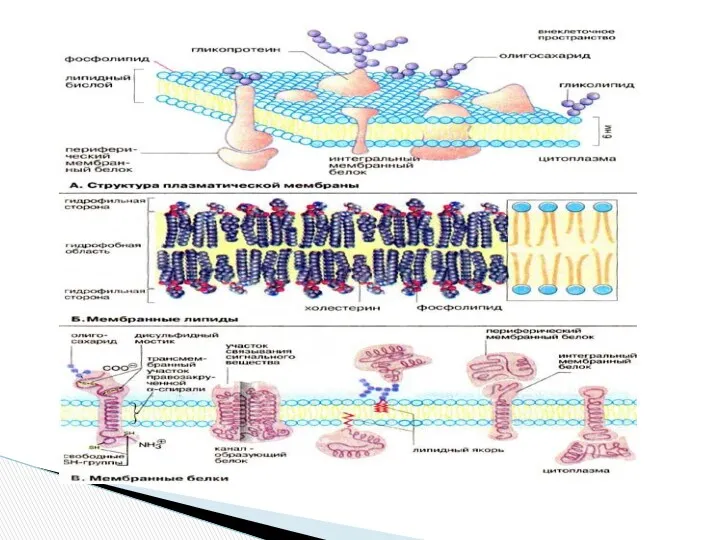
- Рецепторы клетки

Содержание
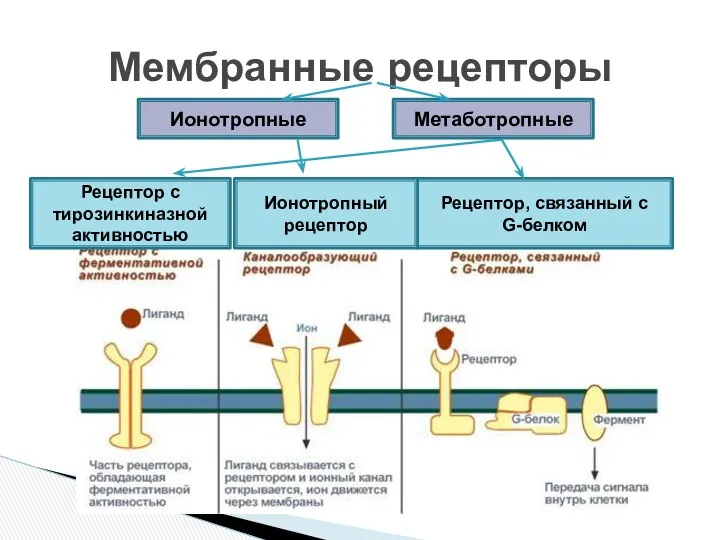
- 3. Мембранные рецепторы Рецептор с тирозинкиназной активностью Рецептор, связанный с G-белком Ионотропный рецептор Метаботропные Ионотропные
- 4. Представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом, что приводит к ионному току
- 5. Нервные импульсы вызывают высвобождение ацетилхолина в синаптическую щель, после чего он взаимодействует с холинорецепторами. Соединение ацетилхолина
- 6. ИОНОТРОПНЫЕ РЕЦЕПТОРЫ N-холинорецептор
- 7. Метаботропные рецепторы
- 8. Участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Виды «вторичных посредников»: циклические нуклеотиды (цАМФ и
- 9. Представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и внутриклеточный C-конец. Сайт связывания с
- 10. G-связанный рецептор (схема строения)
- 11. АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА ФОСФОИНОЗИТИДНАЯ СИСТЕМА Компоненты аденилатциклазной системы: мембранный белок-рецептор G-белок Аденилатциклаза (фермент) Гуанозинтрифосфат (ГТФ) Протеинкиназы АТФ.
- 12. До момента действия гормона компоненты находятся в диссоциированнном состоянии. После образования комплекса сигнальной молекулы (гормон) с
- 13. ц-АМФ активирует ферменты – протеинкиназы , которые катализируют реакции фосфорилирования различных белков (ферменты, ядерные белки, мембранные
- 14. Работа адренорецептора
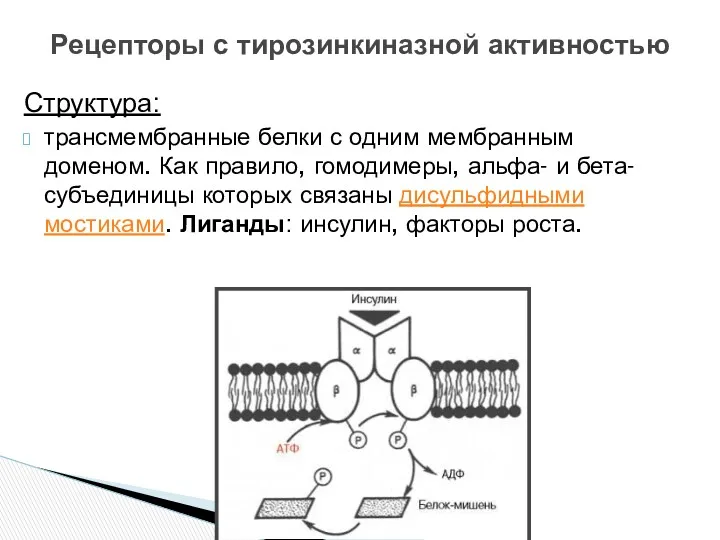
- 15. Структура: трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, альфа- и бета-субъединицы которых связаны дисульфидными
- 16. Инсулин с высокой специфичностью связывается и распознаётся α-субъединицей рецептора, которая при присоединении гормона изменяет свою конформацию.
- 17. Работа тирозинкиназного рецептора
- 18. растворимые ДНК-связывающие белки не связаны с плазматической мембраной, локализованы внутри ядра клетки!!! Лиганды: глюкокортикостероидные и тиреоидные
- 19. проникновение гормона через билипидный слой мембраны в клетку образование комплекса гормон-рецептор, который перемещается в ядро клетки
- 21. Скачать презентацию
Мембранные рецепторы
Рецептор с тирозинкиназной активностью
Рецептор, связанный с
G-белком
Ионотропный
рецептор
Метаботропные
Ионотропные
Мембранные рецепторы
Рецептор с тирозинкиназной активностью
Рецептор, связанный с
G-белком
Ионотропный
рецептор
Метаботропные
Ионотропные

Представляют собой мембранные каналы, открываемые или закрываемые при связывании с лигандом,

Нервные импульсы вызывают высвобождение ацетилхолина в синаптическую щель, после чего он
Нервные импульсы вызывают высвобождение ацетилхолина в синаптическую щель, после чего он
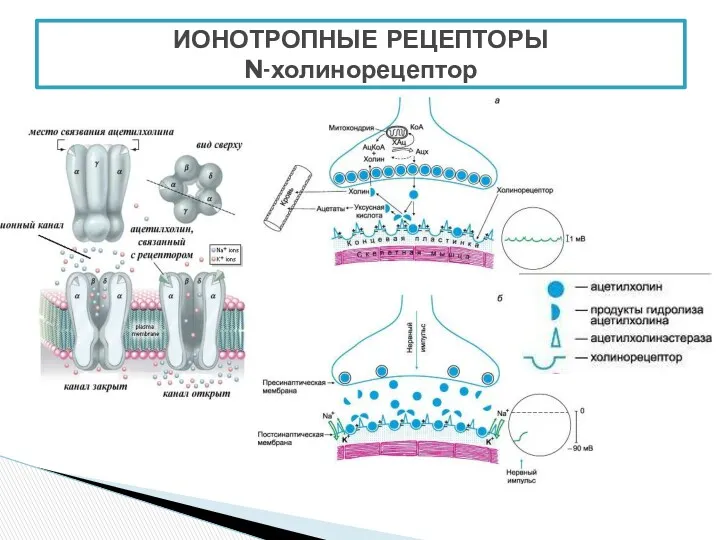
ИОНОТРОПНЫЕ РЕЦЕПТОРЫ
N-холинорецептор
ИОНОТРОПНЫЕ РЕЦЕПТОРЫ
N-холинорецептор

Метаботропные рецепторы
Метаботропные рецепторы

Участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Виды «вторичных посредников»:
циклические
Участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Виды «вторичных посредников»:
циклические
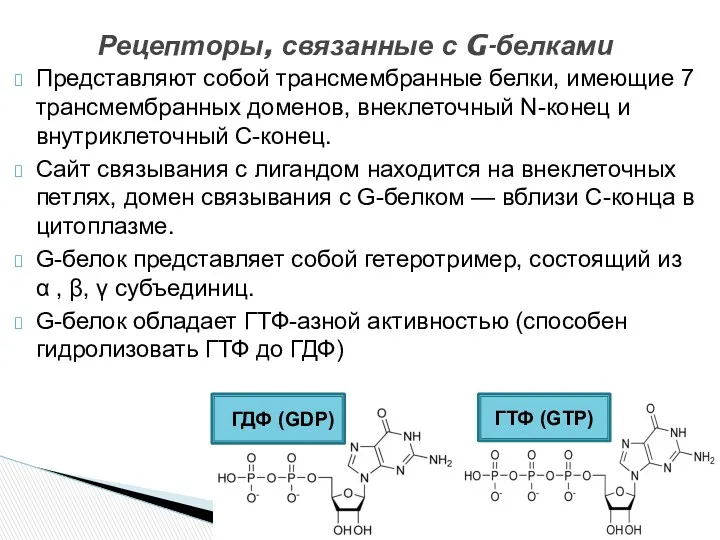
Представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и
Представляют собой трансмембранные белки, имеющие 7 трансмембранных доменов, внеклеточный N-конец и
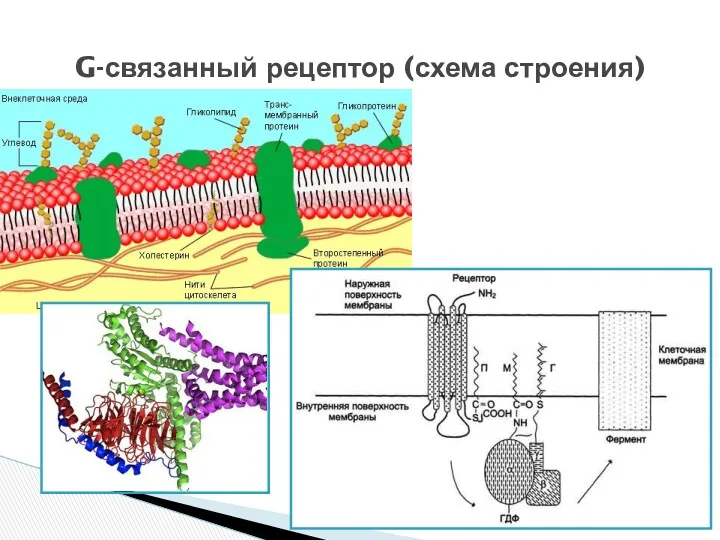
G-связанный рецептор (схема строения)
G-связанный рецептор (схема строения)

АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА
ФОСФОИНОЗИТИДНАЯ СИСТЕМА
Компоненты аденилатциклазной системы:
мембранный белок-рецептор
G-белок
Аденилатциклаза (фермент)
Гуанозинтрифосфат (ГТФ)
Протеинкиназы
АТФ.
Существует два
АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА
ФОСФОИНОЗИТИДНАЯ СИСТЕМА
Компоненты аденилатциклазной системы:
мембранный белок-рецептор
G-белок
Аденилатциклаза (фермент)
Гуанозинтрифосфат (ГТФ)
Протеинкиназы
АТФ.
Существует два

До момента действия гормона компоненты находятся в диссоциированнном состоянии. После образования
До момента действия гормона компоненты находятся в диссоциированнном состоянии. После образования

ц-АМФ активирует ферменты – протеинкиназы , которые катализируют реакции фосфорилирования различных
ц-АМФ активирует ферменты – протеинкиназы , которые катализируют реакции фосфорилирования различных
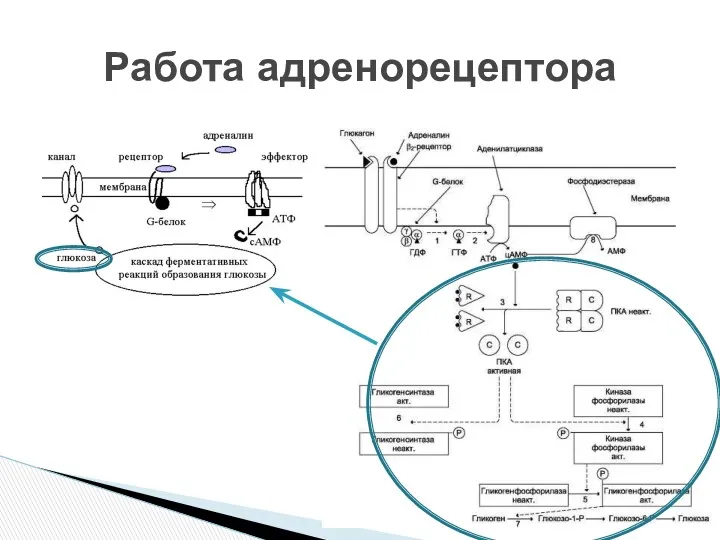
Работа адренорецептора
Работа адренорецептора
Структура:
трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, альфа- и
Структура:
трансмембранные белки с одним мембранным доменом. Как правило, гомодимеры, альфа- и

Инсулин с высокой специфичностью связывается и распознаётся α-субъединицей рецептора, которая при
Инсулин с высокой специфичностью связывается и распознаётся α-субъединицей рецептора, которая при
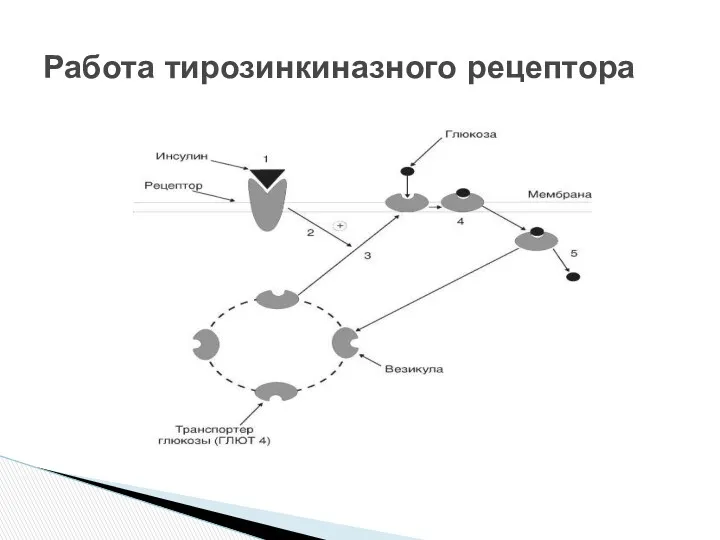
Работа тирозинкиназного рецептора
Работа тирозинкиназного рецептора

растворимые ДНК-связывающие белки
не связаны с плазматической мембраной, локализованы внутри ядра клетки!!!
Лиганды:
растворимые ДНК-связывающие белки
не связаны с плазматической мембраной, локализованы внутри ядра клетки!!!
Лиганды:

проникновение гормона через билипидный слой мембраны в клетку
образование комплекса гормон-рецептор, который
проникновение гормона через билипидный слой мембраны в клетку
образование комплекса гормон-рецептор, который
 Личинки животных
Личинки животных Ч.Дарвиннің. Жердегі тіршіліктің эволюциясы теориясының негізгі қағидалары,эволюцияның синтетикалық теориясы
Ч.Дарвиннің. Жердегі тіршіліктің эволюциясы теориясының негізгі қағидалары,эволюцияның синтетикалық теориясы Для нимфальной стадии клеща
Для нимфальной стадии клеща Физиология и биохимия микроорганизмов
Физиология и биохимия микроорганизмов Нервная система. Спинной мозг. Рефлектроная дуга
Нервная система. Спинной мозг. Рефлектроная дуга Выделение. Мочевыделительная система
Выделение. Мочевыделительная система Болезни овощей и фруктов
Болезни овощей и фруктов Надтип Amera. Тип Моллюски, или мягкотелые MOLLUSCA
Надтип Amera. Тип Моллюски, или мягкотелые MOLLUSCA Сон-трава или прострел раскрытый
Сон-трава или прострел раскрытый Передвижение веществ в организме растения
Передвижение веществ в организме растения Развитие тутового шелкопряда
Развитие тутового шелкопряда Царство Растения
Царство Растения Теория существования прокариот
Теория существования прокариот Как появился человек на Земле
Как появился человек на Земле Травы и цветы полей и лугов
Травы и цветы полей и лугов Класс двудольные. Класс однодольные. 6 класс
Класс двудольные. Класс однодольные. 6 класс Условия полноценного развития системы кровообращения
Условия полноценного развития системы кровообращения Железы внутренней и смешанной секреции
Железы внутренней и смешанной секреции Влияние музыки на растения
Влияние музыки на растения Тип Моллюски
Тип Моллюски Закономерности изменчивости
Закономерности изменчивости Кровеносная система
Кровеносная система Polymers and amino acids
Polymers and amino acids Пауки и паутина. Рисование паутины пауков
Пауки и паутина. Рисование паутины пауков Питание китов
Питание китов Презентация по биологии 8 класс темы Внутренняя среда организма
Презентация по биологии 8 класс темы Внутренняя среда организма Генетика пола. Наследование,сцепленное с полом
Генетика пола. Наследование,сцепленное с полом Системна радіобіологія: від молекули до організму. Системна радіобіологія: від організму до популяції
Системна радіобіологія: від молекули до організму. Системна радіобіологія: від організму до популяції