- Питание бактерий

Содержание
- 2. Под питанием понимают процессы поступления и выведения питательных веществ в клетку и из клетки. Питание в
- 3. Для бактерий характерно многообразие источников получения питательных веществ. В зависимости от источника получения углерода бактерии делят
- 4. Процессы питания должны обеспечивать энергетические потребности бактериальной клетки. По источникам энергии микроорганизмы делят на: 1) фототрофы
- 5. По степени гетеротрофности микроорганизмы делятся на: 1) сапрофиты – питаются мертвым органическим материалом; 2) паразиты –
- 6. Факторами роста бактерий являются витамины, аминокислоты, пуриновые и пиримидиновые основания, присутствие которых ускоряет рост. Среди бактерий
- 7. Микроорганизмы ассимилируют питательные вещества в виде небольших молекул, поэтому белки, полисахариды и другие биополимеры могут служить
- 8. Транспорт веществ
- 9. Для того, чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки, они должны проникнуть в клетку
- 10. Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.
- 11. При пассивном переносе вещество проникает в клетку только по градиенту концентрации. Затрат энергии при этом не
- 12. Простая диффузия — неспецифическое проникновение веществ в клетку, при этом решающее значение имеет величина молекул и
- 13. Облегченная диффузия протекает с участием белка-переносчика. Скорость этого способа переноса зависит от концентрации вещества в наружном
- 14. При активном переносе вещество проникает в клетку против градиента концентрации при помощи белка-переносчика — пермеазы. При
- 15. При одном типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) «накачиваются» в клетку и создают концентрацию,
- 16. Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система, составной частью которой является белок-переносчик,
- 17. Поступив в клетку, органический источник углерода и энергии вступает в цепь биохимических реакций, в результате которых
- 18. Конструктивный метаболизм
- 19. Основными видами пластического обмена являются: 1) белковый; 2) углеводный; 3) липидный; 4) нуклеиновый.
- 20. Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают белки под действием протеаз с
- 21. Биосинтез аминокислот. Распад белков в аэробных условиях называется тлением, в анаэробных – гниением. В результате распада
- 22. Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные продукты фруктозодифосфатного (ФДФ) и
- 23. Биосинтез нуклеотидов. Пуриновые и пиримидиновые нуклеотиды — это те строительные блоки, из которых синтезируются нуклеиновые кислоты.
- 24. Биосинтез жиров. Жиры или липиды являются важными компонентами ЦПМ и клеточной стенки грамотрицательных бактерий, а также
- 25. Биосинтез углеводов. Углеводы представлены в бактериальной клетке в виде моно-, ди- и полисахаридов, а также комплексных
- 27. Скачать презентацию

Под питанием понимают процессы поступления и выведения питательных веществ в клетку
Под питанием понимают процессы поступления и выведения питательных веществ в клетку

Для бактерий характерно многообразие источников получения питательных веществ.
В зависимости от источника
Для бактерий характерно многообразие источников получения питательных веществ.
В зависимости от источника

Процессы питания должны обеспечивать энергетические потребности бактериальной клетки. По источникам энергии
Процессы питания должны обеспечивать энергетические потребности бактериальной клетки. По источникам энергии

По степени гетеротрофности микроорганизмы делятся на:
1) сапрофиты – питаются мертвым
По степени гетеротрофности микроорганизмы делятся на:
1) сапрофиты – питаются мертвым

Факторами роста бактерий являются витамины, аминокислоты, пуриновые и пиримидиновые основания, присутствие
Факторами роста бактерий являются витамины, аминокислоты, пуриновые и пиримидиновые основания, присутствие

Микроорганизмы ассимилируют питательные вещества в виде небольших молекул, поэтому белки, полисахариды
Микроорганизмы ассимилируют питательные вещества в виде небольших молекул, поэтому белки, полисахариды

Транспорт веществ
Транспорт веществ

Для того, чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки,
Для того, чтобы питательные вещества могли подвергнуться превращениям в цитоплазме клетки,

Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.
Существует два типа переноса веществ в бактериальную клетку: пассивный и активный.

При пассивном переносе вещество проникает в клетку только по градиенту концентрации.
При пассивном переносе вещество проникает в клетку только по градиенту концентрации.

Простая диффузия — неспецифическое проникновение веществ в клетку, при этом решающее
Простая диффузия — неспецифическое проникновение веществ в клетку, при этом решающее

Облегченная диффузия протекает с участием белка-переносчика. Скорость этого способа переноса зависит
Облегченная диффузия протекает с участием белка-переносчика. Скорость этого способа переноса зависит

При активном переносе вещество проникает в клетку против градиента концентрации при
При активном переносе вещество проникает в клетку против градиента концентрации при

При одном типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) «накачиваются»
При одном типе активного транспорта небольшие молекулы (аминокислоты, некоторые сахара) «накачиваются»

Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система,
Для осуществления этих процессов в бактериальной клетке локализуется специальная фосфотрансферная система,

Поступив в клетку, органический источник углерода и энергии вступает в цепь
Поступив в клетку, органический источник углерода и энергии вступает в цепь

Конструктивный метаболизм
Конструктивный метаболизм

Основными видами пластического обмена являются:
1) белковый;
2) углеводный;
3) липидный;
Основными видами пластического обмена являются:
1) белковый;
2) углеводный;
3) липидный;

Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают
Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают

Биосинтез аминокислот.
Распад белков в аэробных условиях называется тлением, в анаэробных
Биосинтез аминокислот.
Распад белков в аэробных условиях называется тлением, в анаэробных

Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат
Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат

Биосинтез нуклеотидов.
Пуриновые и пиримидиновые нуклеотиды — это те строительные блоки,
Биосинтез нуклеотидов.
Пуриновые и пиримидиновые нуклеотиды — это те строительные блоки,

Биосинтез жиров.
Жиры или липиды являются важными компонентами ЦПМ и клеточной
Биосинтез жиров.
Жиры или липиды являются важными компонентами ЦПМ и клеточной

Биосинтез углеводов.
Углеводы представлены в бактериальной клетке в виде моно-, ди-
Биосинтез углеводов.
Углеводы представлены в бактериальной клетке в виде моно-, ди-
 Птицы все - мои друзья
Птицы все - мои друзья Класс птицы как высокоорганизованная и специализированная группа. Морфофизиологические особенности, биология, распространение
Класс птицы как высокоорганизованная и специализированная группа. Морфофизиологические особенности, биология, распространение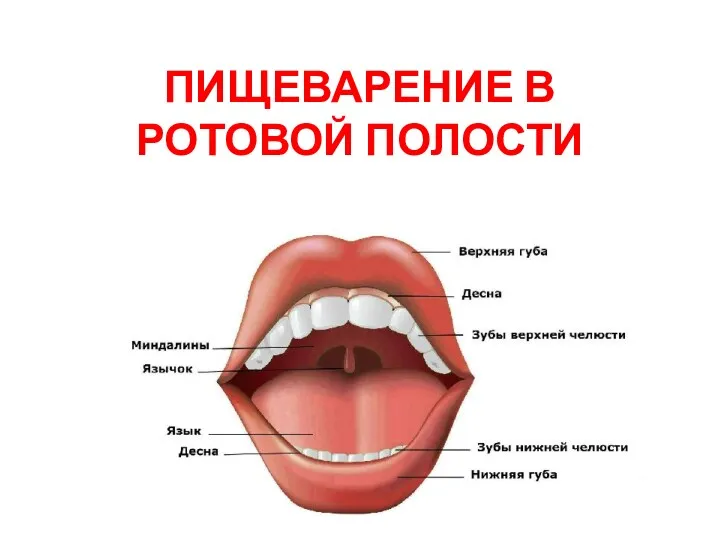 Пищеварение в ротовой полости
Пищеварение в ротовой полости Проверка знаний по теме Передвижение веществ в растении
Проверка знаний по теме Передвижение веществ в растении Общая вирусология. Систематика, морфология и физиология вирусов. Бактериофаги. Использование фагов в медицинской практике
Общая вирусология. Систематика, морфология и физиология вирусов. Бактериофаги. Использование фагов в медицинской практике Далалық және ауылшаруашылық дақылдары
Далалық және ауылшаруашылық дақылдары 6 класс. Побег.
6 класс. Побег. Многообразие организмов
Многообразие организмов Китоподібні - виключно водні ссавці
Китоподібні - виключно водні ссавці Биологическая роль и строение нуклеиновых кислот
Биологическая роль и строение нуклеиновых кислот Esophagus. Esophageal Structure
Esophagus. Esophageal Structure Класс насекомые
Класс насекомые Зелёное черенкование с закрытой корневой системой
Зелёное черенкование с закрытой корневой системой Биохимия соединительной ткани
Биохимия соединительной ткани Ас қорыту жүйесі
Ас қорыту жүйесі Сенсорные системы. Общая характеристика сенсорных рецепторов. Зрительная система
Сенсорные системы. Общая характеристика сенсорных рецепторов. Зрительная система מבנה התא
מבנה התא Деление клетки. Митоз
Деление клетки. Митоз Обобщающий урок по теме: Класс млекопитающие, 7 класс
Обобщающий урок по теме: Класс млекопитающие, 7 класс Биохимия. Ферменты (Лекция№2)
Биохимия. Ферменты (Лекция№2) Состав крови
Состав крови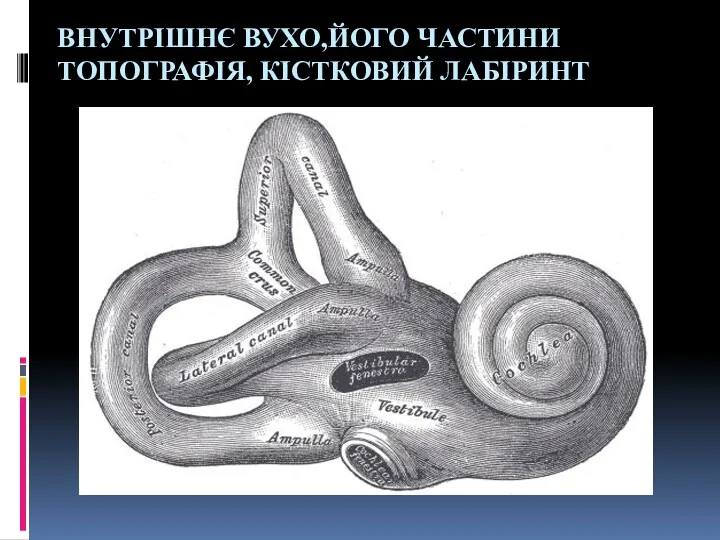 Внутрішнє вухо, його частини, топографія, кістковий лабіринт
Внутрішнє вухо, його частини, топографія, кістковий лабіринт Экологические группы птиц презентация
Экологические группы птиц презентация Обучающая презентация Флора г. Аксая Ростовской области
Обучающая презентация Флора г. Аксая Ростовской области Какая наука изучает историю живых организмов по сохранившимся останкам?
Какая наука изучает историю живых организмов по сохранившимся останкам? Zarys Fizjologii - Układu Krążenia cz. I
Zarys Fizjologii - Układu Krążenia cz. I Пищеварение и усвояемость пищи
Пищеварение и усвояемость пищи Водоросли. Лекция 2
Водоросли. Лекция 2