- Регуляция клеточного деления

Содержание
- 2. Клеточный цикл
- 3. Определение фаз КЦ Период между двумя делениями называется интерфазой. В клетках млекопитающих интерфаза длится от 12
- 4. Сверочные точки КЦ
- 5. G1-сверочная точка Проверка повреждения ДНК Роль белка р53
- 6. G1 сверочные точки G1-postmitotic, G1-ps для G1-pre-S
- 7. G1 сверочные точки
- 8. Сверочные точки КЦ
- 9. Проверка завершенности S-фазы Клетка мониторится на присутствие фрагментов Оказаки
- 10. Проверка повреждений ДНК (на примере дрожжей)-1
- 11. Проверка повреждений ДНК (на примере дрожжей)-2
- 12. Сверочные точки КЦ
- 13. Проверка веретена Формирование центросомы
- 14. Центросома и система микротрубочек в профазной, метафазной и интерфазной клетках Тройное иммунофлуоресцентное окрашивание выявляет микротрубочки (красный
- 15. Упрощенная схема строения центросомы в интерфазных клетках млекопитающих в середине S-фазы клеточного цикла
- 16. Ультраструктура центросомы в митотической клетке млекопитающих
- 17. Роль BRCА1 в формировании центросомы BRCA1 - обладает убиквитин-зависимой лигазной активностью. Контролирует локализацию гамма-тубулина в центросоме
- 18. Регуляция центросом BRCA1-зависимой убиквитиновой лигазой и Aurora A киназой Результат потери функции BRCA1 и переэкспрессии AURRA
- 19. Фазы митоза
- 20. Разделение сестринских хроматид
- 21. Варианты КЦ
- 22. Циклины и их киназы
- 23. Циклины в КЦ
- 24. Циклины и их киназы в КЦ
- 25. Циклины в КЦ
- 26. Киназы в регуляции КЦ
- 27. Циклины и их киназы в КЦ
- 28. CDk, циклины и их субстраты-1
- 29. CDk, циклины и их субстраты-2
- 30. CDk, циклины и их субстраты-3
- 31. Активность CDk в тканях человека
- 32. Модель контроля G1 в стволовых клетках
- 33. Структура CDk2 человека, связанной с АТP Небольшая N-терминальная доля содержит В-полоску и PSTAIRE спираль. Показан большой
- 34. Структура циклина А(а) и Н(в) человека
- 35. Структура Cdk2-циклин А1-АТP комплекса человека Cdk2 расположена слева. Циклин А – справа. Т-петля обозначена черной стрелкой.
- 36. Регуляция Cdk активности фосфорилированием
- 37. Регуляция митоза
- 38. Polo-киназа в регуляции митоза
- 39. Ингибиторы Cdks 2 Семейства CKI: Cip/Kip (Cdk ингибиторные белки) р21, р27,р57 – ингибируют Cdk2 и Cdk4/6
- 40. Структура связи комплекса Сdk2-циклин А1 с усеченным пептидом р27 Пептид р27 (затемнен) протянут через вершину комплекса,
- 41. Контроль митоза киназой М фазы
- 42. Cdk-комплексы в G1 и S фазы
- 43. Регуляция циклинов - Транскрипция (E2F стимулирует ЦА и Е в S фазе) или ЦD RAS-RAF-MAP киназный
- 44. Структура протеосомы Коровая частица (CP) СР состоит из 2 копий 14 различных белков. Они объединены в
- 45. Убиквитинация
- 46. Выход из деления-1
- 47. Выход из деления-2
- 48. Деструкция митотических циклинов Циклосома АРС (anaphase promoting complex) – метафаза-анафаза (Cdc20) и выход из митоза (Hct1).
- 49. Регуляция циклинов в КЦ
- 50. Нобелевская премия по физиологии и медицине за 2001 г. Присуждена американцу Леланду Хартвеллу и англичанам Тимоти
- 51. Сверочные точки в КЦ
- 52. Киназы КЦ в развитии рака-1
- 53. Киназы КЦ в развитии рака-2
- 54. Киназы КЦ в развитии рака-3
- 55. Экспрессия киназ КЦ в опухолях человека
- 56. Киназы КЦ в терапии рака
- 57. 2. Раковые супрессорные гены или онкосупрессоры RB р53 (ТР53) Развитие рака происходит из-за потери функции РСГ
- 58. RB Белок
- 59. Развитие ретинобластомы (q14 делеция в 13-й хромосоме)
- 60. Роль вирусных онкогенов
- 61. Комплекс RB с вирусным белком
- 62. RB белок в регуляции КЦ
- 63. RB белок в регуляции КЦ
- 64. Фосфорилирование RB контролируется в КЦ Блокирование КЦ прекращается, когда RB фосфорилируется (в норме) или когда это
- 65. Структура RB белков rb1 (encoding P105) {13q14.1–q14.2}, rb2 (encoding P130) {16q12.2}, rbl1 (retinoblastoma-like-1, encoding P107) {20q11.2}
- 66. Семейство RB белков
- 67. RB белки в клеточном цикле
- 68. Модель регуляции КЦ карманными белками
- 69. Взаимодействие регуляторных белков в G1-фазе
- 70. Семейство E2F млекопитающих
- 71. Экспрессия E2Fs во время КЦ
- 72. E2Fs и рак
- 73. Парадигма взаимодействия E2F-RB в раковых клетках
- 74. В ядре непролиферирующих клеток остается RB гипофосфорилированным и формирует белковый комплекс E2F-RB, т.о. ингибируются гены, регулирующие
- 75. p16 в регуляции КЦ
- 76. Р16 и старение
- 77. p21 и p27 в регуляции КЦ
- 78. Белок p53 – ключевой онкосупрессор. Роль в онкологии
- 79. Структура и функции р53 - ДНК-связывающий белок (впервые описан в 1979 г.) - Фактор транскрипции (активация,
- 80. Родословная семьи с синдромом Li-Fraumeni (1969) Частота: 1:5,000 человек
- 81. Последовательности, узнаваемые р53 PuPuPuC(A/T)(T/A)GPyPyPy
- 82. Доменная структура р53 белка
- 83. Доменная структура р53 белка
- 84. Структура корового домена р53
- 85. 3D структура p53
- 86. р53 регулирует рост
- 87. р53 отвечает на повреждение ДНК
- 88. Неспецифические последовательности ДНК, с которыми может связываться р53
- 89. р53 и апоптоз
- 90. p53 активирует несколько независимых путей. Активация ареста КЦ вместе с геномной нестабильностью – альтернатива апоптозу
- 91. Регуляция активности р53 1. В норме его экспрессия невелика и контролируется убиквинированием и протеосомной деградацией. 2.
- 92. Каждый путь, который активирует р53, вызывает модификацию определенных остатков аминокислот белка
- 94. MDM2 в регуляции р53 1991 г. - впервые показано его взаимодействие с р53
- 95. Структура белка Mdm2
- 96. Mdm2 регулирует p53-зависимый ответ на повреждение ДНК Mdm2 связывается с p53, который регулирует многие гены. До
- 97. Комплексный контроль р53
- 98. Убиквитин-E3-лигазы, регулирующие активность p53
- 99. Активация р53 в ответ на стресс
- 100. Регуляция экспрессии гена Mdm2 У гена Mdm2 два промотора. P1 контролирует базальную экспрессию, P2 промотор использует
- 101. Онкогенные пути усиливают экспрессию Mdm2
- 102. Онкосупрессоры контролируют функцию Mdm2 Белок p14ARF – негативный регулятор Mdm2, он транскрипционно регулируется ФТ семейства E2F.
- 103. Р16 и ARF Ген p16 охватывает локус Ink4b/ARF/Ink4a на хромосоме 9p21. Две различных рамки считывания используются
- 104. РСГ осуществляют контроль КЦ
- 105. Р53 – мишень в химиотерапии В нормальных клетках p53 тонко регулируется. MDM2 стимулирует p53 к протеосомной
- 106. Гены-мишени р53
- 107. 2011 г.: Согласно IARC, выявляется более, чем 26 000 соматических мутаций в (TP53 database version R14
- 108. Наиболее часто встречаемые мутации р53
- 109. Относительная частота герминальных мутаций в p53
- 110. Изоформы р53
- 111. р63 и р53 белки (SAM – sterile alpha motive)
- 112. Экспрессия р63 в опухолях
- 113. Восстановление активности р53 Введение в опухоль дикого p53 с помощью репликативно-дефектного аденовирусного вектора CP-31398 – молекула
- 114. Маркеры РМЖ на 17-хромосоме Wei Zhang and Yingyan Yu «The Important Molecular Markers on Chromosome 17
- 115. Функции маркеров
- 116. Примеры различных РСГ
- 119. Примеры нокаутных мышей с инактивированными РСГ или ПО
- 121. Скачать презентацию
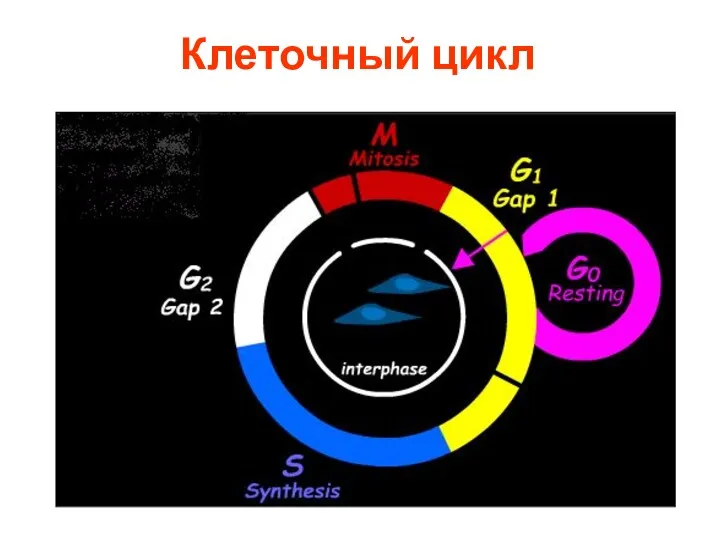
Клеточный цикл
Клеточный цикл

Определение фаз КЦ
Период между двумя делениями называется интерфазой. В клетках млекопитающих
Определение фаз КЦ
Период между двумя делениями называется интерфазой. В клетках млекопитающих
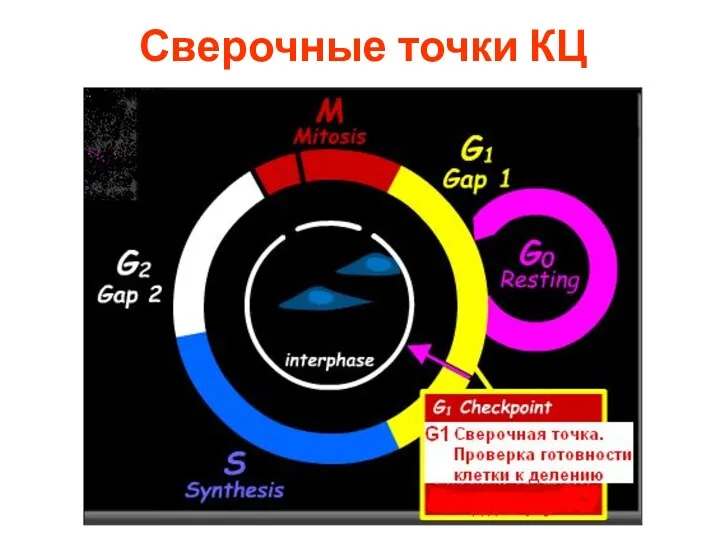
Сверочные точки КЦ
Сверочные точки КЦ

G1-сверочная точка
Проверка повреждения ДНК
Роль белка р53
G1-сверочная точка
Проверка повреждения ДНК
Роль белка р53
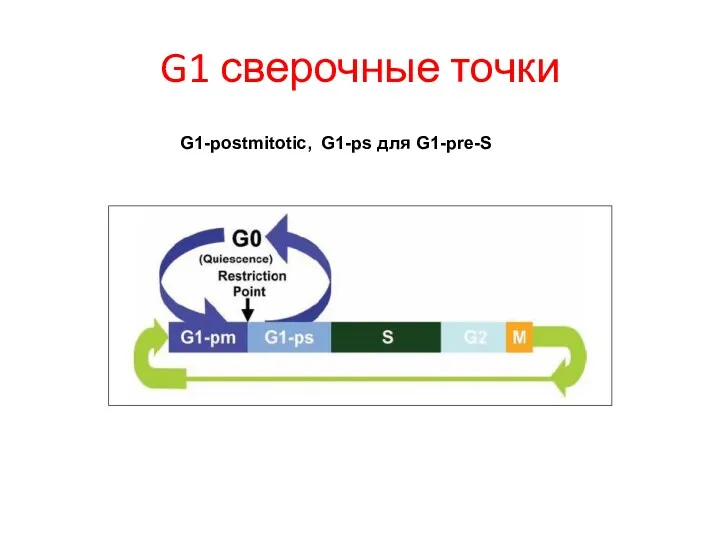
G1 сверочные точки
G1-postmitotic, G1-ps для G1-pre-S
G1 сверочные точки
G1-postmitotic, G1-ps для G1-pre-S
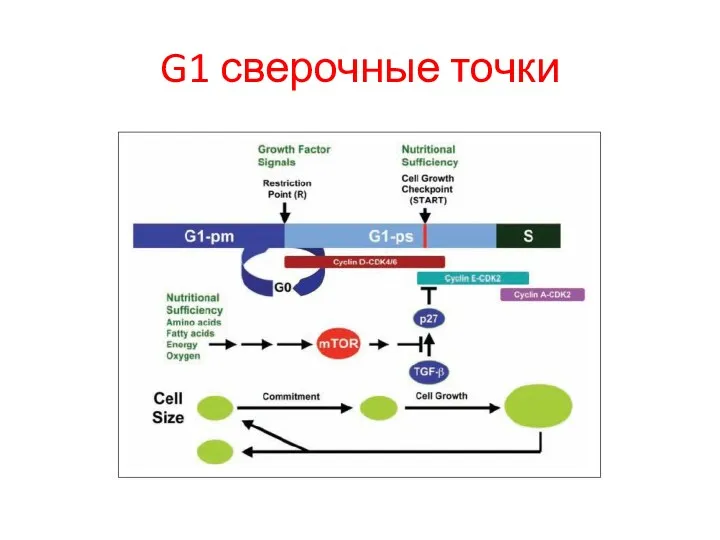
G1 сверочные точки
G1 сверочные точки
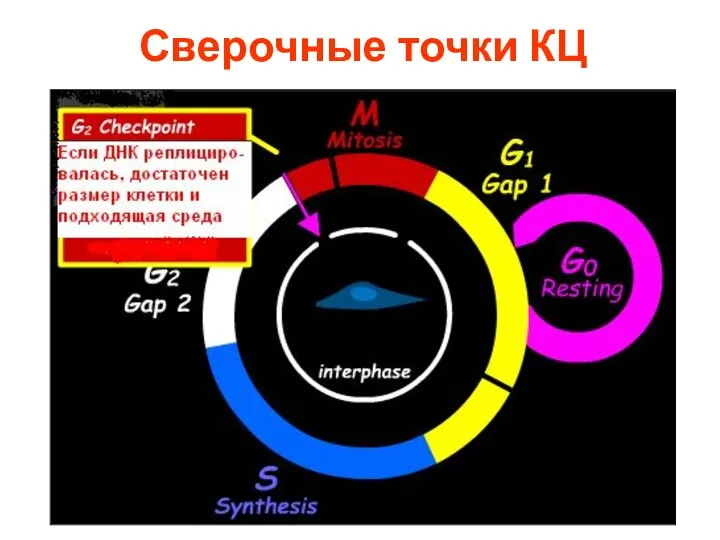
Сверочные точки КЦ
Сверочные точки КЦ
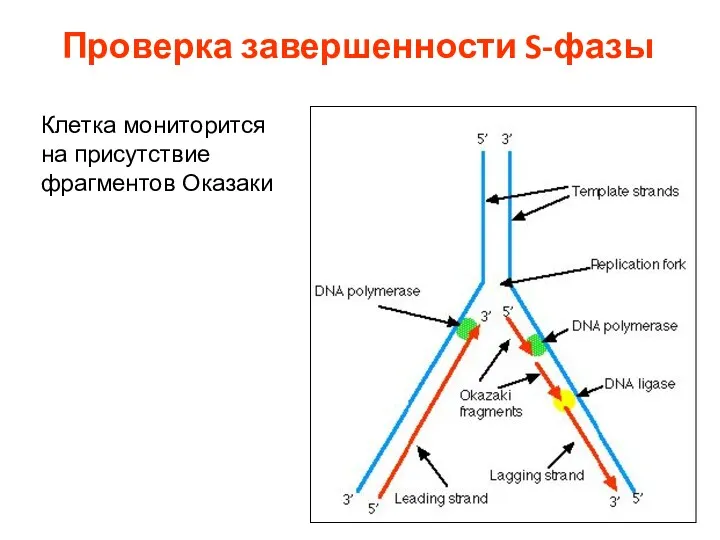
Проверка завершенности S-фазы
Клетка мониторится на присутствие фрагментов Оказаки
Проверка завершенности S-фазы
Клетка мониторится на присутствие фрагментов Оказаки
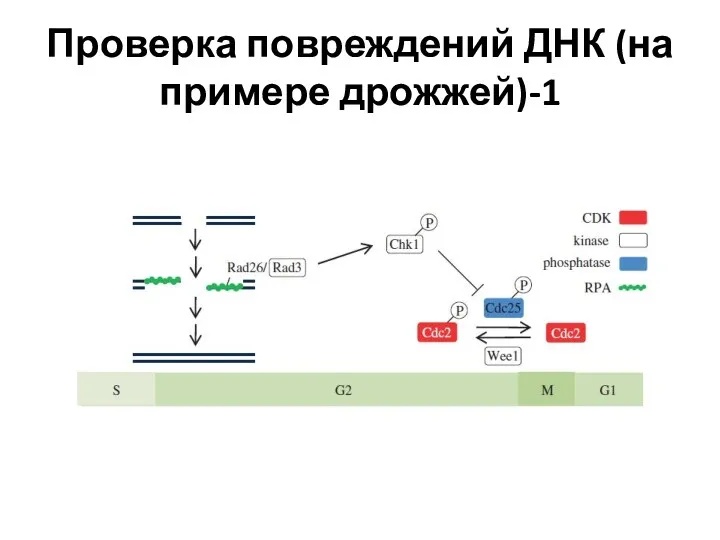
Проверка повреждений ДНК (на примере дрожжей)-1
Проверка повреждений ДНК (на примере дрожжей)-1
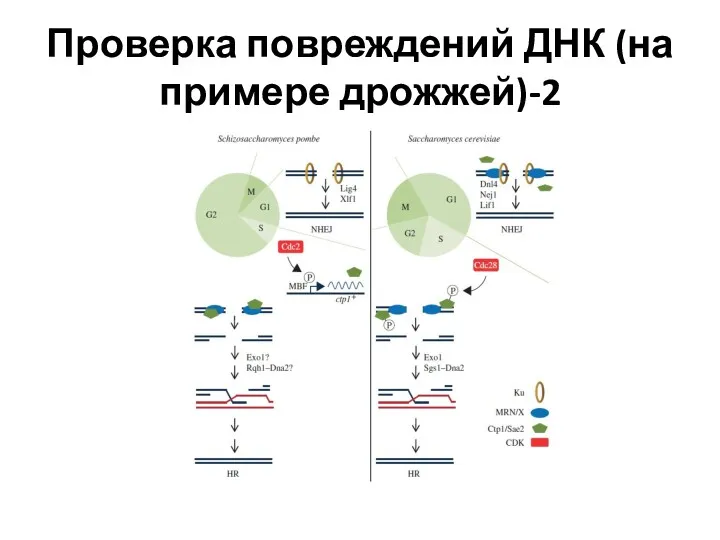
Проверка повреждений ДНК (на примере дрожжей)-2
Проверка повреждений ДНК (на примере дрожжей)-2
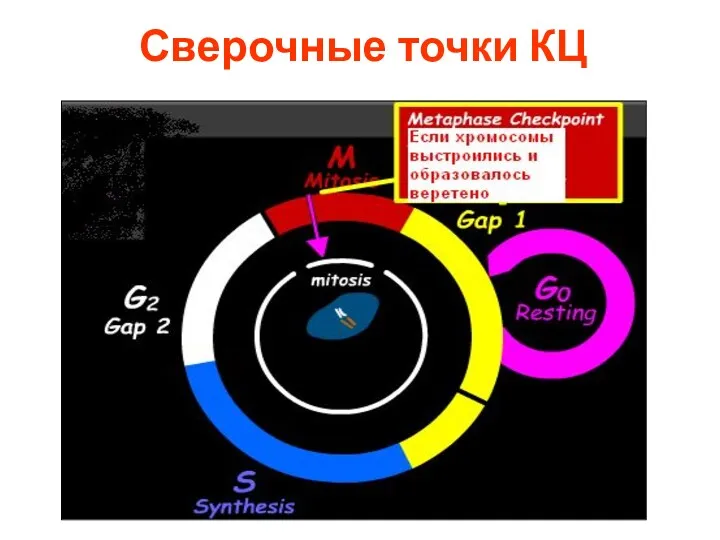
Сверочные точки КЦ
Сверочные точки КЦ
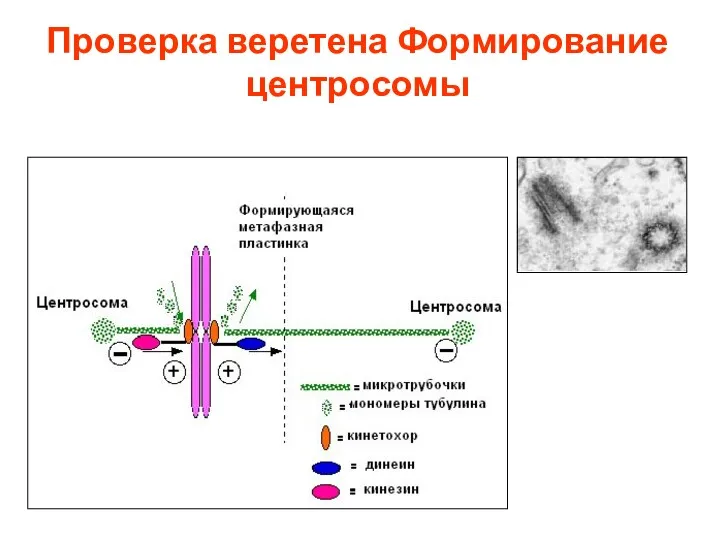
Проверка веретена Формирование центросомы
Проверка веретена Формирование центросомы
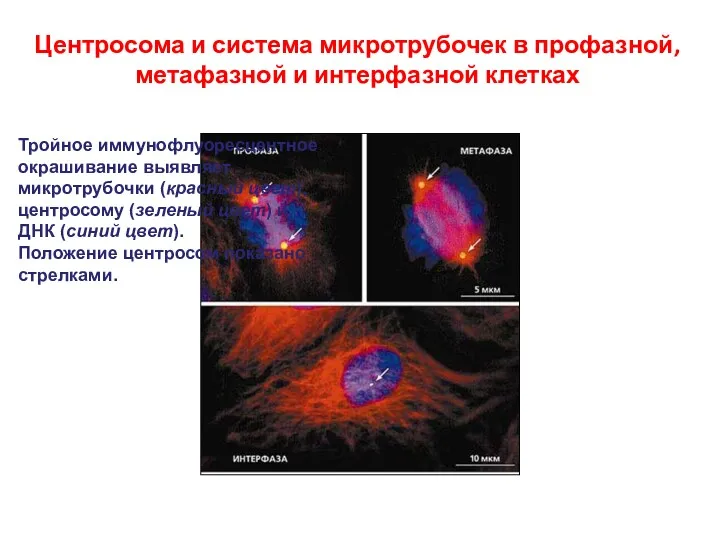
Центросома и система микротрубочек в профазной, метафазной и интерфазной клетках
Тройное иммунофлуоресцентное
Центросома и система микротрубочек в профазной, метафазной и интерфазной клетках
Тройное иммунофлуоресцентное
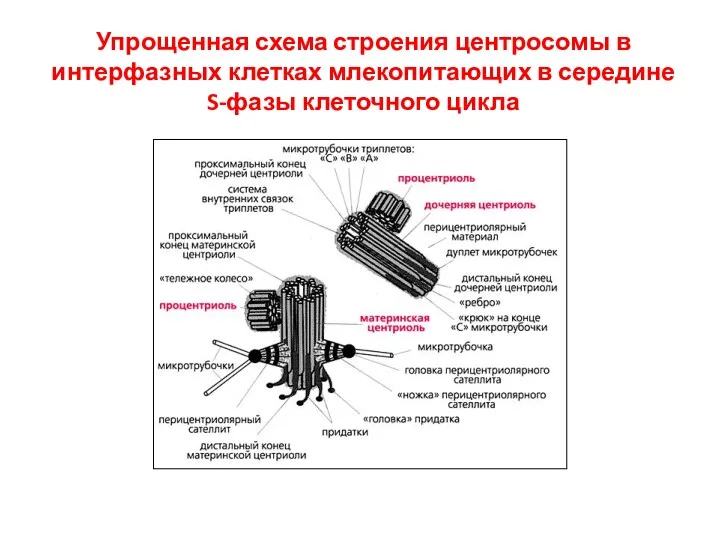
Упрощенная схема строения центросомы в интерфазных клетках млекопитающих в середине S-фазы
Упрощенная схема строения центросомы в интерфазных клетках млекопитающих в середине S-фазы
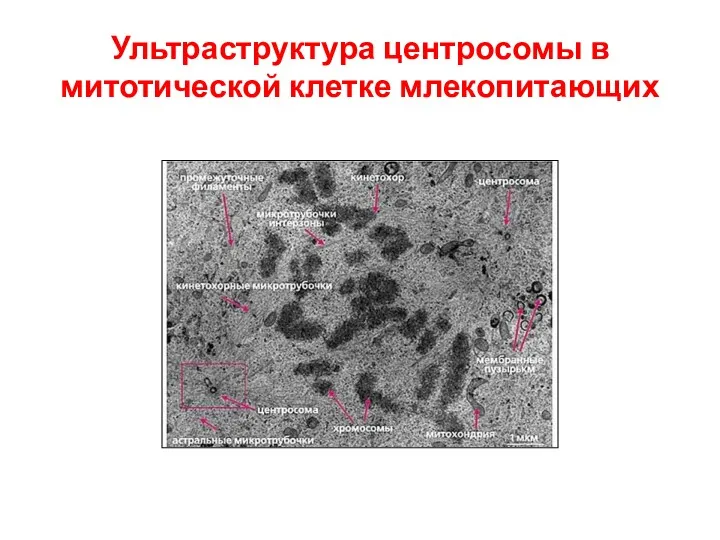
Ультраструктура центросомы в митотической клетке млекопитающих
Ультраструктура центросомы в митотической клетке млекопитающих

Роль BRCА1 в формировании центросомы
BRCA1 - обладает убиквитин-зависимой лигазной активностью.
Роль BRCА1 в формировании центросомы
BRCA1 - обладает убиквитин-зависимой лигазной активностью.

Регуляция центросом BRCA1-зависимой убиквитиновой лигазой и
Aurora A киназой
Результат потери функции
Регуляция центросом BRCA1-зависимой убиквитиновой лигазой и
Aurora A киназой
Результат потери функции
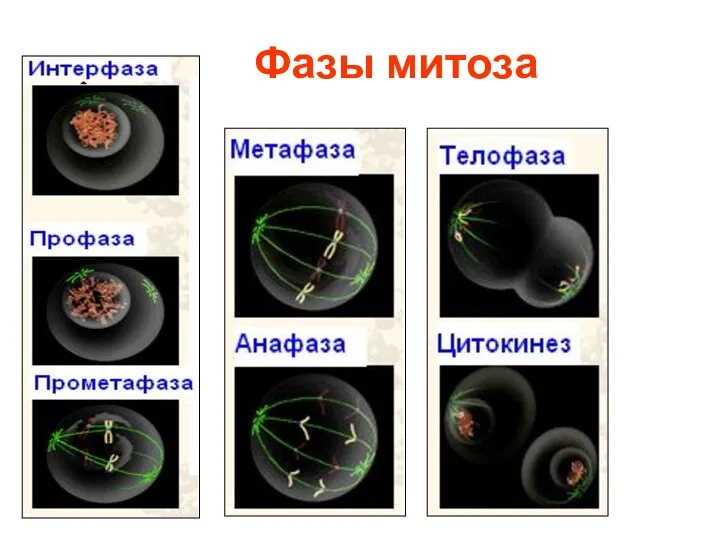
Фазы митоза
Фазы митоза
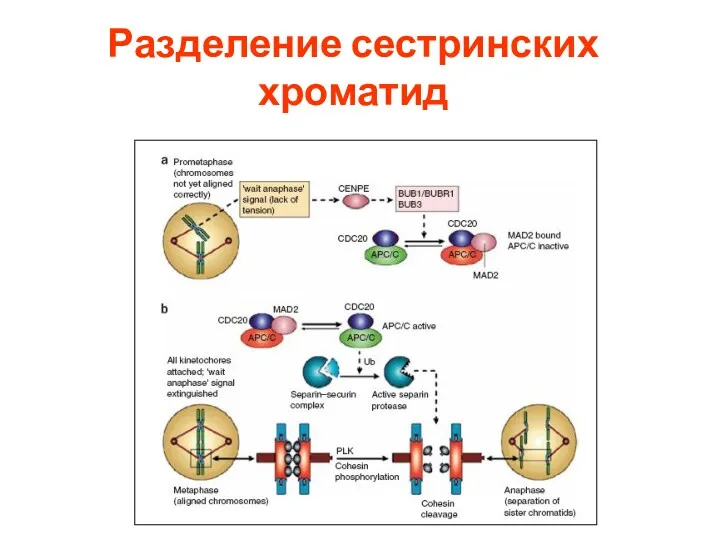
Разделение сестринских хроматид
Разделение сестринских хроматид
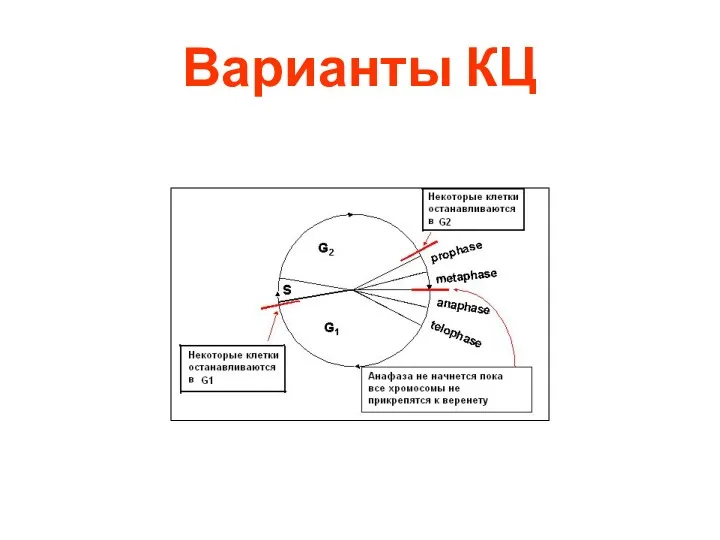
Варианты КЦ
Варианты КЦ
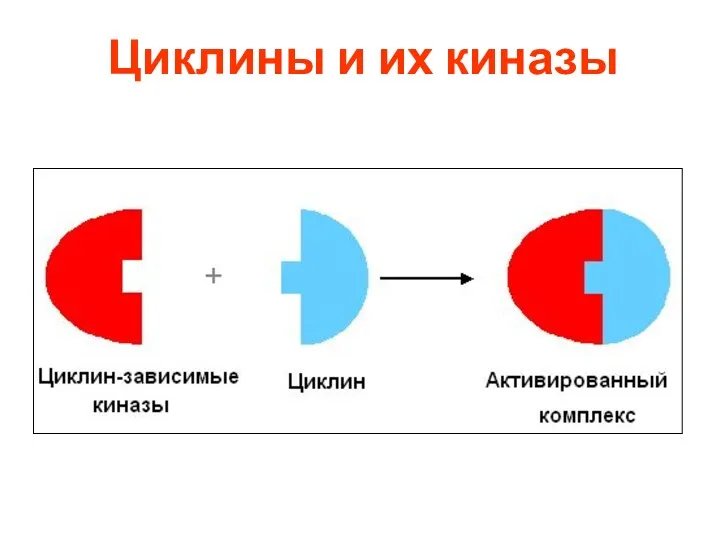
Циклины и их киназы
Циклины и их киназы
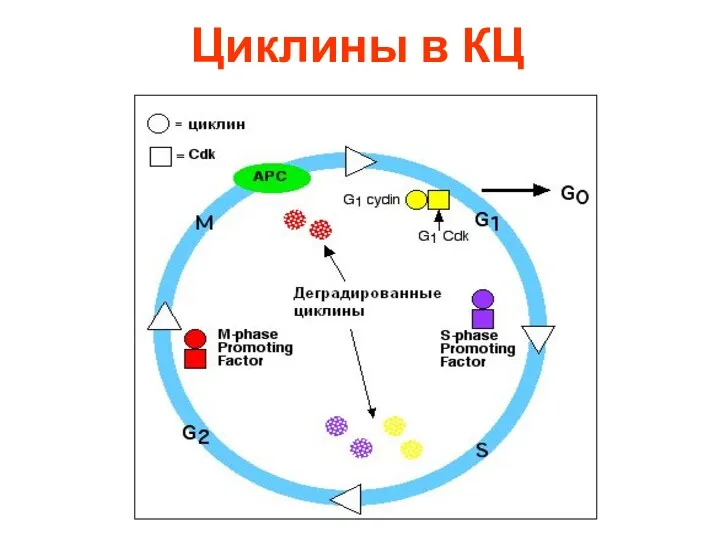
Циклины в КЦ
Циклины в КЦ
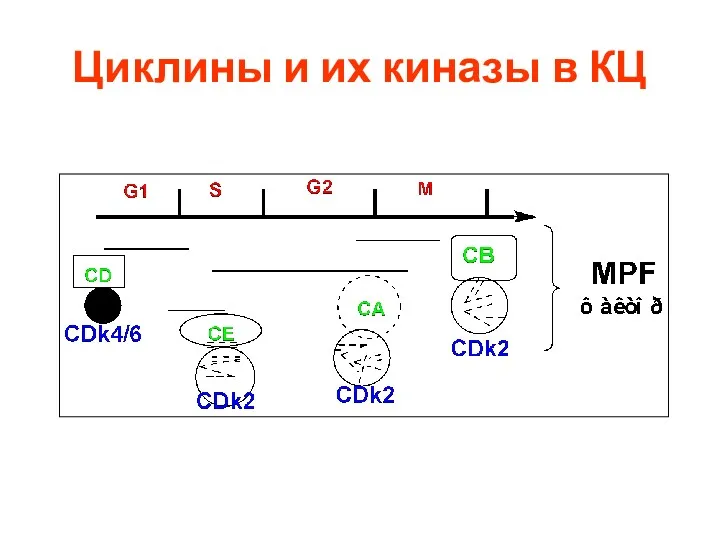
Циклины и их киназы в КЦ
Циклины и их киназы в КЦ
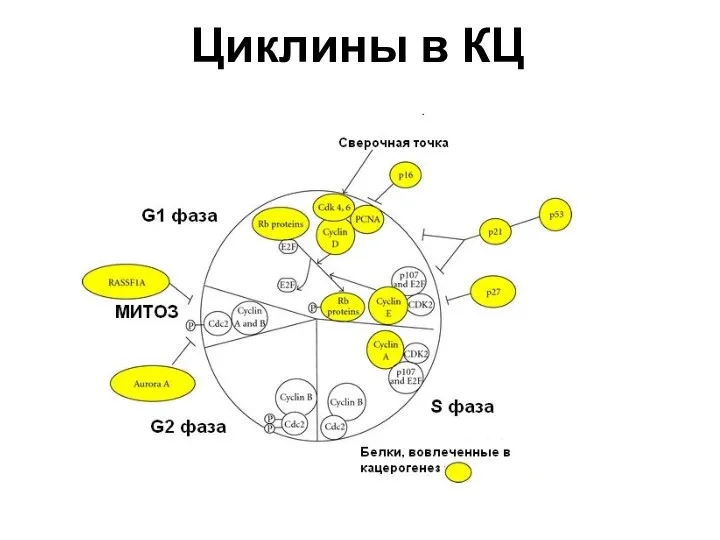
Циклины в КЦ
Циклины в КЦ
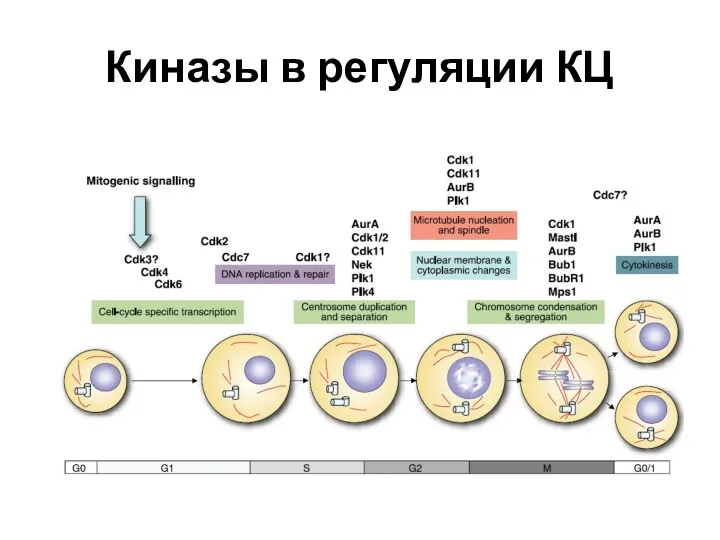
Киназы в регуляции КЦ
Киназы в регуляции КЦ
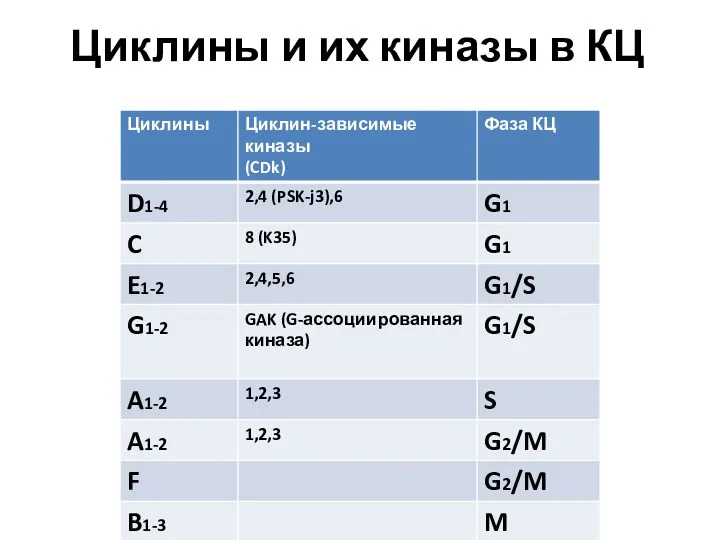
Циклины и их киназы в КЦ
Циклины и их киназы в КЦ

CDk, циклины и их субстраты-1
CDk, циклины и их субстраты-1
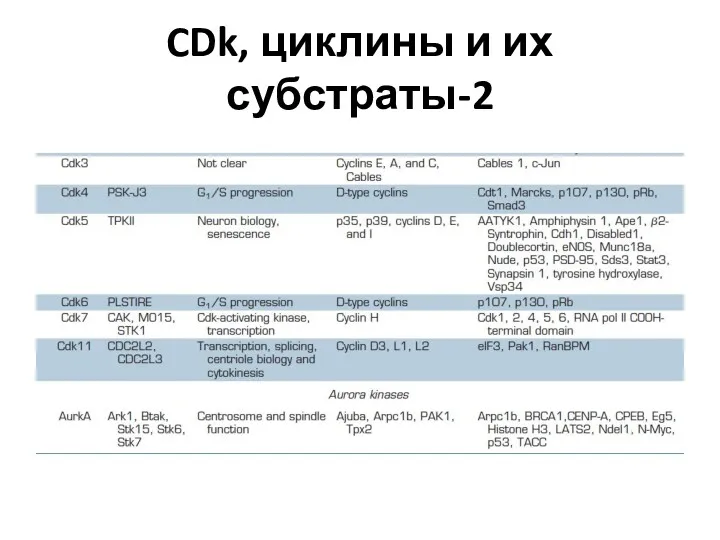
CDk, циклины и их субстраты-2
CDk, циклины и их субстраты-2
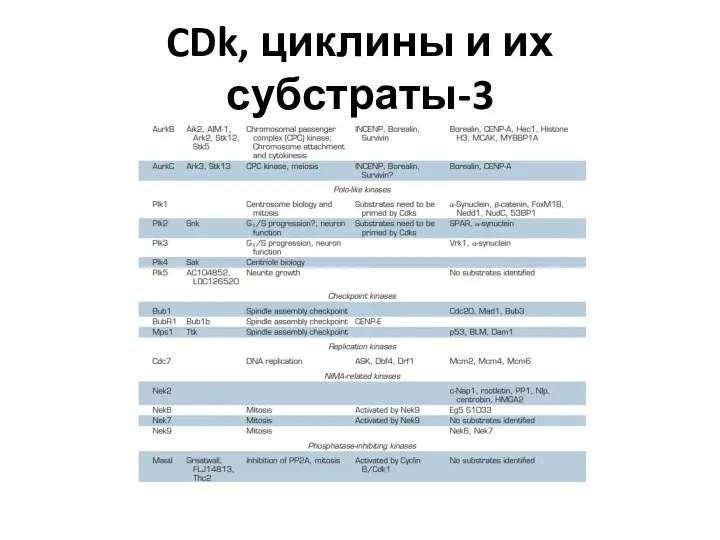
CDk, циклины и их субстраты-3
CDk, циклины и их субстраты-3
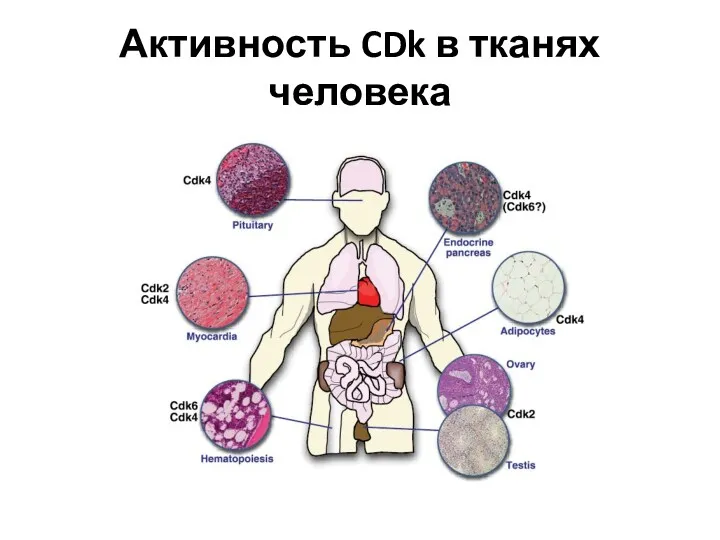
Активность CDk в тканях человека
Активность CDk в тканях человека
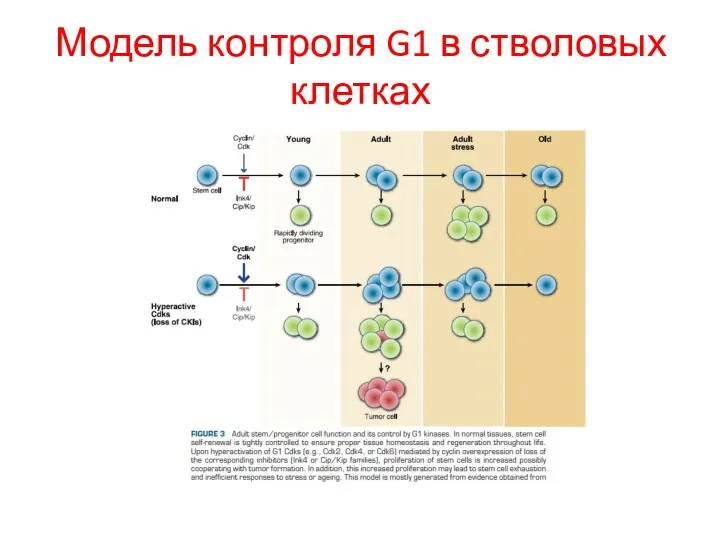
Модель контроля G1 в стволовых клетках
Модель контроля G1 в стволовых клетках
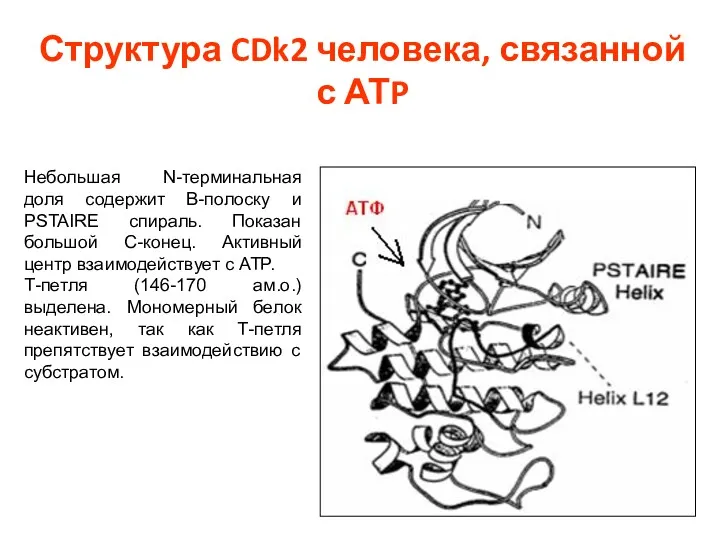
Структура CDk2 человека, связанной с АТP
Небольшая N-терминальная доля содержит В-полоску и
Структура CDk2 человека, связанной с АТP
Небольшая N-терминальная доля содержит В-полоску и
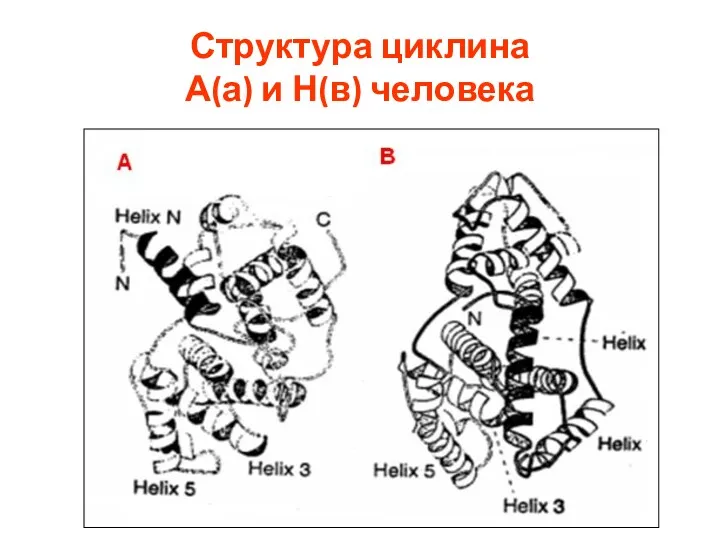
Структура циклина
А(а) и Н(в) человека
Структура циклина
А(а) и Н(в) человека

Структура Cdk2-циклин
А1-АТP комплекса человека
Cdk2 расположена слева. Циклин А –
Структура Cdk2-циклин
А1-АТP комплекса человека
Cdk2 расположена слева. Циклин А –
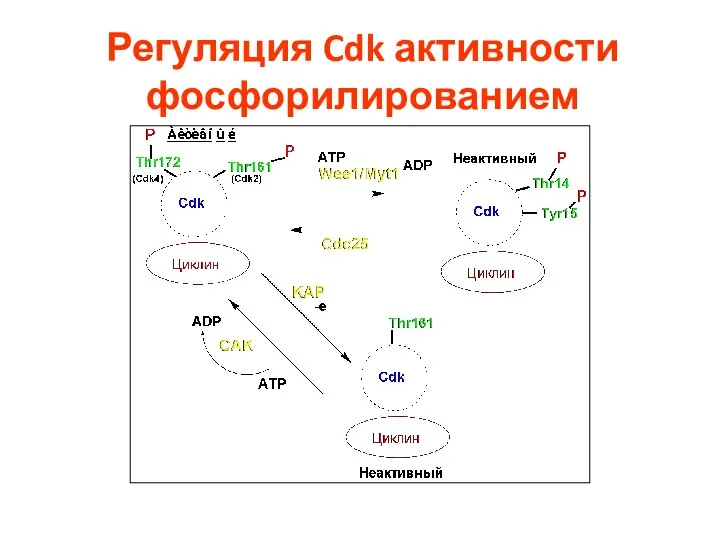
Регуляция Cdk активности фосфорилированием
Регуляция Cdk активности фосфорилированием
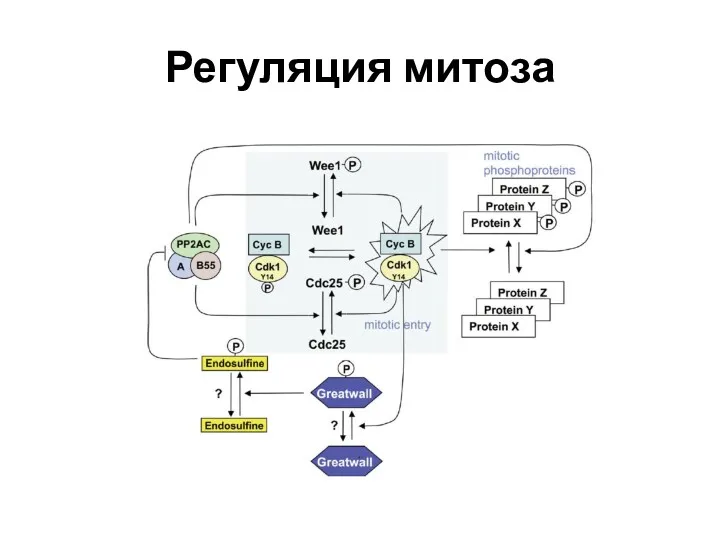
Регуляция митоза
Регуляция митоза
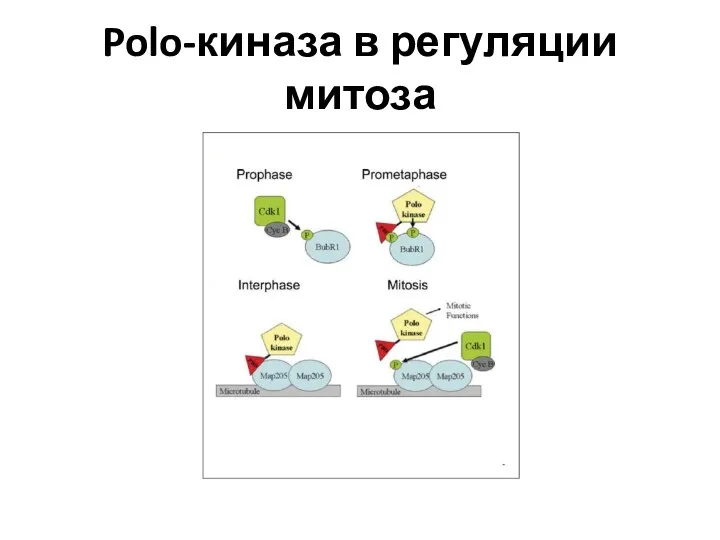
Polo-киназа в регуляции митоза
Polo-киназа в регуляции митоза

Ингибиторы Cdks
2 Семейства CKI:
Cip/Kip (Cdk ингибиторные белки) р21, р27,р57 – ингибируют
Ингибиторы Cdks
2 Семейства CKI:
Cip/Kip (Cdk ингибиторные белки) р21, р27,р57 – ингибируют
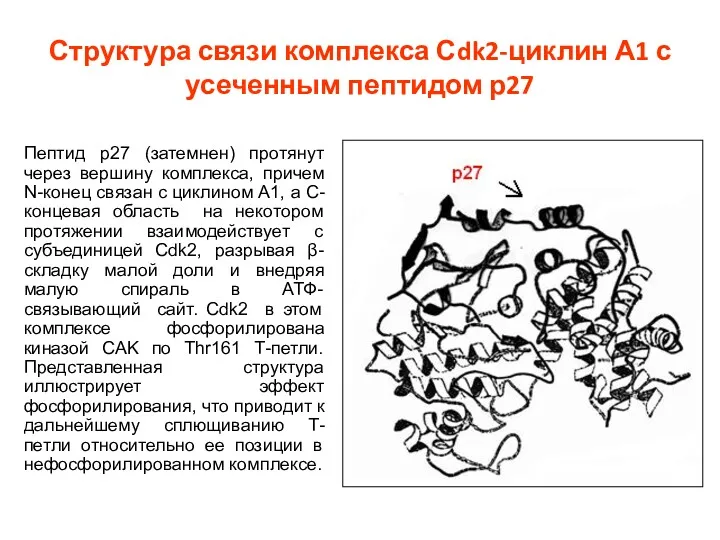
Структура связи комплекса Сdk2-циклин А1 с усеченным пептидом р27
Пептид р27
Структура связи комплекса Сdk2-циклин А1 с усеченным пептидом р27
Пептид р27
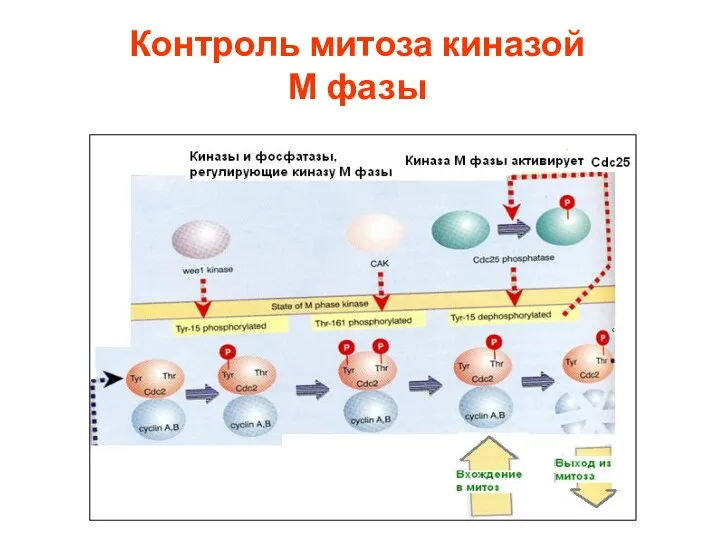
Контроль митоза киназой
М фазы
Контроль митоза киназой
М фазы
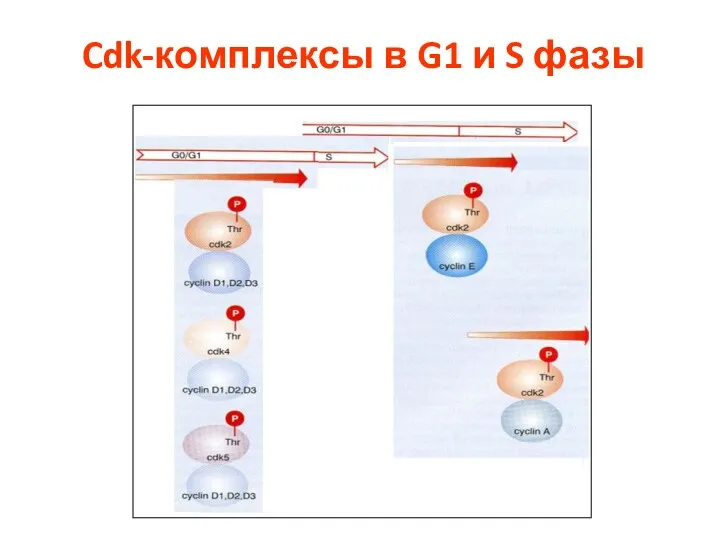
Cdk-комплексы в G1 и S фазы
Cdk-комплексы в G1 и S фазы

Регуляция циклинов
- Транскрипция (E2F стимулирует ЦА и Е в S фазе)
Регуляция циклинов
- Транскрипция (E2F стимулирует ЦА и Е в S фазе)
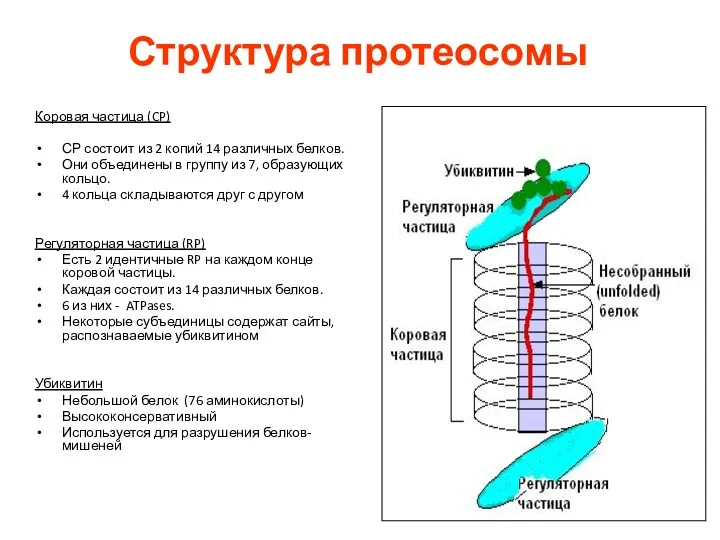
Структура протеосомы
Коровая частица (CP)
СР состоит из 2 копий 14 различных белков.
Они
Структура протеосомы
Коровая частица (CP)
СР состоит из 2 копий 14 различных белков.
Они
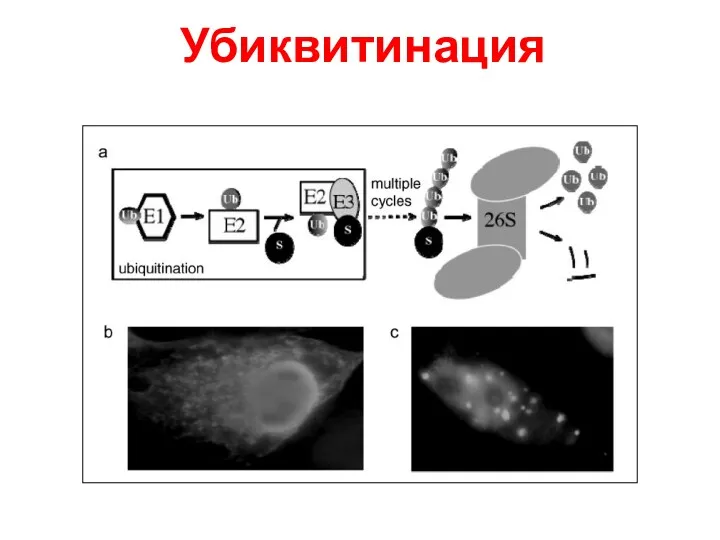
Убиквитинация
Убиквитинация
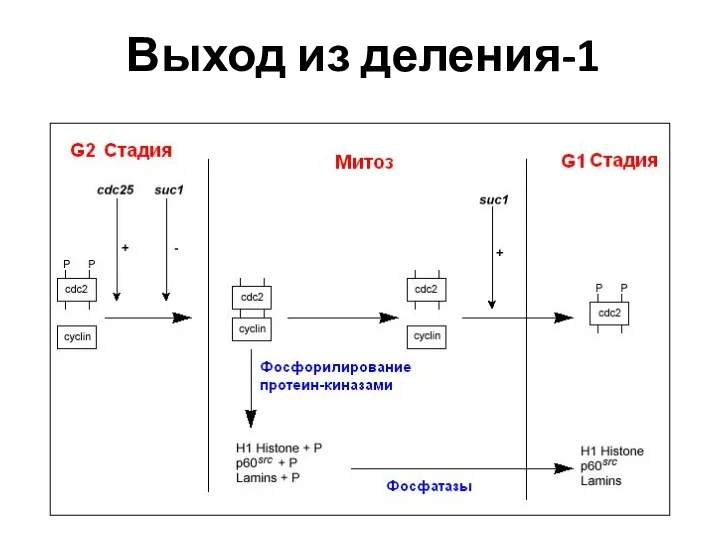
Выход из деления-1
Выход из деления-1
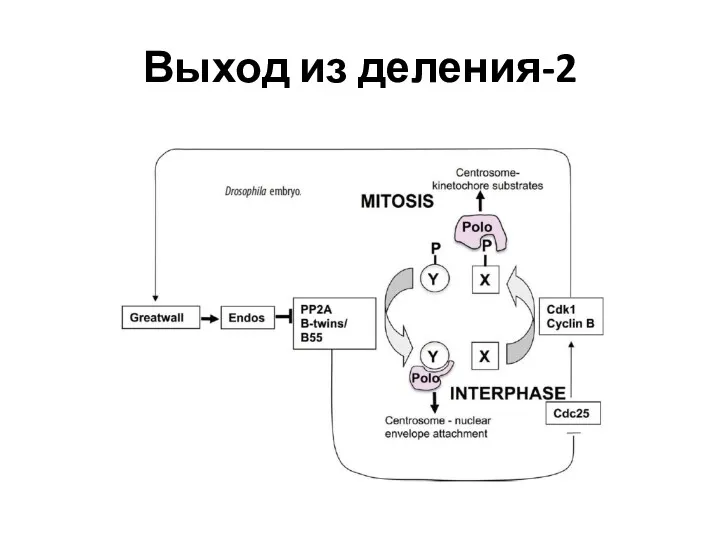
Выход из деления-2
Выход из деления-2

Деструкция митотических циклинов
Циклосома АРС (anaphase promoting complex) – метафаза-анафаза (Cdc20)
Деструкция митотических циклинов
Циклосома АРС (anaphase promoting complex) – метафаза-анафаза (Cdc20)
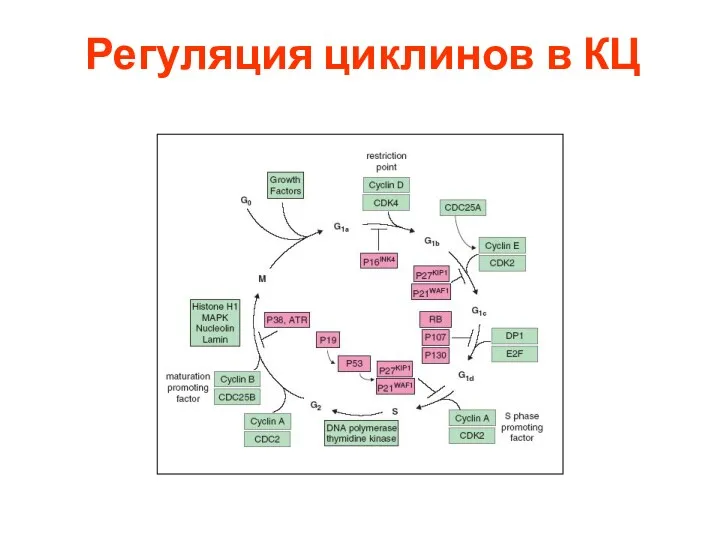
Регуляция циклинов в КЦ
Регуляция циклинов в КЦ

Нобелевская премия по физиологии и медицине за 2001 г.
Присуждена американцу Леланду
Нобелевская премия по физиологии и медицине за 2001 г.
Присуждена американцу Леланду
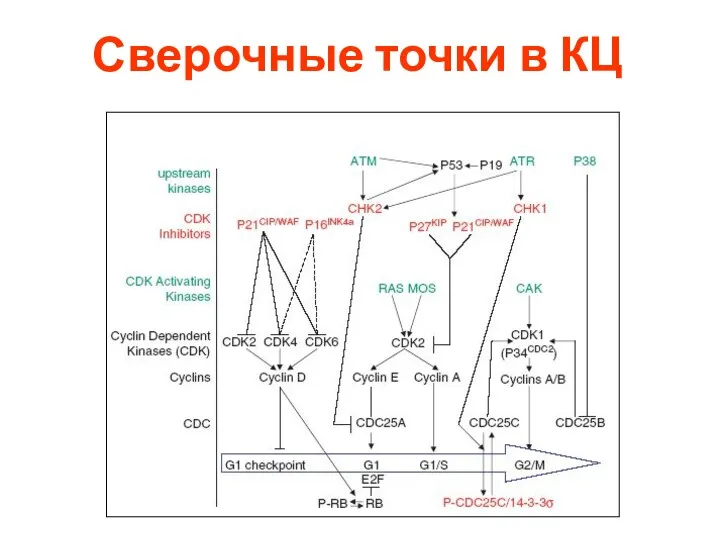
Сверочные точки в КЦ
Сверочные точки в КЦ
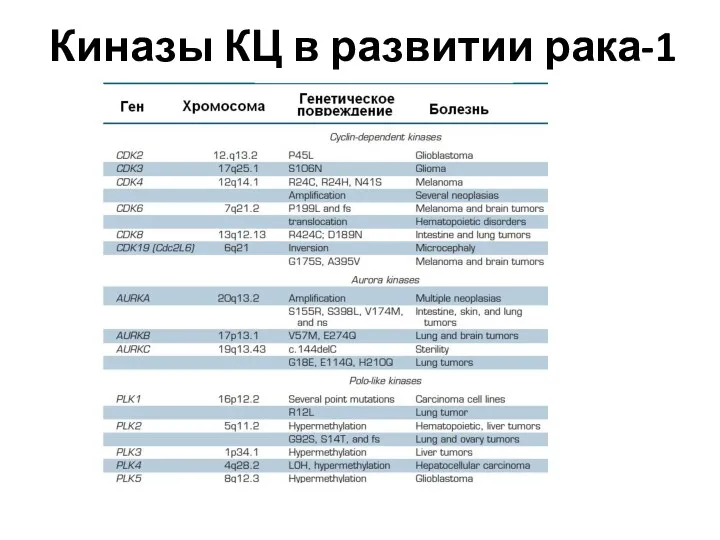
Киназы КЦ в развитии рака-1
Киназы КЦ в развитии рака-1
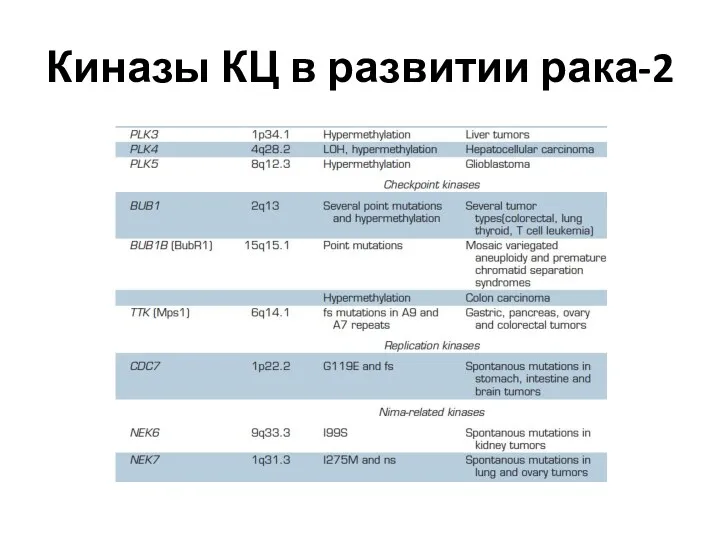
Киназы КЦ в развитии рака-2
Киназы КЦ в развитии рака-2
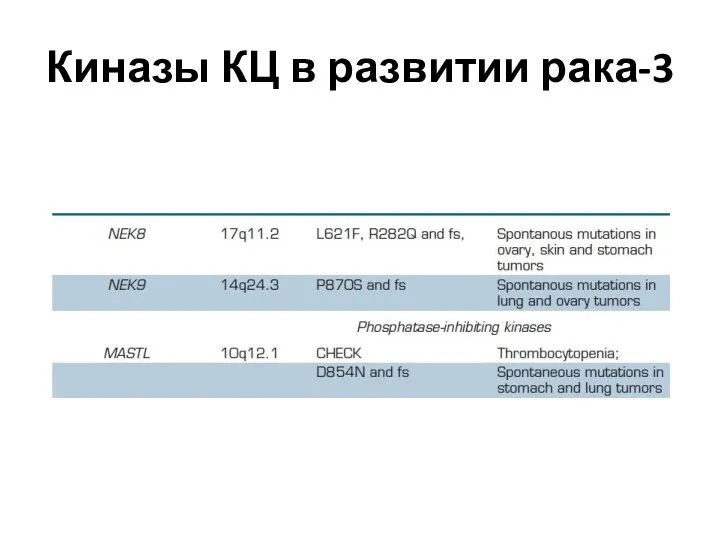
Киназы КЦ в развитии рака-3
Киназы КЦ в развитии рака-3
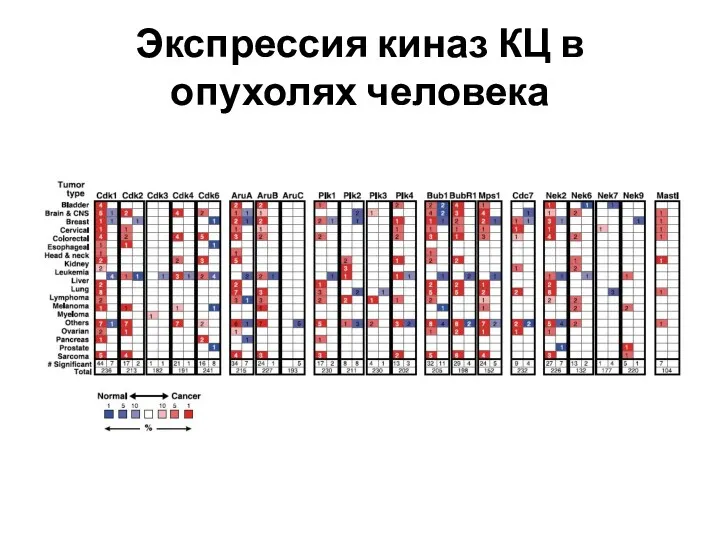
Экспрессия киназ КЦ в опухолях человека
Экспрессия киназ КЦ в опухолях человека
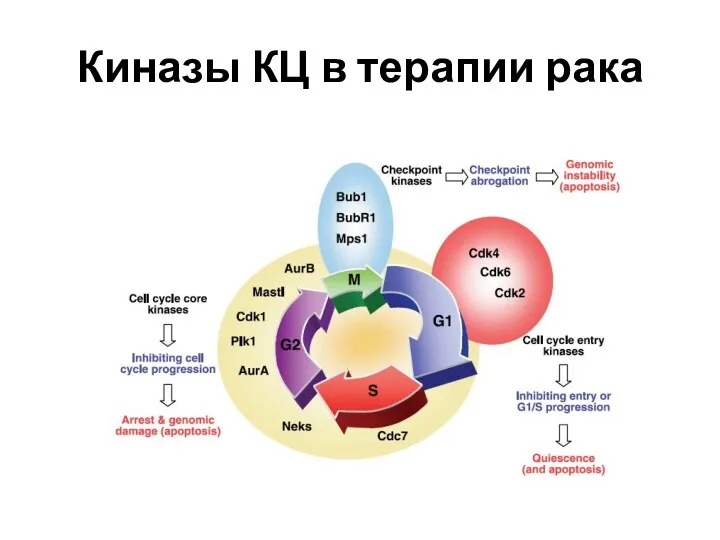
Киназы КЦ в терапии рака
Киназы КЦ в терапии рака

2. Раковые супрессорные гены или онкосупрессоры
RB
р53 (ТР53)
Развитие рака происходит
2. Раковые супрессорные гены или онкосупрессоры
RB
р53 (ТР53)
Развитие рака происходит

RB Белок
RB Белок
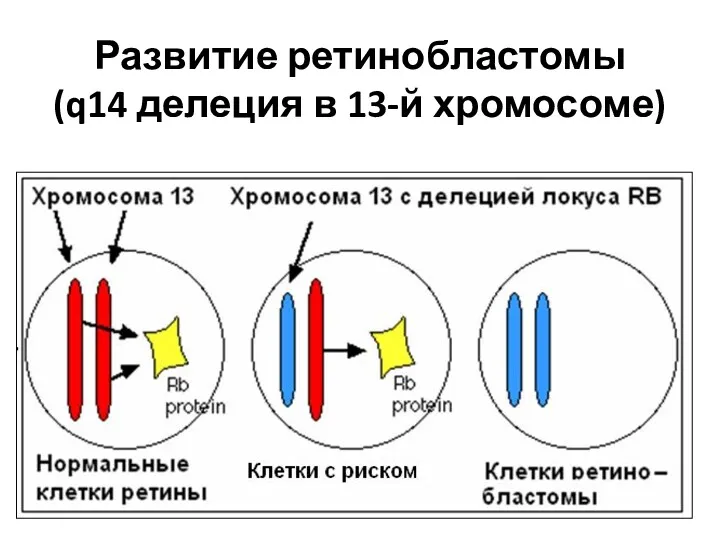
Развитие ретинобластомы
(q14 делеция в 13-й хромосоме)
Развитие ретинобластомы
(q14 делеция в 13-й хромосоме)
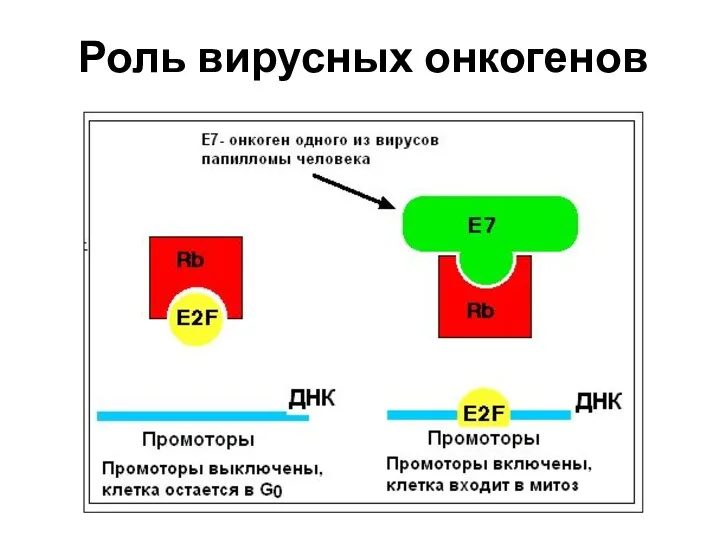
Роль вирусных онкогенов
Роль вирусных онкогенов
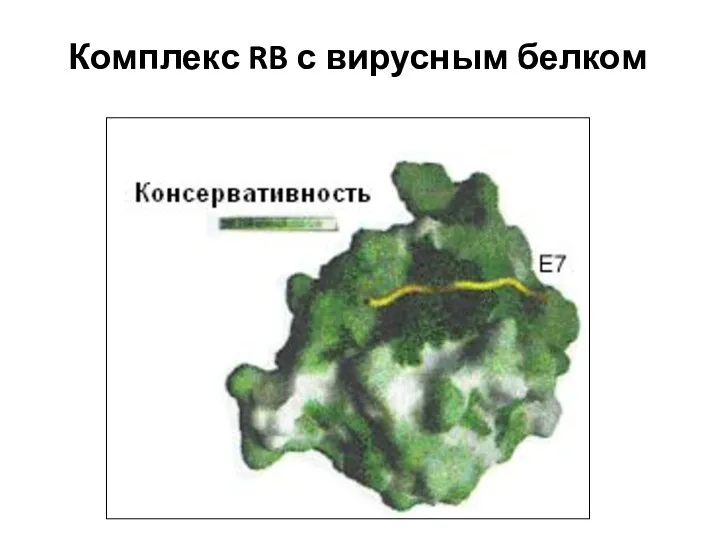
Комплекс RB с вирусным белком
Комплекс RB с вирусным белком
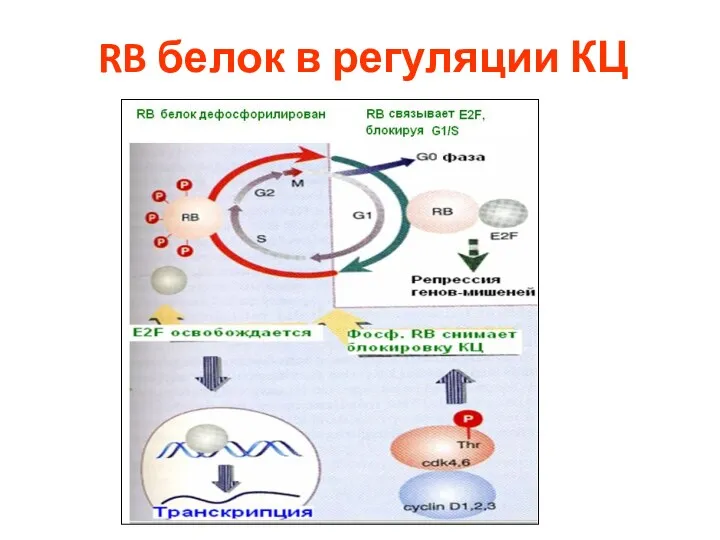
RB белок в регуляции КЦ
RB белок в регуляции КЦ
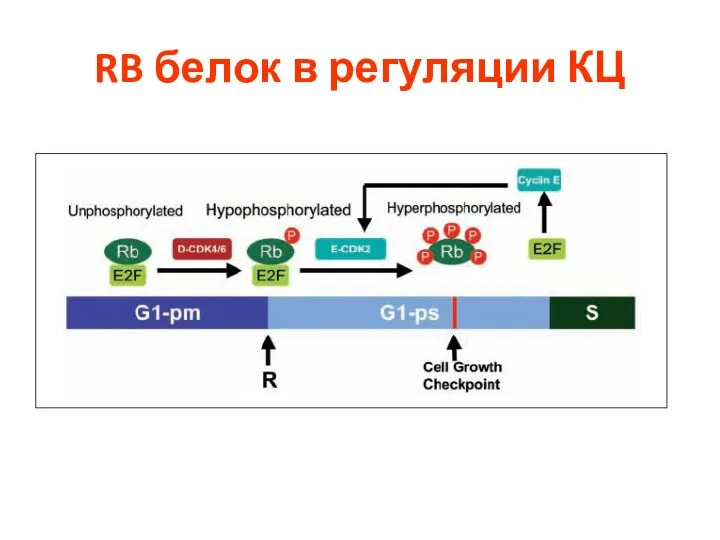
RB белок в регуляции КЦ
RB белок в регуляции КЦ
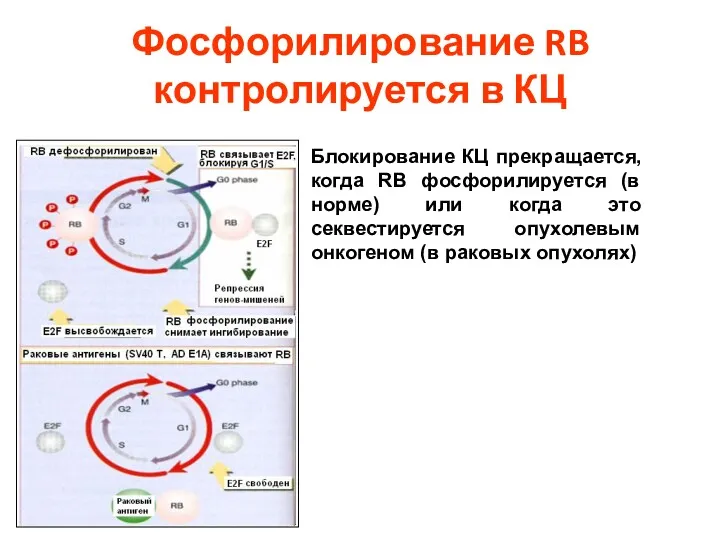
Фосфорилирование RB контролируется в КЦ
Блокирование КЦ прекращается, когда RB фосфорилируется (в
Фосфорилирование RB контролируется в КЦ
Блокирование КЦ прекращается, когда RB фосфорилируется (в
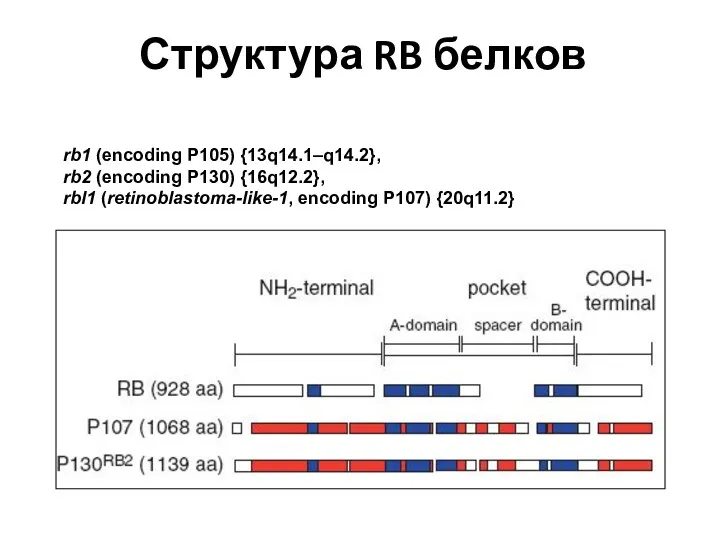
Структура RB белков
rb1 (encoding P105) {13q14.1–q14.2},
rb2 (encoding P130) {16q12.2},
rbl1
Структура RB белков
rb1 (encoding P105) {13q14.1–q14.2},
rb2 (encoding P130) {16q12.2},
rbl1
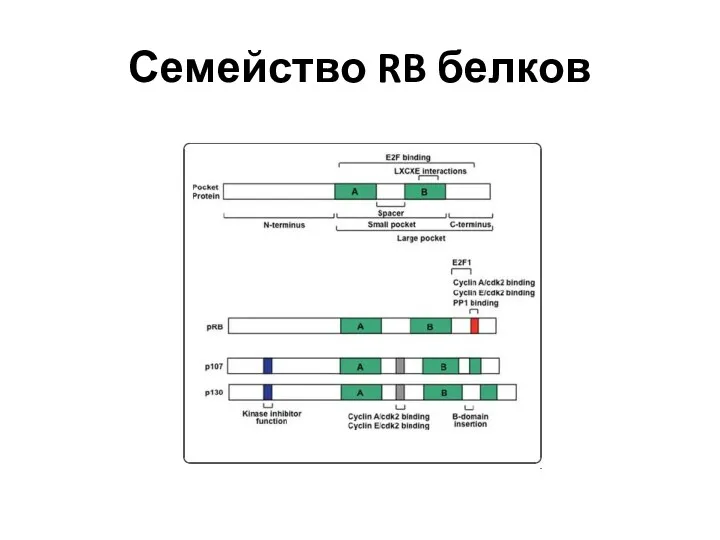
Семейство RB белков
Семейство RB белков
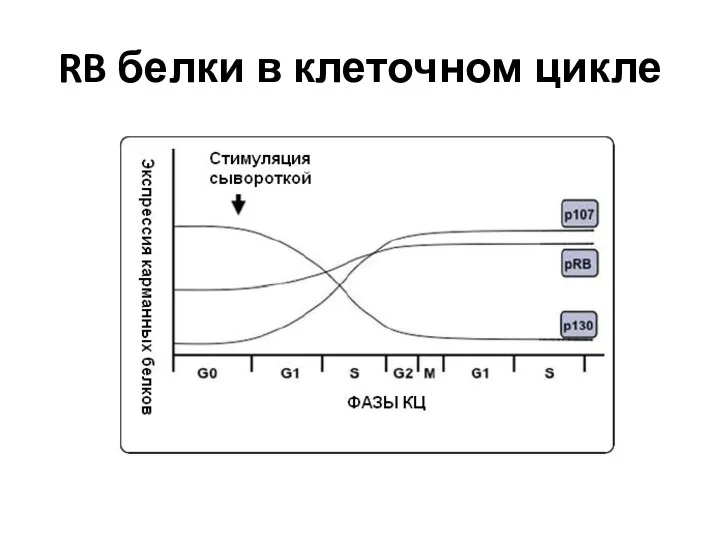
RB белки в клеточном цикле
RB белки в клеточном цикле
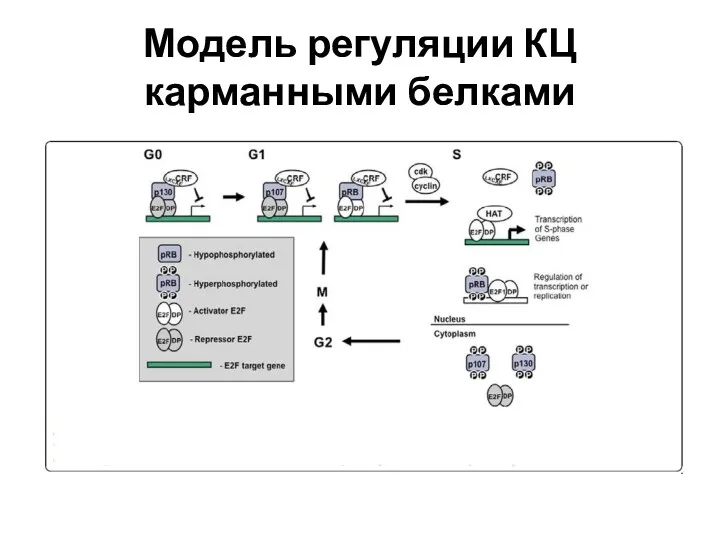
Модель регуляции КЦ карманными белками
Модель регуляции КЦ карманными белками
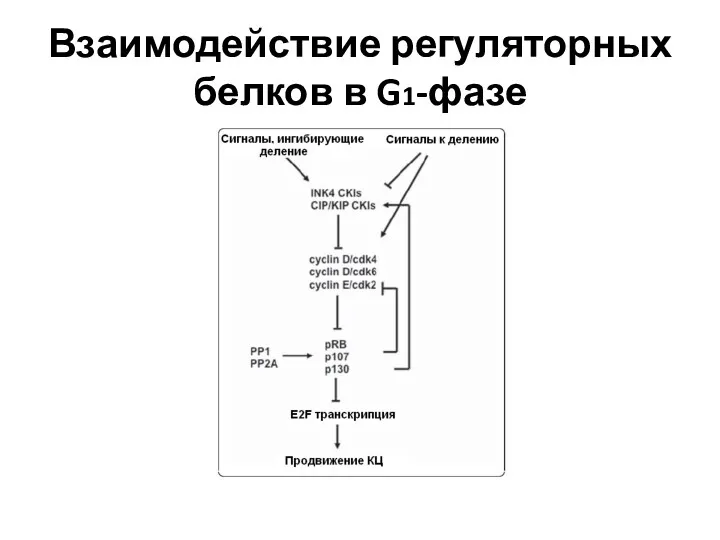
Взаимодействие регуляторных белков в G1-фазе
Взаимодействие регуляторных белков в G1-фазе
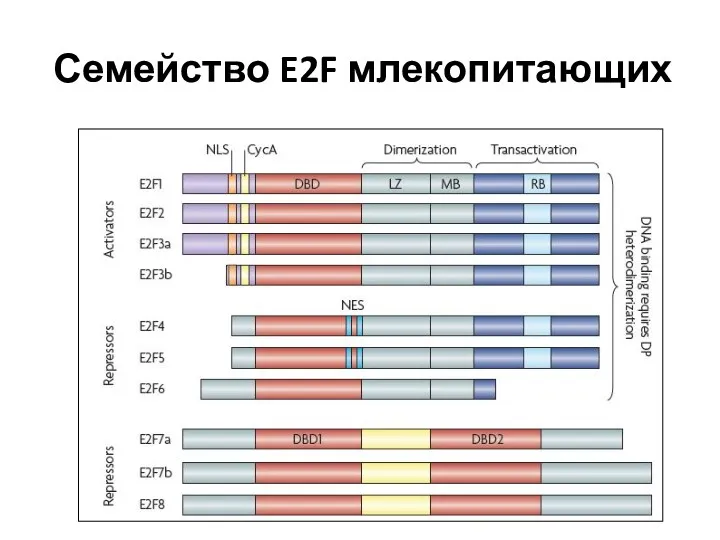
Семейство E2F млекопитающих
Семейство E2F млекопитающих
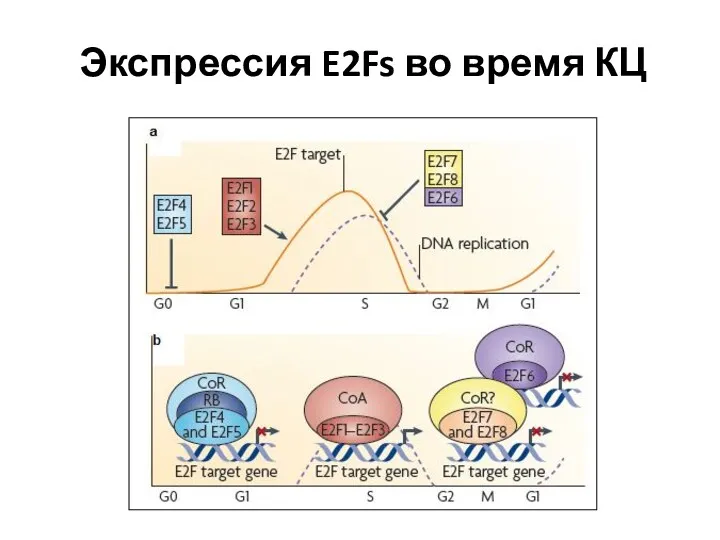
Экспрессия E2Fs во время КЦ
Экспрессия E2Fs во время КЦ
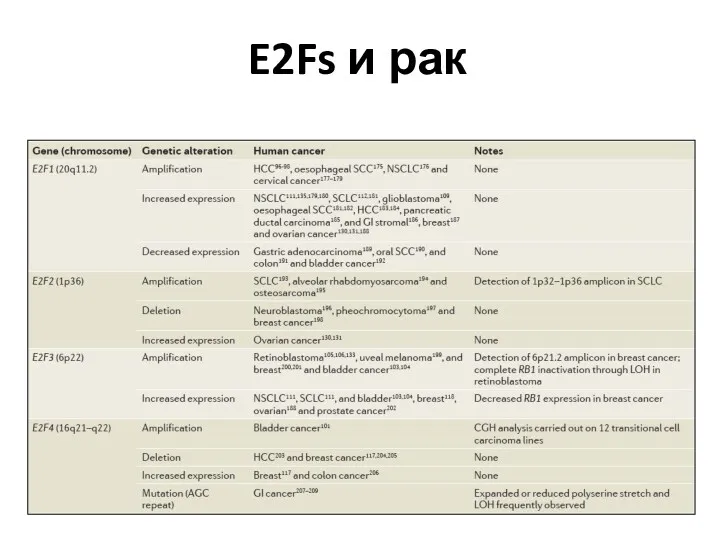
E2Fs и рак
E2Fs и рак
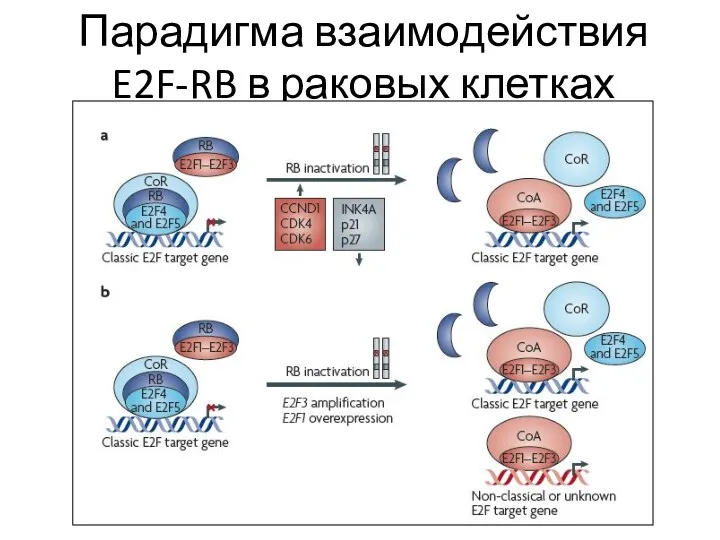
Парадигма взаимодействия E2F-RB в раковых клетках
Парадигма взаимодействия E2F-RB в раковых клетках

В ядре непролиферирующих клеток остается RB гипофосфорилированным и формирует белковый комплекс
В ядре непролиферирующих клеток остается RB гипофосфорилированным и формирует белковый комплекс
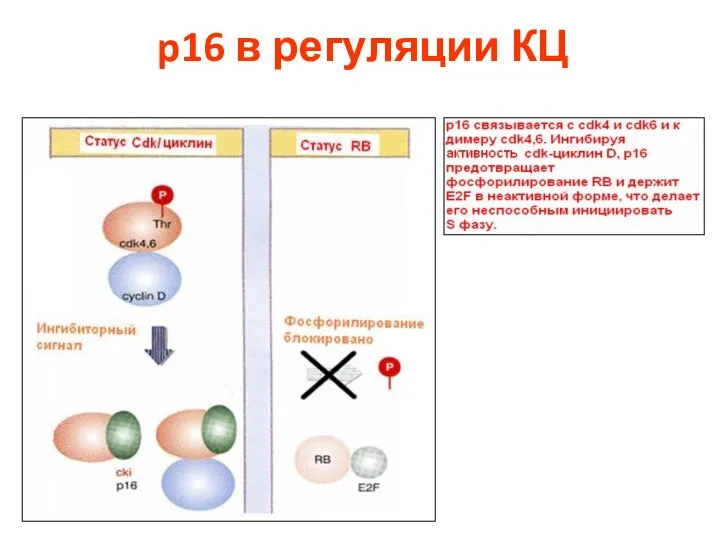
p16 в регуляции КЦ
p16 в регуляции КЦ
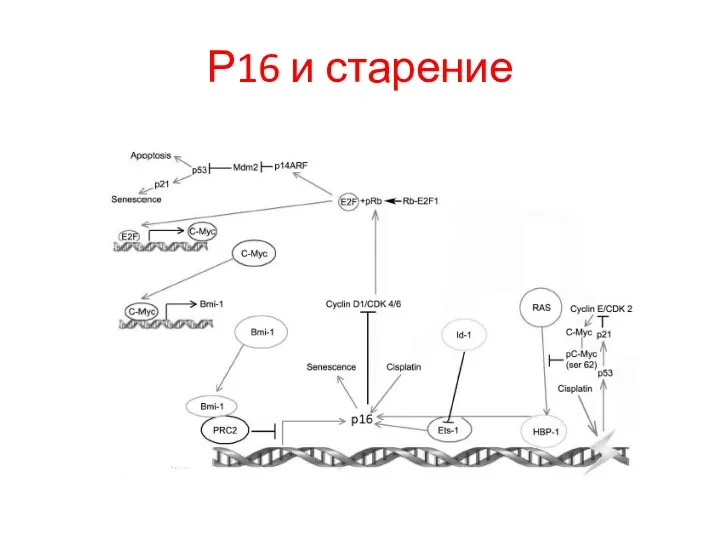
Р16 и старение
Р16 и старение
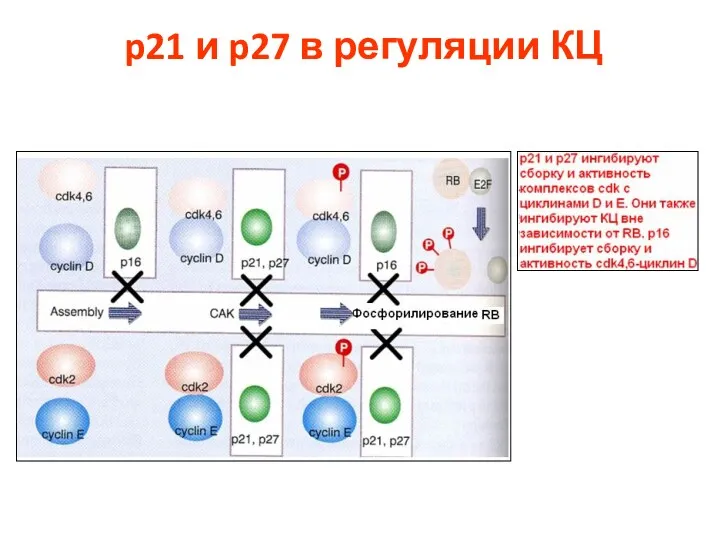
p21 и p27 в регуляции КЦ
p21 и p27 в регуляции КЦ

Белок p53 – ключевой онкосупрессор. Роль в онкологии
Белок p53 – ключевой онкосупрессор. Роль в онкологии

Структура и функции р53
- ДНК-связывающий белок (впервые описан в 1979 г.)
-
Структура и функции р53
- ДНК-связывающий белок (впервые описан в 1979 г.)
-
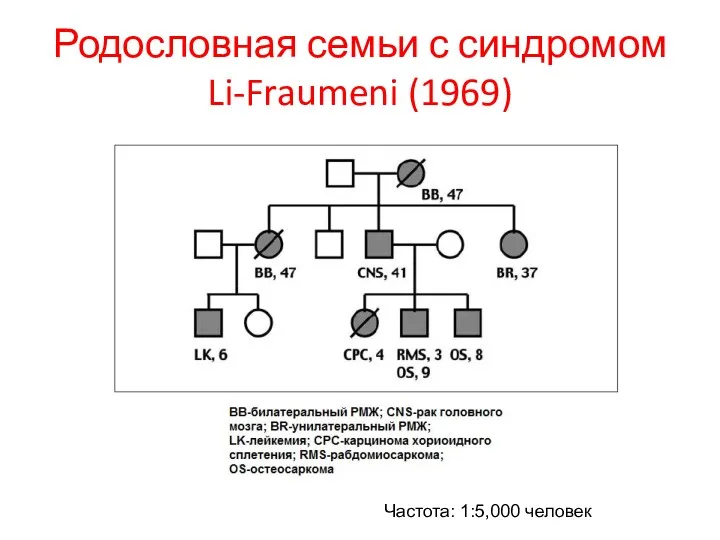
Родословная семьи с синдромом Li-Fraumeni (1969)
Частота: 1:5,000 человек
Родословная семьи с синдромом Li-Fraumeni (1969)
Частота: 1:5,000 человек

Последовательности, узнаваемые р53
PuPuPuC(A/T)(T/A)GPyPyPy
Последовательности, узнаваемые р53
PuPuPuC(A/T)(T/A)GPyPyPy
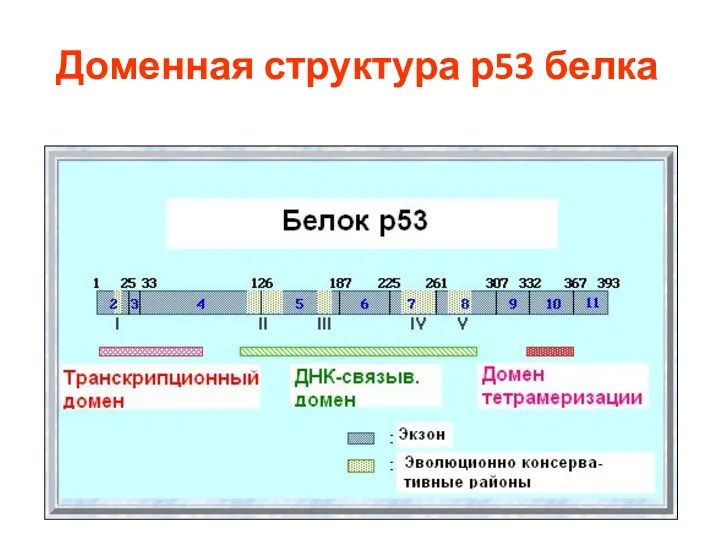
Доменная структура р53 белка
Доменная структура р53 белка
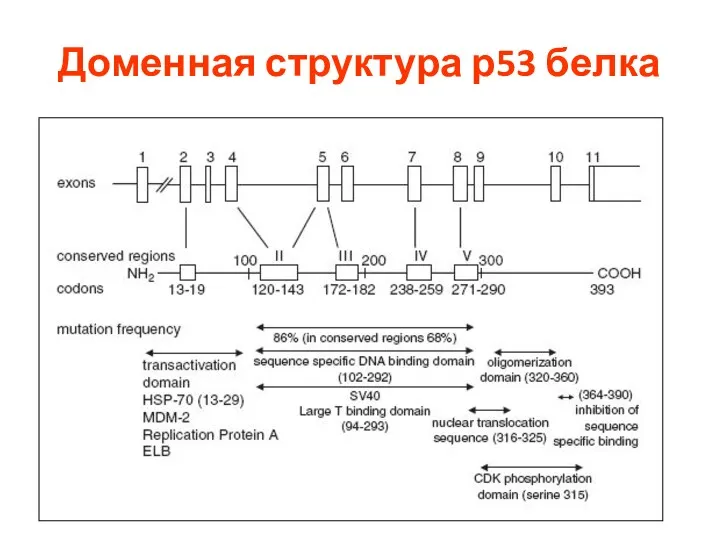
Доменная структура р53 белка
Доменная структура р53 белка
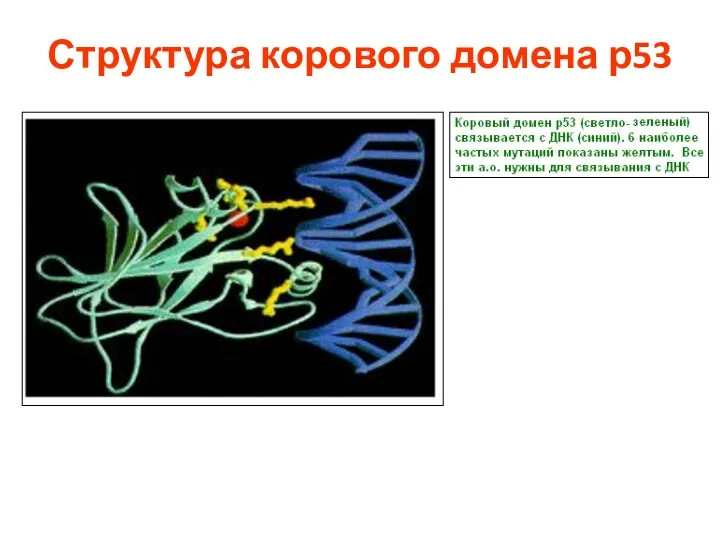
Структура корового домена р53
Структура корового домена р53
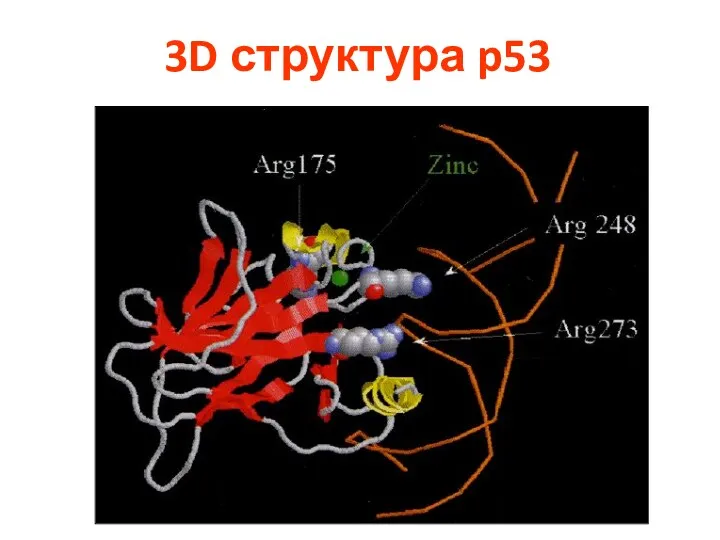
3D структура p53
3D структура p53
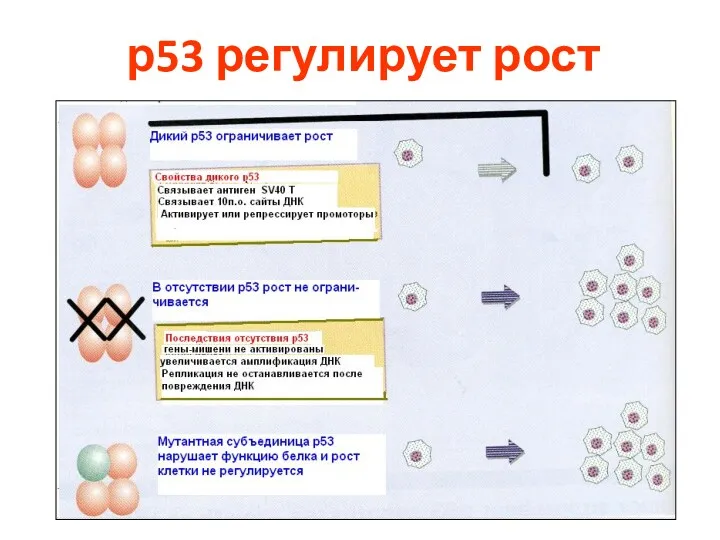
р53 регулирует рост
р53 регулирует рост
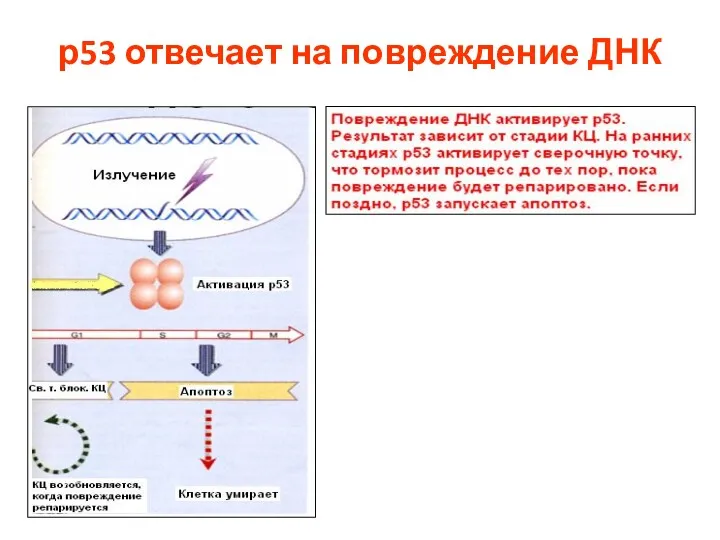
р53 отвечает на повреждение ДНК
р53 отвечает на повреждение ДНК
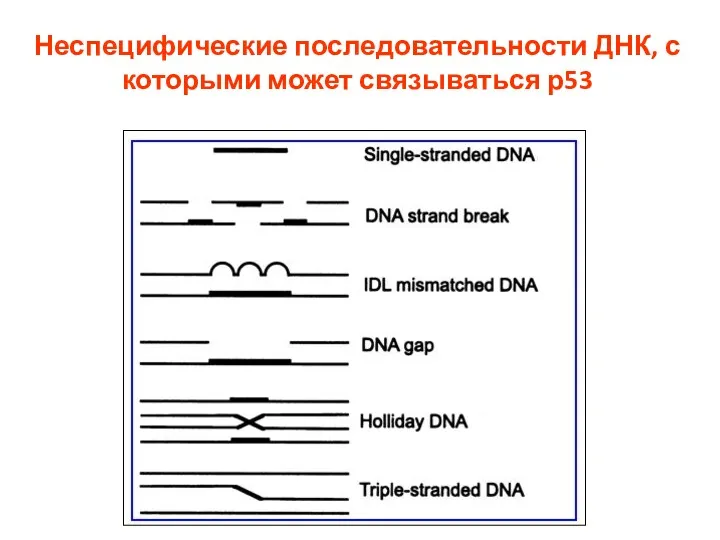
Неспецифические последовательности ДНК, с которыми может связываться р53
Неспецифические последовательности ДНК, с которыми может связываться р53
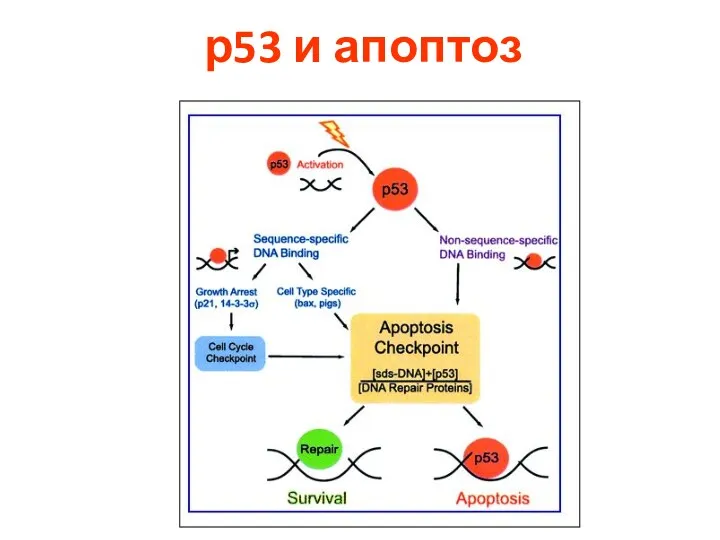
р53 и апоптоз
р53 и апоптоз
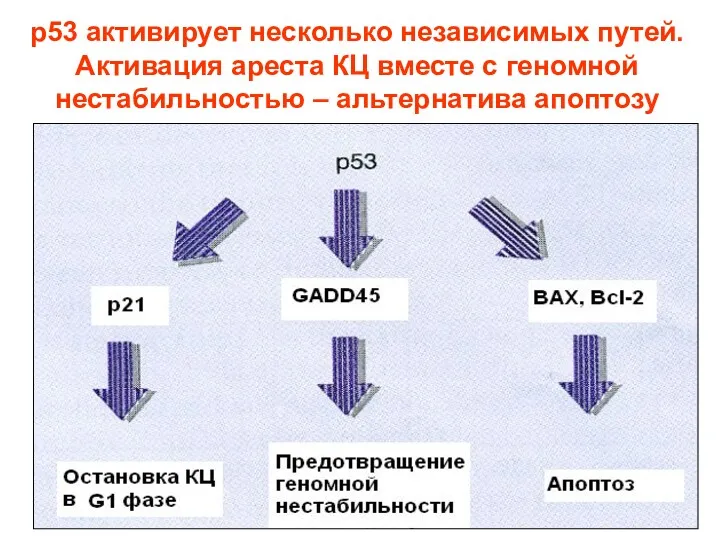
p53 активирует несколько независимых путей. Активация ареста КЦ вместе с геномной
p53 активирует несколько независимых путей. Активация ареста КЦ вместе с геномной

Регуляция активности р53
1. В норме его экспрессия невелика и контролируется убиквинированием
Регуляция активности р53
1. В норме его экспрессия невелика и контролируется убиквинированием
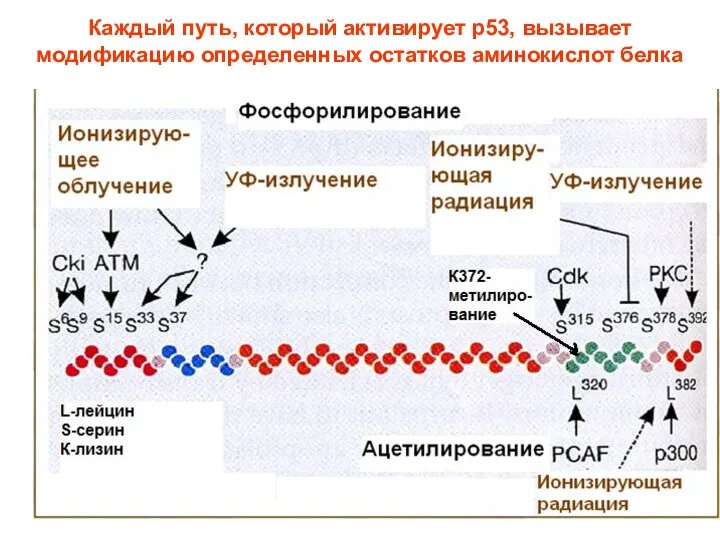
Каждый путь, который активирует р53, вызывает модификацию определенных остатков аминокислот белка
Каждый путь, который активирует р53, вызывает модификацию определенных остатков аминокислот белка
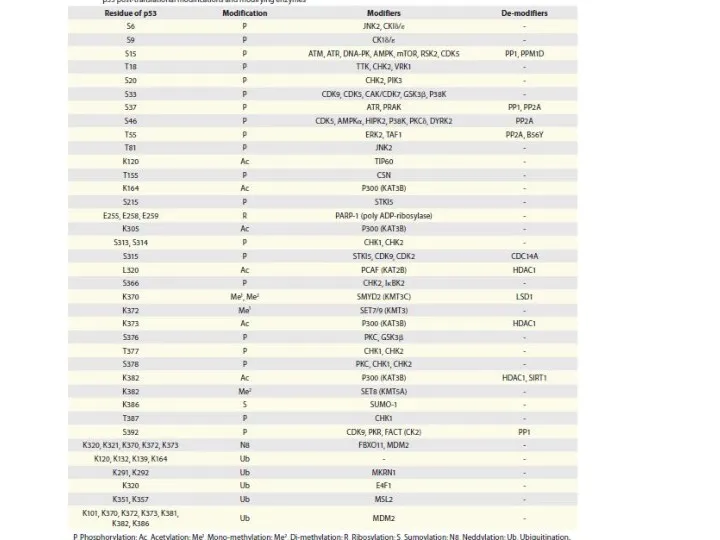

MDM2 в регуляции р53
1991 г. - впервые показано его взаимодействие с
MDM2 в регуляции р53
1991 г. - впервые показано его взаимодействие с

Структура белка Mdm2
Структура белка Mdm2
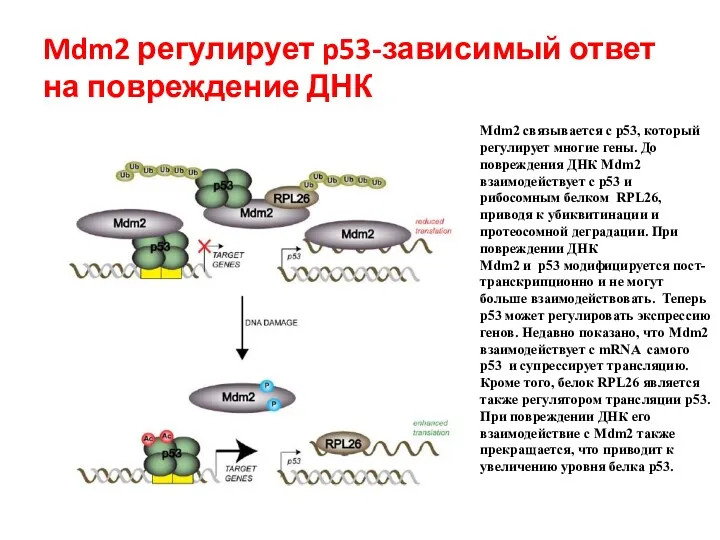
Mdm2 регулирует p53-зависимый ответ на повреждение ДНК
Mdm2 связывается с p53, который
Mdm2 регулирует p53-зависимый ответ на повреждение ДНК
Mdm2 связывается с p53, который
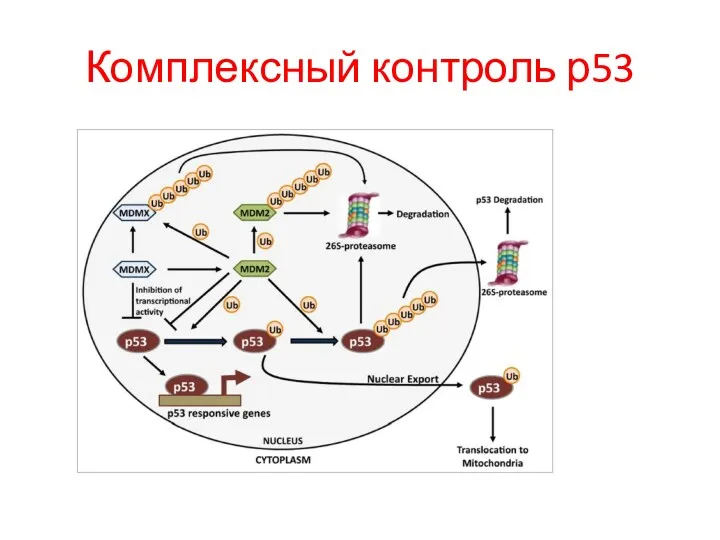
Комплексный контроль р53
Комплексный контроль р53
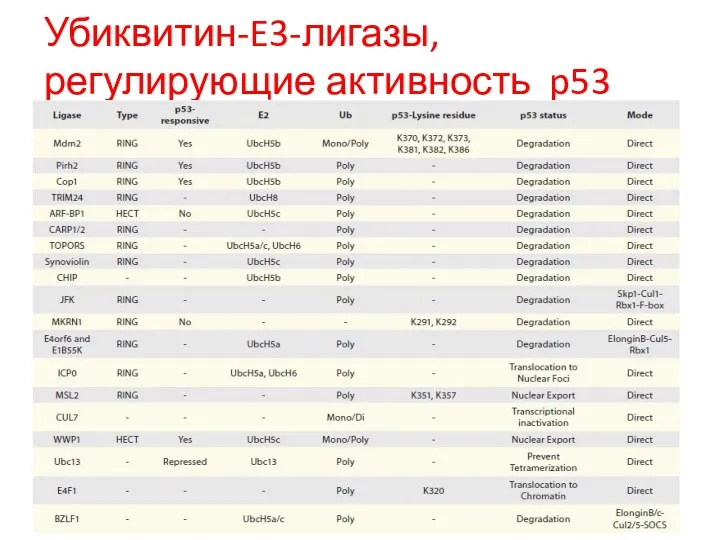
Убиквитин-E3-лигазы, регулирующие активность p53
Убиквитин-E3-лигазы, регулирующие активность p53
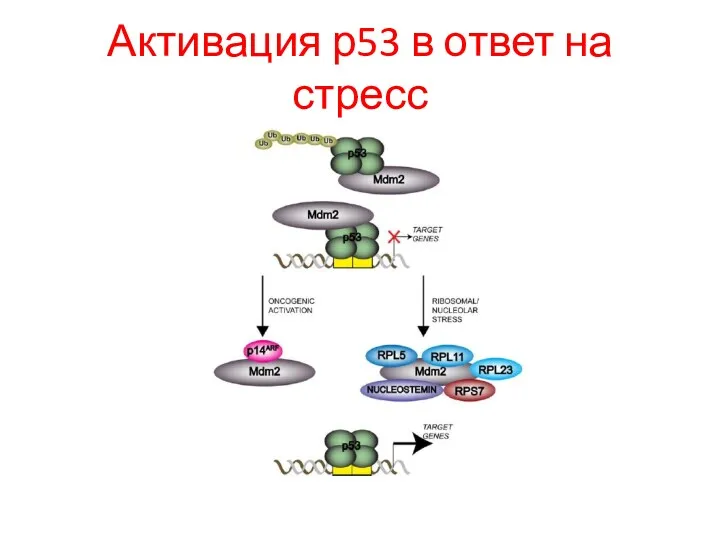
Активация р53 в ответ на стресс
Активация р53 в ответ на стресс
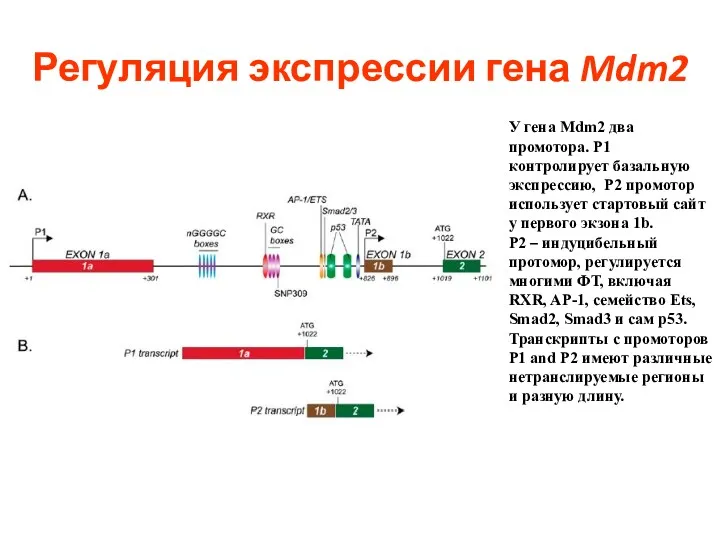
Регуляция экспрессии гена Mdm2
У гена Mdm2 два промотора. P1 контролирует базальную
Регуляция экспрессии гена Mdm2
У гена Mdm2 два промотора. P1 контролирует базальную
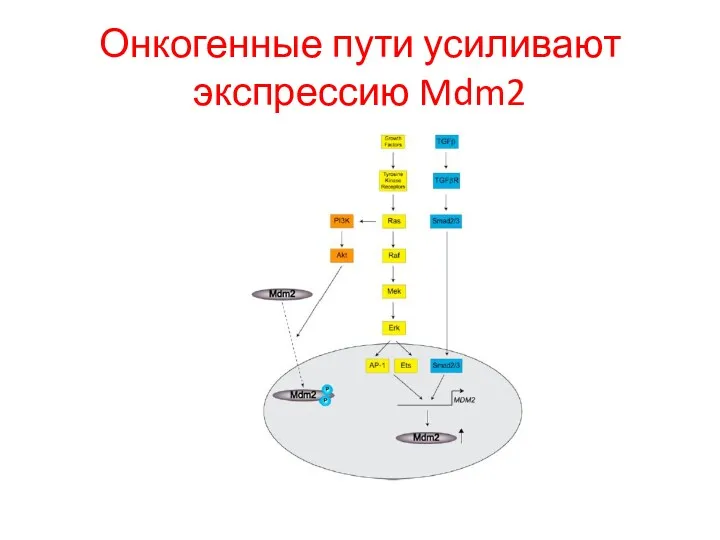
Онкогенные пути усиливают экспрессию Mdm2
Онкогенные пути усиливают экспрессию Mdm2
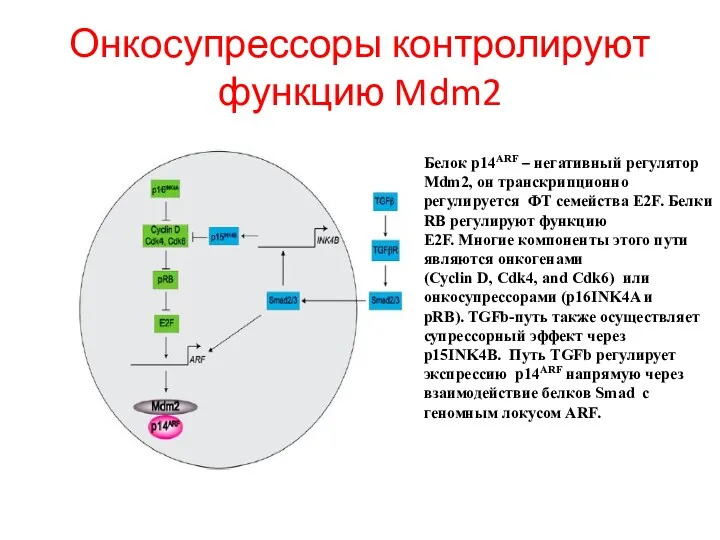
Онкосупрессоры контролируют функцию Mdm2
Белок p14ARF – негативный регулятор Mdm2, он транскрипционно
Онкосупрессоры контролируют функцию Mdm2
Белок p14ARF – негативный регулятор Mdm2, он транскрипционно
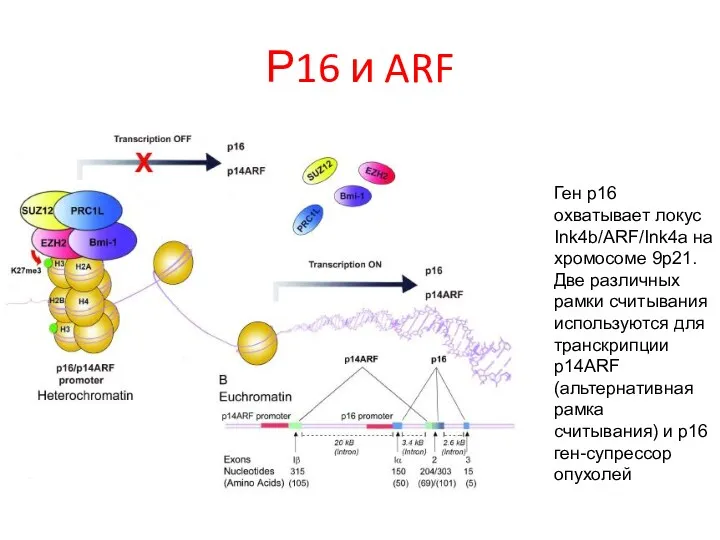
Р16 и ARF
Ген p16 охватывает локус Ink4b/ARF/Ink4a на хромосоме 9p21. Две
Р16 и ARF
Ген p16 охватывает локус Ink4b/ARF/Ink4a на хромосоме 9p21. Две
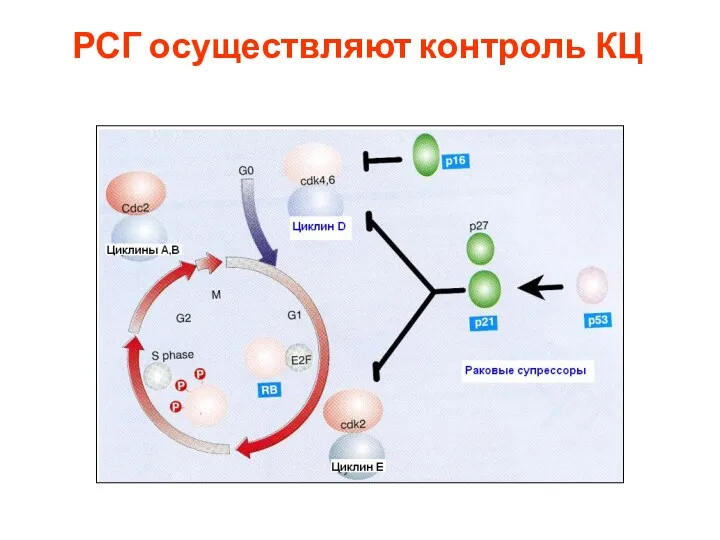
РСГ осуществляют контроль КЦ
РСГ осуществляют контроль КЦ
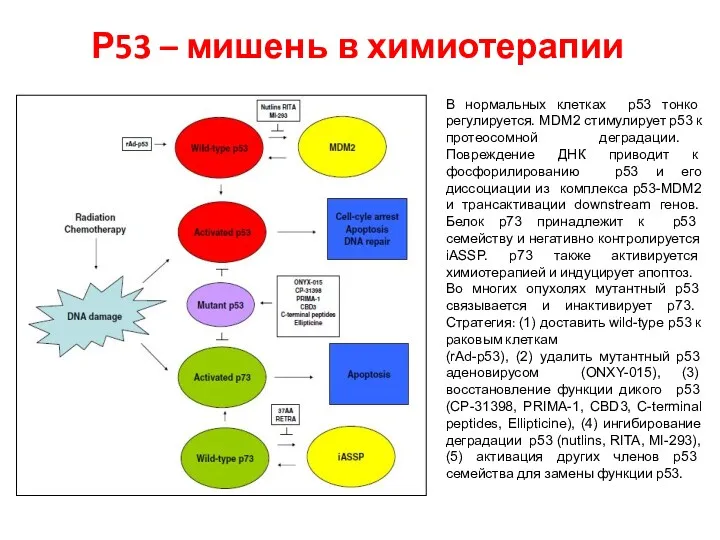
Р53 – мишень в химиотерапии
В нормальных клетках p53 тонко регулируется. MDM2
Р53 – мишень в химиотерапии
В нормальных клетках p53 тонко регулируется. MDM2
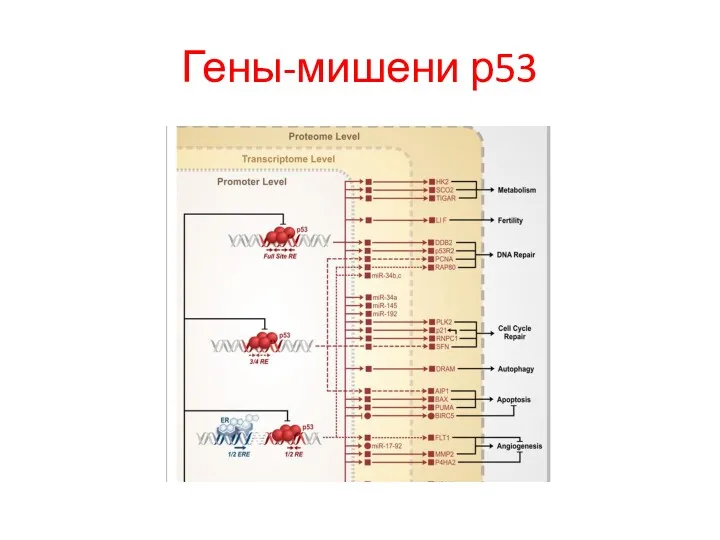
Гены-мишени р53
Гены-мишени р53

2011 г.: Согласно IARC, выявляется более, чем 26 000 соматических мутаций
2011 г.: Согласно IARC, выявляется более, чем 26 000 соматических мутаций
Наиболее часто встречаемые мутации р53
Наиболее часто встречаемые мутации р53

Относительная частота герминальных мутаций в p53
Относительная частота герминальных мутаций в p53
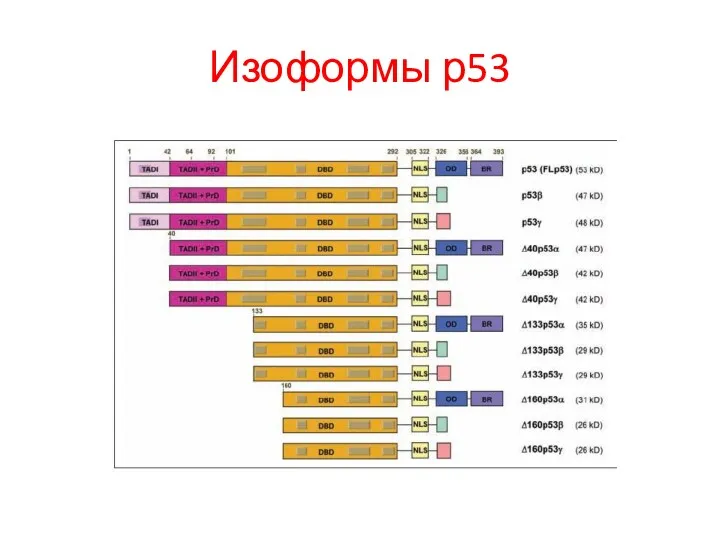
Изоформы р53
Изоформы р53
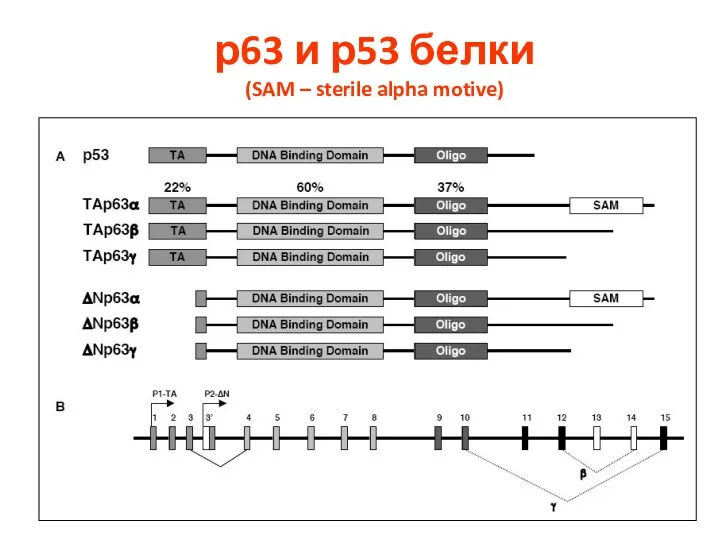
р63 и р53 белки
(SAM – sterile alpha motive)
р63 и р53 белки
(SAM – sterile alpha motive)

Экспрессия р63 в опухолях
Экспрессия р63 в опухолях

Восстановление активности р53
Введение в опухоль дикого p53 с помощью репликативно-дефектного аденовирусного
Восстановление активности р53
Введение в опухоль дикого p53 с помощью репликативно-дефектного аденовирусного

Маркеры РМЖ на 17-хромосоме
Wei Zhang and Yingyan Yu «The Important Molecular
Маркеры РМЖ на 17-хромосоме
Wei Zhang and Yingyan Yu «The Important Molecular

Функции маркеров
Функции маркеров
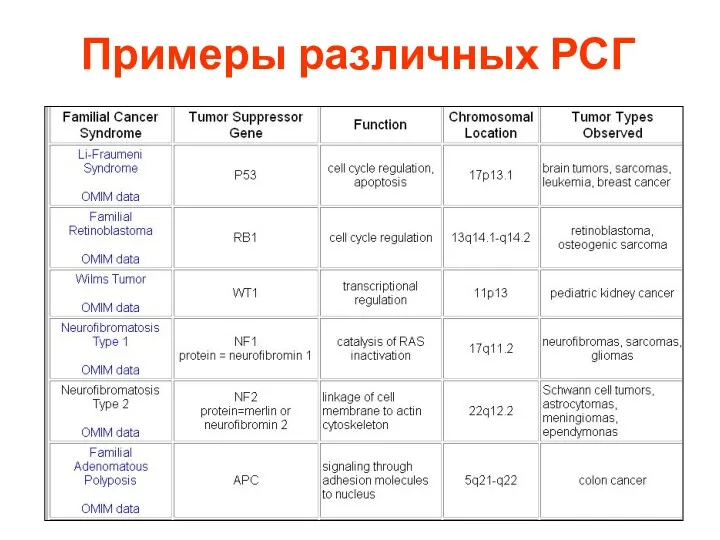
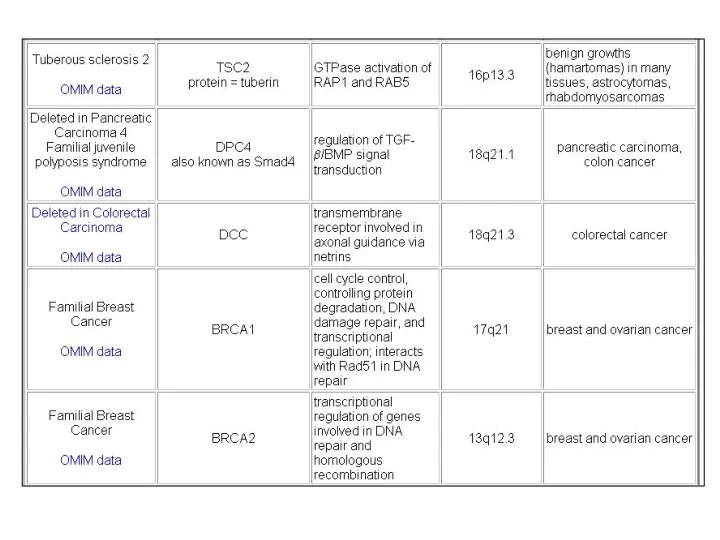
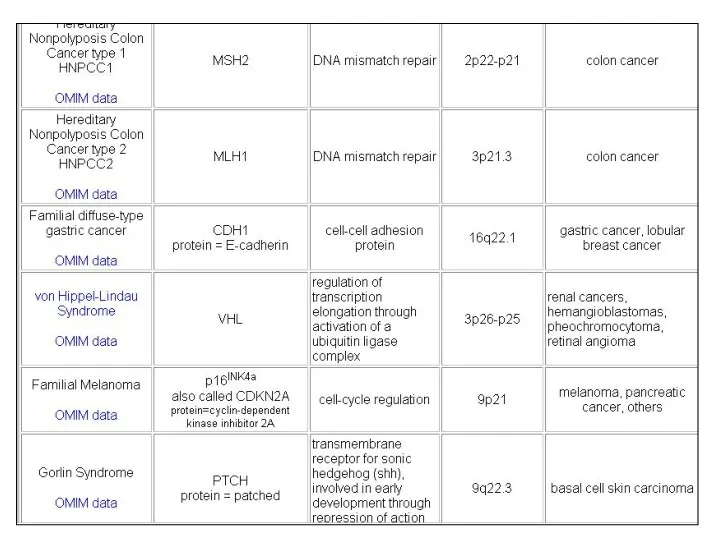
Примеры различных РСГ
Примеры различных РСГ

Примеры нокаутных мышей с инактивированными РСГ или ПО
Примеры нокаутных мышей с инактивированными РСГ или ПО
 Методы экологических исследований. Лекция 6
Методы экологических исследований. Лекция 6 Скелет. Строение, состав и соединение костей
Скелет. Строение, состав и соединение костей День птиц
День птиц Комнатные растения в интерьере
Комнатные растения в интерьере Царство Phyta (растения)
Царство Phyta (растения) Тип: моллюски. Класс: двустворчатые. Устрица
Тип: моллюски. Класс: двустворчатые. Устрица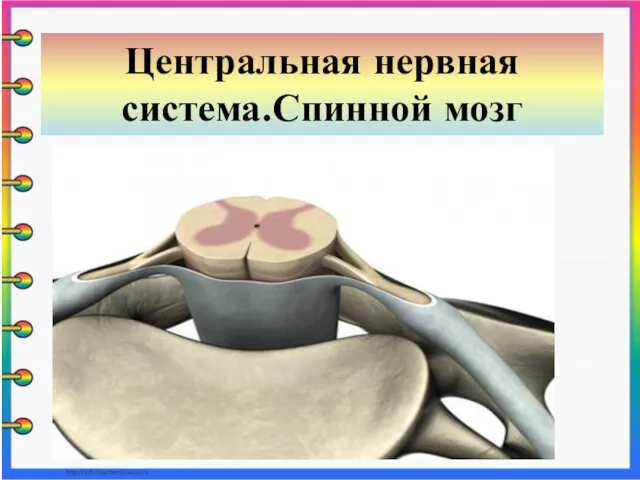 Строение и функции спинного мозга
Строение и функции спинного мозга Презентация к уроку биологии 8 кл.
Презентация к уроку биологии 8 кл. Самые необычные животные планеты
Самые необычные животные планеты Строение тела человека и его функциональные системы
Строение тела человека и его функциональные системы Разнообразие грибов. Значение грибов в природе и жизни человека
Разнообразие грибов. Значение грибов в природе и жизни человека Отряд Чешуекрылые, или Бабочки
Отряд Чешуекрылые, или Бабочки Общие вопросы анатомии и физиологии человека
Общие вопросы анатомии и физиологии человека Витамины. Определение, классификация
Витамины. Определение, классификация Растения-хищники
Растения-хищники Урок биологии в 10 классе Дигибридное скрещивание
Урок биологии в 10 классе Дигибридное скрещивание Бактерии. Строение клетки
Бактерии. Строение клетки Класс пресмыкающиеся
Класс пресмыкающиеся Нанобиотехнологии прокариот. Экпрессия генов
Нанобиотехнологии прокариот. Экпрессия генов Строение, размножение и развитие рыб
Строение, размножение и развитие рыб Эволюция опорно – двигательной системы у животных
Эволюция опорно – двигательной системы у животных Мифологические лекарственные растения
Мифологические лекарственные растения Презентация по теме Видоизменения корней. 6 класс. ФГОС ООО.
Презентация по теме Видоизменения корней. 6 класс. ФГОС ООО. Жердің даму тарихы
Жердің даму тарихы Строение цветка
Строение цветка Происхождение человека. Стадии антропосоциогенеза. Адаптация человека
Происхождение человека. Стадии антропосоциогенеза. Адаптация человека Экзотические птицы
Экзотические птицы Строение растительной клетки
Строение растительной клетки