- Развитие до имплантации. Мышь от оплодотворения до имплантации

Содержание
- 2. Мышь от оплодотворения до имплантации: 1 – 5 сутки развития
- 3. Эмбрионы мыши и человека
- 4. Важнейшие событие преимплантационного развития - переход управления развитием эмбриона от материнского генома (генома ооцита) к геному
- 5. У большинства видов животных процесс MZT медленный: в течение многих клеточных циклов (до 2-3 десятков циклов)
- 6. Lee et.al.Annu Rev Cell Dev Biol. Author manuscript; available in PMC 2015 August 11. Возрастание уровня
- 7. Изменение экспрессии в раннем развитии эмбрионов мыши
- 8. Активация генома эмбриона - участие материнских генов Филогенетическая дистанция между белками ZAR1 у разных видов позвоночных:
- 9. Профиль экспрессии Zscan4 во время доимплантационного развития: А: whole mount in situ hybridization; В: qRT-PCR analysis
- 10. Expressions of Zscan4 and Pou5f1 in the blastocysts, blastocyst outgrowth, and ES cells by the whole
- 11. Определение 1: Тотипотентность – способность клетки развиваться в целый организм Определение 2: Тотипотентность - способность клетки
- 12. После оплодотворения некоторые эпигенетические факторы материнского происхождения (накопленные во время оогенеза), (в том числе DNMTs, Ago2)
- 13. Выбор судьбы клетками раннего эмбриона: решение в 2 этапа 1 этап: внутри или снаружи (вопрос: за
- 15. Дифференцировка бластоцисты: ОСТ-4
- 16. 32 бластомера и более Компактизация и формирование морулы
- 17. Период компактизации Переход от 8 к 16 Переход от 16 к 32 Клеток мало для формирования
- 18. Компактизация и поляризация клеток эмбриона: как круглые клетки становятся плоскими? Fierro-González et al., Cadherin-dependent filopodia control
- 19. Протеин-киназа Hippo (у млекопитающих называется Mst1/2); Этот каскад контролирует пролиферацию и апоптоз; Мутации в этом гене
- 20. Hippo сигналинг: преобразование позиционной информации в молекулярный каскад Gata3 – ранний маркер клеток ТЭ Oct4, Sox2
- 21. Становление полярности эмбриона на стадиях дробления: апикально-базальные различия в строении кортикального слоя. Ajduk and Zernicka-Goetz Molecular
- 22. Hippo сигналинг недостаточен, необходима «внутренняя» среда Щелевые контакты Адгезионные контакты → сигналиг через цитоплазматические тирозин-киназы Фокальные
- 23. Дробление клеток раннего эмбриона: насколько клетки идентичны? Эксперименты по исследованию асимметрии ооцита и раннего эмбриона овцы
- 24. Cleavage patterns of mouse embryos at 2- to 4-cell transition. Two-cell stage mouse blastomeres divide either
- 25. Асимметричная локализация в клетках: aPKC, Par3 и ezrin ezrin aPKC aPKC Par3 Louvet et al., 1996
- 26. Межклеточные контакты в дробящихся эмбрионах, моруле и бластоцисте Gardner, 2002
- 27. Поддержание плюрипотентности ES клеток в бластоцисте – сигнальные пути REGULATORY NETWORKS IN EMBRYO-DERIVED PLURIPOTENT STEM CELLS
- 28. Na/K-ATPаза, AQP (аквапорины) и белки плотных контактов (TJ) играют основную роль в кавитации и формировании бластоцисты:
- 29. Классические представления: В ранней бластоцисте все клетки ВКМ с самого начала различаются по положению, в результате
- 30. FGFR1 Gata6, FGFR1-4 Преимплантационный эпибласт Примитивная эндодерма 2008: PrE - предшественники первичной энтодермы (гипобласта) мигрируют к
- 31. Bischoff et al., 2008 События в жизни преимплантационного эмбриона: что влияет на время их наступления? Эмбрион
- 32. Тотипотентность – способность развиваться как в клетки зародышевых, так и внезародышевых структур:(клетки эмбриона до компактизации ).
- 34. Скачать презентацию
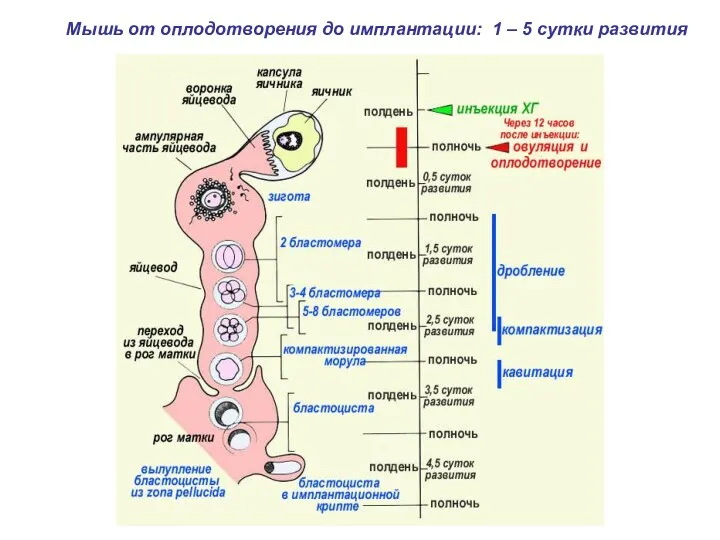
Мышь от оплодотворения до имплантации: 1 – 5 сутки развития
Мышь от оплодотворения до имплантации: 1 – 5 сутки развития

Эмбрионы мыши и человека
Эмбрионы мыши и человека
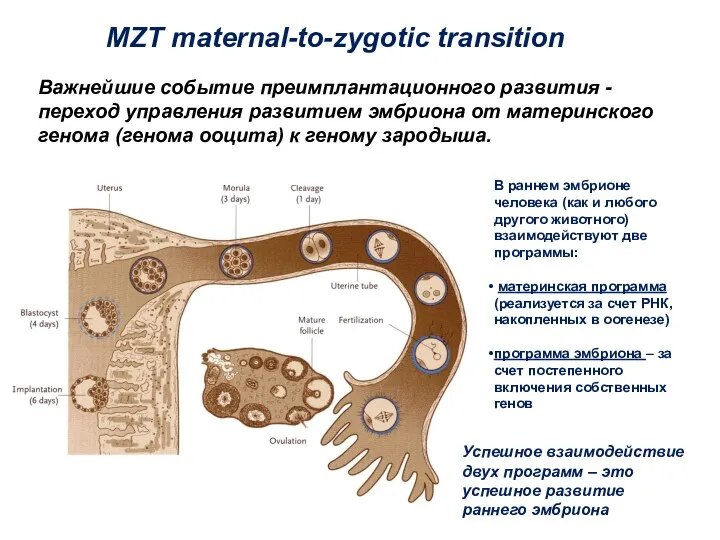
Важнейшие событие преимплантационного развития - переход управления развитием эмбриона от материнского
Важнейшие событие преимплантационного развития - переход управления развитием эмбриона от материнского
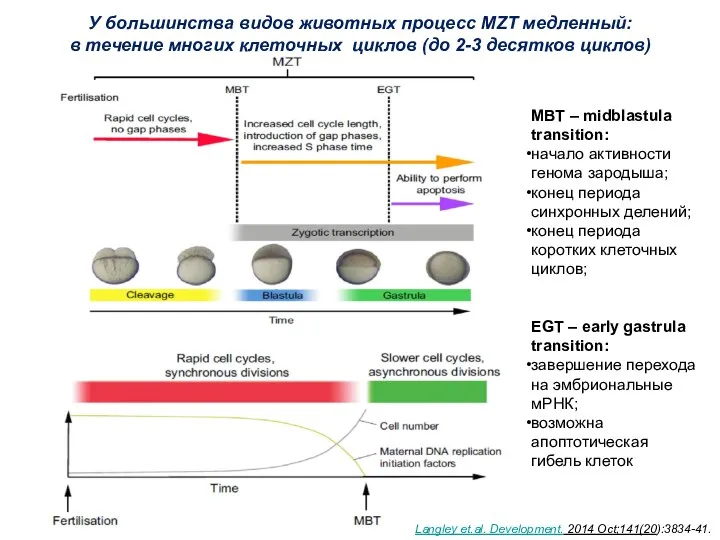
У большинства видов животных процесс MZT медленный:
в течение многих клеточных
У большинства видов животных процесс MZT медленный:
в течение многих клеточных
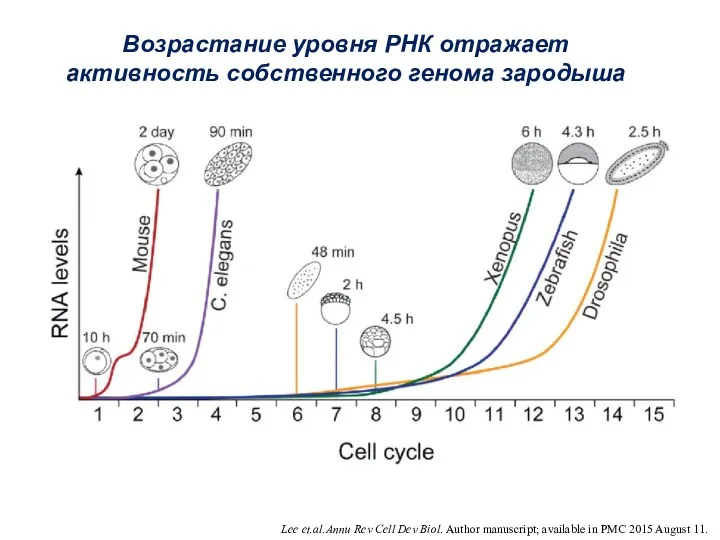
Lee et.al.Annu Rev Cell Dev Biol. Author manuscript; available in PMC
Lee et.al.Annu Rev Cell Dev Biol. Author manuscript; available in PMC
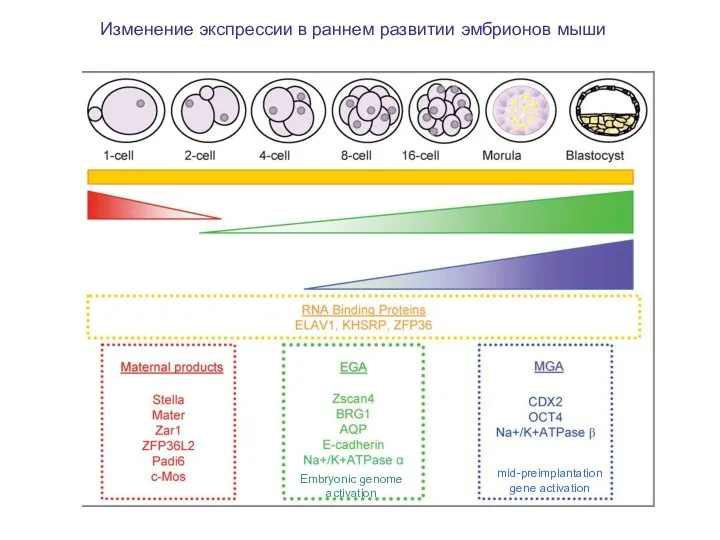
Изменение экспрессии в раннем развитии эмбрионов мыши
Изменение экспрессии в раннем развитии эмбрионов мыши

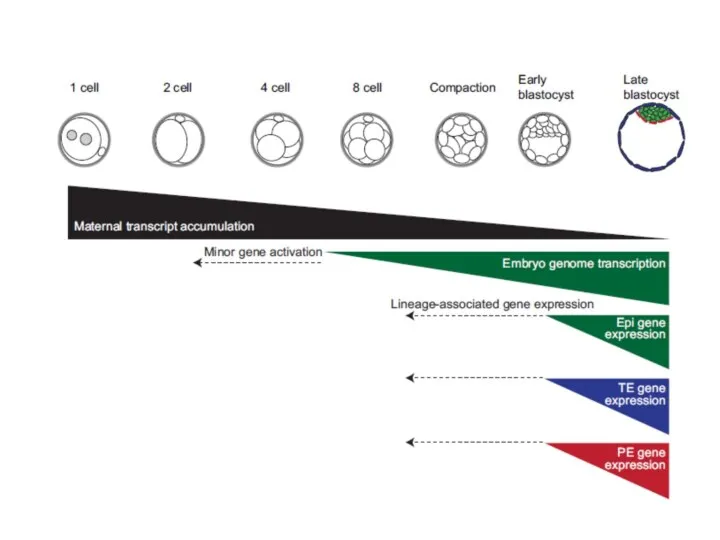
Активация генома эмбриона -
участие материнских генов
Филогенетическая дистанция между белками ZAR1 у
Активация генома эмбриона -
участие материнских генов
Филогенетическая дистанция между белками ZAR1 у
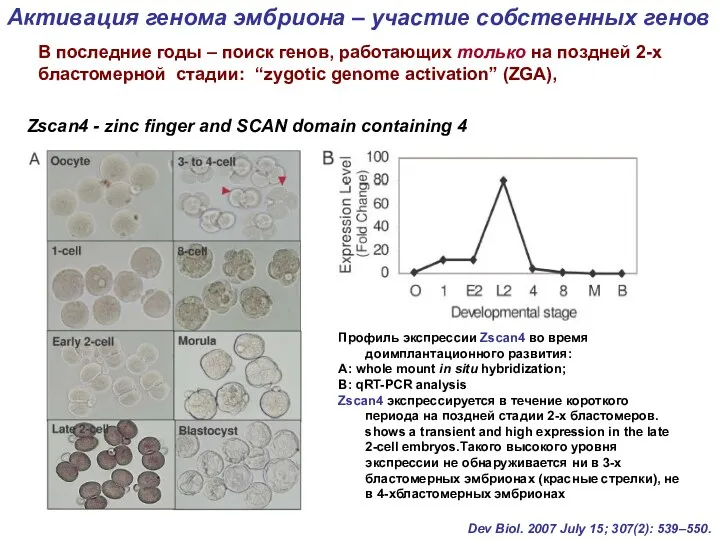
Профиль экспрессии Zscan4 во время доимплантационного развития:
А: whole mount in situ
Профиль экспрессии Zscan4 во время доимплантационного развития:
А: whole mount in situ
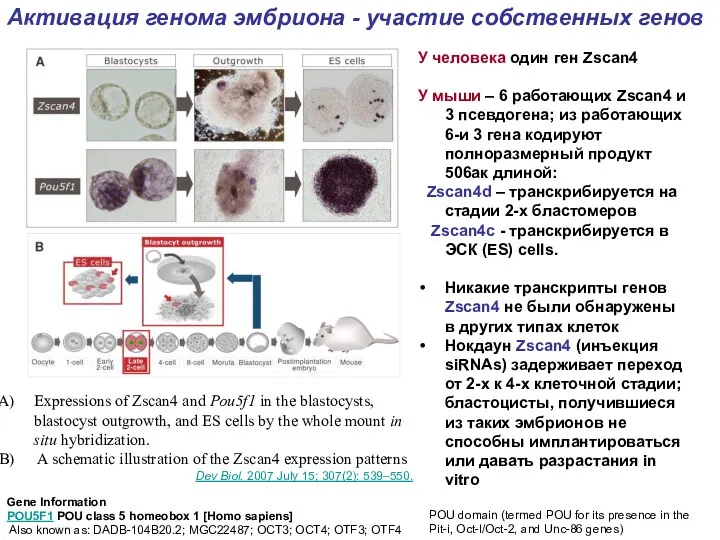
Expressions of Zscan4 and Pou5f1 in the blastocysts, blastocyst outgrowth, and
Expressions of Zscan4 and Pou5f1 in the blastocysts, blastocyst outgrowth, and

Определение 1: Тотипотентность – способность клетки развиваться в целый организм
Определение 2:
Определение 1: Тотипотентность – способность клетки развиваться в целый организм
Определение 2:
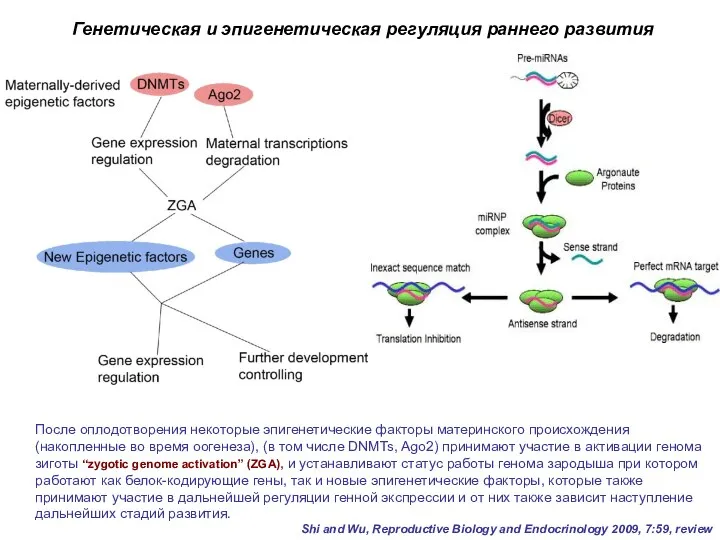
После оплодотворения некоторые эпигенетические факторы материнского происхождения (накопленные во время оогенеза),
После оплодотворения некоторые эпигенетические факторы материнского происхождения (накопленные во время оогенеза),
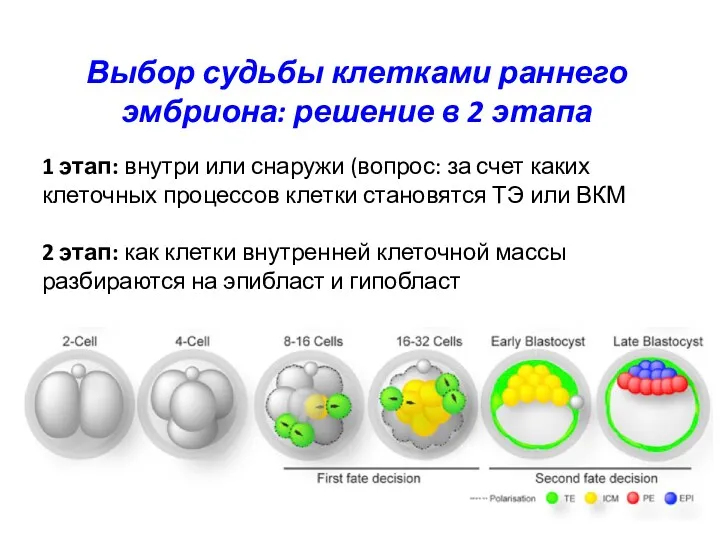
Выбор судьбы клетками раннего эмбриона: решение в 2 этапа
1 этап: внутри
Выбор судьбы клетками раннего эмбриона: решение в 2 этапа
1 этап: внутри
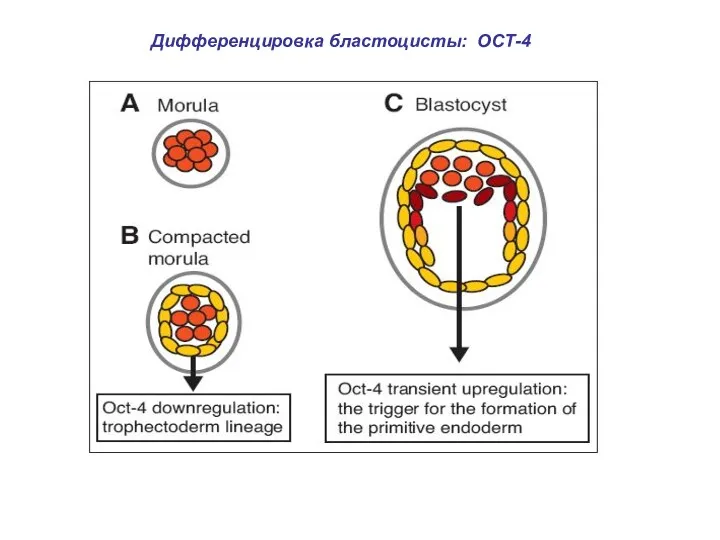
Дифференцировка бластоцисты: ОСТ-4
Дифференцировка бластоцисты: ОСТ-4
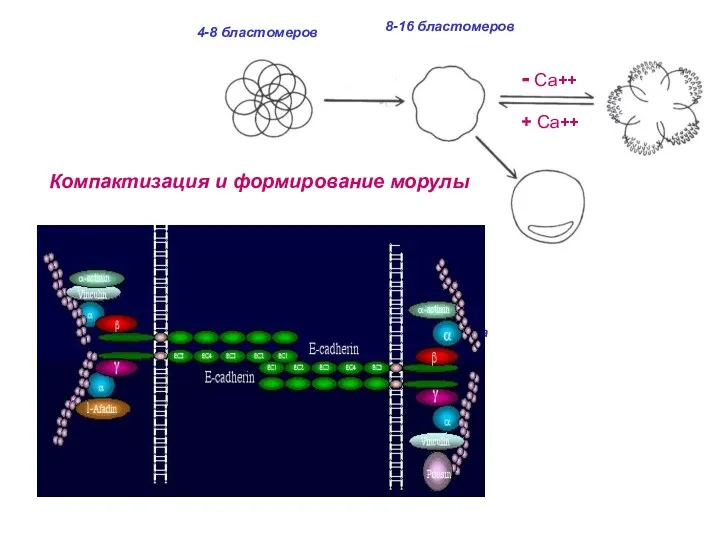
32 бластомера
и более
Компактизация и формирование морулы
32 бластомера
и более
Компактизация и формирование морулы
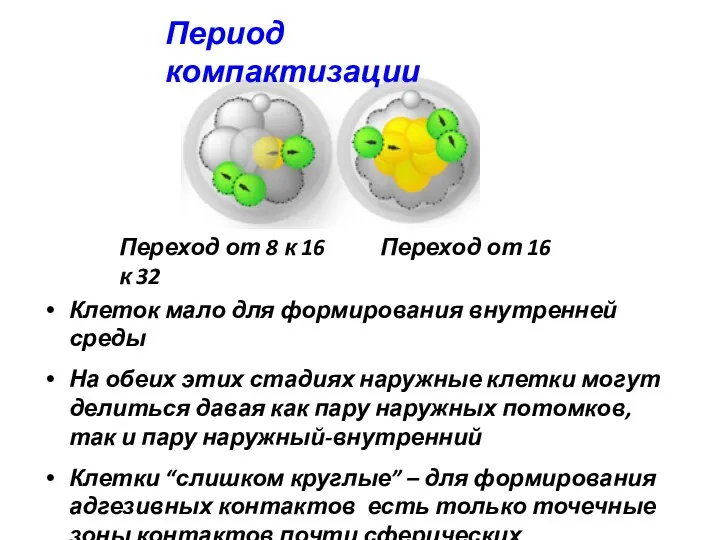
Период компактизации
Переход от 8 к 16 Переход от 16 к 32
Клеток
Период компактизации
Переход от 8 к 16 Переход от 16 к 32
Клеток
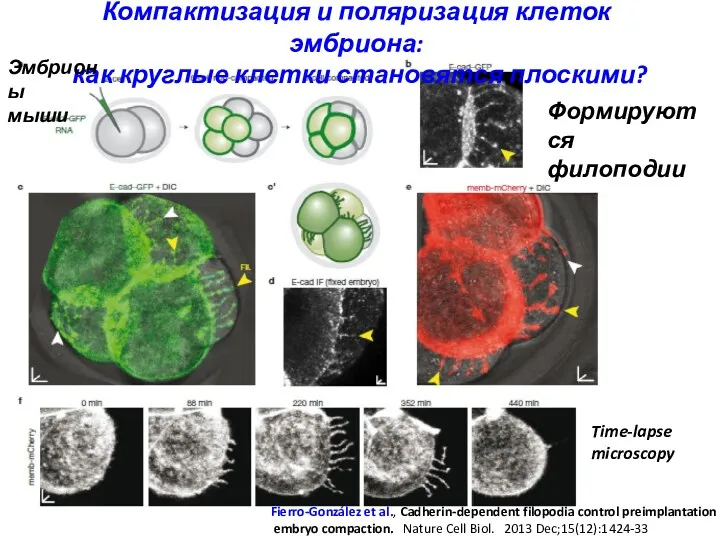
Компактизация и поляризация клеток эмбриона:
как круглые клетки становятся плоскими?
Fierro-González et
Компактизация и поляризация клеток эмбриона:
как круглые клетки становятся плоскими?
Fierro-González et
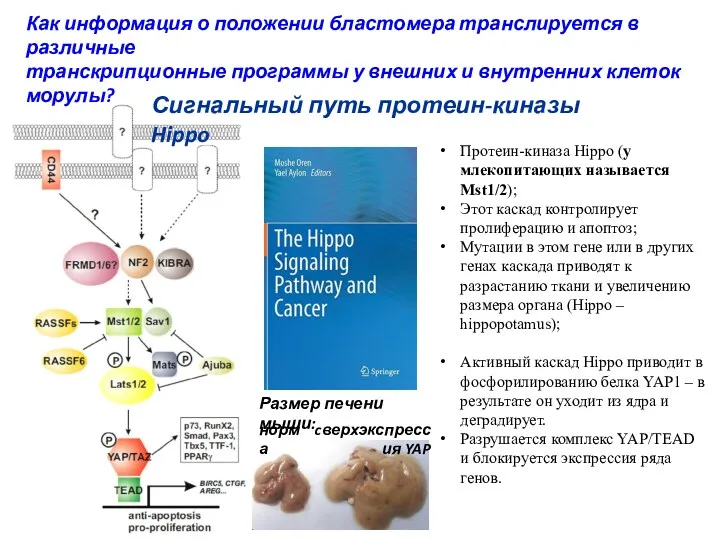
Протеин-киназа Hippo (у млекопитающих называется Mst1/2);
Этот каскад контролирует пролиферацию и апоптоз;
Протеин-киназа Hippo (у млекопитающих называется Mst1/2);
Этот каскад контролирует пролиферацию и апоптоз;
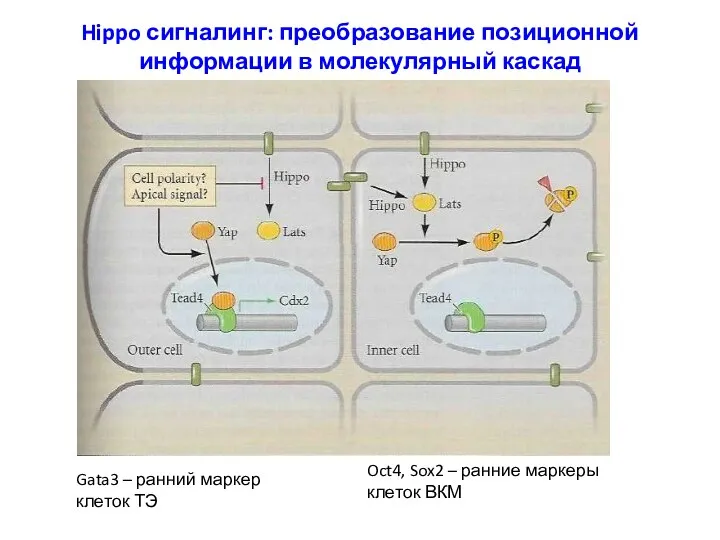
Hippo сигналинг: преобразование позиционной информации в молекулярный каскад
Gata3 – ранний маркер
Hippo сигналинг: преобразование позиционной информации в молекулярный каскад
Gata3 – ранний маркер

Становление полярности эмбриона на стадиях дробления:
апикально-базальные различия в строении кортикального
Становление полярности эмбриона на стадиях дробления:
апикально-базальные различия в строении кортикального
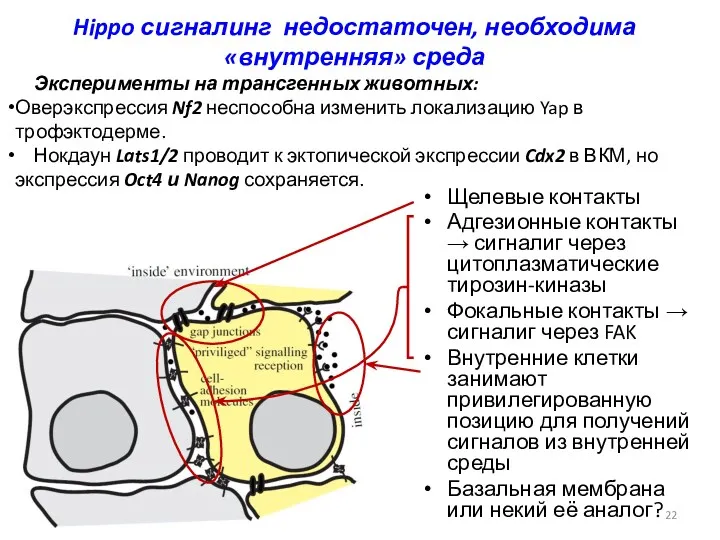
Hippo сигналинг недостаточен, необходима «внутренняя» среда
Щелевые контакты
Адгезионные контакты → сигналиг через
Hippo сигналинг недостаточен, необходима «внутренняя» среда
Щелевые контакты
Адгезионные контакты → сигналиг через
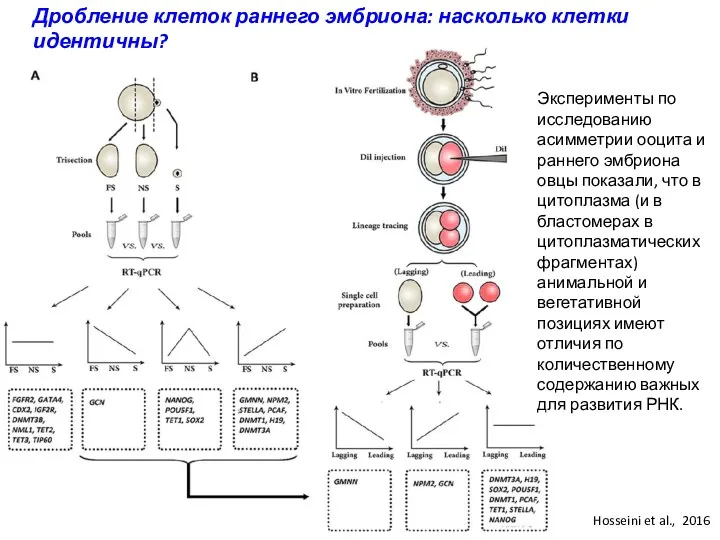
Дробление клеток раннего эмбриона: насколько клетки идентичны?
Эксперименты по исследованию асимметрии ооцита
Дробление клеток раннего эмбриона: насколько клетки идентичны?
Эксперименты по исследованию асимметрии ооцита
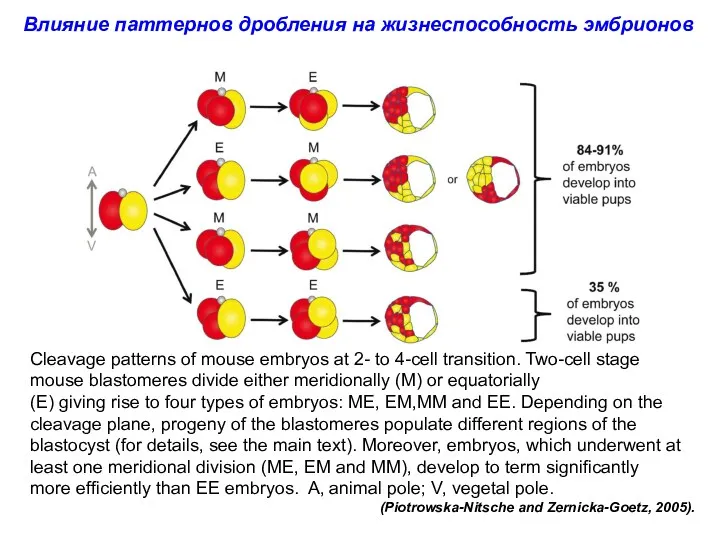
Cleavage patterns of mouse embryos at 2- to 4-cell transition. Two-cell
Cleavage patterns of mouse embryos at 2- to 4-cell transition. Two-cell
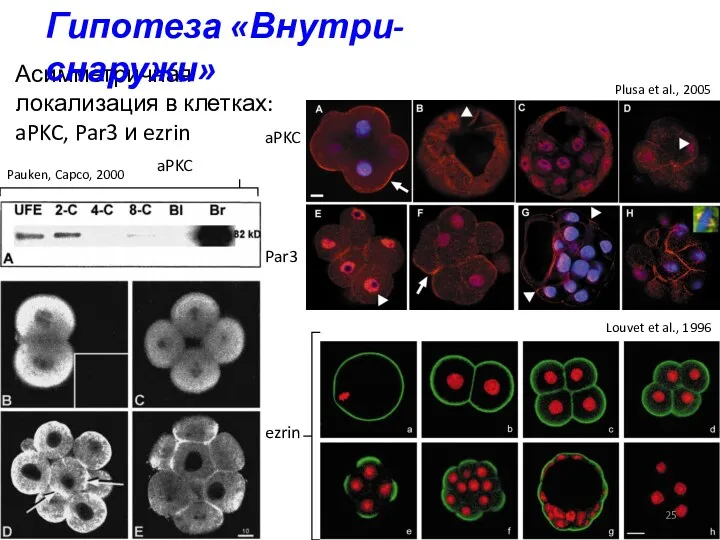
Асимметричная локализация в клетках: aPKC, Par3 и ezrin
ezrin
aPKC
aPKC
Par3
Louvet et al., 1996
Pauken,
Асимметричная локализация в клетках: aPKC, Par3 и ezrin
ezrin
aPKC
aPKC
Par3
Louvet et al., 1996
Pauken,
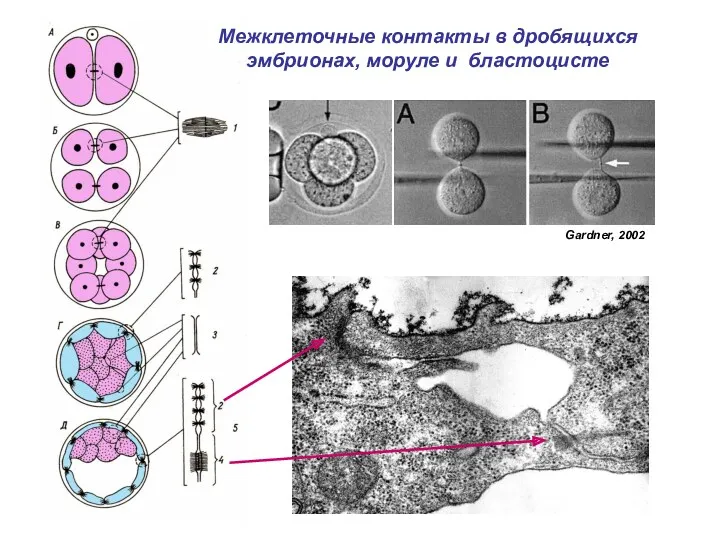
Межклеточные контакты в дробящихся эмбрионах, моруле и бластоцисте
Gardner, 2002
Межклеточные контакты в дробящихся эмбрионах, моруле и бластоцисте
Gardner, 2002
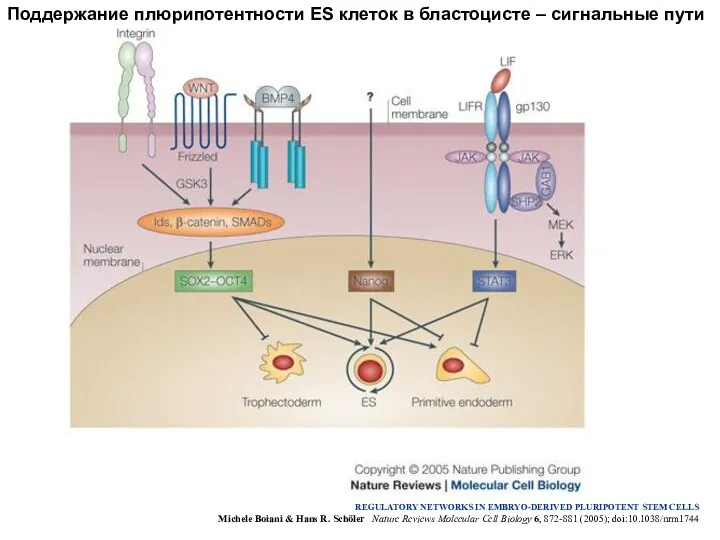
Поддержание плюрипотентности ES клеток в бластоцисте – сигнальные пути
REGULATORY NETWORKS IN
Поддержание плюрипотентности ES клеток в бластоцисте – сигнальные пути
REGULATORY NETWORKS IN
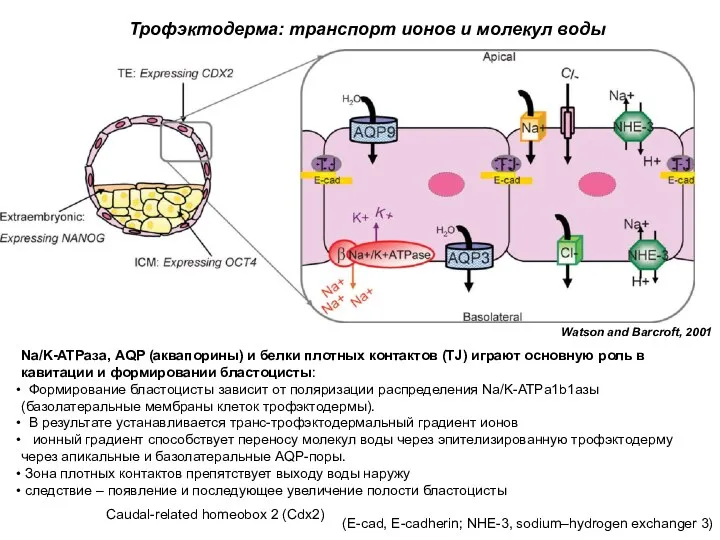
Na/K-ATPаза, AQP (аквапорины) и белки плотных контактов (TJ) играют основную роль
Na/K-ATPаза, AQP (аквапорины) и белки плотных контактов (TJ) играют основную роль

Классические представления:
В ранней бластоцисте все клетки ВКМ с самого начала
Классические представления:
В ранней бластоцисте все клетки ВКМ с самого начала
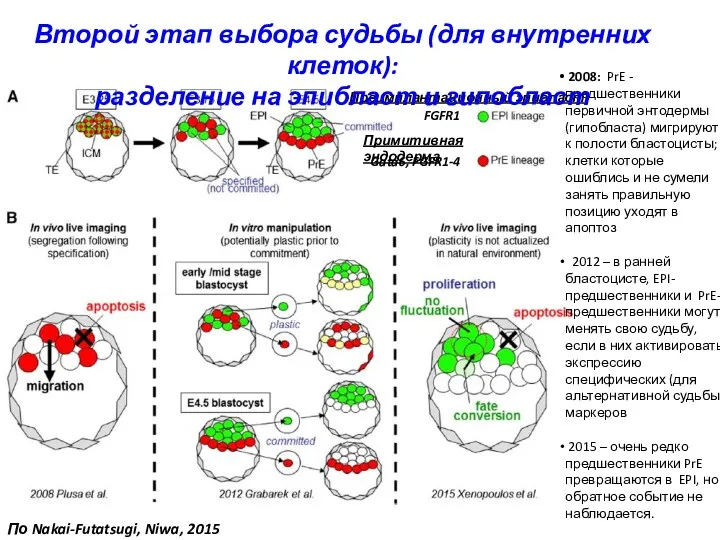
FGFR1
Gata6, FGFR1-4
Преимплантационный эпибласт
Примитивная эндодерма
2008: PrE - предшественники первичной энтодермы
FGFR1
Gata6, FGFR1-4
Преимплантационный эпибласт
Примитивная эндодерма
2008: PrE - предшественники первичной энтодермы
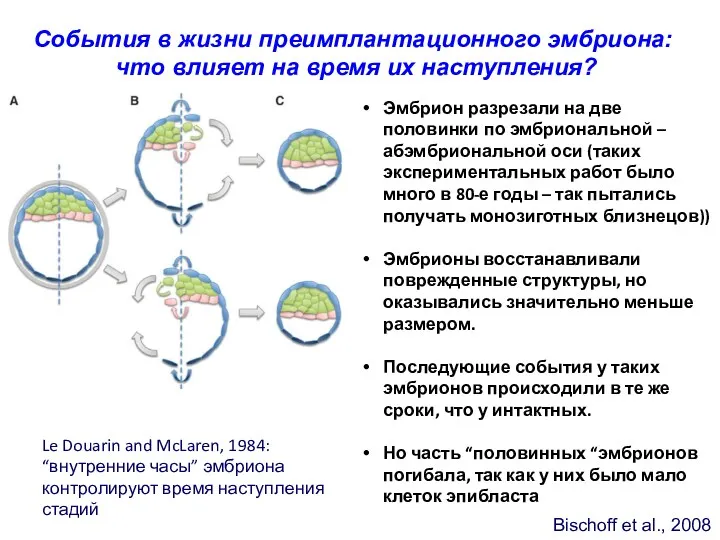
Bischoff et al., 2008
События в жизни преимплантационного эмбриона:
что влияет на
Bischoff et al., 2008
События в жизни преимплантационного эмбриона:
что влияет на
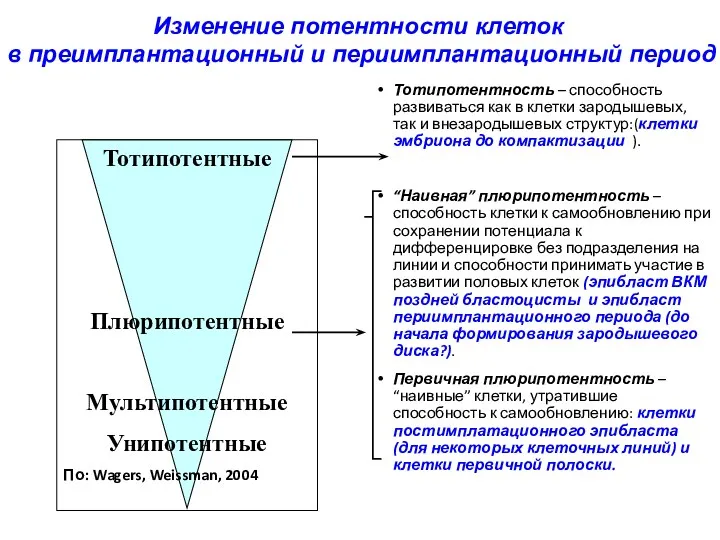
Тотипотентность – способность развиваться как в клетки зародышевых, так и внезародышевых
Тотипотентность – способность развиваться как в клетки зародышевых, так и внезародышевых
 Пигменты, от которых зависит жизнь.
Пигменты, от которых зависит жизнь. Красная книга по исчезающим видам животных и растений Алькеевского района
Красная книга по исчезающим видам животных и растений Алькеевского района Пищеварительная система и процесс пищеварения у человека
Пищеварительная система и процесс пищеварения у человека Растения, вызывающие преимущественно поражение органов дыхания и пищеварительного тракта
Растения, вызывающие преимущественно поражение органов дыхания и пищеварительного тракта Черепные нервы IX-XII
Черепные нервы IX-XII 7 самых ядовитых растений России
7 самых ядовитых растений России Структурные компоненты нуклеиновых кислот. Уровни организации ДНК и РНК
Структурные компоненты нуклеиновых кислот. Уровни организации ДНК и РНК Популяция – структурная единица вида; основная единица эволюции
Популяция – структурная единица вида; основная единица эволюции Липиды. Классификация липидов
Липиды. Классификация липидов Белки, жиры и углеводы
Белки, жиры и углеводы АСҚ пайдаланылу жолдары
АСҚ пайдаланылу жолдары Течение и вязкость жидкостей
Течение и вязкость жидкостей Основные оложения клеточной теории. 10 кл.
Основные оложения клеточной теории. 10 кл. Дробление. Деление зиготы
Дробление. Деление зиготы Головной мозг. Черепные нервы
Головной мозг. Черепные нервы От микроскопа к клетке.
От микроскопа к клетке. Историческое и индивидуальное в развитии
Историческое и индивидуальное в развитии Нервная ткань
Нервная ткань Урок биологии 9 класс Звёздный час – Человек
Урок биологии 9 класс Звёздный час – Человек Антропология, расы, конституции
Антропология, расы, конституции Устройство увеличительных приборов
Устройство увеличительных приборов Глюконеогенез
Глюконеогенез Физиологический процесс дыхание
Физиологический процесс дыхание Обмен веществ. Метаболизм и его функции
Обмен веществ. Метаболизм и его функции Ткани растений
Ткани растений Акция Помоги птице зимой!
Акция Помоги птице зимой! Далалық және ауылшаруашылық дақылдары
Далалық және ауылшаруашылық дақылдары Внутрішня будова птахів
Внутрішня будова птахів