н+
6-Фосфоглюконатдегидрогеназа
СО2
Реакции окислительного этапа пентозофосфатного пути
 Физиология микроорганизмов
Физиология микроорганизмов Разработка урока для 8 класса Пищеварение в ротовой полости
Разработка урока для 8 класса Пищеварение в ротовой полости Кабинет биологии
Кабинет биологии Возрастная физиология и анатомия
Возрастная физиология и анатомия Як комахи використовують різні знаряддя праці
Як комахи використовують різні знаряддя праці Приспособленность живых организмов к среде обитания
Приспособленность живых организмов к среде обитания Физиологические основы голода и насыщения. Пищеварение в полости рта. Лекция 1
Физиологические основы голода и насыщения. Пищеварение в полости рта. Лекция 1 Немембранные органоиды
Немембранные органоиды Игра В мире животных
Игра В мире животных Sistemul nervos
Sistemul nervos Эволюционное учение Ч. Дарвина
Эволюционное учение Ч. Дарвина Предмет и задачи генетики
Предмет и задачи генетики Многообразие паукообразных, их роль в природе
Многообразие паукообразных, их роль в природе Общий курс дрессировки
Общий курс дрессировки Материал для подготовки к ОГЭ по биологии для 9 класса
Материал для подготовки к ОГЭ по биологии для 9 класса Ядро, прокариоты
Ядро, прокариоты Типы конституций и телосложения человека. Влияние осанки на фигуру человека
Типы конституций и телосложения человека. Влияние осанки на фигуру человека Определение видов растений и животных
Определение видов растений и животных Сон и сновидения
Сон и сновидения Методы изучения наследственности человека
Методы изучения наследственности человека Значение опорно-двигательной системы. Строение костей. 8 класс
Значение опорно-двигательной системы. Строение костей. 8 класс Популяция. Основные характеристики популяций
Популяция. Основные характеристики популяций Остеоартросиндесмология. Виды соединения костей
Остеоартросиндесмология. Виды соединения костей Свойства белков. Преврщение белков в организме
Свойства белков. Преврщение белков в организме Тірі адамнан, практикалық маңызды, қаңқаның шығыңқы бөліктерінің орнын сипап анықтау
Тірі адамнан, практикалық маңызды, қаңқаның шығыңқы бөліктерінің орнын сипап анықтау Адаптации. Классификация адаптаций
Адаптации. Классификация адаптаций Строение и свойства белков
Строение и свойства белков Тип членистоногие. Класс паукообразные
Тип членистоногие. Класс паукообразные