- Short history of post-transcriptional gene silencing

Содержание
- 2. What is RNA interference /PTGS? dsRNA needs to be directed against an exon, not an intron
- 3. double-stranded RNAs are produced by: – transcription of inverted repeats – viral replication – transcription of
- 4. RNAi can be induced by:
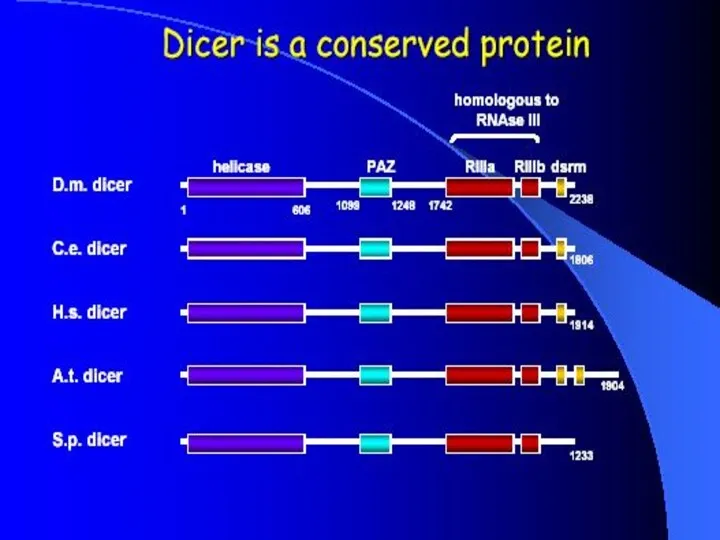
- 8. Dicer Double-stranded RNA triggers processed into siRNAs by enzyme RNAseIII family, specifically the Dicer family Processive
- 10. RISC complex RISC is a large (~500-kDa) RNA-multiprotein complex, which triggers mRNA degradation in response to
- 11. Different classes of small RNA molecules During dsRNA cleavage, different RNA classes are produced: – siRNA
- 12. siRNAs Small interfering RNAs that have an integral role in the phenomenon of RNA interference(RNAi), a
- 13. miRNAs/stRNAs micro/small temporal RNAs derive from ~70 nt ssRNA (single-stranded RNA), which forms a stemloop; processed
- 14. MEM MEM )
- 15. Why is PTGS important? Most widely held view is that RNAi evolved to protect the genome
- 16. Recent applications of RNAi Modulation of HIV-1 replication by RNA interference. Hannon(2002). Potent and specific inhibition
- 17. Possible future improvements of RNAi applications Already developed: in vitro synthesis of siRNAs using T7 RNA
- 18. Conclusions begun in worms, flies, and plants - as an accidental observation. general applications in mammalian
- 19. Регуляция экспрессии генов с помощью miRNA
- 20. DNA-интерференция DNA-guided DNA interference by a prokaryotic Argonaute. Swarts DC, Jore MM, Westra ER, Zhu Y,
- 21. Функции siРНК Сайленсинг мобильных генетических элементов; Сайленсинг гетерохроматиновых повторов; Сайленсинг генетического материала вирусного происхождения; Ограничение степени
- 22. При выделение фракций коротких РНК (19-25 нуклеотидов) из различных организмов обнаружен еще один класс малых РНК
- 23. Функция miРНК Обеспечивают сайленсинг различных генов, обычно, за счет частично комплементарного связывания с мРНК, в результате
- 24. Продукт dsРНК, закодированных в уникальных генах геномов многоклеточных организмов (>1% от всех генов у человека); мРНК
- 25. созданы библиотеки коротких РНК и ДНК-векторов, кодирующих короткие РНК, мишенями которых является около 8000 генов генома
- 26. Fig. 3. Structural preference of miRNA–miRNA* asymmetry in miRNA-induced gene silencing complex (RISC) in vivo. Different
- 27. In vivo gene-silencing effects of anti- b -catenin miRNA and anti-noggin miRNA ( d ) on
- 28. In vivo effects of anti-tyrosinase ( Tyr ) miRNA on the mouse pigment production of local
- 29. Morphological and genetic properties of mirPSCs. ( a ) A morphological comparison between a morula-staged rat
- 30. Mechanism of miR-302–mediated tumor suppression in human iPSCs. miR-302 not only concurrently suppresses G1-phase checkpoint regulators
- 32. What is RNA interference (RNAi)? “The Process by which dsRNA silences gene expression...” Degradation of mRNA
- 33. What are sense and antisense RNA? Messenger RNA (mRNA) is single-stranded, called "sense" because it results
- 34. What are sense and antisense RNA? Antisense molecules interact with complementary strands of nucleic acids, modifying
- 35. RNAi terms dsRNA: double stranded RNA, longer than 30 nt miRNA: microRNA, 21-25 nt. Encoded by
- 36. RNAi like phenomena Plants Petunias Fungi Neurospora Animals Caenorhabditis elegans Alternate terms to RNAi PTGS (Posttranscriptional
- 37. 1990-Petunias Napoli et al. defined an RNAi-like phenomenon and called it “cosupression.” chalcone synthase (CHS), a
- 38. Overexpression of chalcone synthase in petunias unexpectedly resulted in white petunias The levels of endogenous as
- 39. 1992-The mold Carlo Cogoni and Guiseppe Macino of the Università di Roma La Sapienza in Italy
- 40. 1995-The worm Guo and Kemphues studied par-1 gene during embryogenesis The worm, C. elegans has a
- 41. 1995- The worm Guo and Kemphues first studied Par-1 gene mutants Division: Asymmetric?symmetric P-granule distribution
- 42. Guo and Kemphues, 1995
- 43. Both the antisense and sense strands effectively silenced wildtype Par-1 RNAi
- 44. ‘Antisense’ Technology? Sense RNA silences yet no hybridization of sense RNA with sense mRNA is expected!
- 45. Craig Mello In 1996, C. Mello and his student S. Driver also reported that sense RNAs
- 46. 1998-Fire et al and Mello Gel-purified ssRNA Used purified ssRNA (antisense and sense) separately and also
- 47. Unc-22 (Uncoordinated 22) Codes for a non essential myofilament It is present several thousand copies/cell
- 48. Injection for RNAi 6-10 adult hermaphrodites were injected with 0.5x106-1x106 molecules into each gonadal arm.
- 49. Unc-22 phenotype 4-6 hours after injection, eggs collected. Screened for phenotypic changes twiching
- 50. Mex-3 mex-3 encodes two RNA binding proteins; in the early embryo, maternally provided Mex-3 is required
- 51. Mex-3 RNAi b, Embryo from uninjected parent (showing normal pattern of endogenous mex-3 RNA20). c, Embryo
- 52. RNAi concentration and dose response 3.6x106 molecules/gonad Sense phenocopied 1% of progeny Antisense phenocopied 11% of
- 53. Quantitative Assays
- 54. Other possibilities Sense+antisense in low salt Rapid sequential injection of sense & antisense Both cause interference
- 55. Conclusions www.nobelprize.org
- 56. Conclusions www.nobelprize.org
- 57. Ways to induce silent phenotypes Timmons and Fire showed that feeding dsRNA works! Reversible and gene-specific
- 58. Ways to induce silent phenotypes Tabarra, Grishok, and Mello in 1998 demonstrated that soaking in dsRNA
- 59. Mechanisms revealed 25bp species of dsRNA found in plants with co-suppression [Hamilton and Baulcombe, 1999] Sequence
- 60. RNAi: two phases Initiation Generation of mature siRNA or miRNA Execution Silencing of target gene Degradation
- 61. How does RNAi work? www.nobelprize.org
- 62. siRNA biogenesis Dicer (type III RNAse III) cleaves long dsRNA into siRNA 21-25nt dsRNA from exogenous
- 63. RNA Induced Silencing Complex (RISC) RNAi effector complex Preferentially incorporates one strand of unwound RNA [Khvorova
- 64. siRNA design Mittal, 2004
- 65. Custom-made siRNAs
- 66. siRNA libraries Generation of a feeding clone Tuschl, 2003 siRNA libraries Result: 16 757 bacterial strains
- 67. Endogenous RNAi-miRNA We have hundreds of different genes that encode small RNA (collectively, microRNA) whose precursors
- 68. Defense Against Viruses www.nobelprize.org Indeed, Baulcombe, Vance, and others have shown that, in the continuing evolutionary
- 69. Mammalian RNAi McManus and Sharp, 2002
- 70. Getting Around the Problem siRNA (21-22nt) mediate mammalian RNAi Introducing siRNA instead of dsRNA prevents non-specific
- 71. Some applications of RNAi Therapy Candidate genes, drug discovery, and therapy Genome-wide RNAi screens Gene function
- 72. Genome-wide RNAi Only 11% genes showed detectable RNAi phenotype Between 600-800 genes are required for early
- 73. Systems Biology and RNAi Cellular systems act as networks of interacting components (genes, RNA, protein, metabolites,…).
- 74. Networks of Early Embryogenesis Protein-protein interaction dataset: binary physical interactions between 3,848 C. elegans proteins Transcriptome
- 75. Systems Biology Approach: Three networks in one
- 76. The embryogenesis network
- 77. Discovery Project
- 78. Defense against transposons RNAi may also help keep the transposable elements that litter genomes from jumping
- 80. Why use RNAi? 1. The most powerful way to inhibit gene expression and acquire info about
- 84. Скачать презентацию

What is RNA interference /PTGS?
dsRNA needs to be directed
What is RNA interference /PTGS?
dsRNA needs to be directed

double-stranded RNAs are produced by:
– transcription of inverted repeats
– viral
double-stranded RNAs are produced by:
– transcription of inverted repeats
– viral
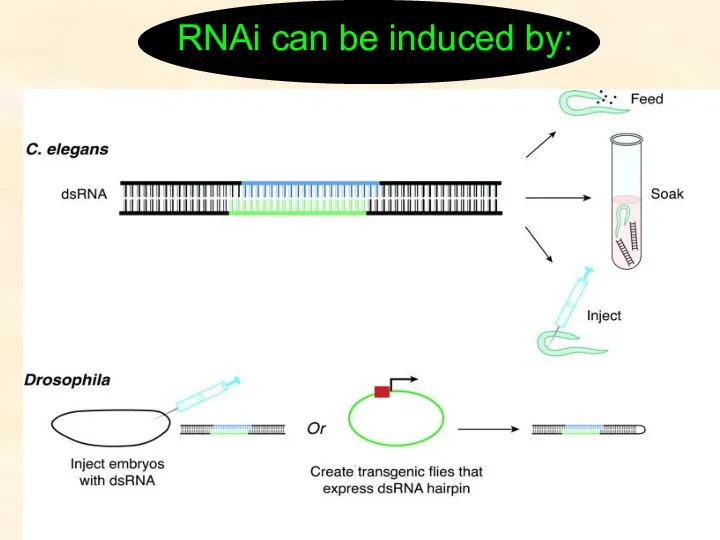
RNAi can be induced by:
RNAi can be induced by:
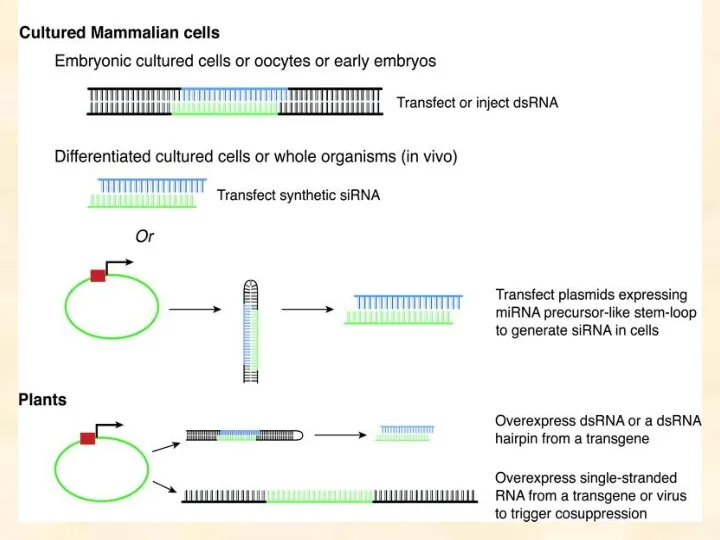
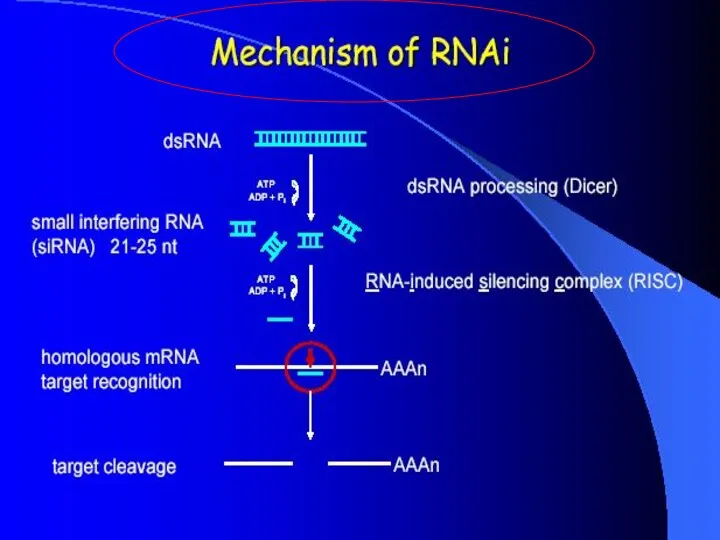
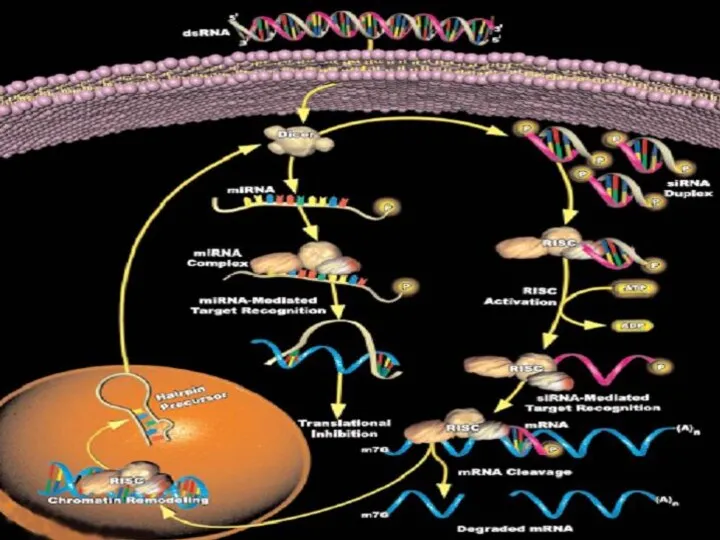

Dicer
Double-stranded RNA triggers processed into siRNAs by enzyme RNAseIII family,
Dicer
Double-stranded RNA triggers processed into siRNAs by enzyme RNAseIII family,

RISC complex
RISC is a large (~500-kDa) RNA-multiprotein complex, which
RISC complex
RISC is a large (~500-kDa) RNA-multiprotein complex, which

Different classes of small RNA molecules
During dsRNA cleavage, different RNA classes
Different classes of small RNA molecules
During dsRNA cleavage, different RNA classes

siRNAs
Small interfering RNAs that have an integral role in
the
siRNAs
Small interfering RNAs that have an integral role in
the

miRNAs/stRNAs
micro/small temporal RNAs derive from ~70 nt ssRNA (single-stranded RNA),
miRNAs/stRNAs
micro/small temporal RNAs derive from ~70 nt ssRNA (single-stranded RNA),

MEM
MEM
)
MEM
MEM
)

Why is PTGS important?
Most widely held view is that RNAi evolved
Why is PTGS important?
Most widely held view is that RNAi evolved

Recent applications of RNAi
Modulation of HIV-1 replication by RNA interference.
Hannon(2002).
Recent applications of RNAi
Modulation of HIV-1 replication by RNA interference.
Hannon(2002).

Possible future improvements of RNAi applications
Already developed:
in vitro synthesis of siRNAs
Possible future improvements of RNAi applications
Already developed:
in vitro synthesis of siRNAs

Conclusions
begun in worms, flies, and plants - as an accidental
Conclusions
begun in worms, flies, and plants - as an accidental
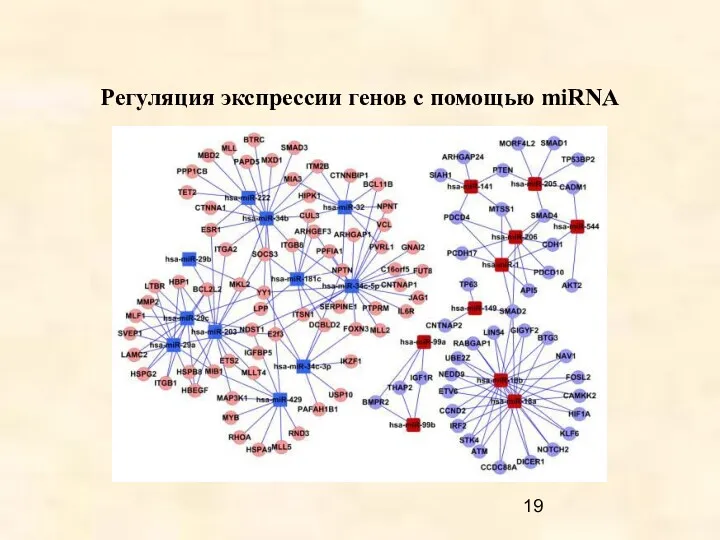
Регуляция экспрессии генов с помощью miRNA
Регуляция экспрессии генов с помощью miRNA

DNA-интерференция DNA-guided DNA interference by a prokaryotic Argonaute. Swarts DC, Jore

Функции siРНК
Сайленсинг мобильных генетических элементов;
Сайленсинг гетерохроматиновых повторов;
Сайленсинг генетического материала вирусного
Функции siРНК
Сайленсинг мобильных генетических элементов;
Сайленсинг гетерохроматиновых повторов;
Сайленсинг генетического материала вирусного

При выделение фракций коротких РНК (19-25 нуклеотидов) из различных организмов обнаружен
При выделение фракций коротких РНК (19-25 нуклеотидов) из различных организмов обнаружен

Функция miРНК
Обеспечивают сайленсинг различных генов, обычно, за счет частично комплементарного связывания
Функция miРНК
Обеспечивают сайленсинг различных генов, обычно, за счет частично комплементарного связывания

Продукт dsРНК, закодированных в уникальных генах геномов многоклеточных организмов (>1% от

созданы библиотеки коротких РНК и ДНК-векторов, кодирующих короткие РНК, мишенями
созданы библиотеки коротких РНК и ДНК-векторов, кодирующих короткие РНК, мишенями
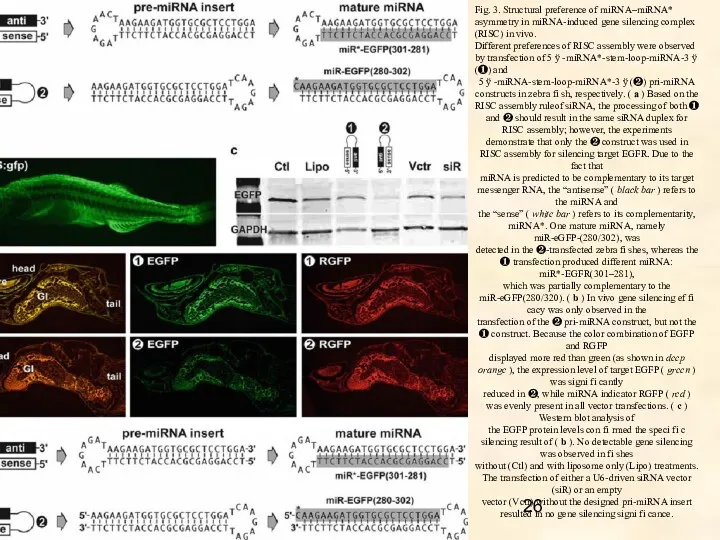
Fig. 3. Structural preference of miRNA–miRNA* asymmetry in miRNA-induced gene silencing
Fig. 3. Structural preference of miRNA–miRNA* asymmetry in miRNA-induced gene silencing
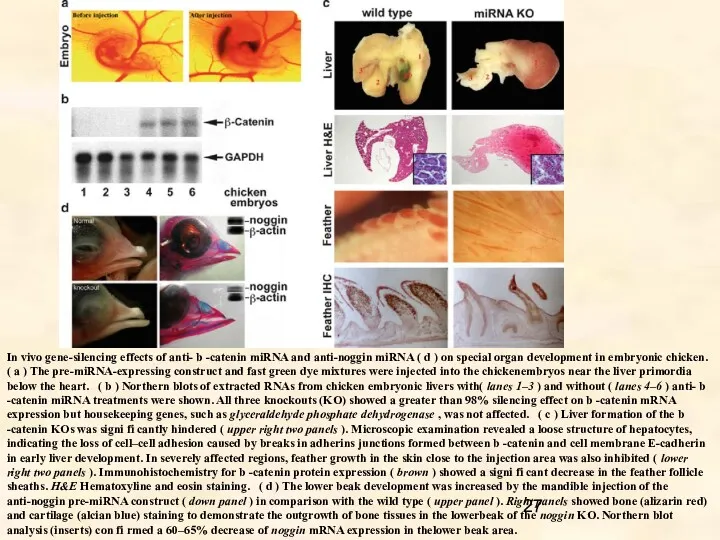
In vivo gene-silencing effects of anti- b -catenin miRNA and anti-noggin
In vivo gene-silencing effects of anti- b -catenin miRNA and anti-noggin
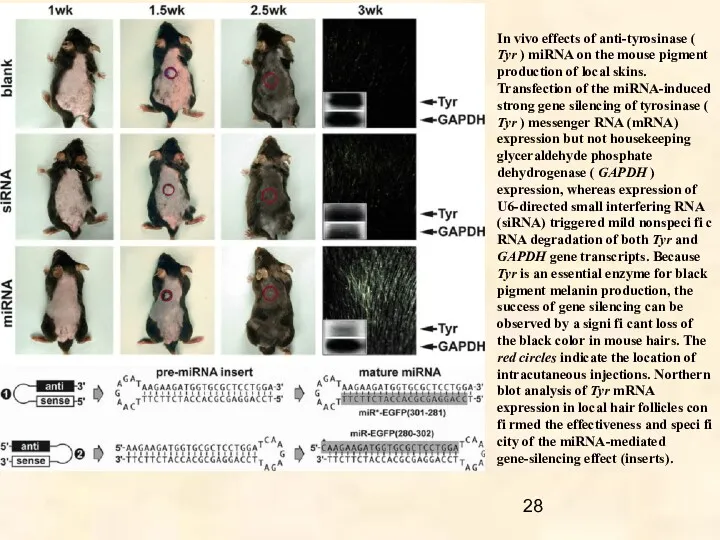
In vivo effects of anti-tyrosinase ( Tyr ) miRNA on the
In vivo effects of anti-tyrosinase ( Tyr ) miRNA on the
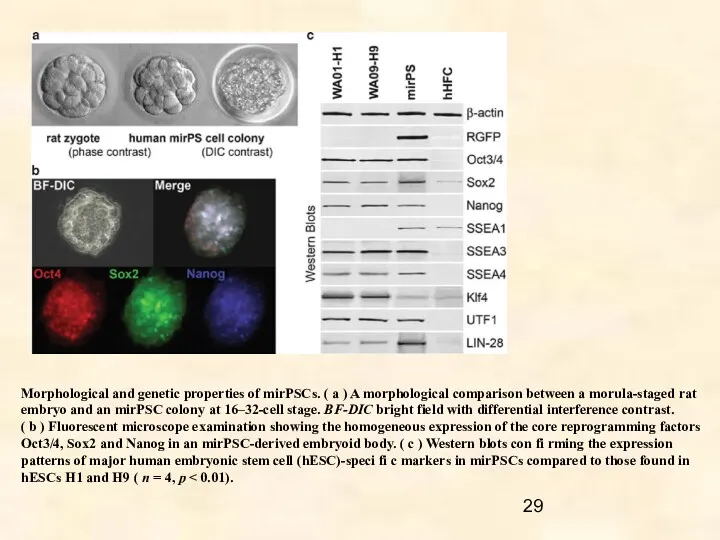
Morphological and genetic properties of mirPSCs. ( a ) A morphological
Morphological and genetic properties of mirPSCs. ( a ) A morphological
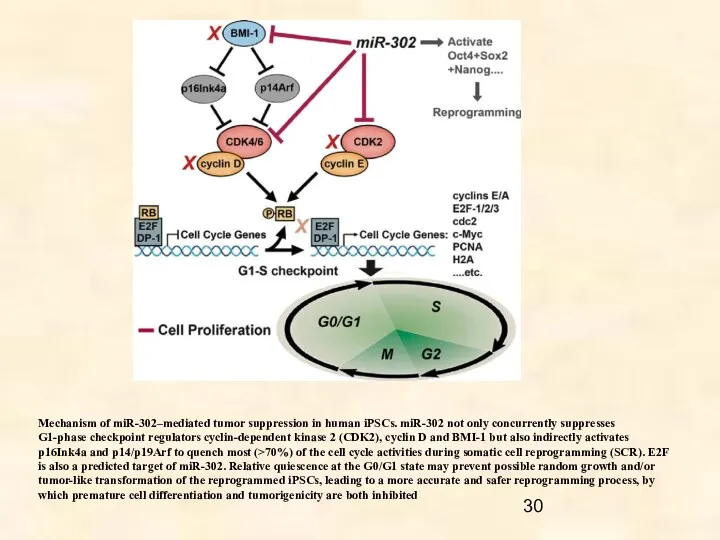
Mechanism of miR-302–mediated tumor suppression in human iPSCs. miR-302 not only
Mechanism of miR-302–mediated tumor suppression in human iPSCs. miR-302 not only

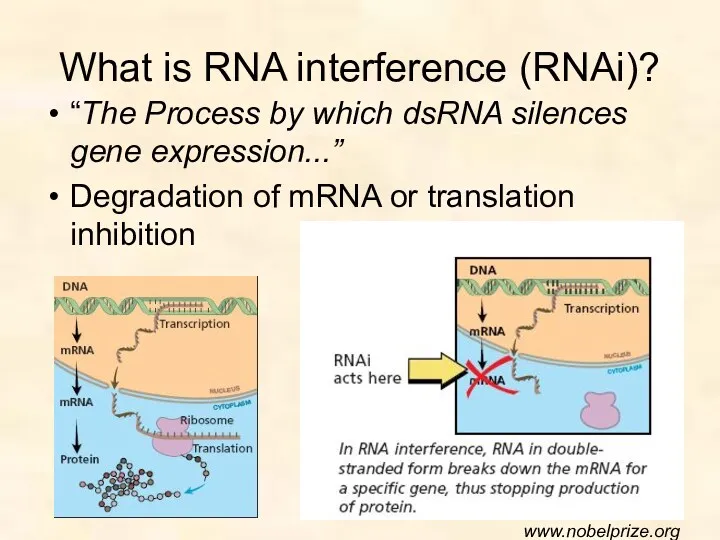
What is RNA interference (RNAi)?
“The Process by which dsRNA silences gene
What is RNA interference (RNAi)?
“The Process by which dsRNA silences gene
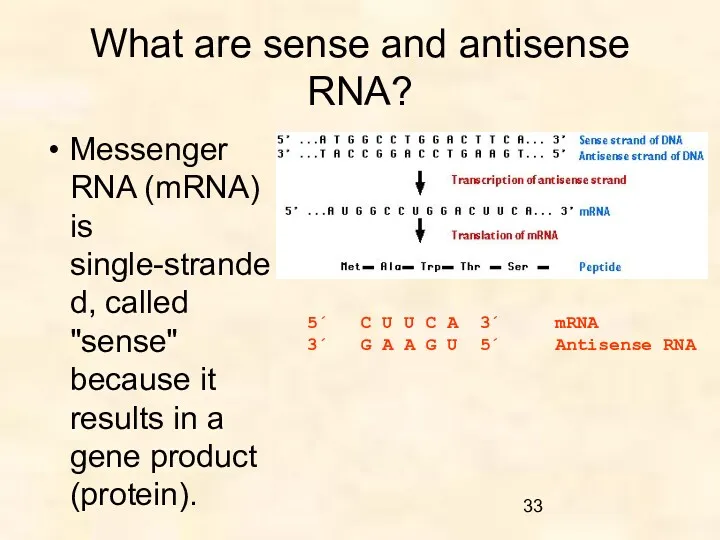
What are sense and antisense RNA?
Messenger RNA (mRNA) is single-stranded, called
What are sense and antisense RNA?
Messenger RNA (mRNA) is single-stranded, called
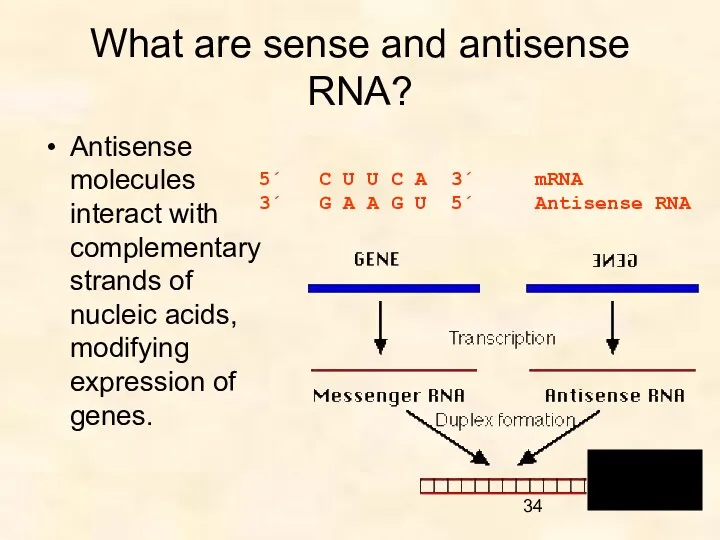
What are sense and antisense RNA?
Antisense molecules interact with complementary strands
What are sense and antisense RNA?
Antisense molecules interact with complementary strands

RNAi terms
dsRNA: double stranded RNA, longer than 30 nt
miRNA: microRNA, 21-25
RNAi terms
dsRNA: double stranded RNA, longer than 30 nt
miRNA: microRNA, 21-25

RNAi like phenomena
Plants
Petunias
Fungi
Neurospora
Animals
Caenorhabditis elegans
Alternate terms to RNAi
PTGS (Posttranscriptional Gene Silencing)
Cosuppression
Quelling
Virus-induced
Plants
Petunias
Fungi
Neurospora
Animals
Caenorhabditis elegans
Alternate terms to RNAi
PTGS (Posttranscriptional Gene Silencing)
Cosuppression
Quelling
Virus-induced

1990-Petunias
Napoli et al. defined an RNAi-like phenomenon and called it “cosupression.”
chalcone
1990-Petunias
Napoli et al. defined an RNAi-like phenomenon and called it “cosupression.”
chalcone

Overexpression of chalcone synthase in petunias unexpectedly resulted in white petunias
The
Overexpression of chalcone synthase in petunias unexpectedly resulted in white petunias
The

1992-The mold
Carlo Cogoni and Guiseppe Macino of the Università di Roma
1992-The mold
Carlo Cogoni and Guiseppe Macino of the Università di Roma
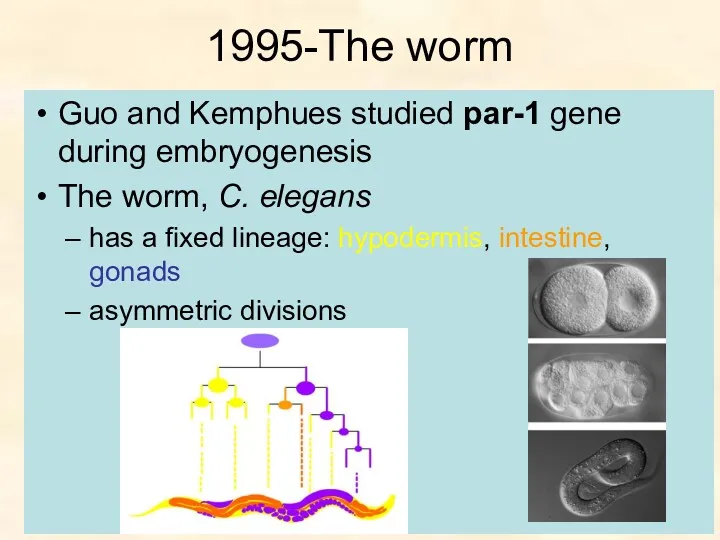
1995-The worm
Guo and Kemphues studied par-1 gene during embryogenesis
The worm, C.
1995-The worm
Guo and Kemphues studied par-1 gene during embryogenesis
The worm, C.
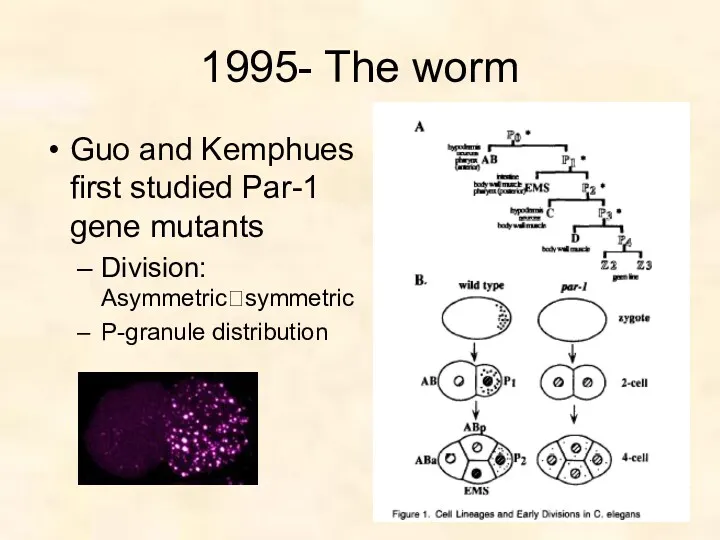
1995- The worm
Guo and Kemphues first studied Par-1 gene mutants
Division: Asymmetric?symmetric
P-granule
1995- The worm
Guo and Kemphues first studied Par-1 gene mutants
Division: Asymmetric?symmetric
P-granule

Guo and Kemphues, 1995
Guo and Kemphues, 1995
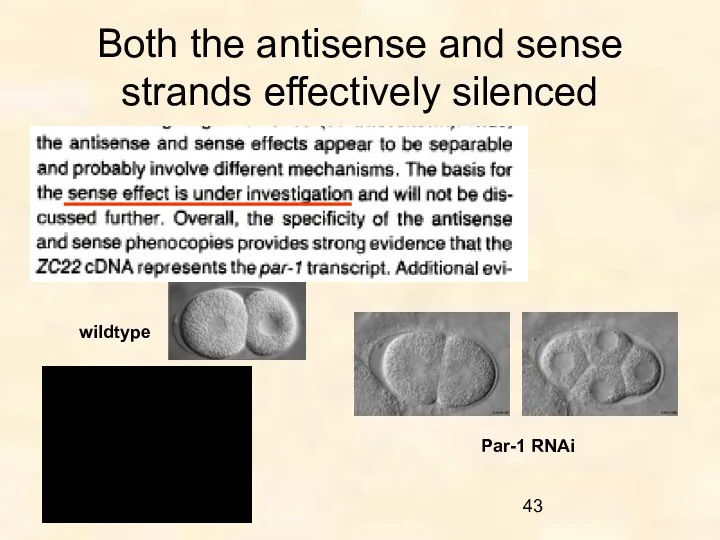
Both the antisense and sense strands effectively silenced
wildtype
Par-1 RNAi
Both the antisense and sense strands effectively silenced
wildtype
Par-1 RNAi

‘Antisense’ Technology?
Sense RNA silences yet no hybridization of sense RNA with
‘Antisense’ Technology?
Sense RNA silences yet no hybridization of sense RNA with

Craig Mello
In 1996, C. Mello and his student S. Driver also
Craig Mello
In 1996, C. Mello and his student S. Driver also

1998-Fire et al and Mello
Gel-purified ssRNA
Used purified ssRNA (antisense and
1998-Fire et al and Mello
Gel-purified ssRNA
Used purified ssRNA (antisense and

Unc-22 (Uncoordinated 22)
Codes for a non essential myofilament
It is present several
Unc-22 (Uncoordinated 22)
Codes for a non essential myofilament
It is present several
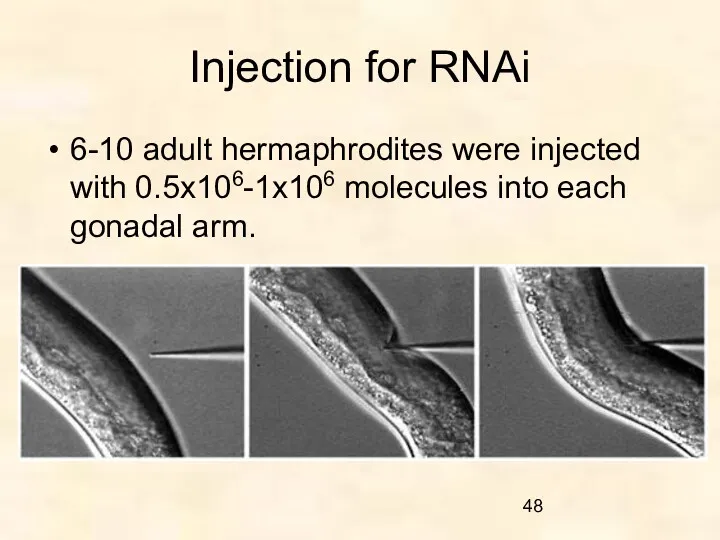
Injection for RNAi
6-10 adult hermaphrodites were injected with 0.5x106-1x106 molecules into
Injection for RNAi
6-10 adult hermaphrodites were injected with 0.5x106-1x106 molecules into
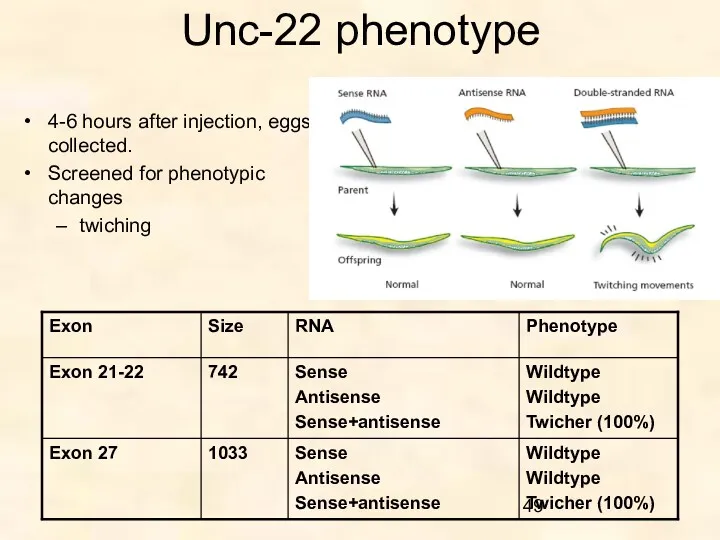
Unc-22 phenotype
4-6 hours after injection, eggs collected.
Screened for phenotypic changes
twiching
Unc-22 phenotype
4-6 hours after injection, eggs collected.
Screened for phenotypic changes
twiching

Mex-3
mex-3 encodes two RNA binding proteins; in the early embryo, maternally
Mex-3
mex-3 encodes two RNA binding proteins; in the early embryo, maternally
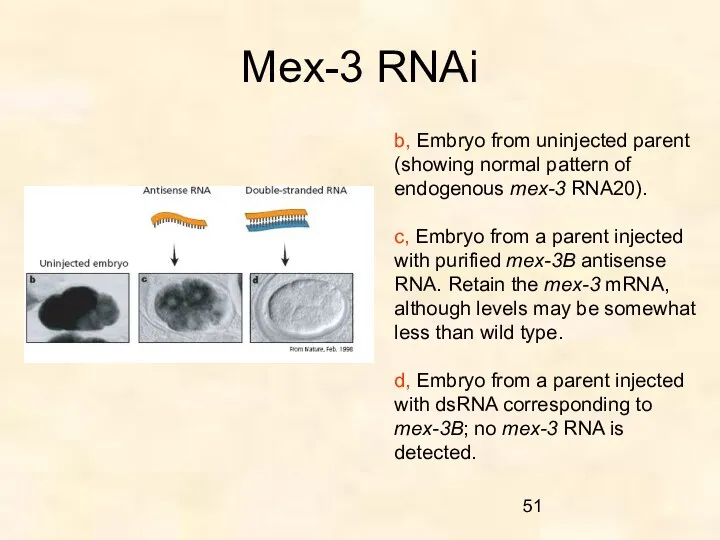
Mex-3 RNAi
b, Embryo from uninjected parent (showing normal pattern of endogenous
Mex-3 RNAi
b, Embryo from uninjected parent (showing normal pattern of endogenous

RNAi concentration and dose response
3.6x106 molecules/gonad
Sense phenocopied 1% of progeny
Antisense phenocopied
RNAi concentration and dose response
3.6x106 molecules/gonad
Sense phenocopied 1% of progeny
Antisense phenocopied
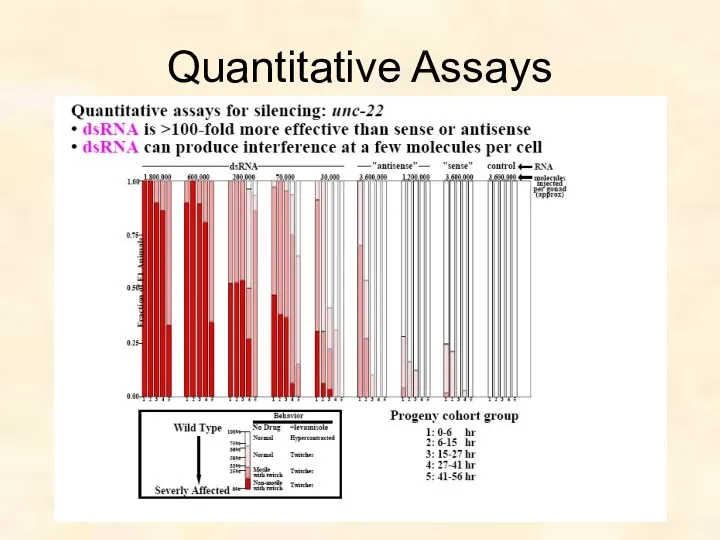
Quantitative Assays
Quantitative Assays

Other possibilities
Sense+antisense in low salt
Rapid sequential injection of sense & antisense
Both
Other possibilities
Sense+antisense in low salt
Rapid sequential injection of sense & antisense
Both

Conclusions
www.nobelprize.org
Conclusions
www.nobelprize.org

Conclusions
www.nobelprize.org
Conclusions
www.nobelprize.org
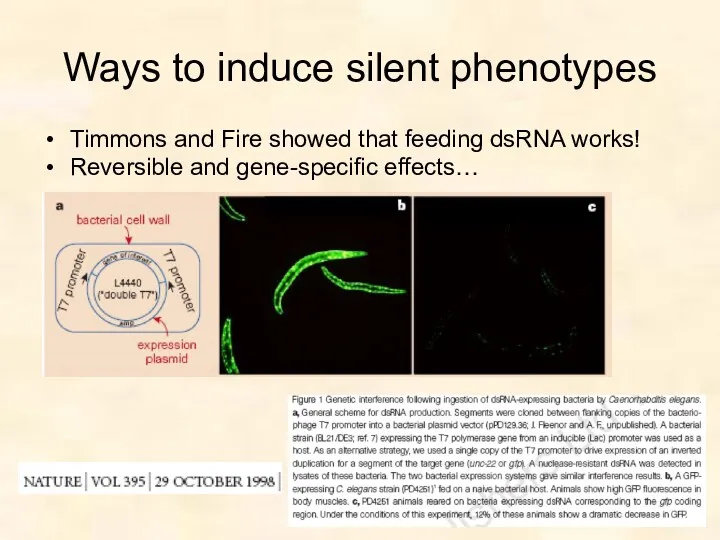
Ways to induce silent phenotypes
Timmons and Fire showed that feeding dsRNA
Ways to induce silent phenotypes
Timmons and Fire showed that feeding dsRNA
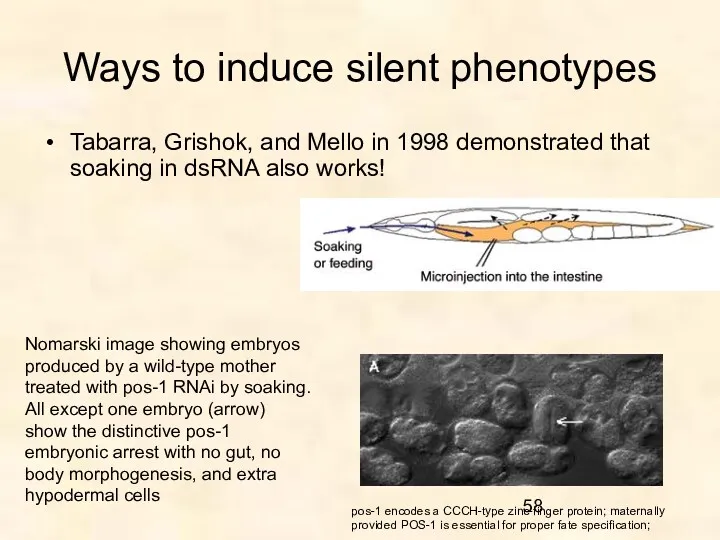
Ways to induce silent phenotypes
Tabarra, Grishok, and Mello in 1998 demonstrated
Ways to induce silent phenotypes
Tabarra, Grishok, and Mello in 1998 demonstrated

Mechanisms revealed
25bp species of dsRNA found in plants with co-suppression [Hamilton
Mechanisms revealed
25bp species of dsRNA found in plants with co-suppression [Hamilton

RNAi: two phases
Initiation
Generation of mature siRNA or miRNA
Execution
Silencing of target gene
Degradation
RNAi: two phases
Initiation
Generation of mature siRNA or miRNA
Execution
Silencing of target gene
Degradation
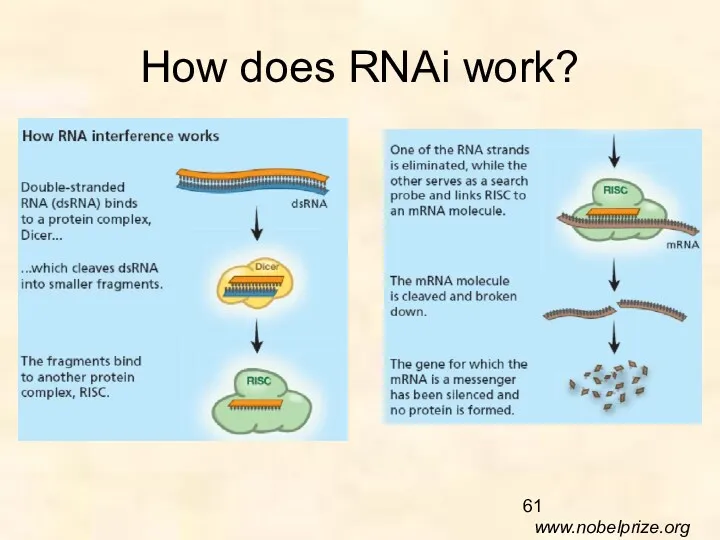
How does RNAi work?
www.nobelprize.org
How does RNAi work?
www.nobelprize.org
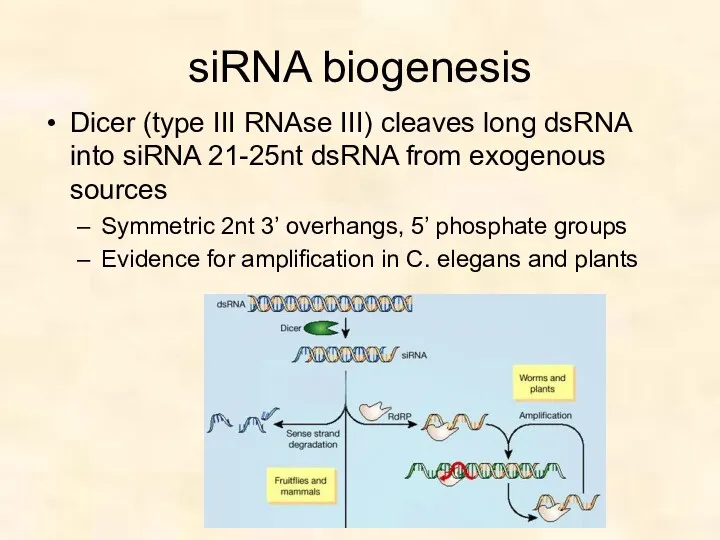
siRNA biogenesis
Dicer (type III RNAse III) cleaves long dsRNA into siRNA
siRNA biogenesis
Dicer (type III RNAse III) cleaves long dsRNA into siRNA

RNA Induced Silencing Complex (RISC)
RNAi effector complex
Preferentially incorporates one strand of
RNA Induced Silencing Complex (RISC)
RNAi effector complex
Preferentially incorporates one strand of
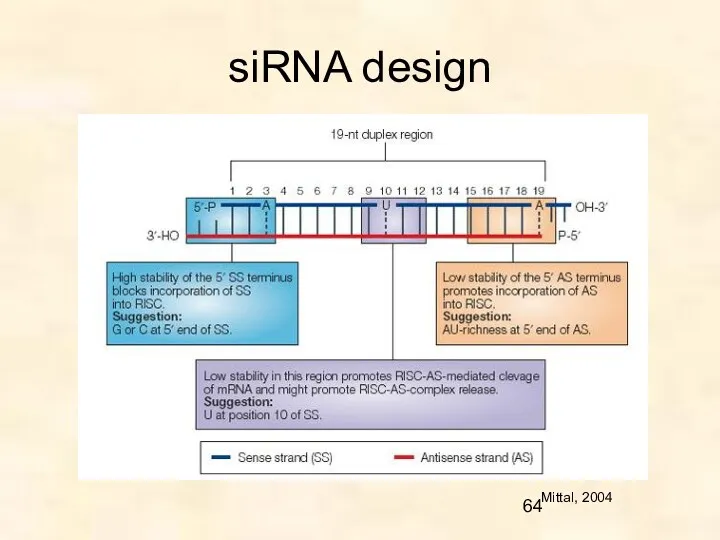
siRNA design
Mittal, 2004
siRNA design
Mittal, 2004
Custom-made siRNAs
Custom-made siRNAs
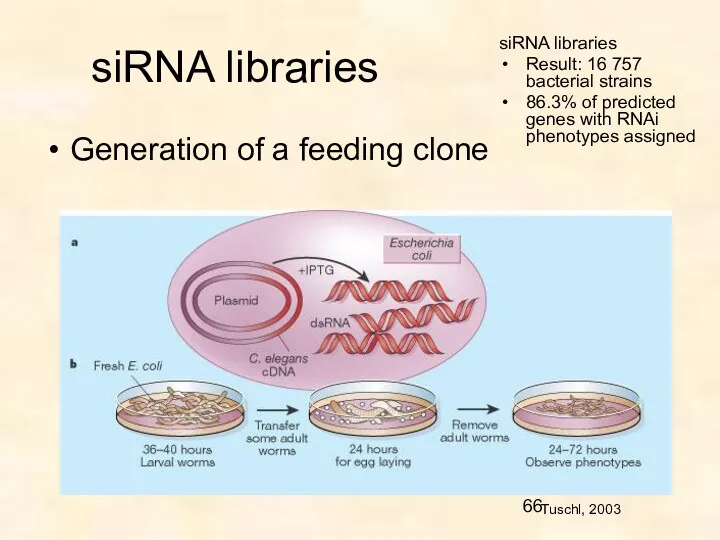
siRNA libraries
Generation of a feeding clone
Tuschl, 2003
siRNA libraries
Result: 16 757 bacterial
siRNA libraries
Generation of a feeding clone
Tuschl, 2003
siRNA libraries
Result: 16 757 bacterial
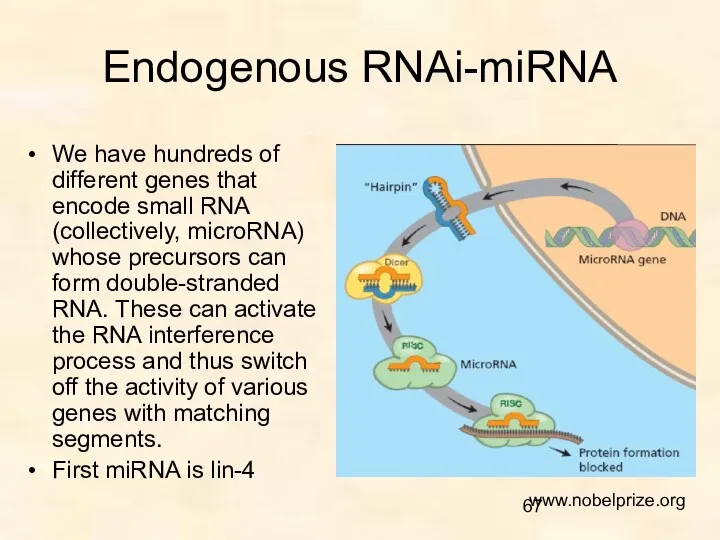
Endogenous RNAi-miRNA
We have hundreds of different genes that encode small RNA
Endogenous RNAi-miRNA
We have hundreds of different genes that encode small RNA

Defense Against Viruses
www.nobelprize.org
Indeed, Baulcombe, Vance, and others have shown that, in
Defense Against Viruses
www.nobelprize.org
Indeed, Baulcombe, Vance, and others have shown that, in
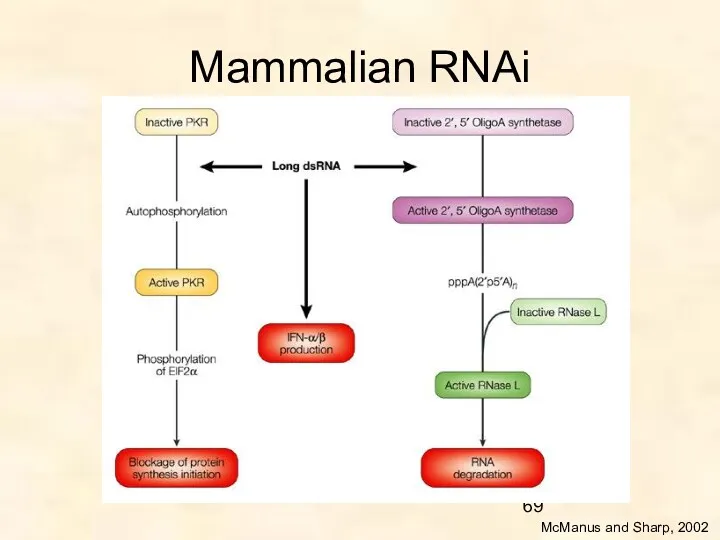
Mammalian RNAi
McManus and Sharp, 2002
Mammalian RNAi
McManus and Sharp, 2002

Getting Around the Problem
siRNA (21-22nt) mediate mammalian RNAi
Introducing siRNA instead of
Getting Around the Problem
siRNA (21-22nt) mediate mammalian RNAi
Introducing siRNA instead of

Some applications of RNAi
Therapy
Candidate genes, drug discovery, and therapy
Genome-wide RNAi screens
Gene
Some applications of RNAi
Therapy
Candidate genes, drug discovery, and therapy
Genome-wide RNAi screens
Gene
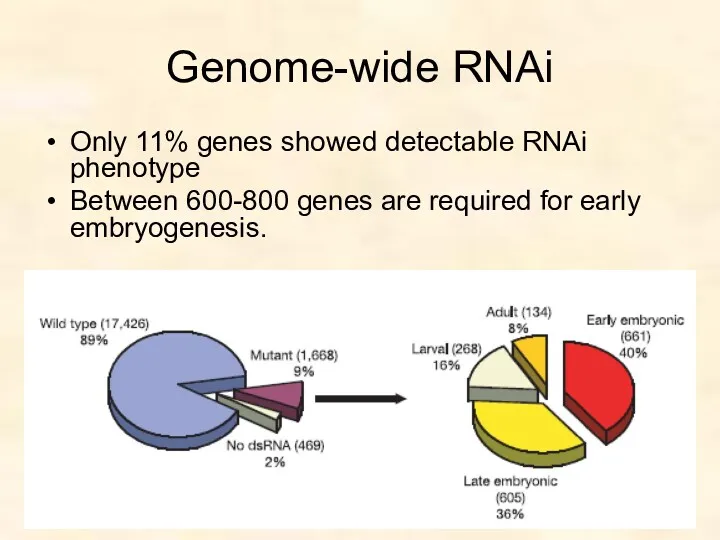
Genome-wide RNAi
Only 11% genes showed detectable RNAi phenotype
Between 600-800 genes are
Genome-wide RNAi
Only 11% genes showed detectable RNAi phenotype
Between 600-800 genes are

Systems Biology and RNAi
Cellular systems act as networks of interacting components
Systems Biology and RNAi
Cellular systems act as networks of interacting components

Networks of Early Embryogenesis
Protein-protein interaction dataset: binary physical interactions between 3,848
Networks of Early Embryogenesis
Protein-protein interaction dataset: binary physical interactions between 3,848
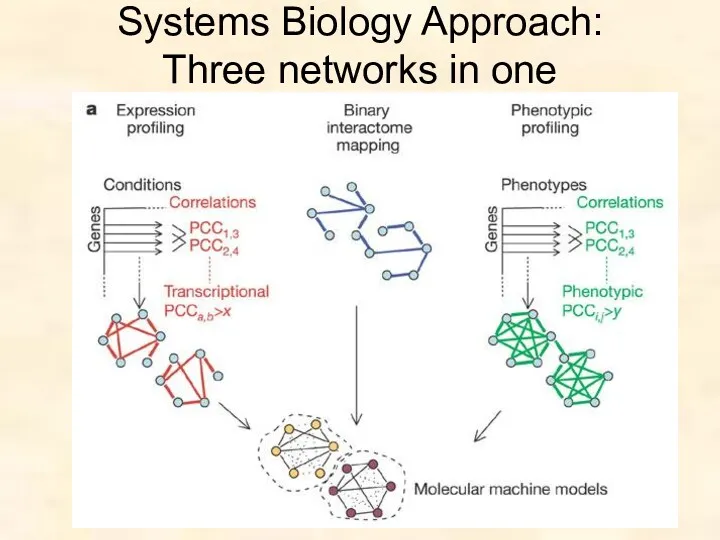
Systems Biology Approach:
Three networks in one
Systems Biology Approach:
Three networks in one
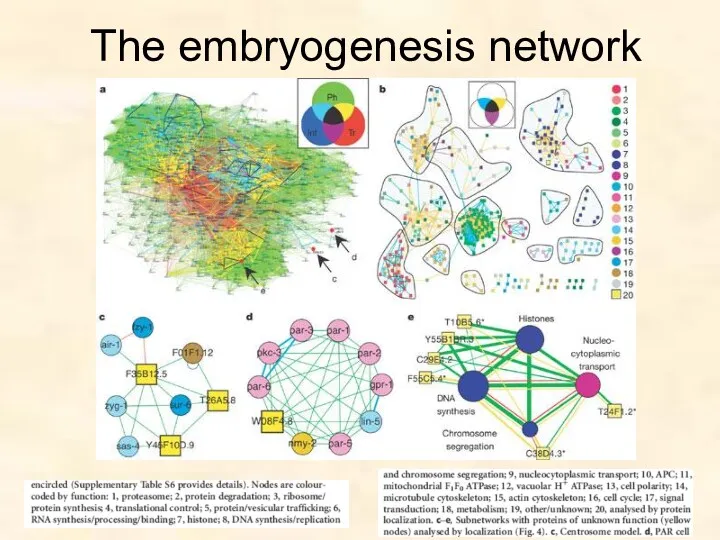
The embryogenesis network
The embryogenesis network
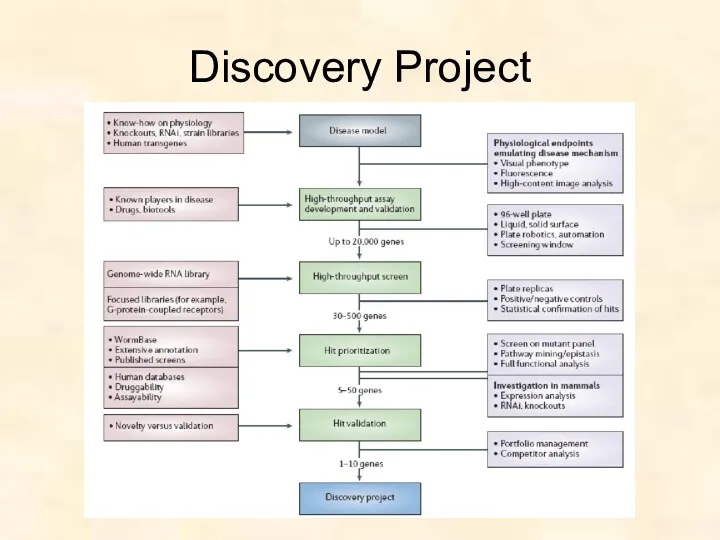
Discovery Project
Discovery Project

Defense against transposons
RNAi may also help keep the transposable elements that
Defense against transposons
RNAi may also help keep the transposable elements that


Why use RNAi?
1. The most powerful way to inhibit gene expression
Why use RNAi?
1. The most powerful way to inhibit gene expression

 Углеводы и липиды. Строение и функции
Углеводы и липиды. Строение и функции Приспособление организмов к среде обитания
Приспособление организмов к среде обитания Птица мандаринка
Птица мандаринка Хищные (лат. Carnivora — плотоядные)
Хищные (лат. Carnivora — плотоядные) Презентация по биологии для учащихся 7 класса по теме Класс Пресмыкающиеся
Презентация по биологии для учащихся 7 класса по теме Класс Пресмыкающиеся Імунітет. Імунна система
Імунітет. Імунна система Питательные среды для культивирования и их классификация
Питательные среды для культивирования и их классификация Эндоплазматическая сеть
Эндоплазматическая сеть Протерозойська ера
Протерозойська ера Вегетативті көбею
Вегетативті көбею Выращивание томатов
Выращивание томатов Клетки – маленькие лаборатории
Клетки – маленькие лаборатории Эволюция низших растений
Эволюция низших растений Бесполое размножение
Бесполое размножение Раннее начало половой жизни: за и против
Раннее начало половой жизни: за и против Уникальные грибы
Уникальные грибы Эволюция растений
Эволюция растений Нутрициология: белки и аминокислоты
Нутрициология: белки и аминокислоты Мышечные и нервная ткани
Мышечные и нервная ткани Гипотезы, объясняющие механизмы старения. Зависимость проявления старения от генотипа, условий и образа жизни
Гипотезы, объясняющие механизмы старения. Зависимость проявления старения от генотипа, условий и образа жизни Полухордовые и хордовые
Полухордовые и хордовые Многообразие форм живых органихмов
Многообразие форм живых органихмов Вопросы по биологии
Вопросы по биологии КИМ для подготовки учащихся 5 класса к ВПР по биологии. 1 вариант
КИМ для подготовки учащихся 5 класса к ВПР по биологии. 1 вариант Видовое разнообразие. Условия устойчивости экосистем
Видовое разнообразие. Условия устойчивости экосистем Красота окружающего мира. Насекомые
Красота окружающего мира. Насекомые Анатомо-физиологические особенности органа зрения
Анатомо-физиологические особенности органа зрения pril1
pril1