- Перенос дыхательных газов кровью

Содержание
- 2. Наиболее важные функции крови Транспорт питательных веществ от пищеварительного тракта к тканям и местам накопления (жировой
- 3. Перенос О2 кровью или гемолимфой в виде растворенного газа (многие беспозвоночные) с помощью дыхательных пигментов, способных
- 4. Молекулярные веса и локализация ( в клетках или в плазме крови) дыхательных пигментов у различных животных
- 5. Упаковка гемоглобина (Hb) в клетку – эритроцит В плазме крови на связь О2 с Hb влияют
- 6. Влияние содержания гемоглобина на вязкость крови Представлено отношение вязкости крови к вязкости чистой воды (принятой за
- 7. ? Размеры эритроцитов ? У саламандр эритроциты в 100-200 раз больше, чем у млекопитающих (3.5-10 мкм).
- 8. У человека средний диаметр эритроцита 7.5 мкм. Как и у других животных, размеры эритроцитов варьируют (6–9
- 9. ? Наличие ядра в эритроцитах? У млекопитающих – безъядерные У др. позвоночных – с ядрами Myodes
- 10. Кривые кислородной диссоциации Hb + O2 → HbO2 Utvjukj,by гемоглобин оксигемоглобин > %O2 → Если %O2
- 11. Кривые кислородной диссоциации для крови голубя I – кривая, полученная при нормальных для организма птицы условиях:
- 12. Разнообразие кривых КД: А. Больше сродство к кислороду (= Hb меньше отдает О2) – сдвиг влево
- 13. Кривые кислородной диссоциации у сусликов, находящихся в обычном состоянии (T b = 38° C) и в
- 14. Кривые кислородной диссоциации для крови голубя (Lutz et al.,1973) I – кривая, полученная при нормальных для
- 15. Связь между величиной эффекта Бора (сдвиг Р50 при ∆pH=1) и размерами тела. Гемоглобин мелких животных обнаруживает
- 16. Кривые кислородной диссоциации для крови рыбы Opsanus при 15оС, I – кривая для цельной крови в
- 17. Влияние органических фосфатов Раствор чистого Нb → > сродства к О2, чем у цельной крови. Если
- 18. Кривые кислородной диссоциации для крови беременной козы и плода Более высокое сродство к кислороду у крови
- 19. После рождения фетальный Нb исчезает и замещается Нb взрослого. Сходный эффект (сдвиг КД влево) – у
- 20. Кривые кислородной диссоциации для крови ламы и викуньи располагаются слева от кривых для других млекопитающих (заштрихованная
- 21. Сродство к О2 играет роль 1) В легких, где Нb связывается с О2 2) В тканях,
- 22. Кривые кислородной диссоциации для крови млекопитающих разных размеров (Schmidt-Nielsen, 1972). У мелких млекопитающих сродство к кислороду
- 23. Вокруг каждого капилляра можно выделить приблизительно цилиндрическую область, в которой ткань получает кислород путем диффузии из
- 24. В реальности: Мышь Лошадь 1) 2000 капилляров/мм2 2) P50 > в 2.5 раз Т.е. оба пути
- 25. Диффузия кислорода и азота в вакуум через слой плазмы крови всегда осуществляется в соотношении 0,5:1. Относительная
- 26. Другие пигменты: Гемоцианин не ускоряет диффузию (очень большие молекулы). ~ Механизм облегчения диффузии О2 связан с
- 27. Увеличение %Hb в тканях беспозвоночных, обитающих в местах с нехваткой О2. Нематоды в кишечнике овцы дышат
- 28. Эволюционный аспект распространения Нb среди представителей разных таксонов
- 29. Основа гема – порфириновое ядро – есть в молекулах ферментов–цитохромов у всех растений и животных. Небольшие
- 30. КД у беспозвоночных У многих – высокое сродство к О2. Р50 Дв. моллюск Phacoides 0.2 мм
- 31. Кривые кислородной диссоциации для гемоэритрина из крови и целомической жидкости сипункулиды Dendrostomum zostericolum (Manwell, 1960). Сипункули́ды
- 32. Кривые кислородной диссоциации для гемоцианина (из крови) и миоглобина (из мышц радулы) боконервного моллюска Cryptochilon (Manwell,
- 33. Перенос СО2 Кровь в тканях – отдает О2 и забирает СО2; Кровь в легких – отдает
- 34. ∑ СО2 в крови >>> CO2, растворенного в жидкости при данном РCO2 . Большая часть диоксида
- 35. Уравнение Хендерсона-Хассельбаха: [H+][A-]/[HA] = K → [H+] = K [HA] /[A-] →[-log] → pH = pK
- 36. ∑ СО2 в крови >>> CO2, растворенного в жидкости при данном РCO2 . Большая часть диоксида
- 37. Важнейшие буферные вещества крови: Система угольная кислота ↔ бикарбонат Фосфаты Белки (содержат группы, которые могут диссоциировать
- 38. Kривые Диссоциации (КД) для СО2 Количество СО2, поглощаемого кровью = F (PCO2) → можно построить КД,
- 39. Кривые диссоциации для СО2, полученные в опытах с кровью млекопитающих (Winton, Bayliss, 1955). Для оксигенированной и
- 40. СО2 при водном дыхании RQ = СО2 /O2 = lim [0.7-1.0] Пусть RQ =1 1) На
- 41. Соотношение между РО2 и РСО2 в воздухе и воде в случае, когда дыхательный коэффициент (ДК) равен
- 42. Концентрация СО2 в крови и рН Принципиальное различие водных и сухопутных животных по % СО2 в
- 43. Величины pH крови позвоночных (представителей всех классов), измеренные при нормальной для каждого животного температуре тела. Штриховкой
- 44. CO2 + H2O ↔ H2CO3 ↔ H+ + HCO3- ↔ [H+ + CO32-] минуты-секунды доли секунды
- 45. Активность карбоангидразы в крови млекопитающих связана с размерами тела: она тем выше, чем меньше животное
- 46. Когда CO2 попадает в кровь, КАРБОАНГИДРАЗА обеспечивает мгновенное образование угольной кислоты. карбоангидраза + CO2 + H2O
- 47. Концентрация О2 в среде и интенсивность метаболизма У млекопитающих МR покоя практически не меняется даже при
- 48. Потребление кислорода у рыб зависит от его парциального давления (РО2) в воде. У иглобрюха и скапа
- 49. При низких концентрациях кислорода его потребление золотой рыбкой прямо пропорционально содержанию его в воде, но в
- 51. Скачать презентацию

Наиболее важные функции крови
Транспорт питательных веществ от пищеварительного тракта к
Наиболее важные функции крови
Транспорт питательных веществ от пищеварительного тракта к
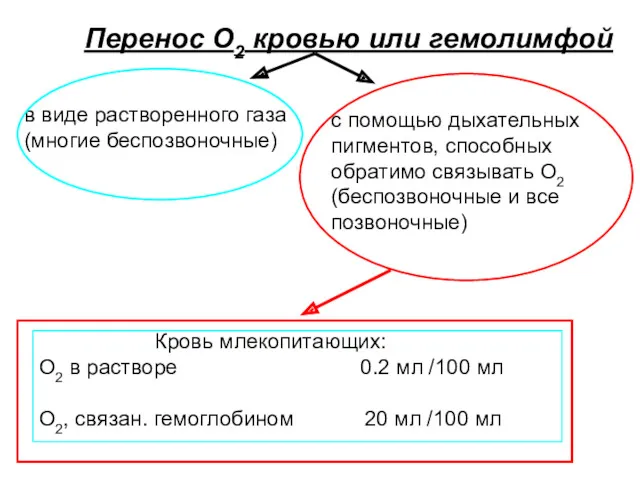
Перенос О2 кровью или гемолимфой
в виде растворенного газа
(многие беспозвоночные)
с помощью дыхательных
Перенос О2 кровью или гемолимфой
в виде растворенного газа
(многие беспозвоночные)
с помощью дыхательных

Молекулярные веса и локализация ( в клетках или в плазме крови)
дыхательных
Молекулярные веса и локализация ( в клетках или в плазме крови)
дыхательных

Упаковка гемоглобина (Hb) в клетку – эритроцит
В плазме крови на
Упаковка гемоглобина (Hb) в клетку – эритроцит
В плазме крови на
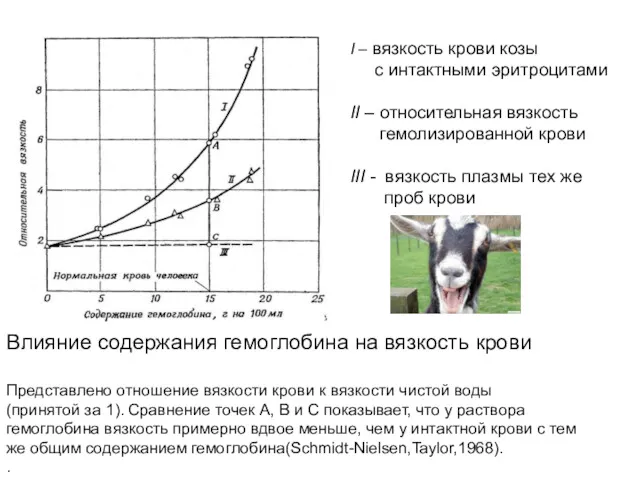
Влияние содержания гемоглобина на вязкость крови
Представлено отношение вязкости крови к
Влияние содержания гемоглобина на вязкость крови
Представлено отношение вязкости крови к

? Размеры эритроцитов ?
У саламандр эритроциты в 100-200 раз больше,
? Размеры эритроцитов ?
У саламандр эритроциты в 100-200 раз больше,

У человека средний диаметр эритроцита 7.5 мкм. Как и у других
У человека средний диаметр эритроцита 7.5 мкм. Как и у других
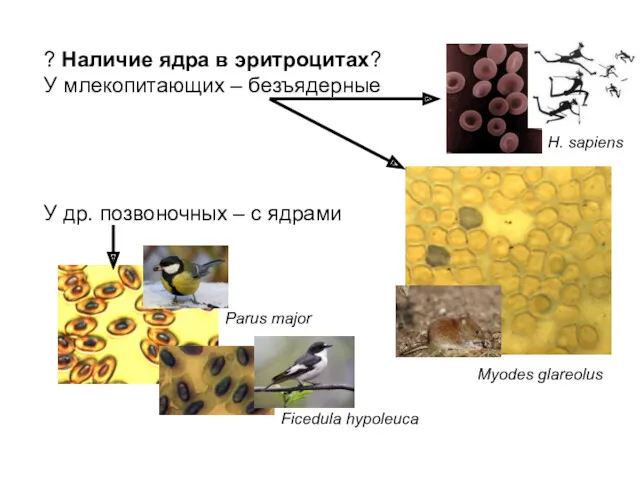
? Наличие ядра в эритроцитах?
У млекопитающих – безъядерные
У др. позвоночных
? Наличие ядра в эритроцитах?
У млекопитающих – безъядерные
У др. позвоночных
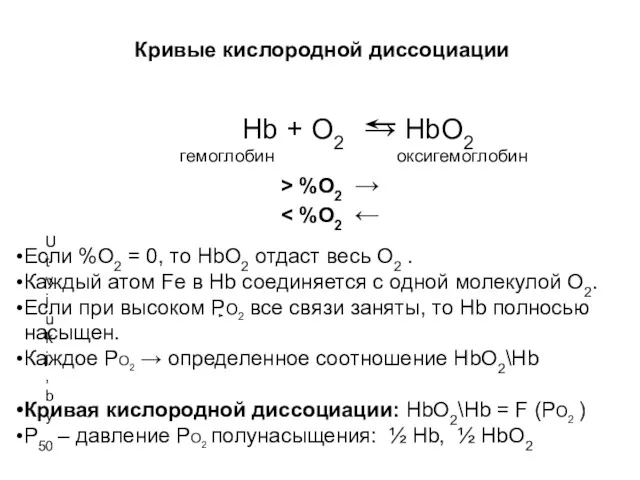
Кривые кислородной диссоциации
Hb + O2 → HbO2
Utvjukj,by
гемоглобин
оксигемоглобин
> %O2 →
< %O2
Кривые кислородной диссоциации
Hb + O2 → HbO2
Utvjukj,by
гемоглобин
оксигемоглобин
> %O2 →
< %O2
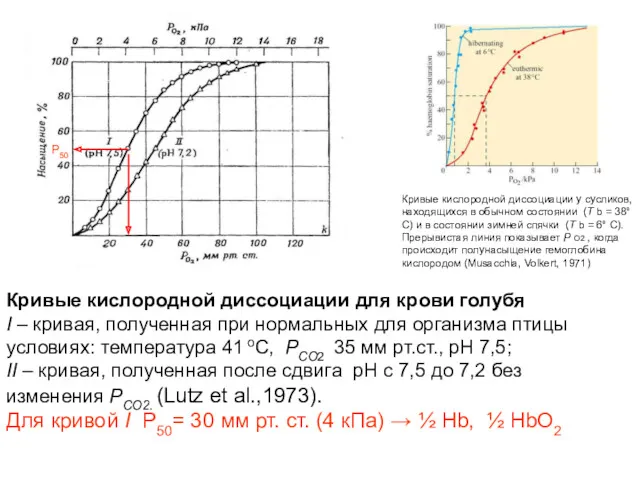
Кривые кислородной диссоциации для крови голубя
I – кривая, полученная при
Кривые кислородной диссоциации для крови голубя
I – кривая, полученная при

Разнообразие кривых КД:
А. Больше сродство к кислороду
(=
Разнообразие кривых КД:
А. Больше сродство к кислороду
(=
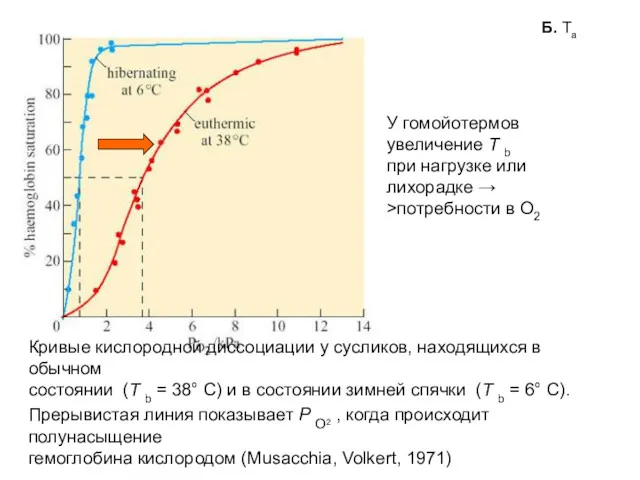
Кривые кислородной диссоциации у сусликов, находящихся в обычном
состоянии (T b =
Кривые кислородной диссоциации у сусликов, находящихся в обычном
состоянии (T b =
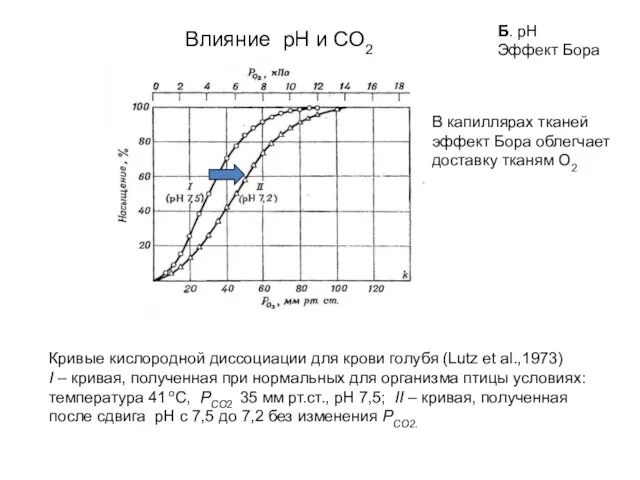
Кривые кислородной диссоциации для крови голубя (Lutz et al.,1973)
I – кривая,
Кривые кислородной диссоциации для крови голубя (Lutz et al.,1973)
I – кривая,
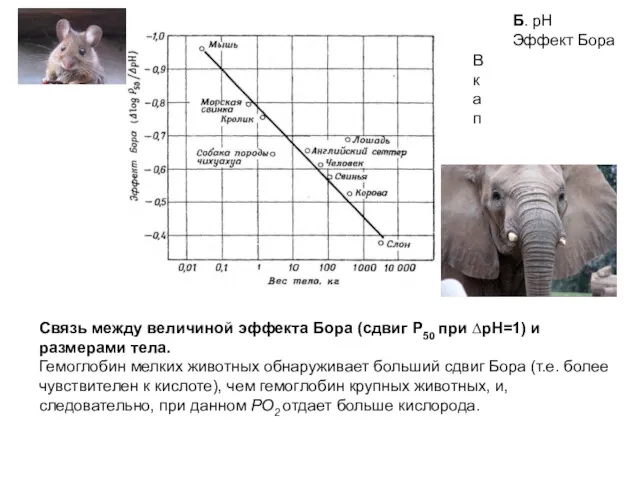
Связь между величиной эффекта Бора (сдвиг Р50 при ∆pH=1) и размерами
Связь между величиной эффекта Бора (сдвиг Р50 при ∆pH=1) и размерами

Кривые кислородной диссоциации для крови рыбы Opsanus при 15оС,
I – кривая
Кривые кислородной диссоциации для крови рыбы Opsanus при 15оС,
I – кривая

Влияние органических фосфатов
Раствор чистого Нb → > сродства к О2,
Влияние органических фосфатов
Раствор чистого Нb → > сродства к О2,
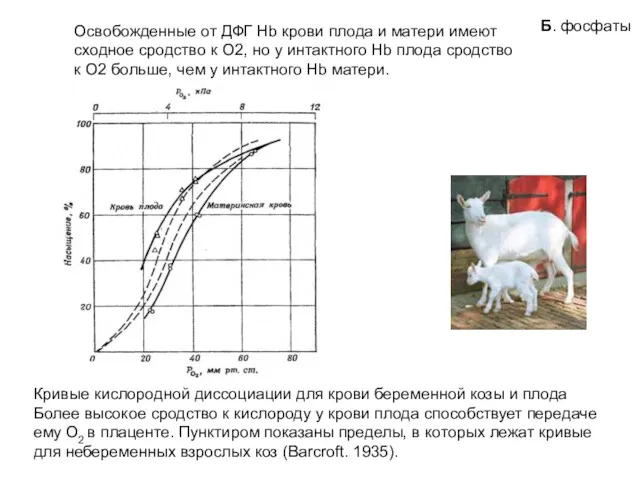
Кривые кислородной диссоциации для крови беременной козы и плода
Более высокое сродство
Кривые кислородной диссоциации для крови беременной козы и плода
Более высокое сродство

После рождения фетальный Нb исчезает и замещается Нb
взрослого. Сходный эффект
После рождения фетальный Нb исчезает и замещается Нb
взрослого. Сходный эффект

Кривые кислородной диссоциации для крови ламы и викуньи располагаются
слева от
Кривые кислородной диссоциации для крови ламы и викуньи располагаются
слева от
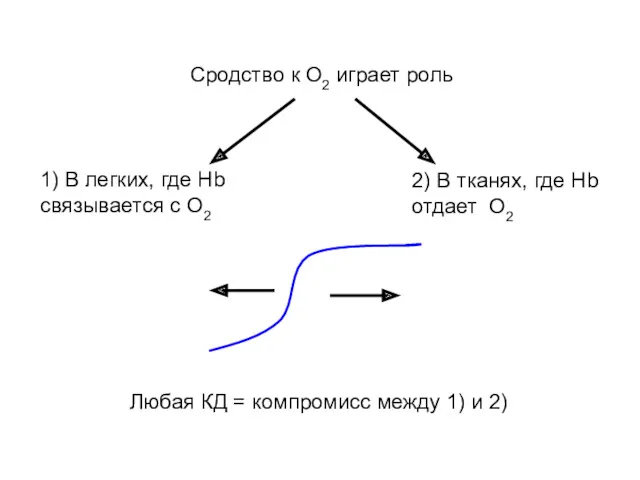
Сродство к О2 играет роль
1) В легких, где Нb связывается с
Сродство к О2 играет роль
1) В легких, где Нb связывается с
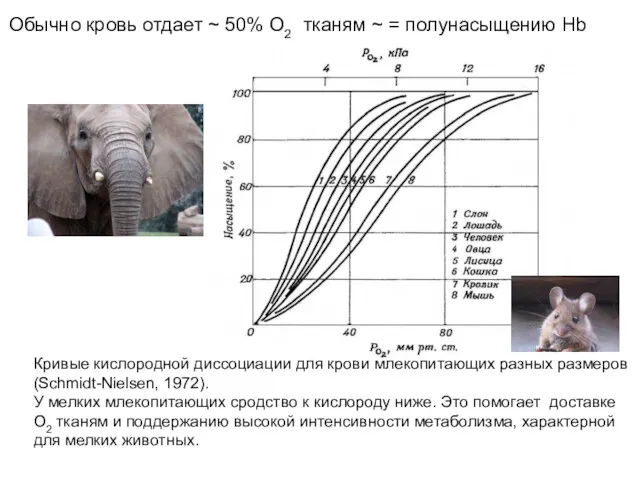
Кривые кислородной диссоциации для крови млекопитающих разных размеров
(Schmidt-Nielsen, 1972).
У мелких
Кривые кислородной диссоциации для крови млекопитающих разных размеров
(Schmidt-Nielsen, 1972).
У мелких
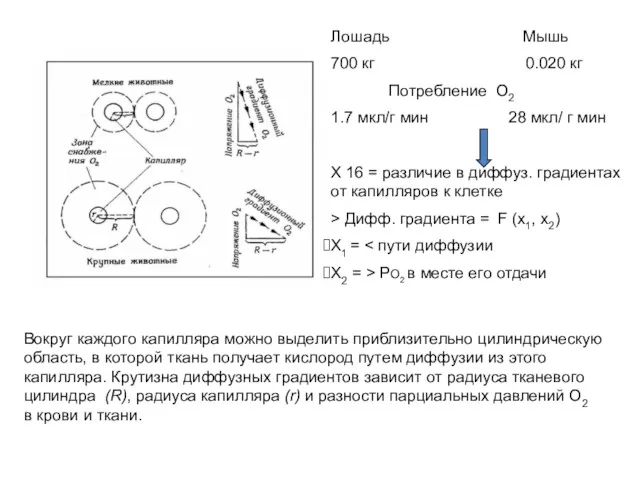
Вокруг каждого капилляра можно выделить приблизительно цилиндрическую
область, в которой ткань получает
Вокруг каждого капилляра можно выделить приблизительно цилиндрическую
область, в которой ткань получает
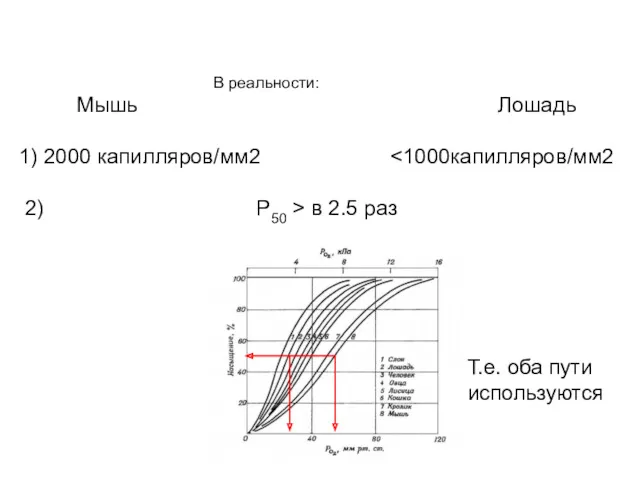
В реальности:
Мышь Лошадь
1) 2000 капилляров/мм2 <1000капилляров/мм2
2)
В реальности:
Мышь Лошадь
1) 2000 капилляров/мм2 <1000капилляров/мм2
2)
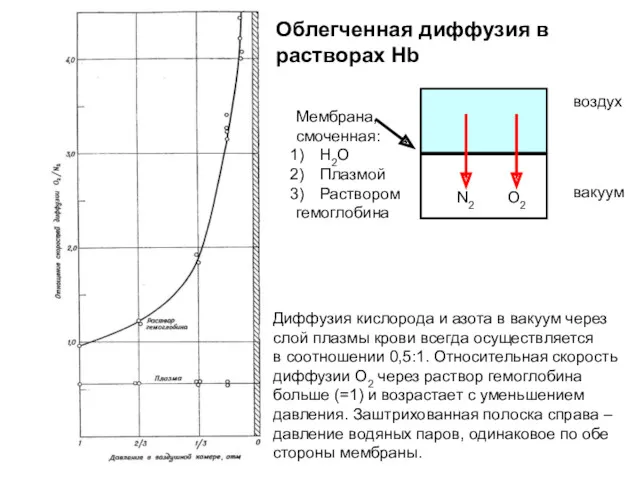
Диффузия кислорода и азота в вакуум через
слой плазмы крови всегда осуществляется
Диффузия кислорода и азота в вакуум через
слой плазмы крови всегда осуществляется

Другие пигменты: Гемоцианин не ускоряет диффузию (очень большие молекулы).
~ Механизм облегчения
Другие пигменты: Гемоцианин не ускоряет диффузию (очень большие молекулы).
~ Механизм облегчения

Увеличение %Hb в тканях беспозвоночных, обитающих в местах с нехваткой О2.
Увеличение %Hb в тканях беспозвоночных, обитающих в местах с нехваткой О2.

Эволюционный аспект распространения Нb среди
представителей разных таксонов
Эволюционный аспект распространения Нb среди
представителей разных таксонов
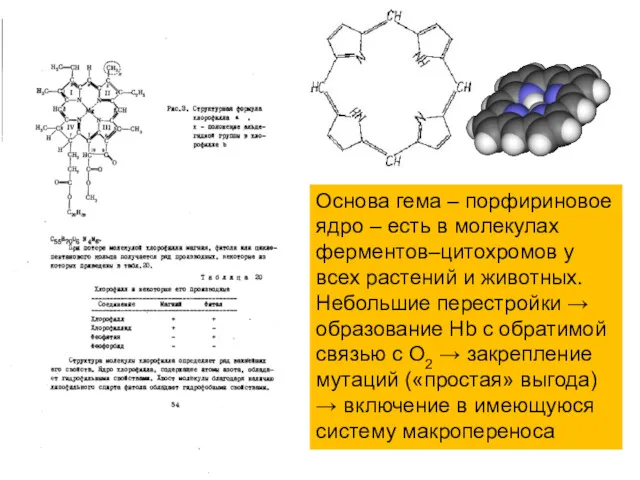
Основа гема – порфириновое ядро – есть в молекулах ферментов–цитохромов у
всех
Основа гема – порфириновое ядро – есть в молекулах ферментов–цитохромов у
всех

КД у беспозвоночных
У многих – высокое сродство к О2.
КД у беспозвоночных
У многих – высокое сродство к О2.
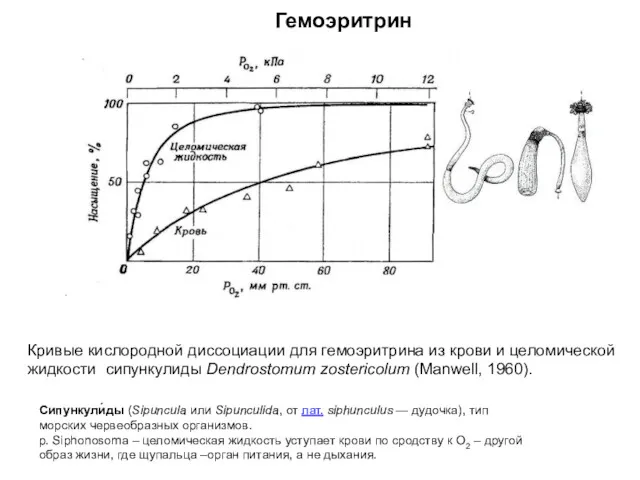
Кривые кислородной диссоциации для гемоэритрина из крови и целомической
жидкости сипункулиды Dendrostomum
Кривые кислородной диссоциации для гемоэритрина из крови и целомической
жидкости сипункулиды Dendrostomum
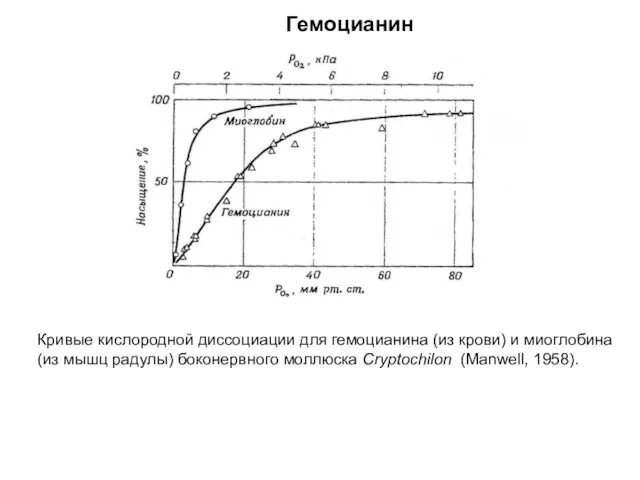
Кривые кислородной диссоциации для гемоцианина (из крови) и миоглобина
(из мышц радулы)
Кривые кислородной диссоциации для гемоцианина (из крови) и миоглобина
(из мышц радулы)

Перенос СО2
Кровь в тканях – отдает О2 и забирает СО2;
Кровь в
Перенос СО2
Кровь в тканях – отдает О2 и забирает СО2;
Кровь в

∑ СО2 в крови >>> CO2, растворенного в жидкости
при данном
∑ СО2 в крови >>> CO2, растворенного в жидкости
при данном
![Уравнение Хендерсона-Хассельбаха: [H+][A-]/[HA] = K → [H+] = K [HA]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/128525/slide-34.jpg)
Уравнение Хендерсона-Хассельбаха:
[H+][A-]/[HA] = K
→ [H+] = K [HA] /[A-] →[-log]
→ pH
Уравнение Хендерсона-Хассельбаха:
[H+][A-]/[HA] = K
→ [H+] = K [HA] /[A-] →[-log]
→ pH

∑ СО2 в крови >>> CO2, растворенного в жидкости
при данном
∑ СО2 в крови >>> CO2, растворенного в жидкости
при данном

Важнейшие буферные вещества крови:
Система угольная кислота ↔ бикарбонат
Фосфаты
Белки (содержат группы, которые
Важнейшие буферные вещества крови:
Система угольная кислота ↔ бикарбонат
Фосфаты
Белки (содержат группы, которые

Kривые Диссоциации (КД) для СО2
Количество СО2, поглощаемого кровью = F (PCO2)
→
Kривые Диссоциации (КД) для СО2
Количество СО2, поглощаемого кровью = F (PCO2)
→
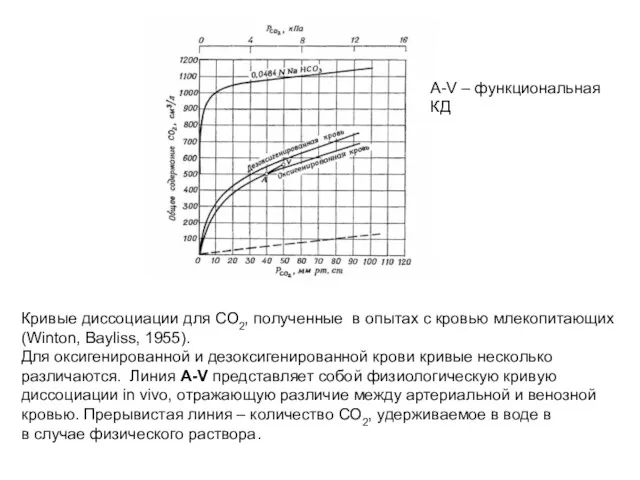
Кривые диссоциации для СО2, полученные в опытах с кровью млекопитающих
(Winton, Bayliss,
Кривые диссоциации для СО2, полученные в опытах с кровью млекопитающих
(Winton, Bayliss,

СО2 при водном дыхании
RQ = СО2 /O2 = lim [0.7-1.0]
Пусть
СО2 при водном дыхании
RQ = СО2 /O2 = lim [0.7-1.0]
Пусть
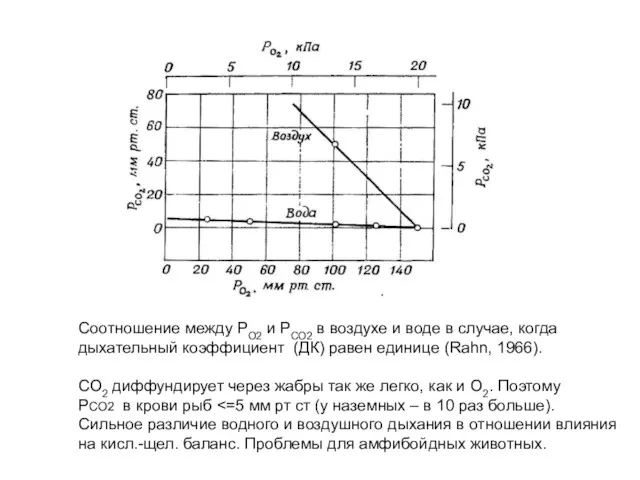
Соотношение между РО2 и РСО2 в воздухе и воде в случае,
Соотношение между РО2 и РСО2 в воздухе и воде в случае,

Концентрация СО2 в крови и рН
Принципиальное различие водных и сухопутных
Концентрация СО2 в крови и рН
Принципиальное различие водных и сухопутных
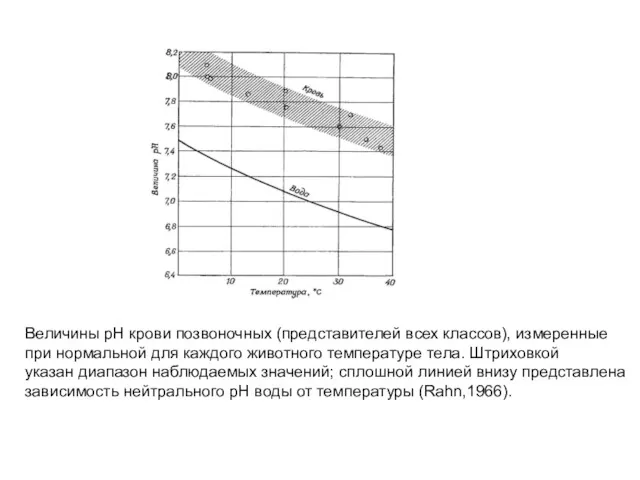
Величины pH крови позвоночных (представителей всех классов), измеренные
при нормальной для каждого
Величины pH крови позвоночных (представителей всех классов), измеренные
при нормальной для каждого

CO2 + H2O ↔ H2CO3 ↔ H+ + HCO3- ↔ [H+

Активность карбоангидразы в крови млекопитающих связана с размерами
тела: она тем выше,
Активность карбоангидразы в крови млекопитающих связана с размерами
тела: она тем выше,

Когда CO2 попадает в кровь, КАРБОАНГИДРАЗА
обеспечивает мгновенное образование угольной кислоты.
обеспечивает мгновенное образование угольной кислоты.

Концентрация О2 в среде и интенсивность
метаболизма
У млекопитающих МR покоя
Концентрация О2 в среде и интенсивность
метаболизма
У млекопитающих МR покоя
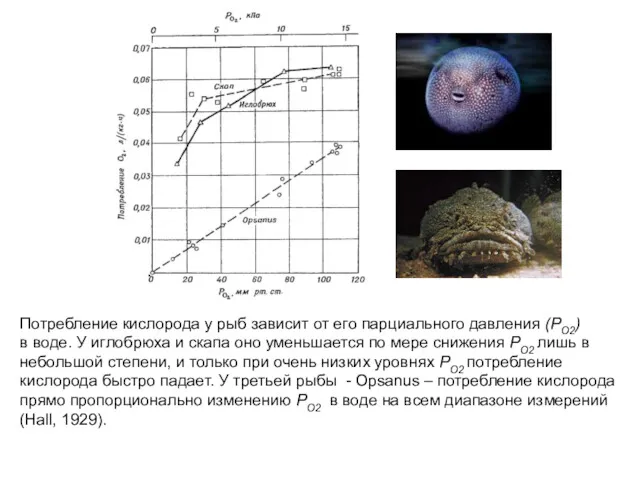
Потребление кислорода у рыб зависит от его парциального давления (РО2)
в воде.
Потребление кислорода у рыб зависит от его парциального давления (РО2)
в воде.

При низких концентрациях кислорода его потребление
золотой рыбкой прямо пропорционально содержанию
При низких концентрациях кислорода его потребление
золотой рыбкой прямо пропорционально содержанию
 Развитие человеческого организма
Развитие человеческого организма Строение и функции белков-2
Строение и функции белков-2 Sequoia sempervirens
Sequoia sempervirens Esophagus. Esophageal Structure
Esophagus. Esophageal Structure Решение задач по общей биологии
Решение задач по общей биологии Папоротеподібні. Загальна характеристика
Папоротеподібні. Загальна характеристика Животные и растения России. Белка и пион
Животные и растения России. Белка и пион General Musculoskeletal Screening: Upper Extremities
General Musculoskeletal Screening: Upper Extremities Презентация Внешнее строение корня
Презентация Внешнее строение корня Класс Млекопитающие. Внешнее строение
Класс Млекопитающие. Внешнее строение Как человек изменял природу
Как человек изменял природу Class Monocotyledones. Division Poales (Graminales)
Class Monocotyledones. Division Poales (Graminales) Методы психогенетики
Методы психогенетики Строение клетки. Клеточная мембрана. Ядро. Цитоплазма. Клеточный центр. Рибосомы
Строение клетки. Клеточная мембрана. Ядро. Цитоплазма. Клеточный центр. Рибосомы Презентация к уроку Здоровье кожи
Презентация к уроку Здоровье кожи Биологические процессы
Биологические процессы Оптические приборы. Дефекты зрения. Очки
Оптические приборы. Дефекты зрения. Очки Тип Членистоногие
Тип Членистоногие Алма шыбыны. Rhagoletis pomonella Walsh
Алма шыбыны. Rhagoletis pomonella Walsh общая биология
общая биология Тип моллюски
Тип моллюски Копротрофы. Паразиты высших растений
Копротрофы. Паразиты высших растений Тип Моллюски или Мягкотелые
Тип Моллюски или Мягкотелые Повторение по биологии 5 класс
Повторение по биологии 5 класс Организм. Системы органов
Организм. Системы органов Тип Кольчатые черви. 7 класс
Тип Кольчатые черви. 7 класс Зоология позвоночных. Анатомия млекопитающих. (Лекция 12)
Зоология позвоночных. Анатомия млекопитающих. (Лекция 12) Plants and animals of North America
Plants and animals of North America