- Основы сравнительной эмбриологии

Содержание
- 2. Закон эмбриональной дивергенции: общее для данного морфологического типа животных образуется в эмбриогенезе раньше, чем специальное (сначала
- 3. Эволюция животного мира: Тип развития – сложившийся в ходе эволюционного развития комплекс взаимосвязанных морфогенетических процессов, как
- 4. РАЗВИТИЕ ИГЛОКОЖИХ лекция 6 (1) Arbacia punctulata Lytechinus pictus
- 5. Раннее развитие: оплодотворение, дробление Дробление голобластическое, радиального типа. Оплодотворение наружное, моноспермное. 1-я и 2-я борозды –
- 6. Раннее развитие: бластула 128-клеточный зародыш находится на стадии бластулы. Бластула у морского ежа однослойна, содержит от
- 7. Раннее развитие: бластула Сопряженность увеличения поверхности бластулы и формирования бластоцеля: осуществляется под действием сил самого бластоцеля:
- 8. Начинается с выселения первичной мезенхимы, PMC (из микромеров, за счёт филоподий). Гаструляция у иглокожих (1) Базальная
- 9. Гаструляция у иглокожих (2) Миграция первичных мезенхимных клеток по внеклеточному матриксу (фибронектин). Клетки вегетативной пластинки движутся
- 10. Гаструляция у иглокожих (3) Вторая фаза инвагинации начинается после паузы. резкое удлинение архентерона (его длина утраивается
- 11. Гаструляция у иглокожих (4) Филоподии клеток вторичной мезенхимы, простираясь через жидкость, содержащуюся в бластоцеле, устанавливают контакт
- 12. Личиночная стадия и метаморфоз иглокожих Личинка конической формы – стадия призмы. сокращения филоподий также участвуют в
- 13. РАЗВИТИЕ ОБОЛОЧНИКОВ И БЕСЧЕРЕПНЫХ лекция 6 (2)
- 14. Развитие оболочников (1) Дробление у асцидий билатеральное: 1-я меридиональная борозда делит яйцо на симметричные правую и
- 15. Развитие оболочников (2) инвагинация: центральные клетки вегетативного полушария (энтодерма); инволюция: клетки по периферии энтодермы зачатка (хордо-мезодерма);
- 16. Развитие ланцетника (1) Олиголецитальные яйцеклетки ланцетника изолецитального типа (~ 100 мкм). Дробление: полное почти равномерное радиальное
- 17. Развитие ланцетника (2) Гаструляция путем инвагинации. разница темпов деления клеток в краевой зоне и в вегетативной
- 18. Развитие ланцетника (3) Стадия нейрулы – дифференцировка зародышевых листков и закладка осевых органов. Нейруляция – смыкание
- 20. Скачать презентацию

Закон эмбриональной дивергенции: общее для данного морфологического типа животных образуется в
Закон эмбриональной дивергенции: общее для данного морфологического типа животных образуется в
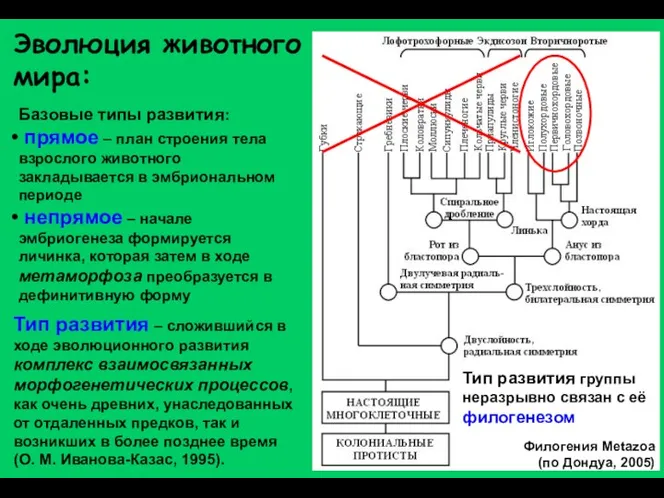
Эволюция животного мира:
Тип развития – сложившийся в ходе эволюционного развития комплекс
Эволюция животного мира:
Тип развития – сложившийся в ходе эволюционного развития комплекс

РАЗВИТИЕ ИГЛОКОЖИХ
лекция 6 (1)
Arbacia punctulata
Lytechinus pictus
РАЗВИТИЕ ИГЛОКОЖИХ
лекция 6 (1)
Arbacia punctulata
Lytechinus pictus

Раннее развитие: оплодотворение, дробление
Дробление голобластическое, радиального типа.
Оплодотворение наружное, моноспермное.
1-я и
Раннее развитие: оплодотворение, дробление
Дробление голобластическое, радиального типа.
Оплодотворение наружное, моноспермное.
1-я и

Раннее развитие: бластула
128-клеточный зародыш находится на стадии бластулы.
Бластула у морского ежа
Раннее развитие: бластула
128-клеточный зародыш находится на стадии бластулы.
Бластула у морского ежа

Раннее развитие: бластула
Сопряженность увеличения поверхности бластулы и формирования бластоцеля:
осуществляется под
Раннее развитие: бластула
Сопряженность увеличения поверхности бластулы и формирования бластоцеля:
осуществляется под
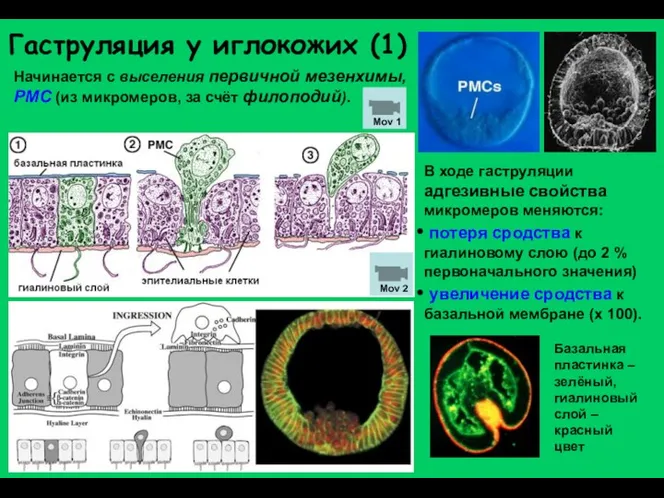
Начинается с выселения первичной мезенхимы, PMC (из микромеров, за счёт филоподий).
Гаструляция
Начинается с выселения первичной мезенхимы, PMC (из микромеров, за счёт филоподий).
Гаструляция

Гаструляция у иглокожих (2)
Миграция первичных мезенхимных клеток по внеклеточному матриксу (фибронектин).
Клетки
Гаструляция у иглокожих (2)
Миграция первичных мезенхимных клеток по внеклеточному матриксу (фибронектин).
Клетки
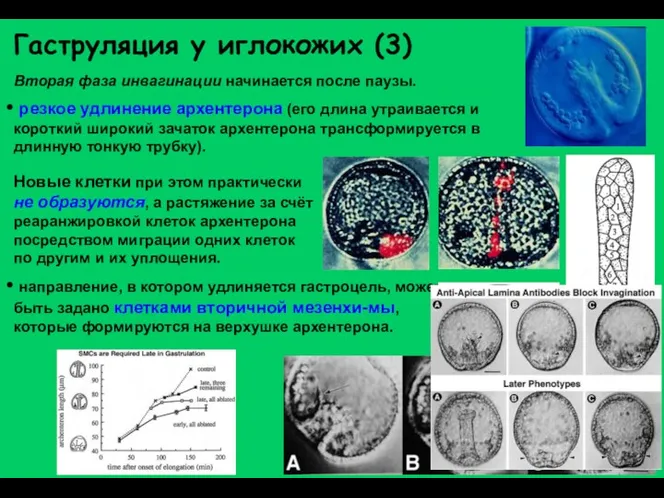
Гаструляция у иглокожих (3)
Вторая фаза инвагинации начинается после паузы.
резкое
Гаструляция у иглокожих (3)
Вторая фаза инвагинации начинается после паузы.
резкое
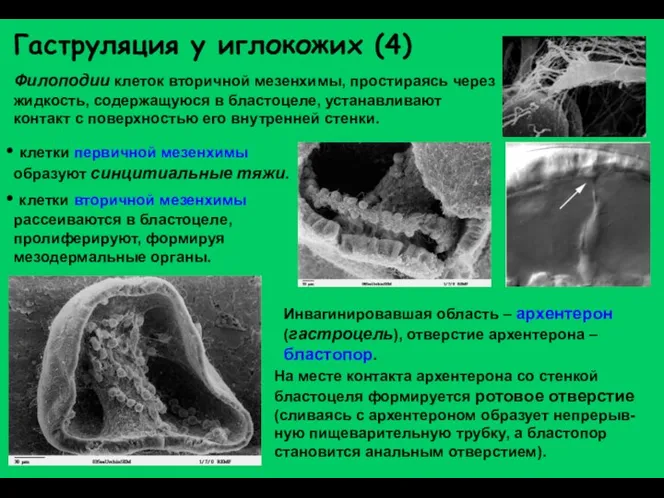
Гаструляция у иглокожих (4)
Филоподии клеток вторичной мезенхимы, простираясь через жидкость, содержащуюся
Гаструляция у иглокожих (4)
Филоподии клеток вторичной мезенхимы, простираясь через жидкость, содержащуюся

Личиночная стадия и метаморфоз иглокожих
Личинка конической формы – стадия призмы.
сокращения
Личиночная стадия и метаморфоз иглокожих
Личинка конической формы – стадия призмы.
сокращения

РАЗВИТИЕ ОБОЛОЧНИКОВ И БЕСЧЕРЕПНЫХ
лекция 6 (2)
РАЗВИТИЕ ОБОЛОЧНИКОВ И БЕСЧЕРЕПНЫХ
лекция 6 (2)
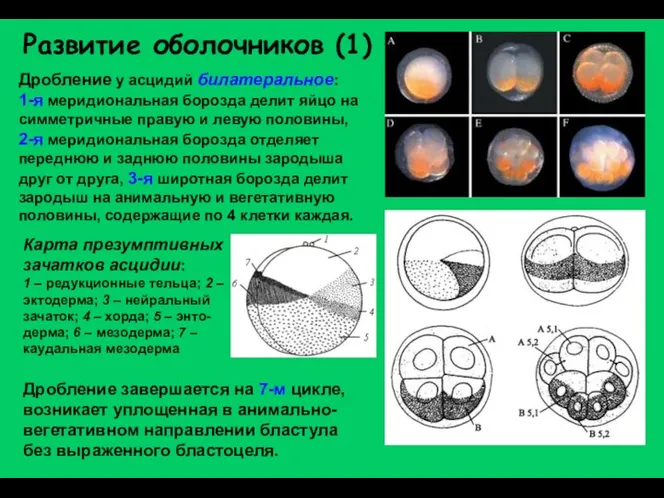
Развитие оболочников (1)
Дробление у асцидий билатеральное:
1-я меридиональная борозда делит яйцо на
Развитие оболочников (1)
Дробление у асцидий билатеральное:
1-я меридиональная борозда делит яйцо на
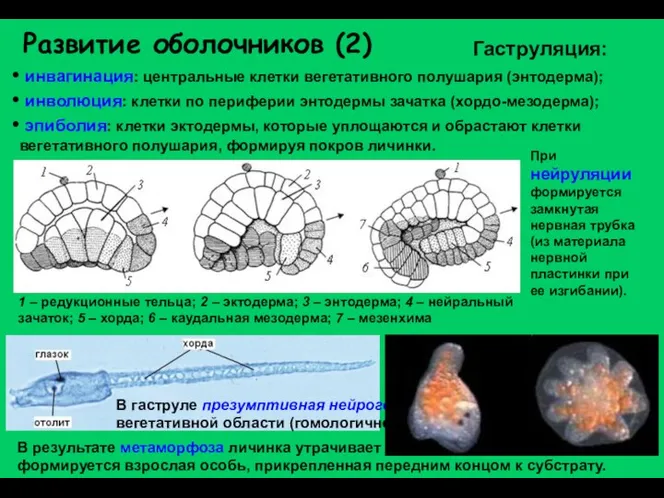
Развитие оболочников (2)
инвагинация: центральные клетки вегетативного полушария (энтодерма);
инволюция: клетки
Развитие оболочников (2)
инвагинация: центральные клетки вегетативного полушария (энтодерма);
инволюция: клетки
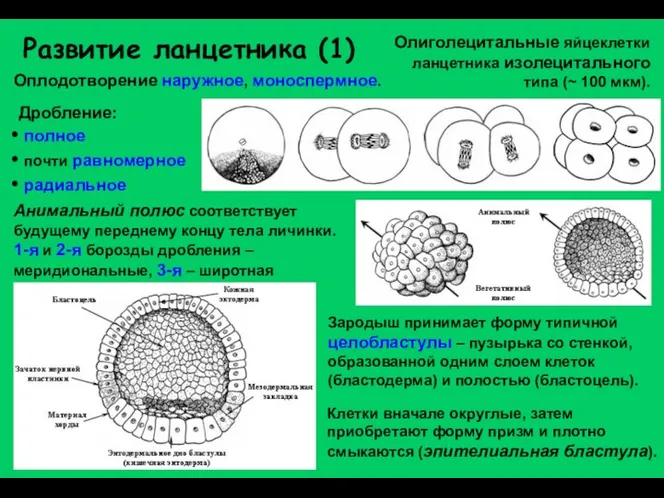
Развитие ланцетника (1)
Олиголецитальные яйцеклетки ланцетника изолецитального типа (~ 100 мкм).
Дробление:
полное
почти
Развитие ланцетника (1)
Олиголецитальные яйцеклетки ланцетника изолецитального типа (~ 100 мкм).
Дробление:
полное
почти
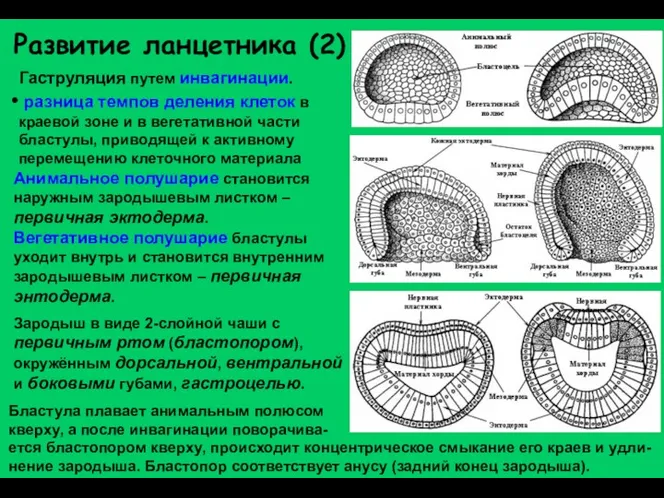
Развитие ланцетника (2)
Гаструляция путем инвагинации.
разница темпов деления клеток в краевой
Развитие ланцетника (2)
Гаструляция путем инвагинации.
разница темпов деления клеток в краевой
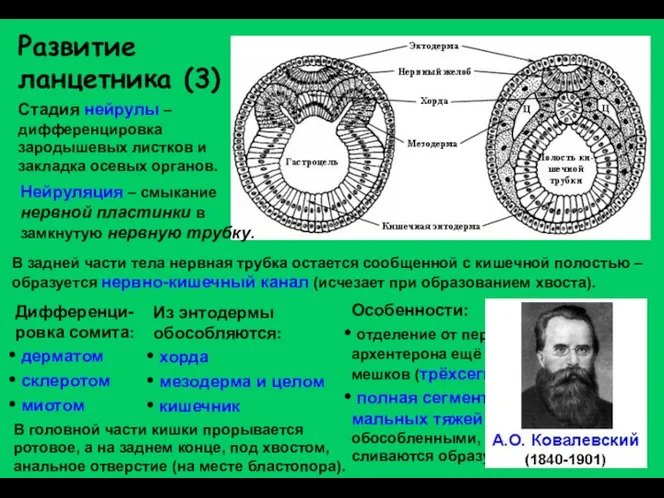
Развитие ланцетника (3)
Стадия нейрулы – дифференцировка зародышевых листков и закладка осевых
Развитие ланцетника (3)
Стадия нейрулы – дифференцировка зародышевых листков и закладка осевых
 Разнообразие растений. (3 класс)
Разнообразие растений. (3 класс) Животный мир Пермского края
Животный мир Пермского края Простейшие. Многообразие, среда обитания
Простейшие. Многообразие, среда обитания Грибы в биосфере и жизни человека
Грибы в биосфере и жизни человека Биохимическая эволюция. Теория Опарина-Холдейна
Биохимическая эволюция. Теория Опарина-Холдейна Селекція ембріонів за статтю
Селекція ембріонів за статтю Презентация: Борьба организма с инфекцией. Иммунитет
Презентация: Борьба организма с инфекцией. Иммунитет Микробиология, как наука. Морфология и ультраструктура бактерий
Микробиология, как наука. Морфология и ультраструктура бактерий Перенос дыхательных газов кровью
Перенос дыхательных газов кровью Красная книга Казахстана
Красная книга Казахстана Водородная химическая связь
Водородная химическая связь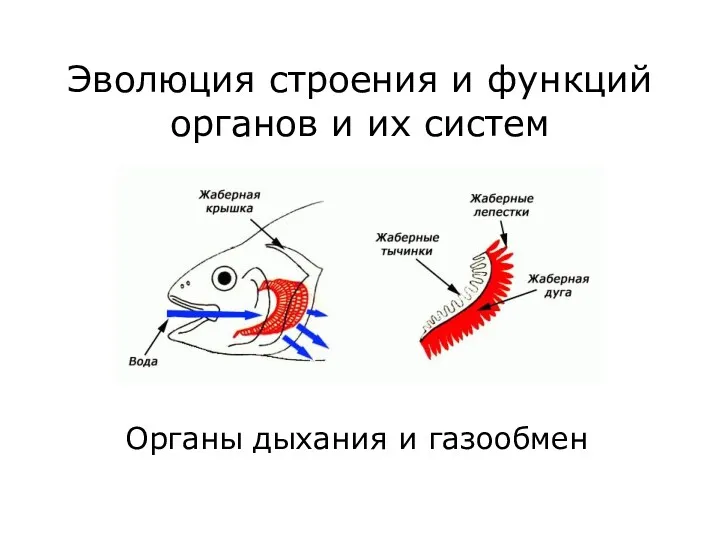 Органы дыхания и газообмен
Органы дыхания и газообмен Леса Белгородской области
Леса Белгородской области Внешнее строение птиц, и строение скелета
Внешнее строение птиц, и строение скелета Дельфины
Дельфины Australian Wildlife
Australian Wildlife Прокаріотичні організми: бактерії. Особливості їхньої організації та функціонування
Прокаріотичні організми: бактерії. Особливості їхньої організації та функціонування Проводящие ткани у растений
Проводящие ткани у растений Многообразие птиц
Многообразие птиц Эндокринная система
Эндокринная система Сущность жизни и свойства живого
Сущность жизни и свойства живого Evolution 3 – Genetic Variation
Evolution 3 – Genetic Variation Биотические связи в природе
Биотические связи в природе Общая характеристика типа Моллюски
Общая характеристика типа Моллюски Влияние витаминов на здоровье человека
Влияние витаминов на здоровье человека Развитие биологии в додарвиновский период. Работы К. Линнея. (11 класс)
Развитие биологии в додарвиновский период. Работы К. Линнея. (11 класс) Размножение живых организмов. Для подготовки к ЕГЭ
Размножение живых организмов. Для подготовки к ЕГЭ Учебный проект Что в образе тебе моём?
Учебный проект Что в образе тебе моём?