Ряд общебиологических обобщений имеет прямое отношение к становлению и развитию современных представлений об ЭБР презентация
- Ряд общебиологических обобщений имеет прямое отношение к становлению и развитию современных представлений об ЭБР

Содержание
- 2. РЕГЛАМЕНТ ЗАНЯТИЙ: 12 Семинаров 10 Лекций 2
- 3. Ряд общебиологических обобщений имеет прямое отношение к становлению и развитию современных представлений об ЭБР: Открытие и
- 4. Теория зародышевых листков 4 ученик Каспара Фридриха Вольфа
- 5. ввёл в характеристику «типов» Ж.Кювье «эмбриологический принцип»: каждый главный тип животной организации следует особому плану развития
- 6. История представлений об эволюционных аспектах эмбриогенеза и онтогенеза Натурфилософский период: самозарождение - креационизм Новое время: преформизм
- 7. «Эмбриология значительно выигрывает в интересе, если мы будем видеть в зародыше более или менее потускневший портрет
- 8. Эрнст Геккель (1834-1919) Фриц Мюллер (1821-1897) «Эволюция – это преобразование онтогенезов. Не зная фундаментальных закономерностей онтогенеза,
- 9. Заслуги Геккеля: Популяризация учения Ч. Дарвина и «создание» питекантропа Формулировка Биогенетического закона Создание концепции происхождения многоклеточных
- 10. Выдержки из монографии Н.Ю. Клюге: Современная систематика насекомых. СПб.: Лань, 2000. – 336с. Для обоснования «биогенетического
- 11. Применимость биогенетического закона к конкретному случаю: «… работа, вызвавшая наибольший резонанс в прессе, касалась одного вида
- 12. Четвериков Сергей Сергеевич (18-19) … факты, служившие основанием для одной концепции, часто могут получать диаметрально противоположное
- 13. История представлений об эволюционных аспектах эмбриогенеза и онтогенеза Натурфилософский период: самозарождение - креационизм Новое время: преформизм
- 14. Анаболия (надставка) – животное проходит через все стадии, свойственные предкам, после чего испытывает дополнительные изменения, добавляя
- 15. Андрей Валентинович Макрушин (род. 1934г.), российский гидробиолог, д.б.н. (г. Борок Ярославской обл.) А. В. Макрушин, В.
- 16. История представлений об эволюционных аспектах эмбриогенеза и онтогенеза Натурфилософский период: самозарождение - креационизм Новое время: преформизм
- 17. Гетеробатмия – неодинаковый уровень специализации различных частей одного целого, достигнутый в процессе биологической эволюции Гетеробатмия (=
- 18. Наиболее широко известны три эволюционных способа изменения эмбрионального и личиночного развития: разобщение (диссоциация) процессов развития во
- 19. Возможности онтогенеза: типы гетерохронии (Gould, 1977; с небольшими изменениями) 19 Стивен Джей Гулд (1941-2002)
- 20. 20 Формы гетерохронии (Рупперт и др, 2008) Параметры обеих классификаций: Длительность развития сомы Скорость развития сомы
- 21. Биологический энциклопедический словарь, 1986 Аббревиация (Б.С.Матвеев, 1930) – выпадение конечных стадий онтогенеза Анаболия (А.Н.Северцов, 1912) –
- 22. Значение изучения полиморфизма Полиморфизм - важный критерий индикации условий обитания и состояния популяции. Различают изменчивость, связанную
- 23. 2. умножение (дупликация) частей с последующей их дивергенцией Впервые на полимеризацию, как ведущий принцип эволюции обратил
- 24. История представлений об эволюционных аспектах эмбриогенеза и онтогенеза Натурфилософский период: самозарождение - креационизм Новое время: преформизм
- 25. Факторы эволюции (теория стабилизирующего отбора). – М.-Л.: Изд. АН СССР, 1948. 396с. Организм как целое в
- 26. Основные постулаты ЭТЭ (Шишкин, 2003): Наследственность — не партнёр естественного отбора, а его продукт, выступающий как
- 27. Эволюция как изменение онтогенезов Вопрос о том, какой биологический или культурно-социальный признак стал определяющим в формировании
- 28. Алексей Петрович Быстров (1899-1959) В 1957 г. в Ленинграде вышла книга выдающегося анатома и палеонтолога Алексея
- 29. 29 Ряд групп животных : черви (как свободноживущие, так и паразитические), паразитические моллюски (см. рис.), иглокожие,
- 30. Рост размеров биосистем – одна из очевидных эволюционных тенденций Этапы усложнения биосистем – это и переход
- 31. Совершенствование метаболического модуля при росте размеров Диффузия и циркуляция могут обеспечить оптимальный метаболизм (А) даже без
- 32. Тенденции в развитии морфологии адсотрофов Геометрически эти тенденции можно свести к небольшому перечню «операций» (удлинение, уплощение,
- 33. Связи уровня метаболизма и массы тела у организмов разных уровней сложности (Рупперт и др., 2008, т.1,
- 34. Исторические трансформации онтогенезов (на примере аммонитов) 34 Трансформации шва аммонитов Появление гетероморфных аммонитов, принимавшихся ранее за
- 35. Вариации пропорций раковин брюхоногих как рефрен Множество неродственных форм брюхоногих, как ныне живущих, так и вымерших,
- 36. Происхождение мелких интерстициальных видов путем педоморфоза (А) Предок лишенного параподий представителя Dorvelliidae с трохусами мог выглядеть
- 37. Схема жизненного цикла многоклеточных животных (Рупперт и др., 2008. т.1., рис. 4.3.) 37 Разнообразие возможностей трансформации
- 38. Динамика вымираний морских животных 1 – вымирание; 2 – первое появление; 3 – оледенения; 4 -
- 39. Ведущие принципы для объяснения причин развития (Белоусов, 1987) Финалистический принцип (целевая причина по Аристотелю) – процесс
- 40. Исторический (эволюционный) принцип в эмбриологии Во 2-й половине 19 века идеи эволюционизма (исторический принцип) дали мощный
- 41. Каузально-аналитический принцип При всей сложности и плодотворности исторического принципа нельзя не заметить, что при таком подходе
- 42. Концептуальное обоснование механики развития благодаря изучению непосредственных причин развития, осмыслению роли физических и химических факторов в
- 43. Продолжение следует: Будем говорить о гомологиях …
- 44. Первые представления о гомологии В биологии понятие гомологии старше, чем аналогии Евклид (-325 - -265) Клавдий
- 45. Леонардо-да-Винчи (1452-1512) Белон Пьер (1517-1564) Антони ван Левенгук (1632-1723)
- 46. Марко Аурелио Северино (1580-1656) Фрэнсис Бэкон (1561-1626) Рэй Ланкастер (1847-1949) Ричард Оуэн (1804-1892)
- 48. Скачать презентацию
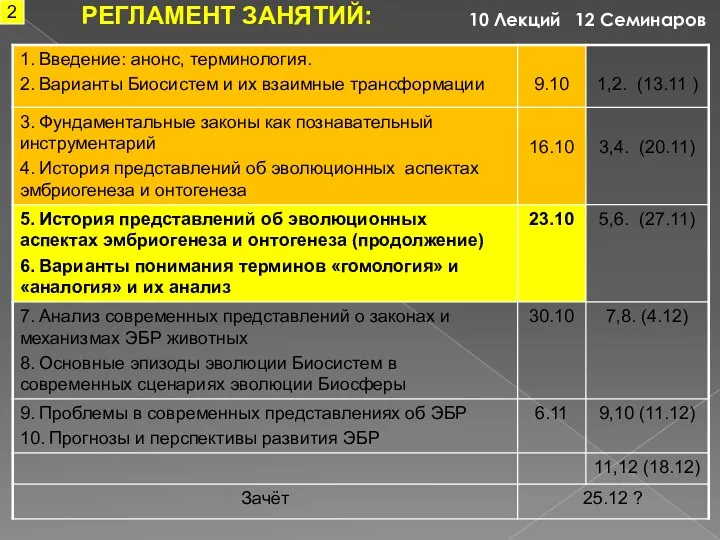
РЕГЛАМЕНТ ЗАНЯТИЙ:
12 Семинаров
10 Лекций
2
РЕГЛАМЕНТ ЗАНЯТИЙ:
12 Семинаров
10 Лекций
2

Ряд общебиологических обобщений имеет прямое отношение
к становлению и развитию
Ряд общебиологических обобщений имеет прямое отношение к становлению и развитию

Теория зародышевых листков
4
ученик Каспара Фридриха Вольфа
Теория зародышевых листков
4
ученик Каспара Фридриха Вольфа

ввёл в характеристику «типов» Ж.Кювье «эмбриологический принцип»: каждый главный тип животной
ввёл в характеристику «типов» Ж.Кювье «эмбриологический принцип»: каждый главный тип животной

История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение
История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение
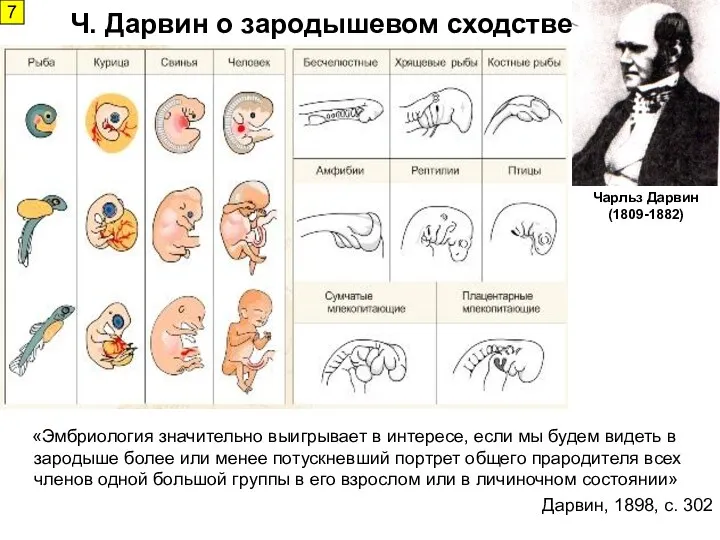
«Эмбриология значительно выигрывает в интересе, если мы будем видеть в
«Эмбриология значительно выигрывает в интересе, если мы будем видеть в
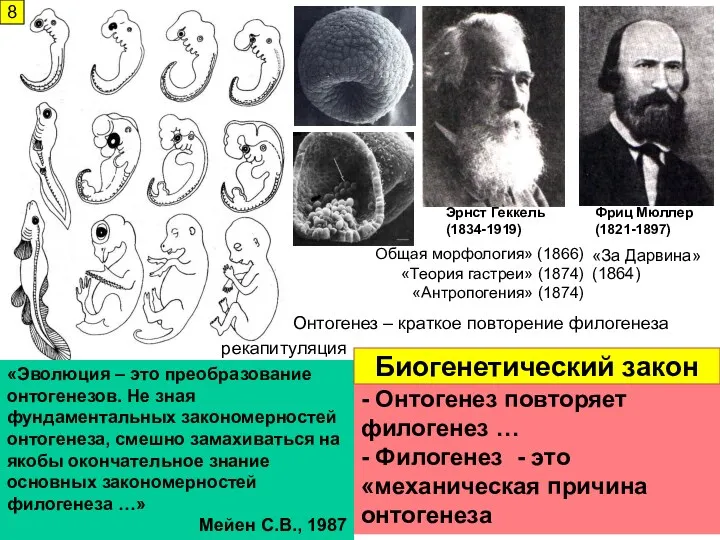
Эрнст Геккель
(1834-1919)
Фриц Мюллер
(1821-1897)
«Эволюция – это преобразование онтогенезов. Не зная фундаментальных закономерностей
Эрнст Геккель
(1834-1919)
Фриц Мюллер
(1821-1897)
«Эволюция – это преобразование онтогенезов. Не зная фундаментальных закономерностей

Заслуги Геккеля:
Популяризация учения Ч. Дарвина и «создание» питекантропа
Формулировка Биогенетического закона
Создание концепции
Заслуги Геккеля:
Популяризация учения Ч. Дарвина и «создание» питекантропа
Формулировка Биогенетического закона
Создание концепции

Выдержки из монографии Н.Ю. Клюге:
Современная систематика насекомых. СПб.: Лань, 2000.
Выдержки из монографии Н.Ю. Клюге: Современная систематика насекомых. СПб.: Лань, 2000.

Применимость биогенетического закона к конкретному случаю:
«… работа, вызвавшая наибольший резонанс
Применимость биогенетического закона к конкретному случаю:
«… работа, вызвавшая наибольший резонанс

Четвериков
Сергей Сергеевич
(18-19)
… факты, служившие основанием для одной концепции, часто могут
Четвериков
Сергей Сергеевич
(18-19)
… факты, служившие основанием для одной концепции, часто могут

История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение
История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение
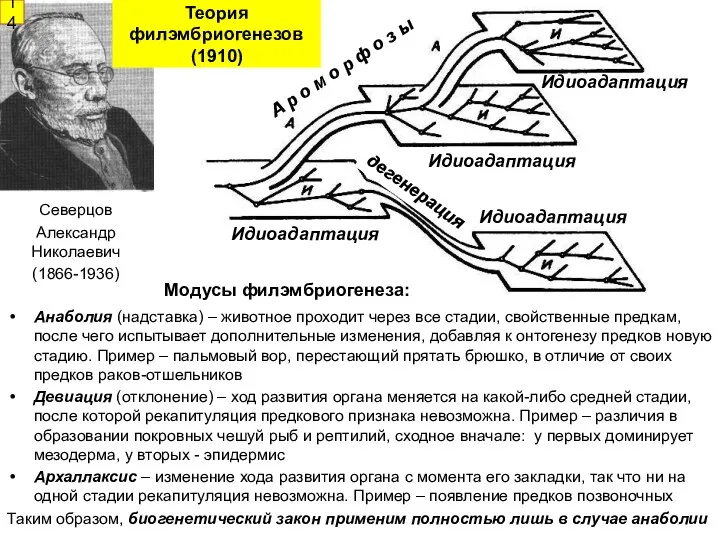
Анаболия (надставка) – животное проходит через все стадии, свойственные предкам, после
Анаболия (надставка) – животное проходит через все стадии, свойственные предкам, после

Андрей Валентинович Макрушин
(род. 1934г.), российский гидробиолог,
д.б.н.
(г. Борок Ярославской обл.)
А. В. Макрушин, В. В.
Андрей Валентинович Макрушин
(род. 1934г.), российский гидробиолог,
д.б.н.
(г. Борок Ярославской обл.)
А. В. Макрушин, В. В.

История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение
История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение

Гетеробатмия – неодинаковый уровень специализации различных частей одного целого, достигнутый в
Гетеробатмия – неодинаковый уровень специализации различных частей одного целого, достигнутый в

Наиболее широко известны три эволюционных способа изменения эмбрионального и личиночного развития:
разобщение
Наиболее широко известны три эволюционных способа изменения эмбрионального и личиночного развития:
разобщение
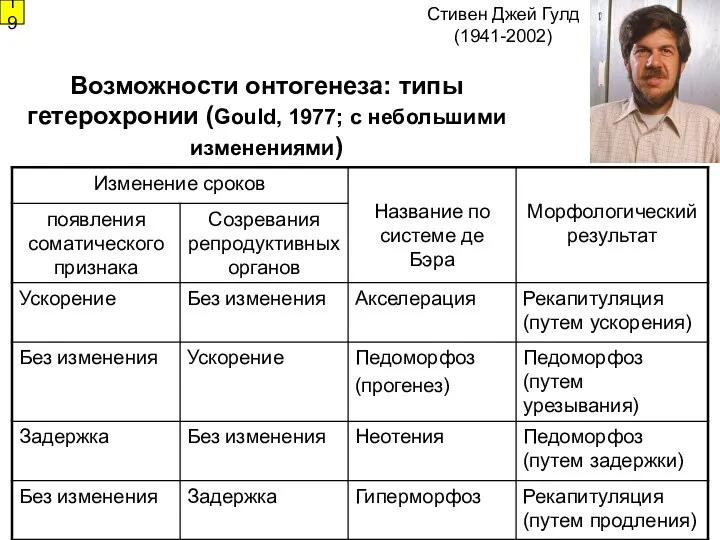
Возможности онтогенеза: типы гетерохронии (Gould, 1977; с небольшими изменениями)
19
Стивен Джей Гулд
(1941-2002)
Возможности онтогенеза: типы гетерохронии (Gould, 1977; с небольшими изменениями)
19
Стивен Джей Гулд
(1941-2002)
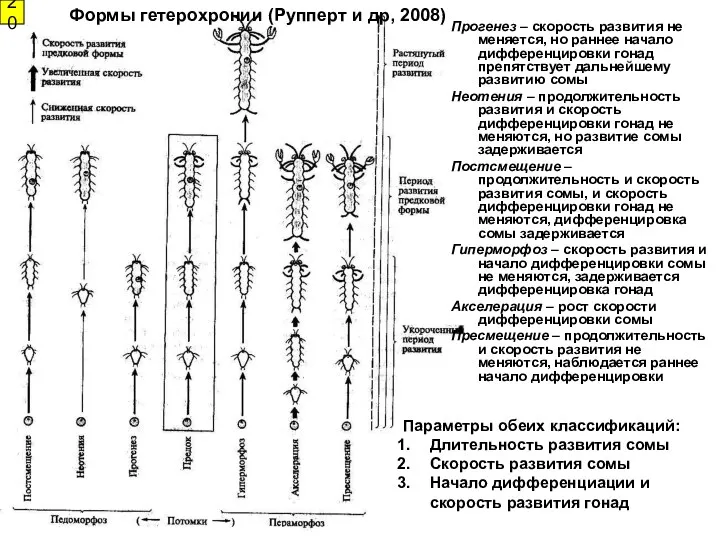
20
Формы гетерохронии (Рупперт и др, 2008)
Параметры обеих классификаций:
Длительность развития сомы
Скорость
20
Формы гетерохронии (Рупперт и др, 2008)
Параметры обеих классификаций:
Длительность развития сомы
Скорость

Биологический энциклопедический словарь, 1986
Аббревиация (Б.С.Матвеев, 1930) – выпадение конечных стадий онтогенеза
Анаболия
Биологический энциклопедический словарь, 1986
Аббревиация (Б.С.Матвеев, 1930) – выпадение конечных стадий онтогенеза
Анаболия

Значение изучения полиморфизма
Полиморфизм - важный критерий индикации условий обитания и состояния
Значение изучения полиморфизма
Полиморфизм - важный критерий индикации условий обитания и состояния

2. умножение (дупликация) частей
с последующей их дивергенцией
Впервые на полимеризацию,
2. умножение (дупликация) частей
с последующей их дивергенцией
Впервые на полимеризацию,

История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение
История представлений об эволюционных аспектах эмбриогенеза и онтогенеза
Натурфилософский период: самозарождение

Факторы эволюции (теория стабилизирующего отбора). – М.-Л.: Изд. АН СССР, 1948.
Факторы эволюции (теория стабилизирующего отбора). – М.-Л.: Изд. АН СССР, 1948.

Основные постулаты ЭТЭ (Шишкин, 2003):
Наследственность — не партнёр естественного отбора, а
Основные постулаты ЭТЭ (Шишкин, 2003):
Наследственность — не партнёр естественного отбора, а

Эволюция как изменение онтогенезов
Вопрос о том, какой биологический или культурно-социальный признак
Эволюция как изменение онтогенезов
Вопрос о том, какой биологический или культурно-социальный признак
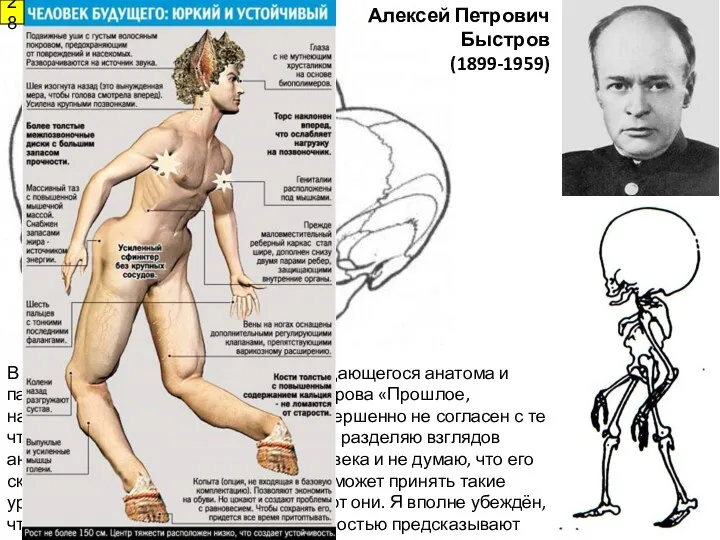
Алексей Петрович Быстров
(1899-1959)
В 1957 г. в Ленинграде вышла книга выдающегося
Алексей Петрович Быстров
(1899-1959)
В 1957 г. в Ленинграде вышла книга выдающегося
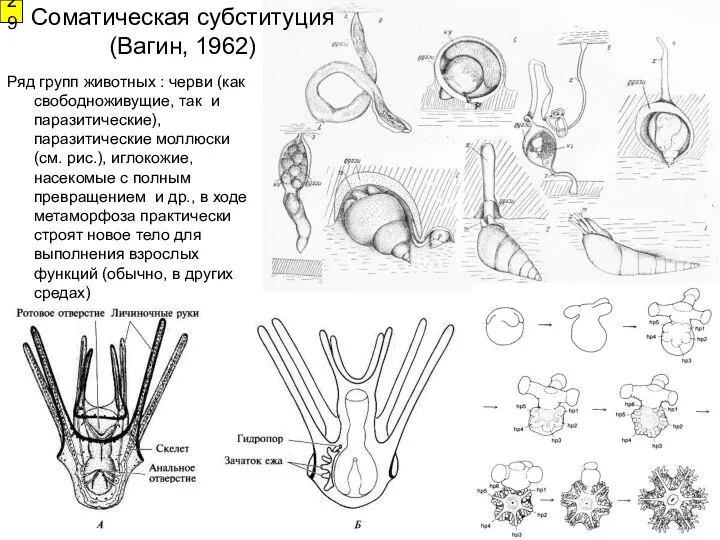
29
Ряд групп животных : черви (как свободноживущие, так и паразитические), паразитические
29
Ряд групп животных : черви (как свободноживущие, так и паразитические), паразитические
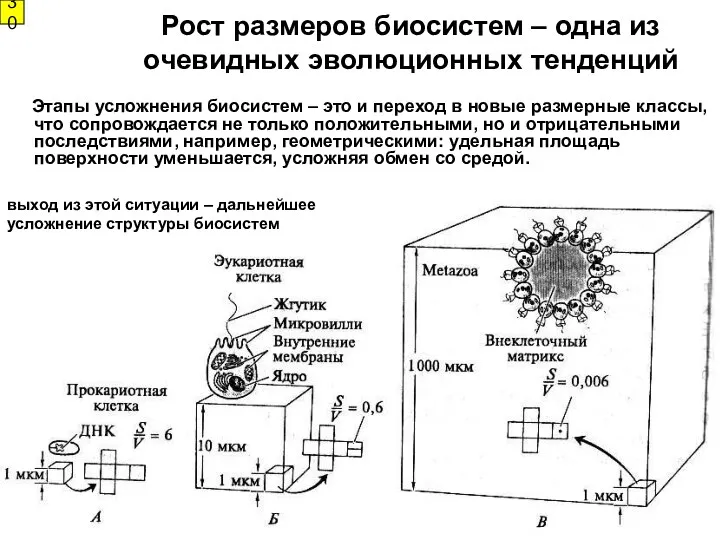
Рост размеров биосистем – одна из очевидных эволюционных тенденций
Этапы усложнения
Рост размеров биосистем – одна из очевидных эволюционных тенденций
Этапы усложнения

Совершенствование метаболического модуля при росте размеров
Диффузия и циркуляция могут обеспечить оптимальный
Совершенствование метаболического модуля при росте размеров
Диффузия и циркуляция могут обеспечить оптимальный
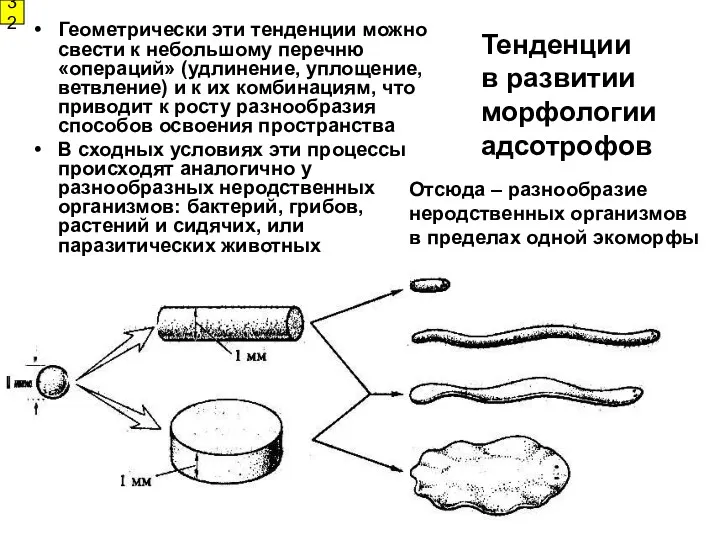
Тенденции
в развитии
морфологии адсотрофов
Геометрически эти тенденции можно свести к небольшому
Тенденции
в развитии
морфологии адсотрофов
Геометрически эти тенденции можно свести к небольшому
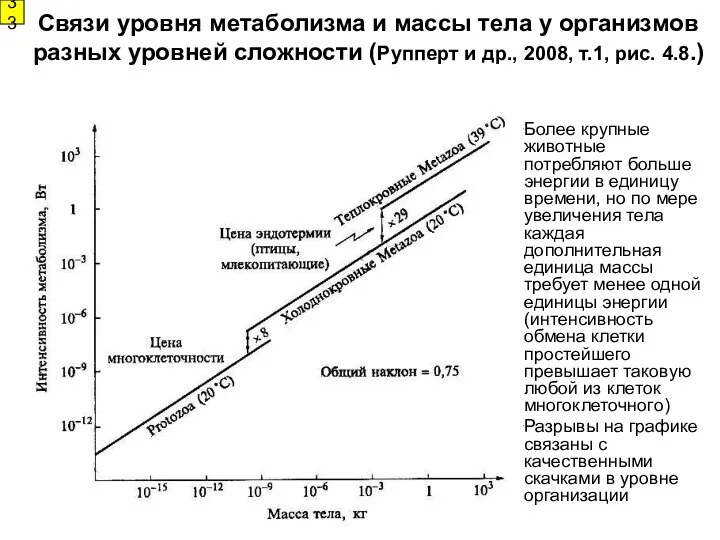
Связи уровня метаболизма и массы тела у организмов разных уровней сложности
Связи уровня метаболизма и массы тела у организмов разных уровней сложности
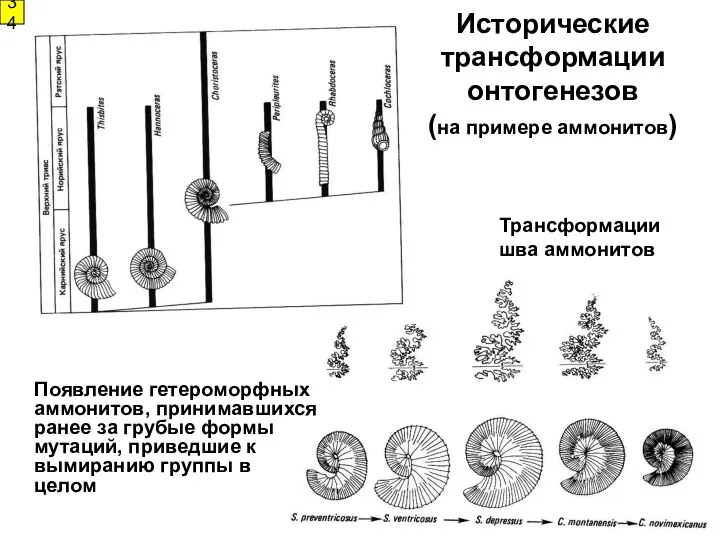
Исторические трансформации онтогенезов
(на примере аммонитов)
34
Трансформации
шва аммонитов
Появление гетероморфных аммонитов,
Исторические трансформации онтогенезов
(на примере аммонитов)
34
Трансформации
шва аммонитов
Появление гетероморфных аммонитов,
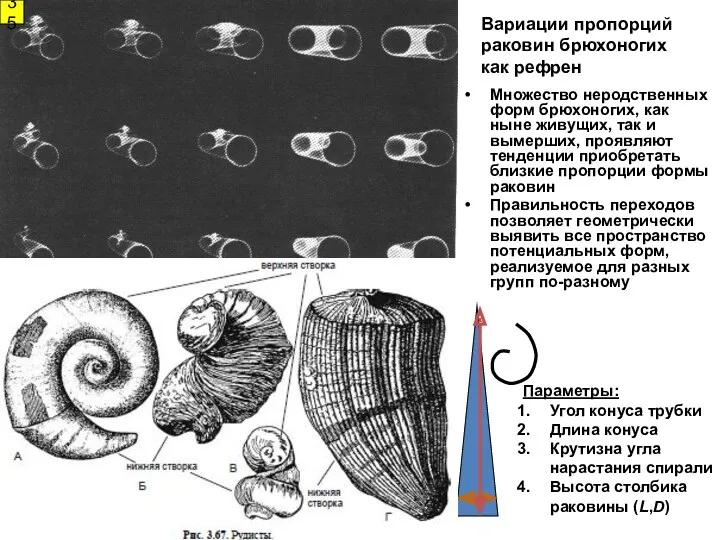
Вариации пропорций раковин брюхоногих
как рефрен
Множество неродственных форм брюхоногих, как ныне
Вариации пропорций раковин брюхоногих
как рефрен
Множество неродственных форм брюхоногих, как ныне
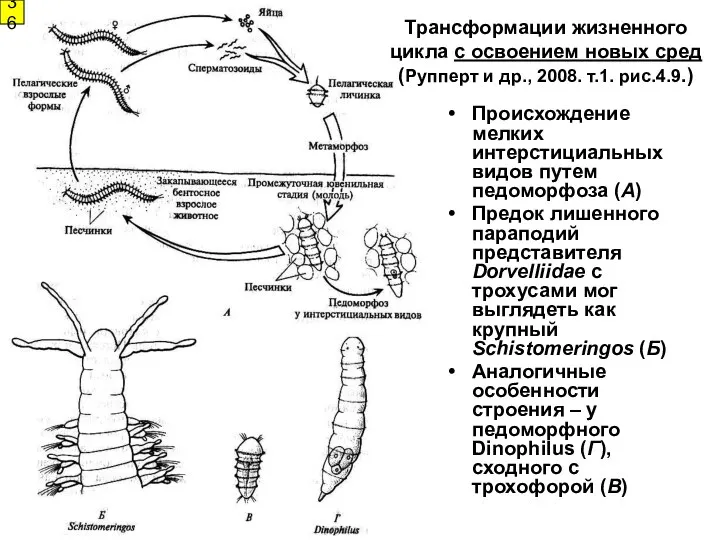
Происхождение мелких интерстициальных видов путем педоморфоза (А)
Предок лишенного параподий представителя Dorvelliidae
Происхождение мелких интерстициальных видов путем педоморфоза (А)
Предок лишенного параподий представителя Dorvelliidae

Схема жизненного цикла многоклеточных животных
(Рупперт и др., 2008. т.1., рис.
Схема жизненного цикла многоклеточных животных (Рупперт и др., 2008. т.1., рис.
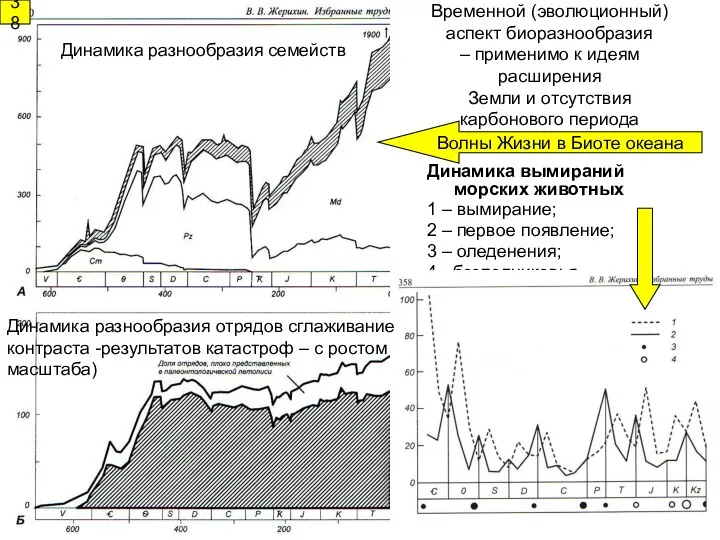
Динамика вымираний морских животных
1 – вымирание;
2 – первое появление;
3
Динамика вымираний морских животных
1 – вымирание;
2 – первое появление;
3

Ведущие принципы для объяснения причин развития (Белоусов, 1987)
Финалистический принцип (целевая причина
Ведущие принципы для объяснения причин развития (Белоусов, 1987)
Финалистический принцип (целевая причина

Исторический (эволюционный) принцип в эмбриологии
Во 2-й половине 19 века идеи эволюционизма
Исторический (эволюционный) принцип в эмбриологии
Во 2-й половине 19 века идеи эволюционизма

Каузально-аналитический принцип
При всей сложности и плодотворности исторического принципа нельзя не заметить,
Каузально-аналитический принцип
При всей сложности и плодотворности исторического принципа нельзя не заметить,

Концептуальное обоснование механики развития благодаря изучению непосредственных причин развития, осмыслению роли
Концептуальное обоснование механики развития благодаря изучению непосредственных причин развития, осмыслению роли

Продолжение следует:
Будем говорить о гомологиях …
Продолжение следует:
Будем говорить о гомологиях …

Первые представления о гомологии
В биологии понятие гомологии старше, чем аналогии
Евклид
(-325
Первые представления о гомологии
В биологии понятие гомологии старше, чем аналогии
Евклид
(-325
Леонардо-да-Винчи
(1452-1512)
Белон Пьер
(1517-1564)
Антони ван Левенгук
(1632-1723)
Леонардо-да-Винчи
(1452-1512)
Белон Пьер
(1517-1564)
Антони ван Левенгук
(1632-1723)

Марко Аурелио Северино (1580-1656)
Фрэнсис Бэкон (1561-1626)
Рэй Ланкастер (1847-1949)
Ричард Оуэн (1804-1892)
Марко Аурелио Северино (1580-1656)
Фрэнсис Бэкон (1561-1626)
Рэй Ланкастер (1847-1949)
Ричард Оуэн (1804-1892)
 Организация генома. Прокариот и эукариот
Организация генома. Прокариот и эукариот Ягоды. Кроссворд
Ягоды. Кроссворд Обитатели тайги
Обитатели тайги Bases of phytocenology
Bases of phytocenology Опорно-двигательная система. Скелет, строение и соединение костей
Опорно-двигательная система. Скелет, строение и соединение костей Мутации – изменения в структуре генов
Мутации – изменения в структуре генов Минеральные вещества. Эндемические заболевания
Минеральные вещества. Эндемические заболевания Охраняемые животные в российской Артике
Охраняемые животные в российской Артике Общая характеристика транспортных процессов в клетке
Общая характеристика транспортных процессов в клетке Размножение и развитие организмов
Размножение и развитие организмов Растения-рекордсмены
Растения-рекордсмены Царство Животные
Царство Животные Зернобобові культури. Лекція 3
Зернобобові культури. Лекція 3 Химический состав растений
Химический состав растений Продукция пчеловодства: мед, прополис, пчелиный яд, маточное молочко, пыльца, воск. Урок 203-204
Продукция пчеловодства: мед, прополис, пчелиный яд, маточное молочко, пыльца, воск. Урок 203-204 Влияние СМИ и компьютера на здоровье подростков
Влияние СМИ и компьютера на здоровье подростков Нервная и гуморальная регуляция функций организма
Нервная и гуморальная регуляция функций организма Растениеводство. Гречиха, как культурное растение
Растениеводство. Гречиха, как культурное растение Биохимия нервной ткани. (Лекция 23)
Биохимия нервной ткани. (Лекция 23) Вирусы. Особенности вирусов
Вирусы. Особенности вирусов Таргетинг генов
Таргетинг генов Лишайники. Разнообразие
Лишайники. Разнообразие Презентация Многообразие биогеоценозов
Презентация Многообразие биогеоценозов Оплодотворение. Этапы
Оплодотворение. Этапы Невидимые нити. Окружающий мир, 2 класс
Невидимые нити. Окружающий мир, 2 класс Биоэнергетика мышечной деятельности
Биоэнергетика мышечной деятельности Дыхательная система человека
Дыхательная система человека Водне середовище життя. Пристосування організмів до життя у воді
Водне середовище життя. Пристосування організмів до життя у воді