- Репликация ДНК. (Лекция 3)

Содержание
- 2. Источники: Курсы лекций и презентации: «Молекулярная биология и генетика» «Молекулярная биология клетки» https://stepik.org https://lectoriy.mipt.ru/course/Biology-Molecular-14L Дейч К.О
- 3. «Кто» такие?
- 4. Антисмысловые РНК (AsRNA) (англ. Antisense RNA) одноцепочечные РНК, которые комплементарны мРНК. AsRNA могут подавлять и активировать
- 5. Псевдогены (англ. pseudogenes) Псевдогены — нефункциональные аналоги структурных генов, утратившие способность кодировать белок и не экспрессирующиеся
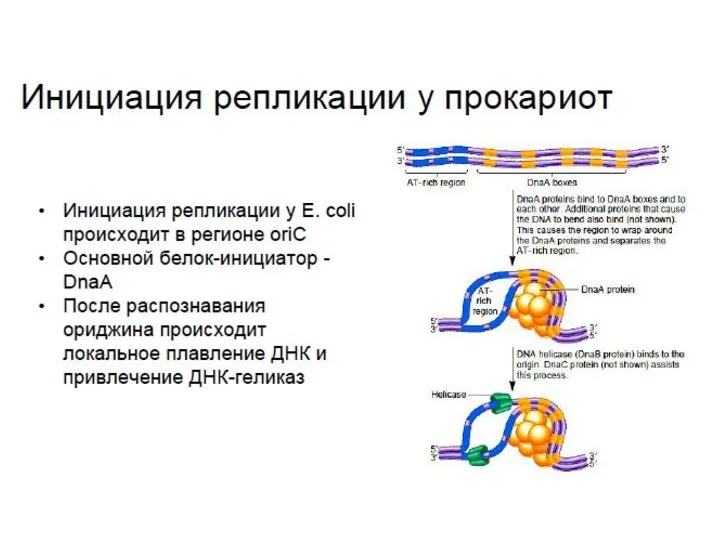
- 6. Лекция 3. Репликация ДНК как инициируется репликация у про- и эукариот, что такое ориджин репликации; почему
- 7. (1957 г.)
- 8. ДНК полимераза В 1956 г. Корнберг выделил из клеток бактерии Е. coli фермент ДНК-полимеразу (ДНК-полимераза I).
- 9. Репликация ДНК
- 10. Основные ферменты репликации ДНК-полимераза - фермент катализирующий полимеризацию дезоксирибонуклеотидов на матрице ДНК по принципу комплементарности ДНК-лигаза
- 12. Количество точек начала репликации в геноме Геном вироидов ( 1 РНК ) - по две точки
- 15. ? Как соотносится число ориджинов репликации в хромосомах бактерий и эукариот? У эукариот на одну хромосому
- 16. ? Выберите все верные утверждения, касающиеся инициации репликации у про- и эукариот: У эукариот инициация репликации
- 17. https://www.youtube.com/watch?v=BmAq-EolVCc
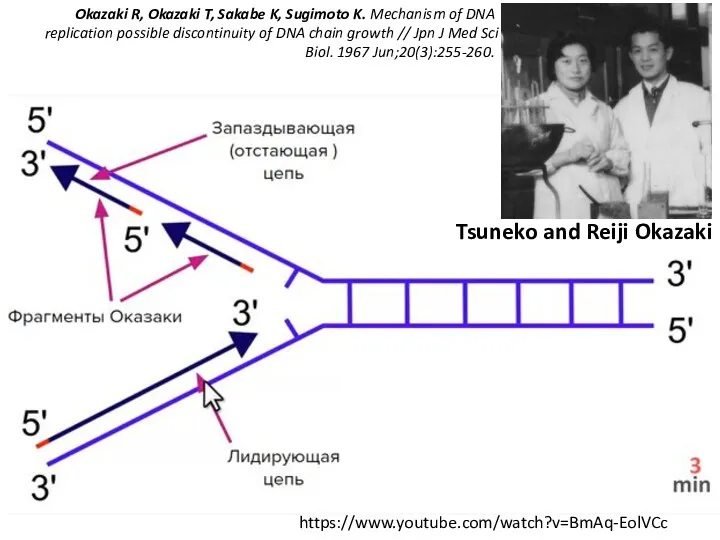
- 18. https://www.youtube.com/watch?v=BmAq-EolVCc Okazaki R, Okazaki T, Sakabe K, Sugimoto K. Mechanism of DNA replication possible discontinuity of
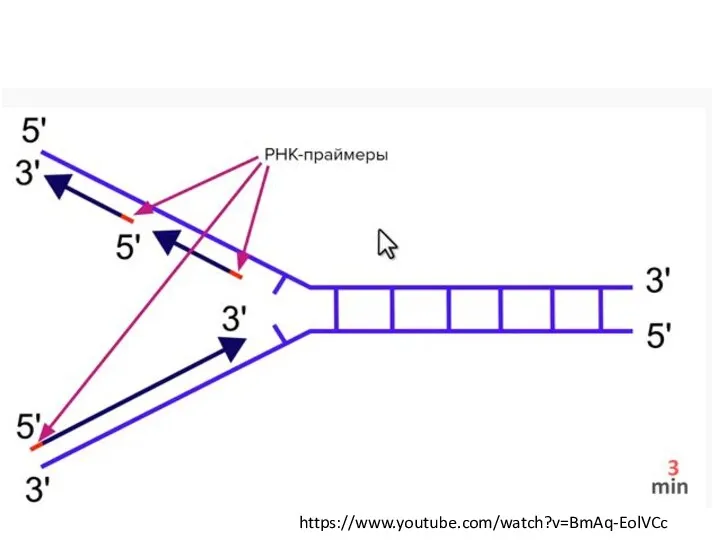
- 19. https://www.youtube.com/watch?v=BmAq-EolVCc
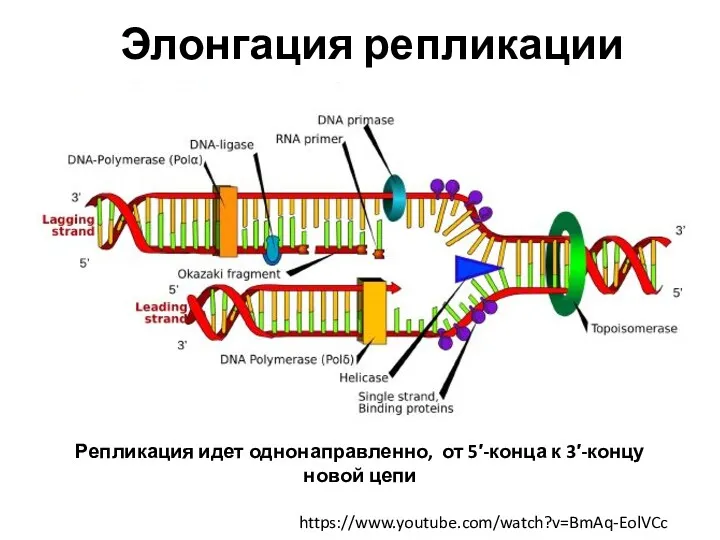
- 20. https://www.youtube.com/watch?v=BmAq-EolVCc Элонгация репликации Репликация идет однонаправленно, от 5′-конца к 3′-концу новой цепи ДНК-лигаза
- 21. ? Почему репликация на одной цепи ДНК идет непрерывно, а на другой — прерывисто? ДНК-полимераза способна
- 25. Репликация ДНК Свойства ДНК-полимераз эукариот
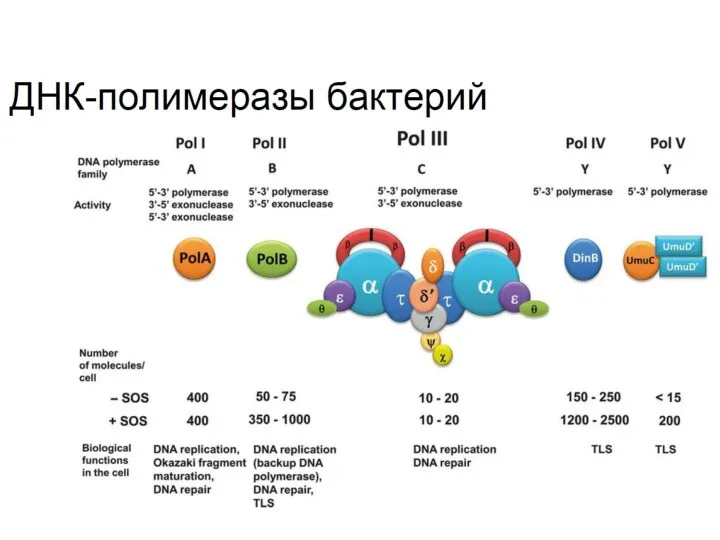
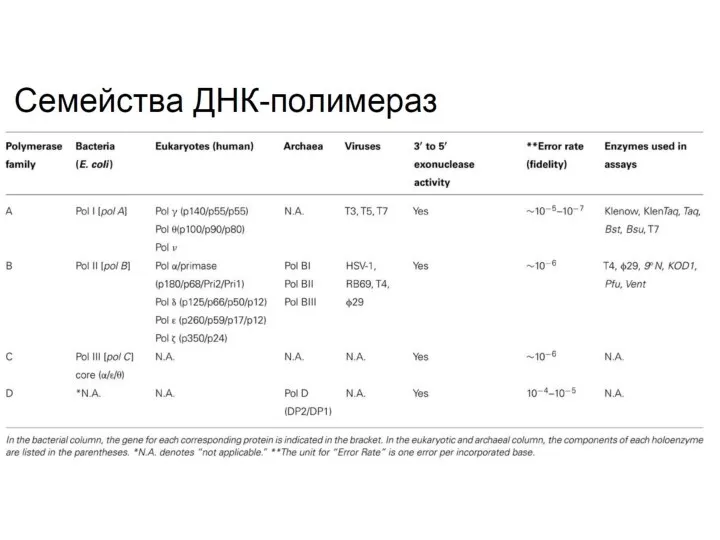
- 26. ? Отметьте верные утверждения, касающиеся ДНК-полимераз бактерий. Процессивность ДНК полимеразы III зависит от бета-субъединицы Все ДНК-полимеразы
- 27. ? Соотнесите разные бактериальные ДНК-полимеразы с их функциями в клетке
- 28. Репликация ДНК у E.coli
- 29. Различия репликации у прокариот и эукариот
- 30. Терминация репликации у прокариот У прокариот есть участок TerC, на котором заканчивается репликация ДНК. На кольцевой
- 31. Терминация репликации у эукариот У эукариот нет специфического сайта для терминации. Терминация происходит, когда сливаются репликационные
- 32. Репликация теломер Оловников А.М.
- 33. Концевые участки хромосом представлены особыми участками, называемыми теломерами. Теломеры представляют собой многочисленные консервативные повторы TG, стабилизированные
- 34. Резюме Репликация ДНК начинается в особом участке, называемом ориджином репликации. В случае прокариотической клетки имеется лишь
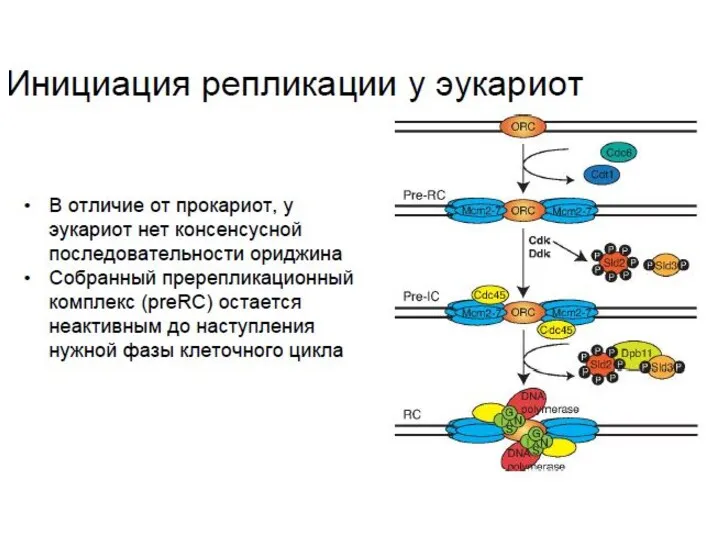
- 35. Резюме В эукариотических клетках инициация репликации начинается со сборки пререпликационного комплекса посредством присоединения к ориджину (без
- 36. Резюме Процесс элонгации вовлекает множество других белков и ферментов. ДНК-полимеразе необходима затравка (праймер) для начала репликации,
- 37. Резюме Основной репликативной полимеразой у бактерий является ДНК-полимераза III — мультисубъединичный комплекс, ДНК-полимераза I задействована в
- 38. Спасибо за внимание!
- 40. Скачать презентацию

Источники:
Курсы лекций и презентации:
«Молекулярная биология и генетика»
«Молекулярная биология клетки»
https://stepik.org
https://lectoriy.mipt.ru/course/Biology-Molecular-14L
Дейч К.О "О
Источники:
Курсы лекций и презентации:
«Молекулярная биология и генетика»
«Молекулярная биология клетки»
https://stepik.org
https://lectoriy.mipt.ru/course/Biology-Molecular-14L
Дейч К.О "О
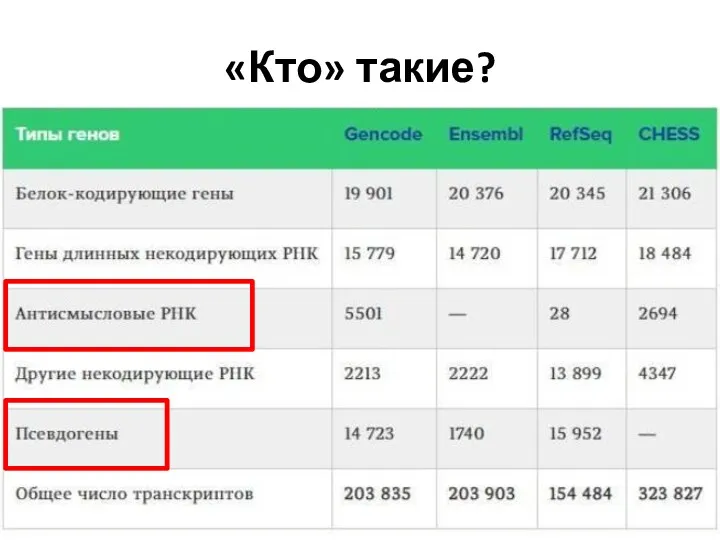
«Кто» такие?
«Кто» такие?
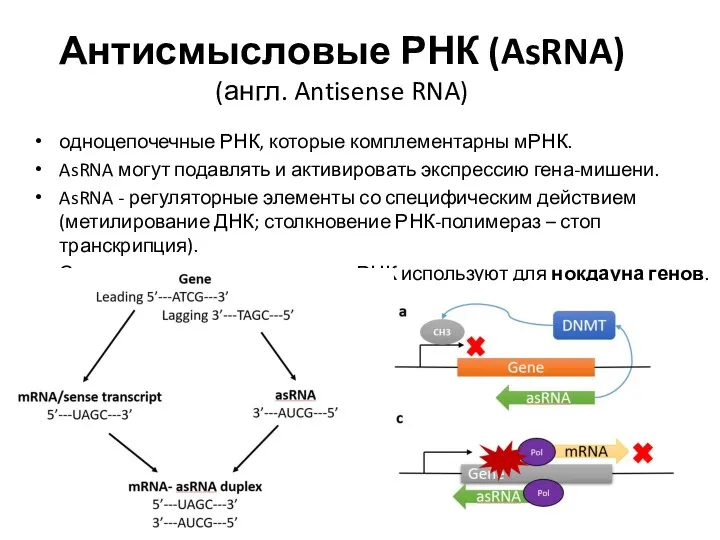
Антисмысловые РНК (AsRNA) (англ. Antisense RNA)
одноцепочечные РНК, которые комплементарны мРНК.
AsRNA
Антисмысловые РНК (AsRNA) (англ. Antisense RNA)
одноцепочечные РНК, которые комплементарны мРНК.
AsRNA

Псевдогены
(англ. pseudogenes)
Псевдогены — нефункциональные аналоги структурных генов, утратившие способность
Псевдогены
(англ. pseudogenes)
Псевдогены — нефункциональные аналоги структурных генов, утратившие способность

Лекция 3.
Репликация ДНК
как инициируется репликация у про- и эукариот, что такое
Лекция 3.
Репликация ДНК
как инициируется репликация у про- и эукариот, что такое
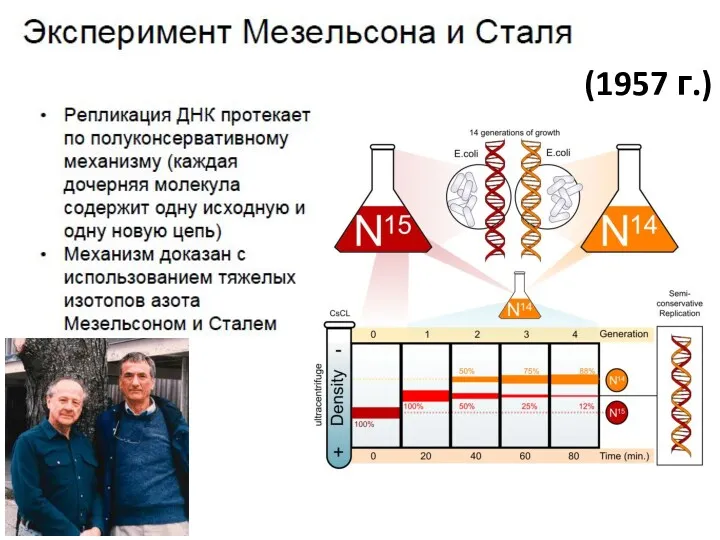
(1957 г.)
(1957 г.)
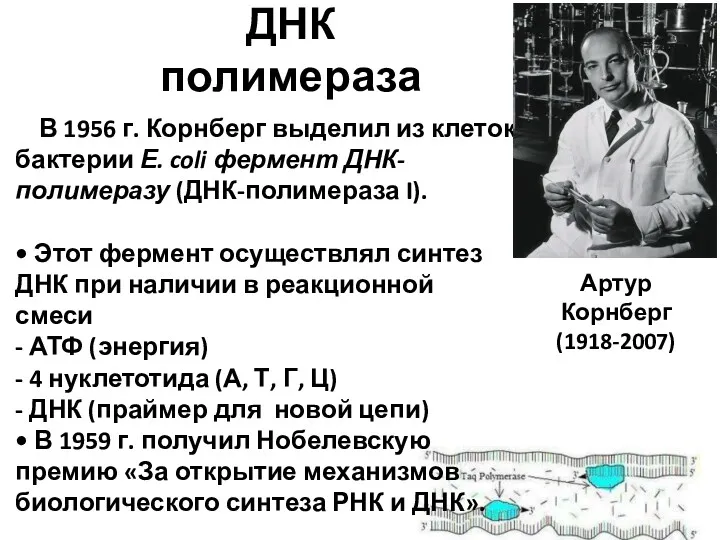
ДНК полимераза
В 1956 г. Корнберг выделил из клеток бактерии Е.
ДНК полимераза
В 1956 г. Корнберг выделил из клеток бактерии Е.
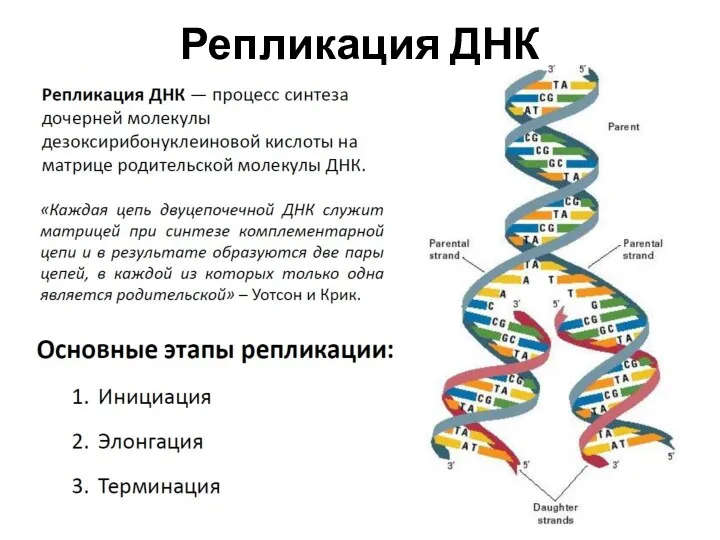
Репликация ДНК
Репликация ДНК

Основные ферменты репликации
ДНК-полимераза - фермент катализирующий полимеризацию дезоксирибонуклеотидов на матрице ДНК
Основные ферменты репликации
ДНК-полимераза - фермент катализирующий полимеризацию дезоксирибонуклеотидов на матрице ДНК
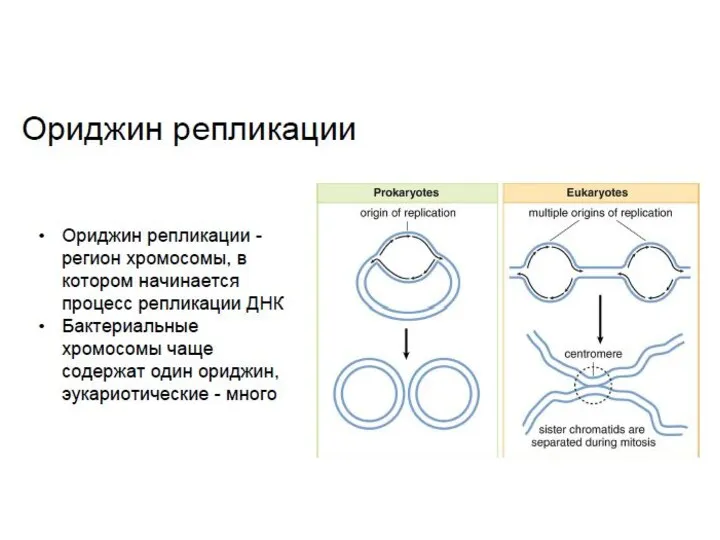

Количество точек
начала репликации в геноме
Геном вироидов ( 1 РНК )
Количество точек
начала репликации в геноме
Геном вироидов ( 1 РНК )

?
Как соотносится число ориджинов репликации в хромосомах бактерий и эукариот?
У эукариот
?
Как соотносится число ориджинов репликации в хромосомах бактерий и эукариот?
У эукариот

?
Выберите все верные утверждения, касающиеся инициации репликации у про- и эукариот:
У
?
Выберите все верные утверждения, касающиеся инициации репликации у про- и эукариот:
У
https://www.youtube.com/watch?v=BmAq-EolVCc
https://www.youtube.com/watch?v=BmAq-EolVCc
https://www.youtube.com/watch?v=BmAq-EolVCc
Okazaki R, Okazaki T, Sakabe K, Sugimoto K. Mechanism of DNA
https://www.youtube.com/watch?v=BmAq-EolVCc
Okazaki R, Okazaki T, Sakabe K, Sugimoto K. Mechanism of DNA
https://www.youtube.com/watch?v=BmAq-EolVCc
https://www.youtube.com/watch?v=BmAq-EolVCc
https://www.youtube.com/watch?v=BmAq-EolVCc
Элонгация репликации
Репликация идет однонаправленно, от 5′-конца к 3′-концу новой цепи
ДНК-лигаза
https://www.youtube.com/watch?v=BmAq-EolVCc
Элонгация репликации
Репликация идет однонаправленно, от 5′-конца к 3′-концу новой цепи
ДНК-лигаза

?
Почему репликация на одной цепи ДНК идет непрерывно, а на другой
?
Почему репликация на одной цепи ДНК идет непрерывно, а на другой
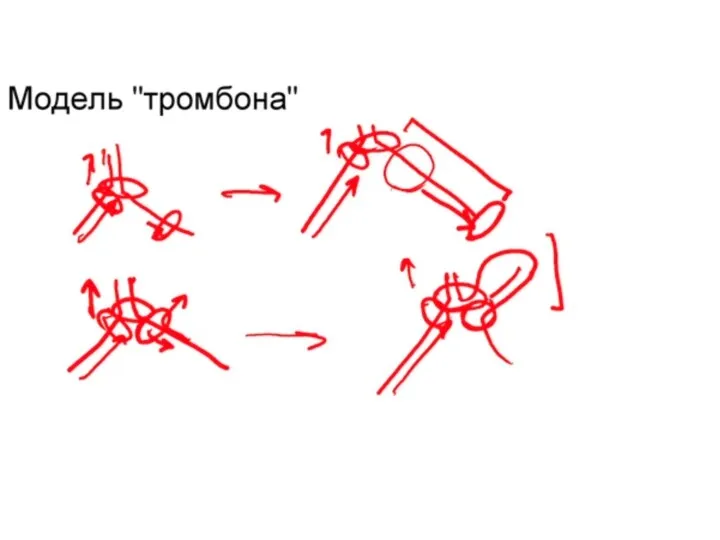
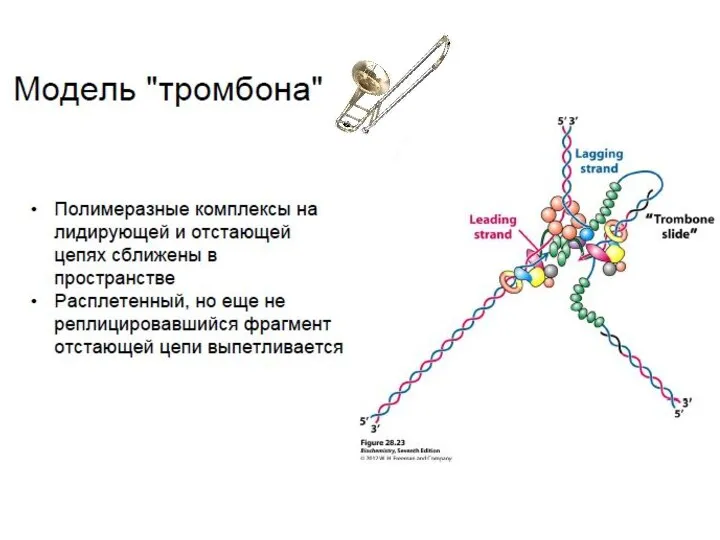
Репликация ДНК
Свойства ДНК-полимераз эукариот
Репликация ДНК
Свойства ДНК-полимераз эукариот

?
Отметьте верные утверждения, касающиеся ДНК-полимераз бактерий.
Процессивность ДНК полимеразы III зависит
?
Отметьте верные утверждения, касающиеся ДНК-полимераз бактерий.
Процессивность ДНК полимеразы III зависит

?
Соотнесите разные бактериальные ДНК-полимеразы с их функциями в клетке
?
Соотнесите разные бактериальные ДНК-полимеразы с их функциями в клетке
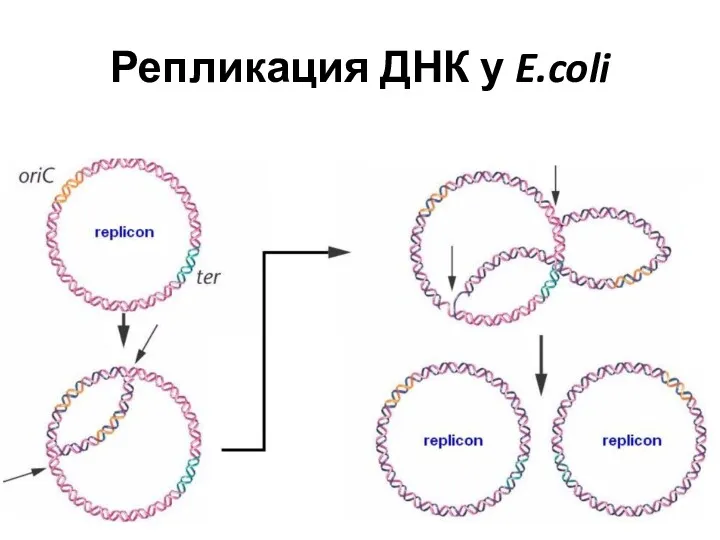
Репликация ДНК у E.coli
Репликация ДНК у E.coli
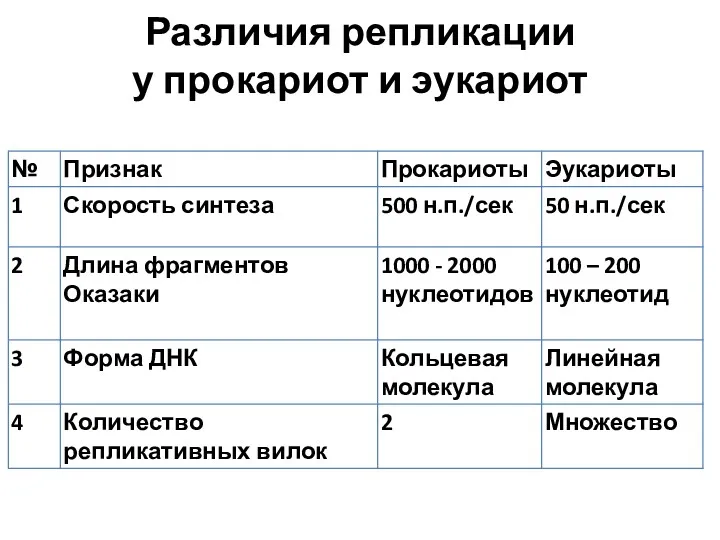
Различия репликации
у прокариот и эукариот
Различия репликации
у прокариот и эукариот
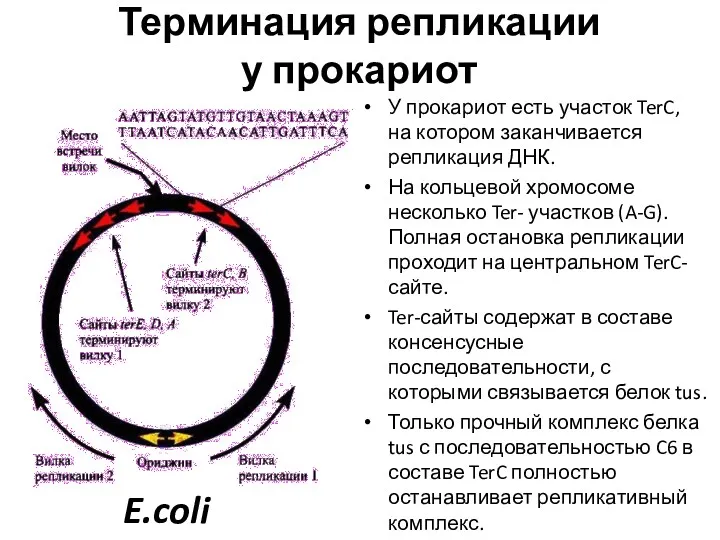
Терминация репликации
у прокариот
У прокариот есть участок TerC, на котором заканчивается
Терминация репликации
у прокариот
У прокариот есть участок TerC, на котором заканчивается

Терминация репликации
у эукариот
У эукариот нет специфического сайта для терминации. Терминация
Терминация репликации
у эукариот
У эукариот нет специфического сайта для терминации. Терминация
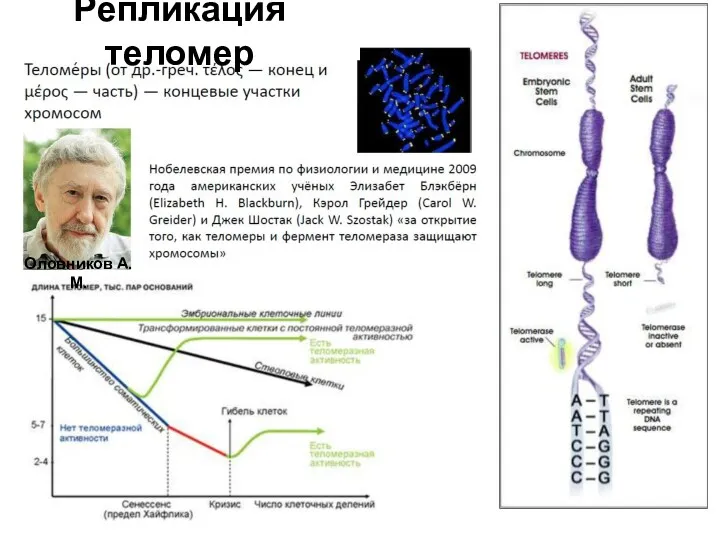
Репликация теломер
Оловников А.М.
Репликация теломер
Оловников А.М.
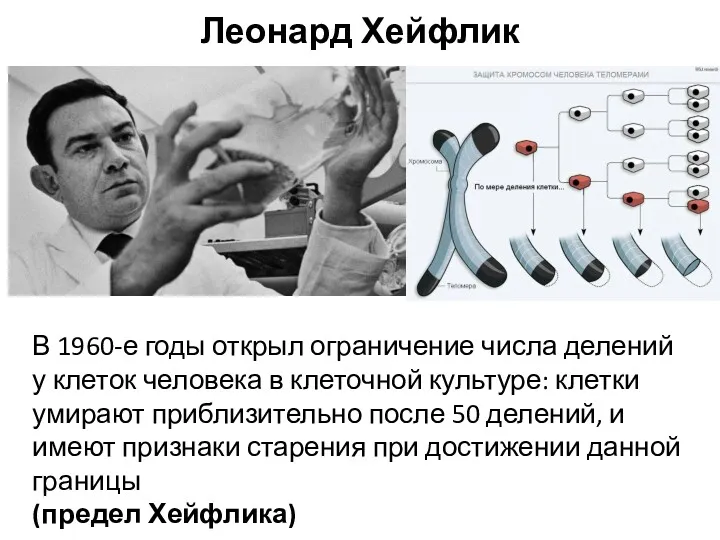
Концевые участки хромосом представлены особыми участками, называемыми теломерами. Теломеры представляют собой
Концевые участки хромосом представлены особыми участками, называемыми теломерами. Теломеры представляют собой

Резюме
Репликация ДНК начинается в особом участке, называемом ориджином репликации. В случае
Резюме
Репликация ДНК начинается в особом участке, называемом ориджином репликации. В случае

Резюме
В эукариотических клетках инициация репликации начинается со сборки пререпликационного комплекса посредством
Резюме
В эукариотических клетках инициация репликации начинается со сборки пререпликационного комплекса посредством

Резюме
Процесс элонгации вовлекает множество других белков и ферментов. ДНК-полимеразе необходима затравка (праймер)
Резюме
Процесс элонгации вовлекает множество других белков и ферментов. ДНК-полимеразе необходима затравка (праймер)

Резюме
Основной репликативной полимеразой у бактерий является ДНК-полимераза III — мультисубъединичный комплекс, ДНК-полимераза
Резюме
Основной репликативной полимеразой у бактерий является ДНК-полимераза III — мультисубъединичный комплекс, ДНК-полимераза

Спасибо за внимание!
Спасибо за внимание!
 Отряд перепончатокрылые
Отряд перепончатокрылые Вопросы, с использованием картинок. ОГЭ по биологии-2019. Открытый банк заданий ФИПИ. Часть 2. Живые организмы
Вопросы, с использованием картинок. ОГЭ по биологии-2019. Открытый банк заданий ФИПИ. Часть 2. Живые организмы Использование возможностей цифровой лаборатории в преподавании предметов естественнонаучного цикла
Использование возможностей цифровой лаборатории в преподавании предметов естественнонаучного цикла Строение и функции клеточной мембраны
Строение и функции клеточной мембраны Покритонасінні. Принципи біологічної систематики
Покритонасінні. Принципи біологічної систематики Игра Насекомые
Игра Насекомые Размножение растений
Размножение растений Презентация ядовитые растения
Презентация ядовитые растения Органы чувств. Регуляция деятельности
Органы чувств. Регуляция деятельности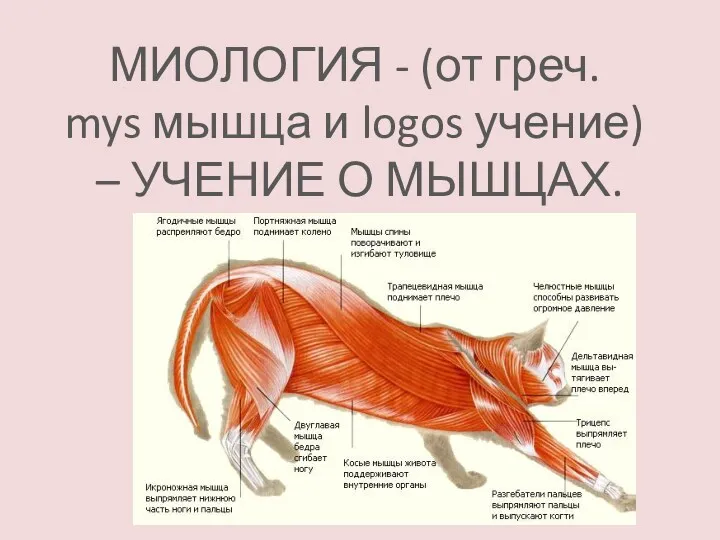 Миология. Мышечная ткань
Миология. Мышечная ткань Влияние экологических факторов на организмы
Влияние экологических факторов на организмы Витамины группы Б
Витамины группы Б История изучения клетки. Клеточная теория
История изучения клетки. Клеточная теория Деревья и кустарники
Деревья и кустарники Биологическое окисление
Биологическое окисление Строение и функции кожи
Строение и функции кожи Дробление. Деление зиготы
Дробление. Деление зиготы презентация к уроку биологии общая характеристика царства растений 7 кл
презентация к уроку биологии общая характеристика царства растений 7 кл Дельфины. (3 класс)
Дельфины. (3 класс)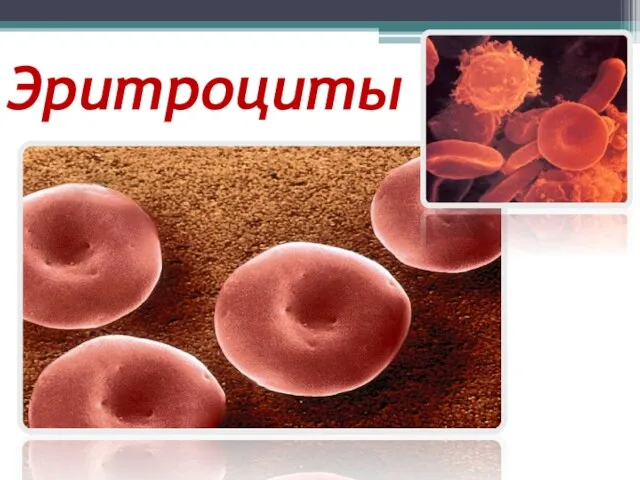 Особенности строения эритроцитов
Особенности строения эритроцитов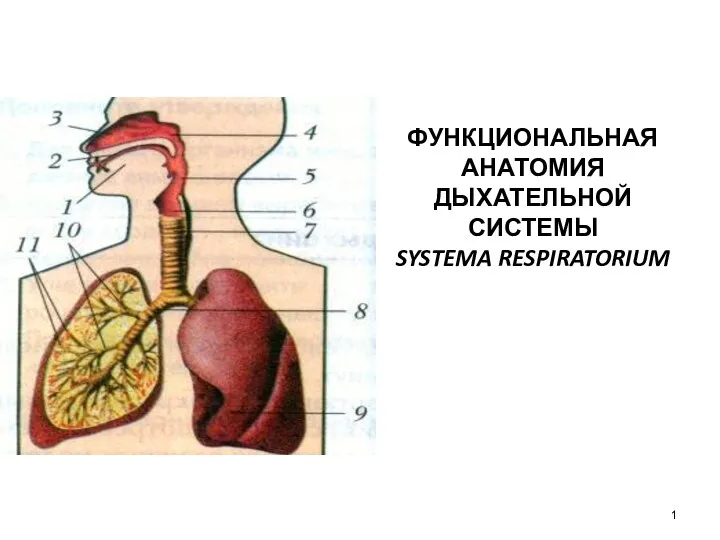 Функциональная анатомия дыхательной системы. Systema respiratorium
Функциональная анатомия дыхательной системы. Systema respiratorium Презентация Иммунитет
Презентация Иммунитет Борьба с цистицеркозом
Борьба с цистицеркозом Тип хордовые. Особенности строения
Тип хордовые. Особенности строения Клеточное строение листа
Клеточное строение листа Краткие комментарии по новой демоверсии КИМ ЕГЭ 2017 года по биологии
Краткие комментарии по новой демоверсии КИМ ЕГЭ 2017 года по биологии Мир животных - мир звуков
Мир животных - мир звуков Есту анализаторы
Есту анализаторы