- Принципы зоологической систематики

Содержание
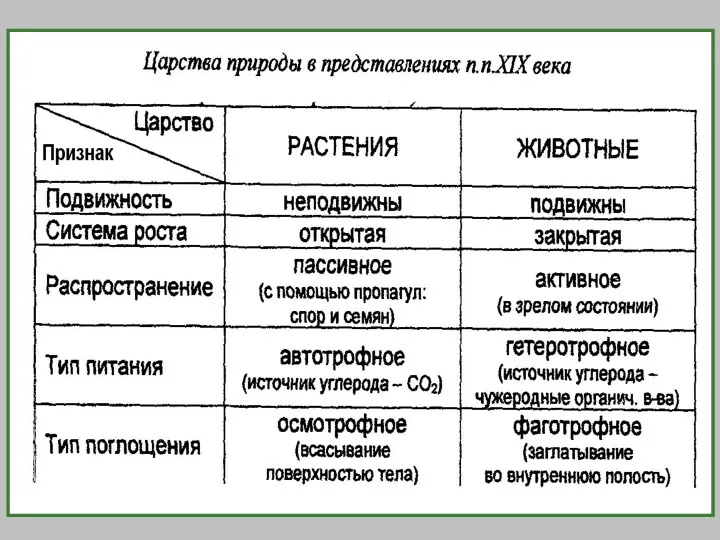
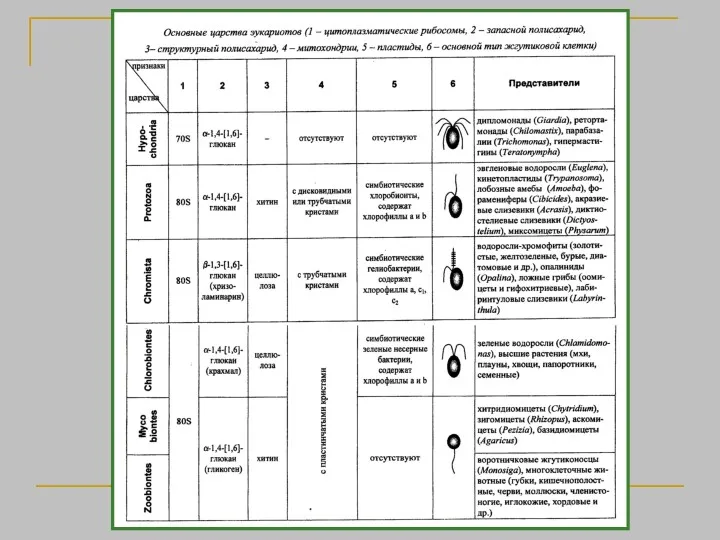
- 2. Биологическая систематика — наука, в задачи которой входит разработка принципов классификации живых организмов и применение этих
- 3. Классификация живых организмов строится по иерархическому принципу. Различные уровни иерархии (ранги) имеют собственные названия. Основу системы
- 4. α-систематика – описание новых видов, их именование и предварительное распределение по родам, β-систематика – выяснение взаимоотношения
- 5. Принципы систематики Концепция вида
- 6. Первым систематиком был Адам. Но он не успел.
- 11. Аристотель, (384 — 322 до н.э. ) Первая естественная система. История животных, О частях животных, О
- 12. По Геккелю филогенетика – наука о путях, закономерностях и причинах исторического развития организмов. Родословное древо Геккеля
- 13. “Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими древесами, на которые я указал
- 14. Традиционная кладистика (Hennig, 1950, 1966) Хенниг предложил строго научные принципы перехода от анализа признаков к реконструкции
- 15. Признаки Негомологичные (гомоплазии) Гомологичные Плезиоморфии Апоморфии Синапоморфии
- 16. Гомоплазии – независимо возникшие признаки. Они не несут никакой информации о филогении 1 - гомоплазия
- 17. Плезиоморфии – древние (исходные; примитивные) гомологичные признаки. Они не несут никакой информации о топологии поздних ветвлений.
- 18. Апоморфия – новый (продвинутый; производный; прогрессивынй) гомологичный признак. Апоморфия является специфическим маркером эволюционной линии. Единичная апоморфия,
- 19. Но если апоморфия возникла до разделения ветвей и передалась в обе ветки, то наличие такой апоморфии
- 20. Для построения филогении трех таксонов (два ветвления) необходимо наличие одной синапоморфии плезиоморфия синапоморфия
- 21. В общем виде для полного разрешения филогении, включающей n ветвлений, необходимо и достаточно n-1 синапоморфий (по
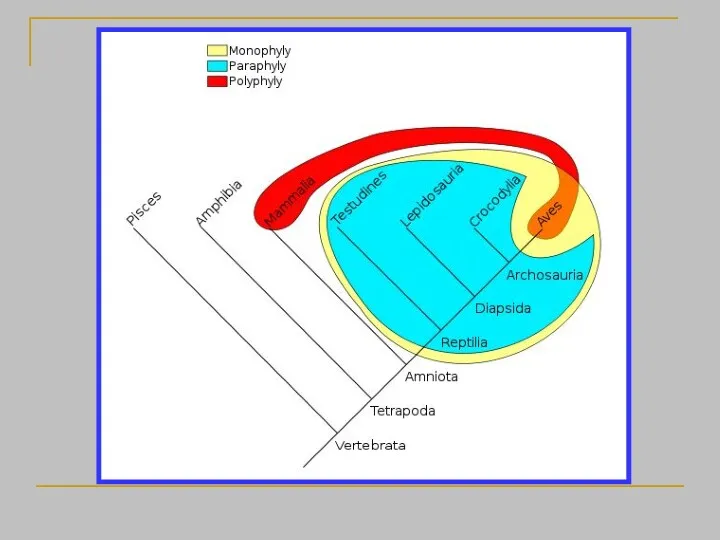
- 22. Филогения строится как система соподчиненных (вложенных одна в другую) клад (монофилетических групп), каждая из которых выявляется
- 23. Модель эволюции в кладистике по Хеннигу Топология - строгая дихотомия Процесс – накопление синапоморфий. Одна истинная
- 24. Картины филогенезов, которуе создает кладистический (по Хеннигу и парсимониальный) анализ, «неполны и однобоки»: Анагенез не учитывается
- 25. Принцип монофилии лежит в самой основе алгоритма построения дерева в хенниговской кладистике. Сипапоморфии однозначно определяют только
- 26. Кладизм объявляет парафилетические группы вне закона просто по той причине, что он не умеет их выявлять
- 27. Проблемы парафилетических таксонов 1+2 = парафилетический таксон. Признак A не уникален, признак B характеризует лишь часть
- 28. Монофилетический таксон - группа, которая включает предка и всех его потомков Монофилетические группы могут иметь синапоморфии
- 30. Три таксономии
- 31. Фенетика основанная на количественной оценке так называемого общего сходства (overall similarity) Отказ от доминирования принципа гомологии
- 32. Эволюционная таксономия Подобно кладистике, при построении системы опирается на эволюционную близость (т. е. общность происхождения), однако
- 33. Традиционная и нумерическая кладистика Увеличение числа признаков приводит к противоречиям между предполагаемыми синапоморфиями, которые свидетельствуют о
- 34. Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения: Традиционная кладистика переисследование материала,
- 35. Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его решения: Нумерическая кладистика использование большого
- 36. Нумерическая кладистика и метод максимальной парсимонии Как выбрать “правильное” дерево? - критерий максимальной парсимонии
- 37. Нет гомоплазий – одно возможное дерево Число шагов (L) = 3 Сайт 4 – инвариантный, сайт
- 38. Первое дерево более парсимониальное, оно короче Происходит голосование “синапоморфиями”
- 40. Скачать презентацию

Биологическая систематика — наука, в задачи которой входит разработка принципов классификации живых организмов и
Биологическая систематика — наука, в задачи которой входит разработка принципов классификации живых организмов и

Классификация живых организмов строится по иерархическому принципу.
Различные уровни иерархии (ранги)
Классификация живых организмов строится по иерархическому принципу.
Различные уровни иерархии (ранги)

α-систематика – описание новых видов, их именование и предварительное распределение по
α-систематика – описание новых видов, их именование и предварительное распределение по

Принципы систематики
Концепция вида
Принципы систематики
Концепция вида
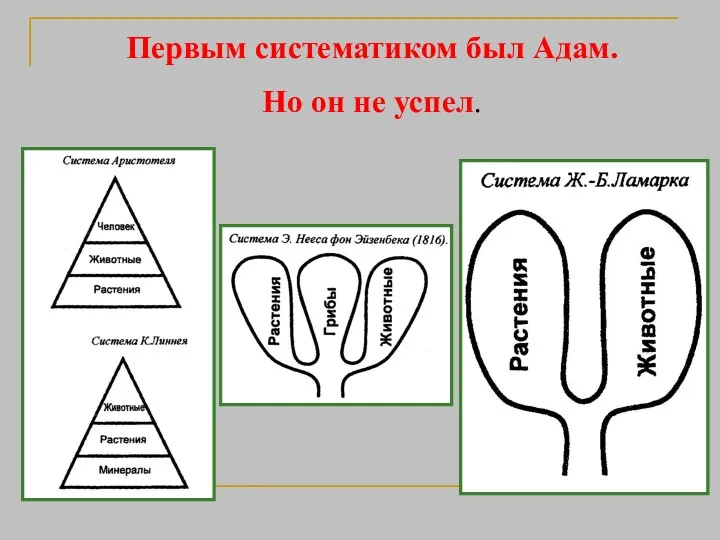
Первым систематиком был Адам.
Но он не успел.
Первым систематиком был Адам.
Но он не успел.
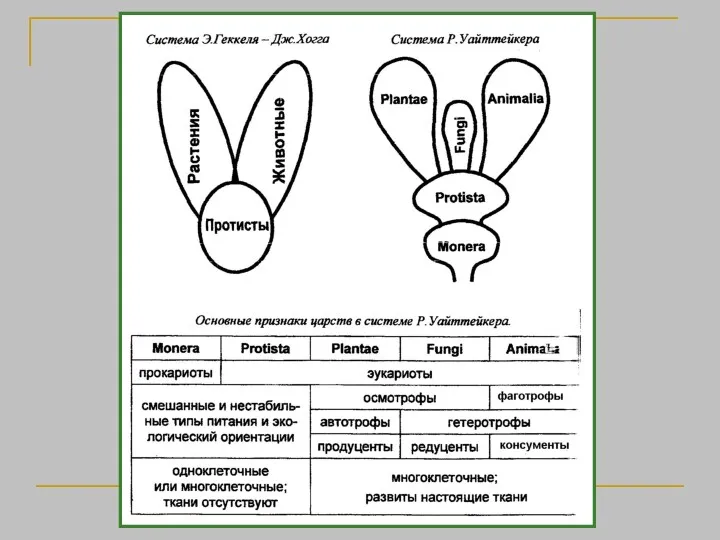
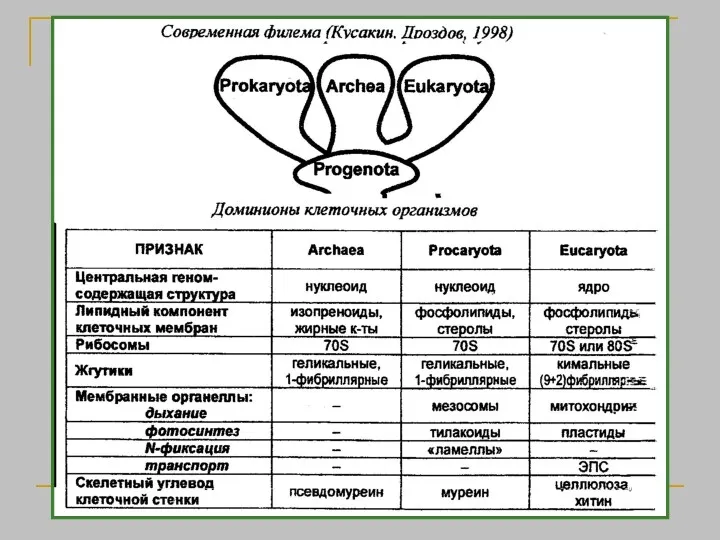

Аристотель, (384 — 322 до н.э. ) Первая естественная система. История животных, О
Аристотель, (384 — 322 до н.э. ) Первая естественная система. История животных, О

По Геккелю филогенетика – наука о путях, закономерностях и причинах исторического
По Геккелю филогенетика – наука о путях, закономерностях и причинах исторического
![“Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/240834/slide-12.jpg)
“Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими
“Недавно в лабораторию [Моргана]пришла почта с произведениями Северцова с многочисленными филогенетическими

Традиционная кладистика (Hennig, 1950, 1966)
Хенниг предложил строго научные принципы перехода от
Традиционная кладистика (Hennig, 1950, 1966) Хенниг предложил строго научные принципы перехода от

Признаки
Негомологичные (гомоплазии)
Гомологичные
Плезиоморфии
Апоморфии
Синапоморфии
Признаки
Негомологичные (гомоплазии)
Гомологичные
Плезиоморфии
Апоморфии
Синапоморфии
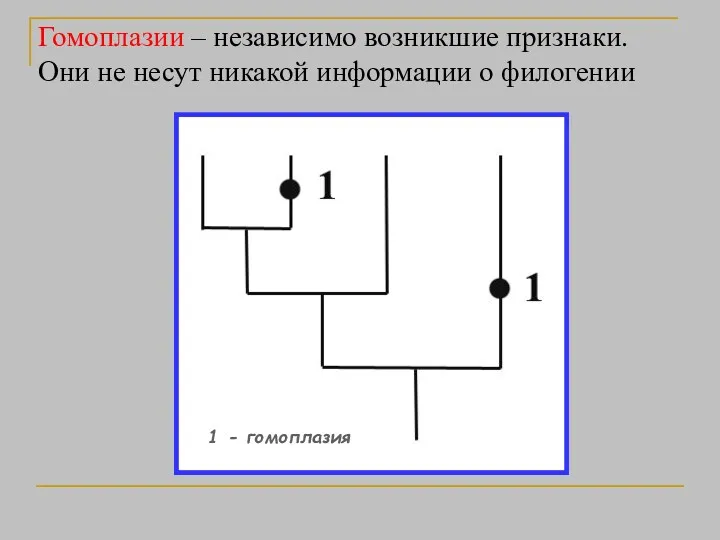
Гомоплазии – независимо возникшие признаки. Они не несут никакой информации о
Гомоплазии – независимо возникшие признаки. Они не несут никакой информации о
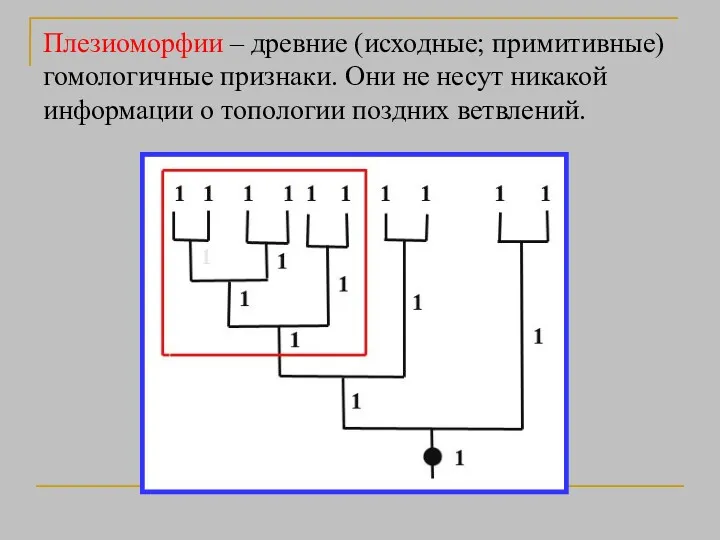
Плезиоморфии – древние (исходные; примитивные) гомологичные признаки. Они не несут никакой
Плезиоморфии – древние (исходные; примитивные) гомологичные признаки. Они не несут никакой
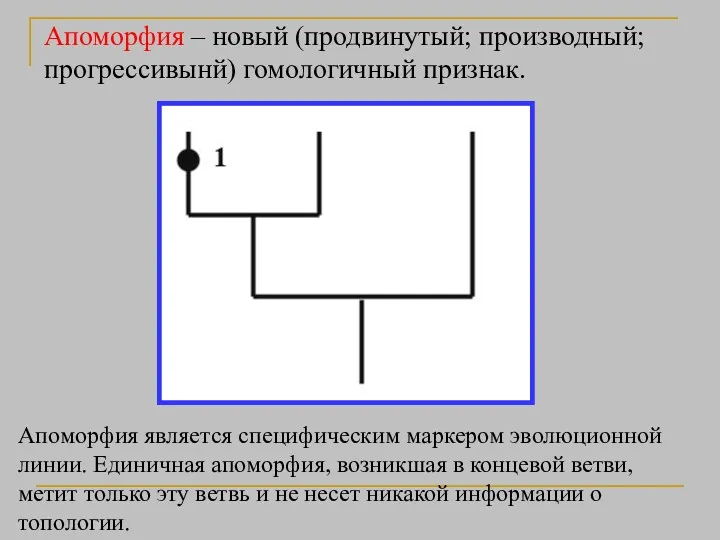
Апоморфия – новый (продвинутый; производный; прогрессивынй) гомологичный признак.
Апоморфия является специфическим
Апоморфия – новый (продвинутый; производный; прогрессивынй) гомологичный признак.
Апоморфия является специфическим
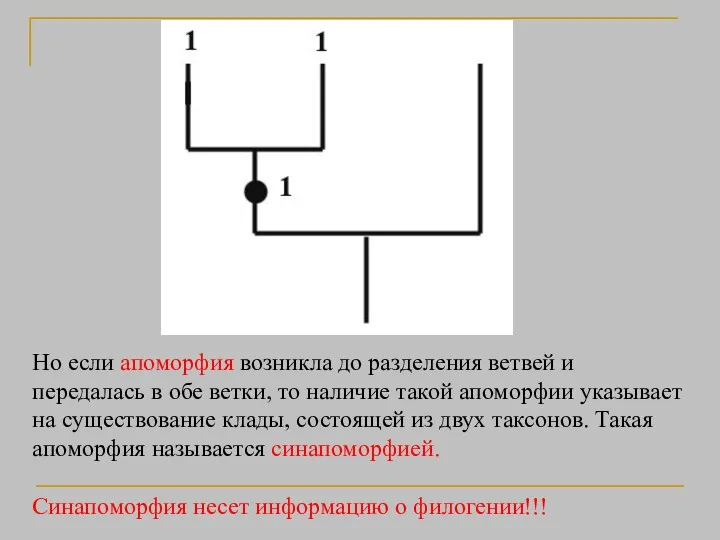
Но если апоморфия возникла до разделения ветвей и передалась в обе
Но если апоморфия возникла до разделения ветвей и передалась в обе
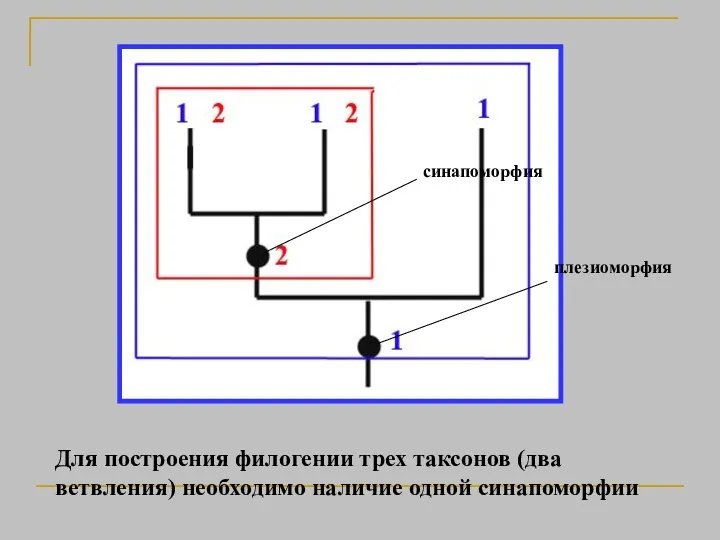
Для построения филогении трех таксонов (два ветвления) необходимо наличие одной синапоморфии
плезиоморфия
синапоморфия
Для построения филогении трех таксонов (два ветвления) необходимо наличие одной синапоморфии
плезиоморфия
синапоморфия
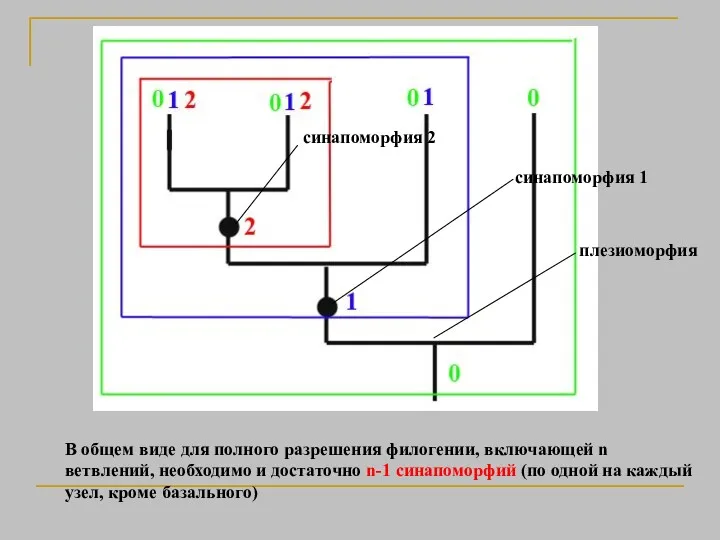
В общем виде для полного разрешения филогении, включающей n ветвлений, необходимо
В общем виде для полного разрешения филогении, включающей n ветвлений, необходимо

Филогения строится как система соподчиненных (вложенных одна в другую) клад (монофилетических
Филогения строится как система соподчиненных (вложенных одна в другую) клад (монофилетических

Модель эволюции в кладистике по Хеннигу
Топология - строгая дихотомия
Процесс – накопление
Модель эволюции в кладистике по Хеннигу
Топология - строгая дихотомия
Процесс – накопление
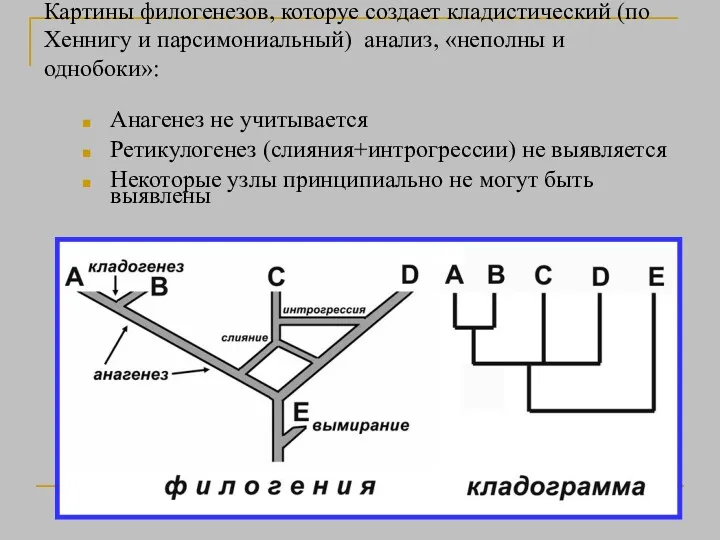
Картины филогенезов, которуе создает кладистический (по Хеннигу и парсимониальный) анализ, «неполны
Картины филогенезов, которуе создает кладистический (по Хеннигу и парсимониальный) анализ, «неполны
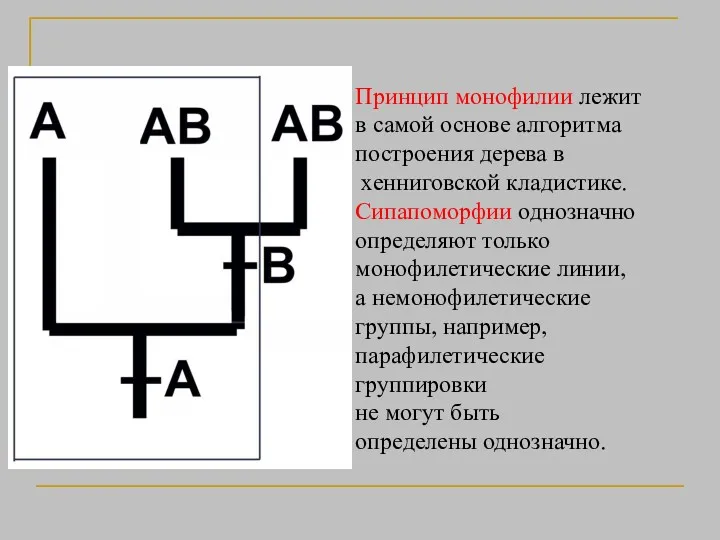
Принцип монофилии лежит
в самой основе алгоритма
построения дерева в
хенниговской
Принцип монофилии лежит
в самой основе алгоритма
построения дерева в
хенниговской
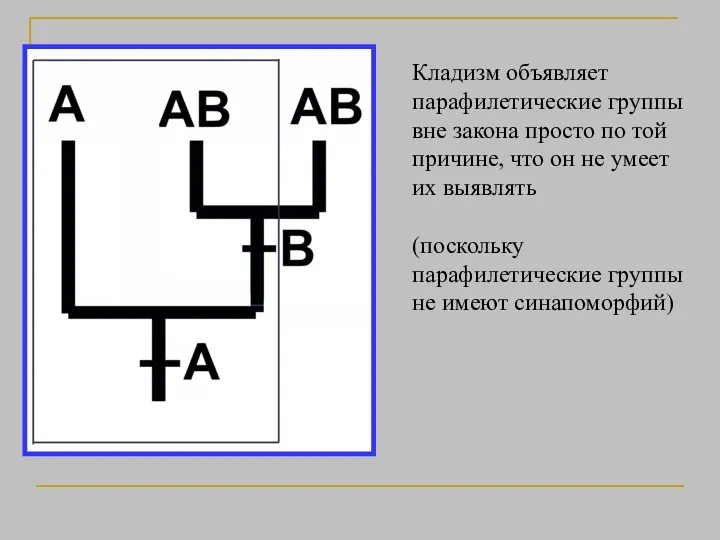
Кладизм объявляет парафилетические группы вне закона просто по той причине, что
Кладизм объявляет парафилетические группы вне закона просто по той причине, что
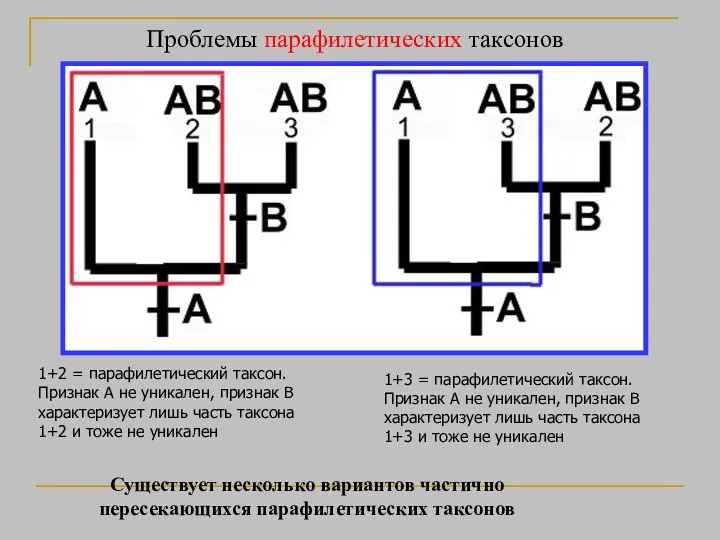
Проблемы парафилетических таксонов
1+2 = парафилетический таксон. Признак A не уникален, признак
Проблемы парафилетических таксонов
1+2 = парафилетический таксон. Признак A не уникален, признак
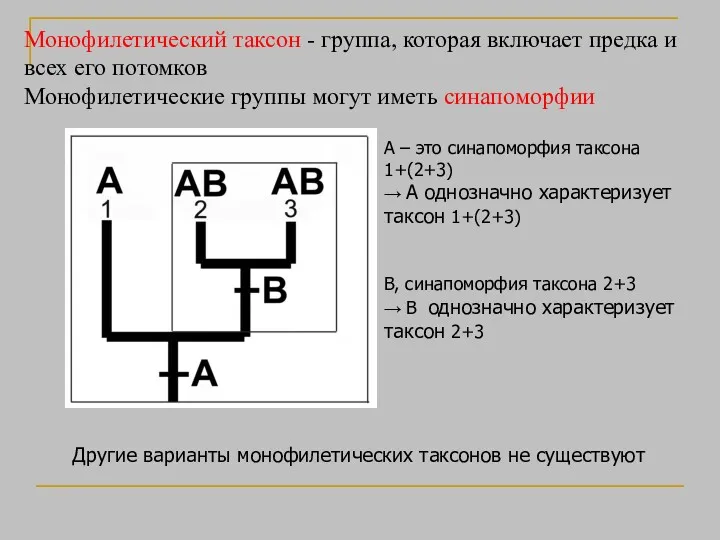
Монофилетический таксон - группа, которая включает предка и всех его потомков
Монофилетический таксон - группа, которая включает предка и всех его потомков
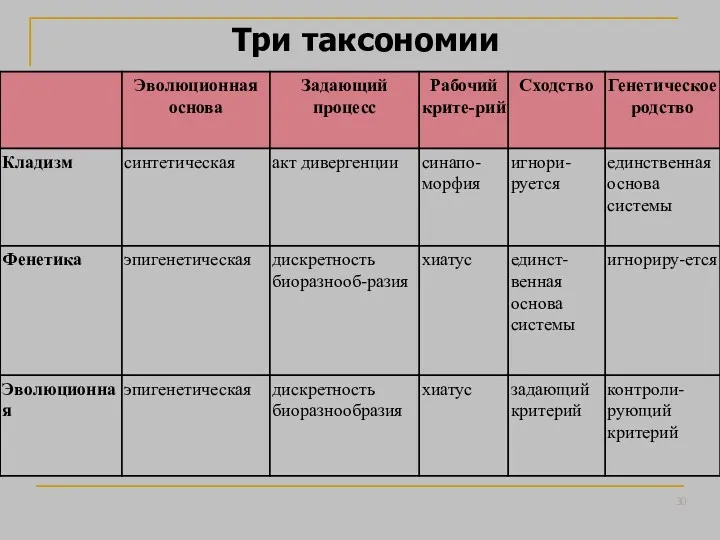
Три таксономии
Три таксономии

Фенетика
основанная на количественной оценке так называемого общего сходства (overall similarity)
Отказ от
Фенетика
основанная на количественной оценке так называемого общего сходства (overall similarity)
Отказ от

Эволюционная таксономия
Подобно кладистике, при построении системы опирается на эволюционную близость (т. е. общность происхождения),
Эволюционная таксономия
Подобно кладистике, при построении системы опирается на эволюционную близость (т. е. общность происхождения),

Традиционная и нумерическая кладистика
Увеличение числа признаков приводит к противоречиям между предполагаемыми
Традиционная и нумерическая кладистика
Увеличение числа признаков приводит к противоречиям между предполагаемыми

Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его
Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его

Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его
Если возникает конфликт между потенциальными синапоморфиями, то есть два пути его

Нумерическая кладистика и
метод максимальной парсимонии
Как выбрать “правильное” дерево?
- критерий максимальной
Нумерическая кладистика и
метод максимальной парсимонии
Как выбрать “правильное” дерево?
- критерий максимальной
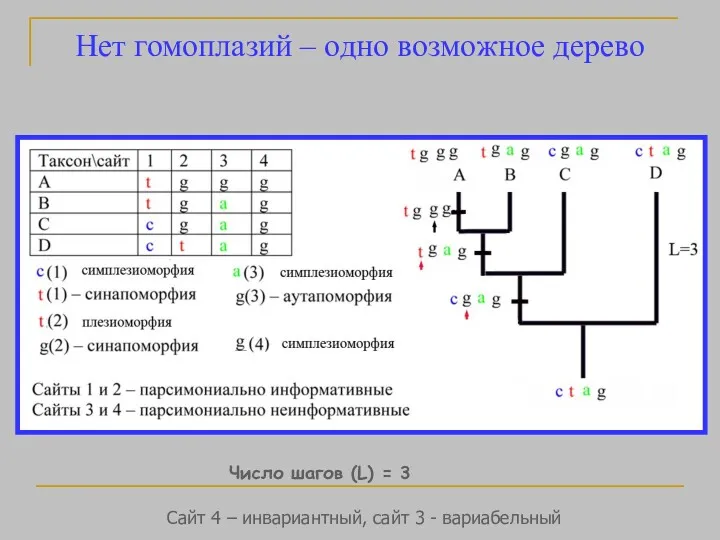
Нет гомоплазий – одно возможное дерево
Число шагов (L) = 3
Сайт 4
Нет гомоплазий – одно возможное дерево
Число шагов (L) = 3
Сайт 4
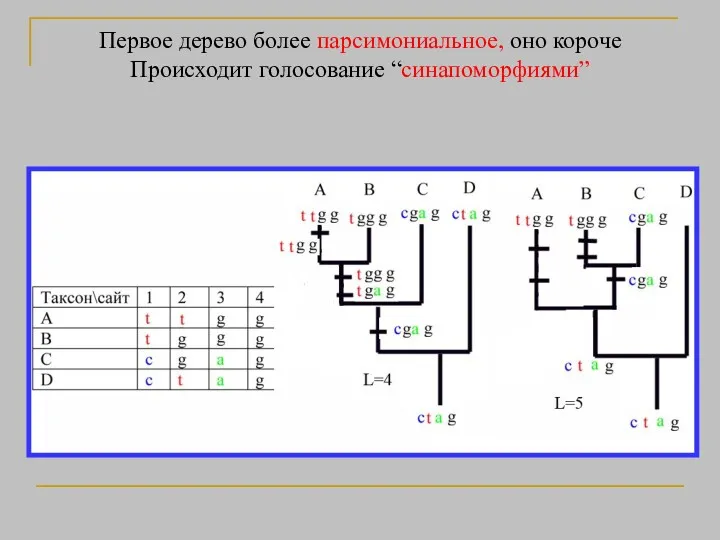
Первое дерево более парсимониальное, оно короче
Происходит голосование “синапоморфиями”
Первое дерево более парсимониальное, оно короче
Происходит голосование “синапоморфиями”
 Костная и мышечная системы. Скелет головы
Костная и мышечная системы. Скелет головы Жасуша ядросы
Жасуша ядросы Онтогенез и филогенез нервной системы
Онтогенез и филогенез нервной системы Вкусовые ощущения
Вкусовые ощущения Биологические ритмы и их влияние на работоспособность человека
Биологические ритмы и их влияние на работоспособность человека Радіобіологія організму людини і тварин. Радіочутливість тканин і органів організму людини і тварин
Радіобіологія організму людини і тварин. Радіочутливість тканин і органів організму людини і тварин Байгабилов Д., 2 курс
Байгабилов Д., 2 курс Семейство пингвины
Семейство пингвины Розмноження на клітинному рівні. (Лекція 3)
Розмноження на клітинному рівні. (Лекція 3) Обмен веществ и превращение энергии в клетке. Воспроизведение клеток.Тема №2
Обмен веществ и превращение энергии в клетке. Воспроизведение клеток.Тема №2 Функции тонкого и толстого кишечника. Роль печени
Функции тонкого и толстого кишечника. Роль печени Стебель, его значение и строение. Многообразие стеблей
Стебель, его значение и строение. Многообразие стеблей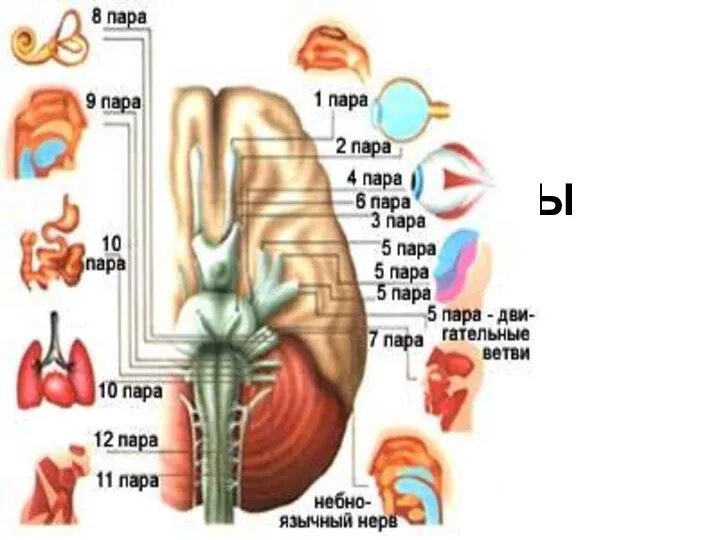 Черепные нервы
Черепные нервы Живі організми як середовище існування
Живі організми як середовище існування Конструкция костей черепа
Конструкция костей черепа презентация по биологии 7 класс
презентация по биологии 7 класс Генная инженерия растений
Генная инженерия растений Цитоплазматическая наследственность
Цитоплазматическая наследственность Структура центра терморегуляции. Роль центра в регуляции теплообмена
Структура центра терморегуляции. Роль центра в регуляции теплообмена Животные (для дошкольников)
Животные (для дошкольников) Мужская половая система
Мужская половая система Генетика – наука, о закономерностях наследственности и изменчивости
Генетика – наука, о закономерностях наследственности и изменчивости Физиология микроорганизмов
Физиология микроорганизмов Протопласты растительных клеток как объекты биологического конструирования
Протопласты растительных клеток как объекты биологического конструирования Теория И.П. Павлова о типах высшей нервной деятельности
Теория И.П. Павлова о типах высшей нервной деятельности Редкие животные
Редкие животные Анатомиялық препараттардан мойын, көкірек, бел, сегізкөз және құйымшақ омыртқаларының ерекшеліктерін оқу
Анатомиялық препараттардан мойын, көкірек, бел, сегізкөз және құйымшақ омыртқаларының ерекшеліктерін оқу Флора бассейна реки Водлы
Флора бассейна реки Водлы