- Синапсы химические, электрические

Содержание
- 2. ЭЛЕКТРИЧЕСКИЙ СИНАПС
- 3. УЛЬТРАСТРУКТУРА НЕКСУСА (ЩЕЛЕВОГО КОНТАКТА)
- 4. ОСОБЕННОСТИ ЭЛЕКТРИЧЕСКИХ СИНАПСОВ КАК ПРАВИЛО – ВОЗБУЖДАЮЩИЕ КАК ПРАВИЛО – ОДНОСТОРОННИЕ ОТСУТСТВИЕ СИНАПТИЧЕСКОЙ ЗАДЕРЖКИ НЕЧУВСТВИТЕЛЬНОСТЬ К
- 5. НЕЙРОХИМИЯ
- 6. Otto Loewi
- 7. ТИПЫ СИНАПТИЧЕСКИХ СВЯЗЕЙ
- 8. ЭЛЕКТРОННАЯ МИКРОФОТОГРАФИЯ НЕРВНО-МЫШЕЧНОГО СОЕДИНЕНИЯ
- 9. СХЕМА ХИМИЧЕСКОГО СИНАПСА
- 10. СИНАПТИЧЕСКАЯ ЗАДЕРЖКА – ПРОМЕЖУТОК ВРЕМЕНИ МЕЖДУ МОМЕНТОМ ПОСТУПЛЕНИЕЯ ПД К ПРЕСИНАПТИЧЕСКОМУ ОКОНЧАНИЮ И НАЧАЛОМ СМЕЩЕНИЯ ЗАРЯДА
- 11. МИНИАТЮРНЫЕ ПОТЕНЦИАЛЫ КОНЦЕВОЙ ПЛАСТИНКИ (МПКП) В СОСТОЯНИИ ПОКОЯ
- 12. ПРИНЦИП ДЕЙЛА ВО ВСЕХ ПРЕСИНАПТИЧЕСКИХ ОКОНЧАНИЯХ ОДНОГО НЕЙРОНА ВЫСВОБОЖДАЕТСЯ ОДИН И ТОТ ЖЕ МЕДИАТОР ИСКЛЮЧЕНИЕ –
- 13. ЭТАПЫ ИДЕНТИФИКАЦИИ МЕДИАТОРА АНАТОМИЧЕСКИЙ – присутствие вещества в пресинаптических окончаниях БИОХИМИЧЕСКИЙ – присутствие ферментов, которые синтезируют
- 14. ЭТАПЫ ИДЕНТИФИКАЦИИ МЕДИАТОРА ФИЗИОЛОГИЧЕСКИЙ – демонстрация того, что физиологическая стимуляция заставляет пресинаптическое окончание выделять данное вещество
- 15. ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ В СИНАПСЕ
- 18. ЖИЗНЕННЫЙ ЦИКЛ СИНАПТИЧЕСКИХ ПУЗЫРЬКОВ
- 25. БИОСИНТЕЗ И ИНАКТИВАЦИЯ АЦЕТИЛХОЛИНА
- 26. РЕЦЕПТОРЫ К АЦЕТИЛХОЛИНУ
- 29. НЕРВНО-МЫШЕЧНЫЙ СИНАПС
- 30. БИОСИНТЕЗ КАТЕХОЛ-АМИНОВ ФМТ – ФЕНИЛЭТАНОЛАМИН – N -МЕТИЛТРАНСФЕРАЗА
- 36. БИОСИНТЕЗ И ИНАКТИВАЦИЯ СЕРОТОНИНА
- 39. АМИНОКИСЛОТЫ-МЕДИАТОРЫ
- 40. НЕЙРОПЕПТИДЫ
- 41. ХАРАКТЕРИСТИКА РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ В ЦНС Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением. Одностороннее
- 42. ПРОЦЕСС ТОРМОЖЕНИЯ В ЦНС Торможение – активный процесс, возникающий в нервной системе и приводящий к ослаблению
- 43. Стимул ВИДЫ ТОРМОЖЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
- 44. РОЛЬ ТОРМОЖЕНИЯ 1.Охранительная. 2.Важная роль в обработке поступающей в ЦНС информации. 3. Важный фактор обеспечения координационной
- 45. ОБЩАЯ ХАРАКТЕРИСТИКА МЫШЦ Виды мышц. Поперечно-полосатые (скелетная, сердечная) и гладкие мышцы (составляют около 50% массы тела).
- 46. Структурно-функциональная характеристика мышечных волокон Мышечное волокно – сократительная единица скелетной мышцы (диаметр 10 – 100 мкм,
- 47. актин миозин А-диск М 1/2 I-диска Z мышца расслаблена мышца сокращена Н-диск А-диск мостик Н ½
- 48. Регуляторная роль тропомиозина и тропонина В расслабленном состоянии тропонин частично блокирует сайты связывания с миозином, находящиеся
- 49. А – нексус межсаркоплазматической области вставочного диска В – участок бокового края волокна, где щель сообщается
- 50. ЭЛЕКТРОННАЯ МИКРОФОТОГРАФИЯ НЕРВНО-МЫШЕЧНОГО СОЕДИНЕНИЯ
- 51. Теория мышечного сокращения и расслабления Электромеханическое сопряжение – распространение ПД по сарколемме в Т- трубочке приводит
- 52. Двигательные единицы - совокупность мышечных волокон, иннервируемых разветвлениями аксона одного мотонейрона . Виды двигательных единиц –
- 53. ТИПЫ СОКРАЩЕНИЙ Изометрическое (изменяется тонус), изотоническое (изменяется длина) и ауксотоническое (изменяются длина и тонус) сокращения. Одиночное
- 54. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ Нервный центр – совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию
- 56. Скачать презентацию
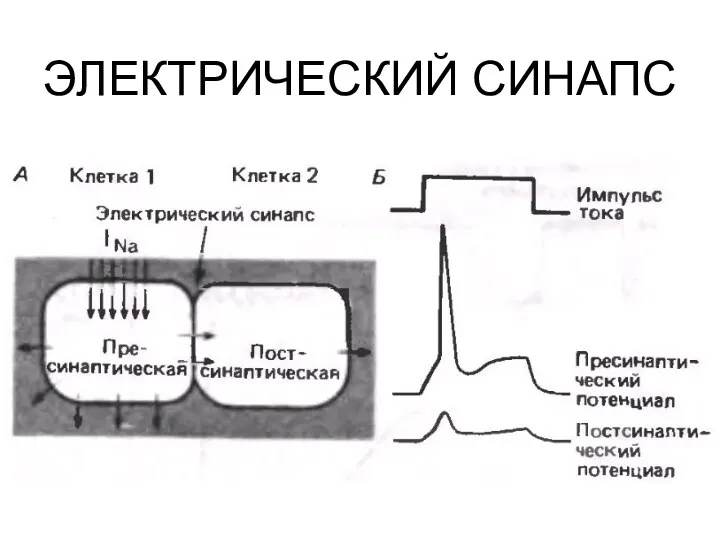
ЭЛЕКТРИЧЕСКИЙ СИНАПС
ЭЛЕКТРИЧЕСКИЙ СИНАПС
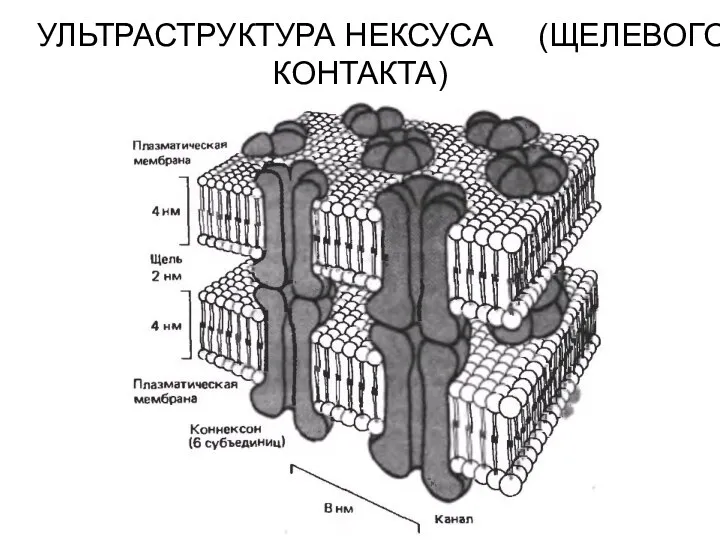
УЛЬТРАСТРУКТУРА НЕКСУСА (ЩЕЛЕВОГО КОНТАКТА)
УЛЬТРАСТРУКТУРА НЕКСУСА (ЩЕЛЕВОГО КОНТАКТА)

ОСОБЕННОСТИ ЭЛЕКТРИЧЕСКИХ СИНАПСОВ
КАК ПРАВИЛО – ВОЗБУЖДАЮЩИЕ
КАК ПРАВИЛО – ОДНОСТОРОННИЕ
ОСОБЕННОСТИ ЭЛЕКТРИЧЕСКИХ СИНАПСОВ
КАК ПРАВИЛО – ВОЗБУЖДАЮЩИЕ
КАК ПРАВИЛО – ОДНОСТОРОННИЕ
НЕЙРОХИМИЯ
НЕЙРОХИМИЯ
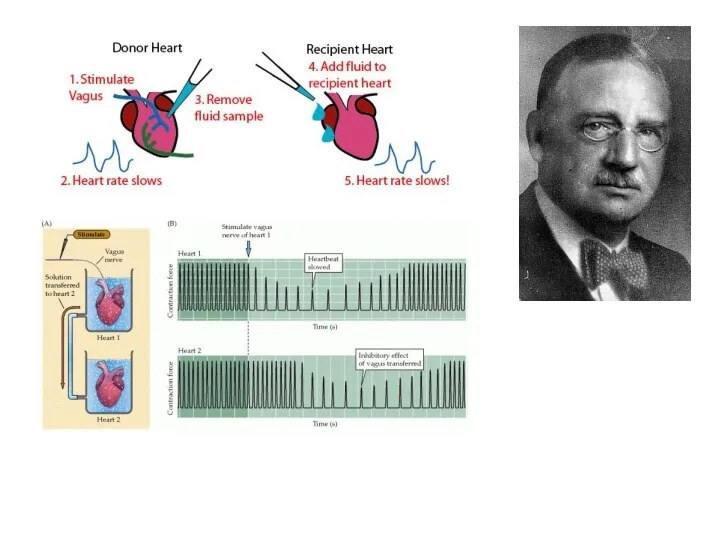
Otto Loewi
Otto Loewi
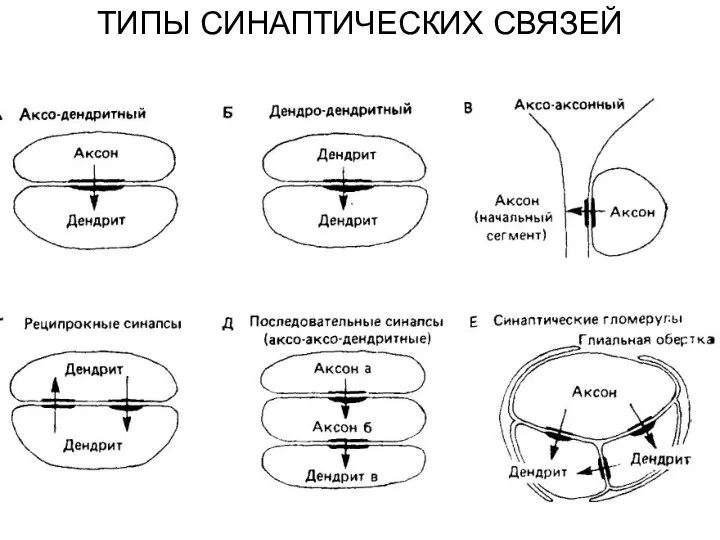
ТИПЫ СИНАПТИЧЕСКИХ СВЯЗЕЙ
ТИПЫ СИНАПТИЧЕСКИХ СВЯЗЕЙ
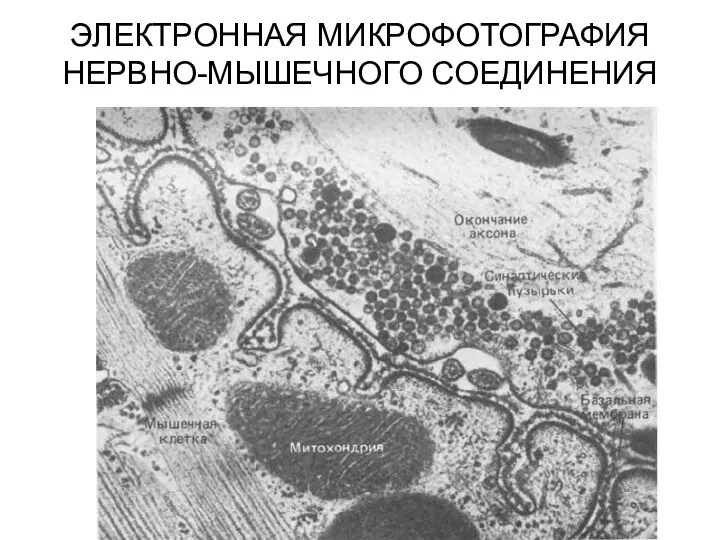
ЭЛЕКТРОННАЯ МИКРОФОТОГРАФИЯ НЕРВНО-МЫШЕЧНОГО СОЕДИНЕНИЯ
ЭЛЕКТРОННАЯ МИКРОФОТОГРАФИЯ НЕРВНО-МЫШЕЧНОГО СОЕДИНЕНИЯ
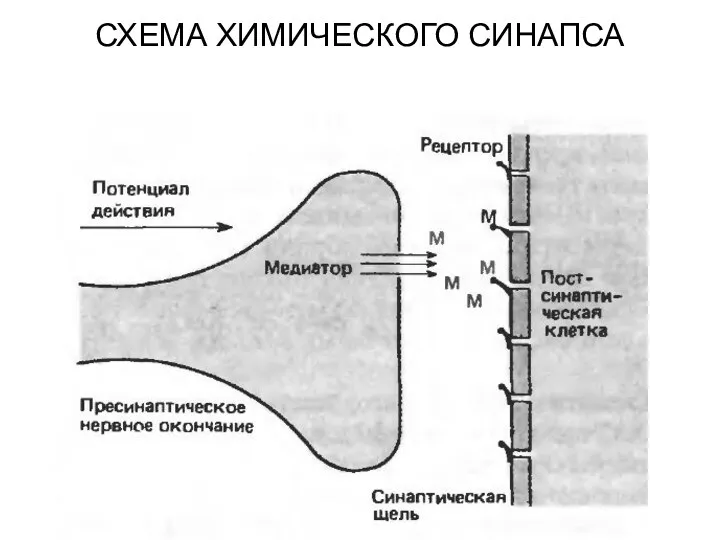
СХЕМА ХИМИЧЕСКОГО СИНАПСА
СХЕМА ХИМИЧЕСКОГО СИНАПСА

СИНАПТИЧЕСКАЯ ЗАДЕРЖКА – ПРОМЕЖУТОК ВРЕМЕНИ МЕЖДУ МОМЕНТОМ ПОСТУПЛЕНИЕЯ ПД К ПРЕСИНАПТИЧЕСКОМУ
СИНАПТИЧЕСКАЯ ЗАДЕРЖКА – ПРОМЕЖУТОК ВРЕМЕНИ МЕЖДУ МОМЕНТОМ ПОСТУПЛЕНИЕЯ ПД К ПРЕСИНАПТИЧЕСКОМУ
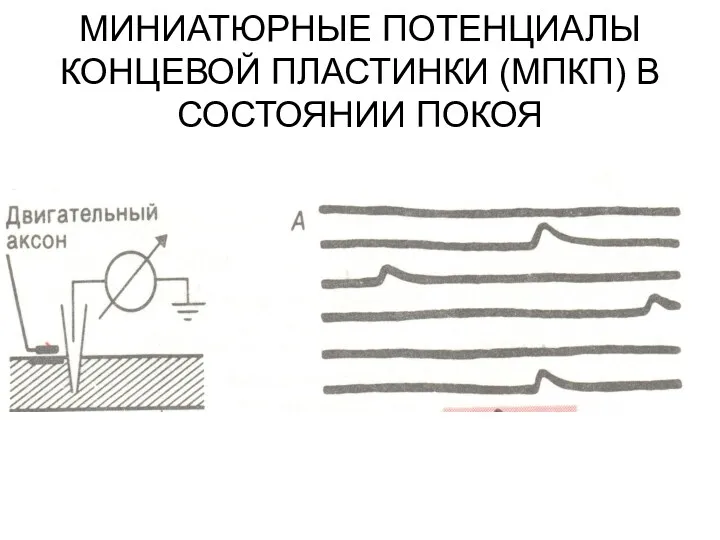
МИНИАТЮРНЫЕ ПОТЕНЦИАЛЫ КОНЦЕВОЙ ПЛАСТИНКИ (МПКП) В СОСТОЯНИИ ПОКОЯ
МИНИАТЮРНЫЕ ПОТЕНЦИАЛЫ КОНЦЕВОЙ ПЛАСТИНКИ (МПКП) В СОСТОЯНИИ ПОКОЯ

ПРИНЦИП ДЕЙЛА
ВО ВСЕХ ПРЕСИНАПТИЧЕСКИХ ОКОНЧАНИЯХ ОДНОГО НЕЙРОНА ВЫСВОБОЖДАЕТСЯ ОДИН И ТОТ
ПРИНЦИП ДЕЙЛА
ВО ВСЕХ ПРЕСИНАПТИЧЕСКИХ ОКОНЧАНИЯХ ОДНОГО НЕЙРОНА ВЫСВОБОЖДАЕТСЯ ОДИН И ТОТ

ЭТАПЫ ИДЕНТИФИКАЦИИ МЕДИАТОРА
АНАТОМИЧЕСКИЙ – присутствие вещества в пресинаптических окончаниях
БИОХИМИЧЕСКИЙ – присутствие
ЭТАПЫ ИДЕНТИФИКАЦИИ МЕДИАТОРА
АНАТОМИЧЕСКИЙ – присутствие вещества в пресинаптических окончаниях
БИОХИМИЧЕСКИЙ – присутствие

ЭТАПЫ ИДЕНТИФИКАЦИИ МЕДИАТОРА
ФИЗИОЛОГИЧЕСКИЙ – демонстрация того, что физиологическая стимуляция заставляет пресинаптическое
ЭТАПЫ ИДЕНТИФИКАЦИИ МЕДИАТОРА
ФИЗИОЛОГИЧЕСКИЙ – демонстрация того, что физиологическая стимуляция заставляет пресинаптическое
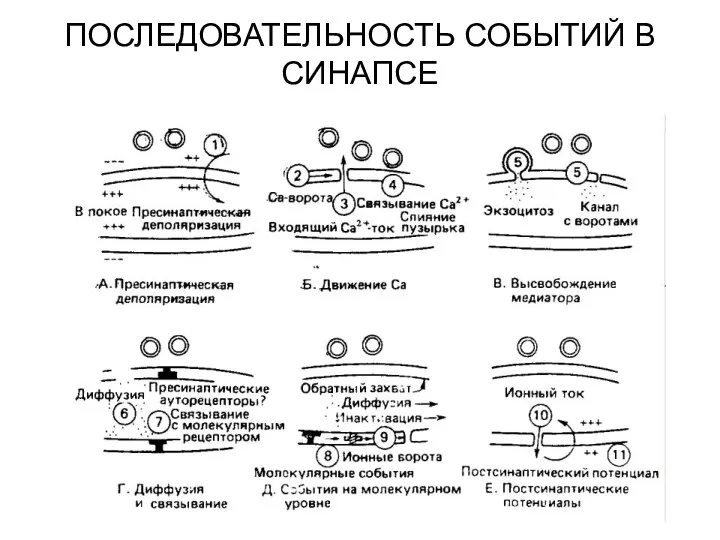
ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ В СИНАПСЕ
ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ В СИНАПСЕ

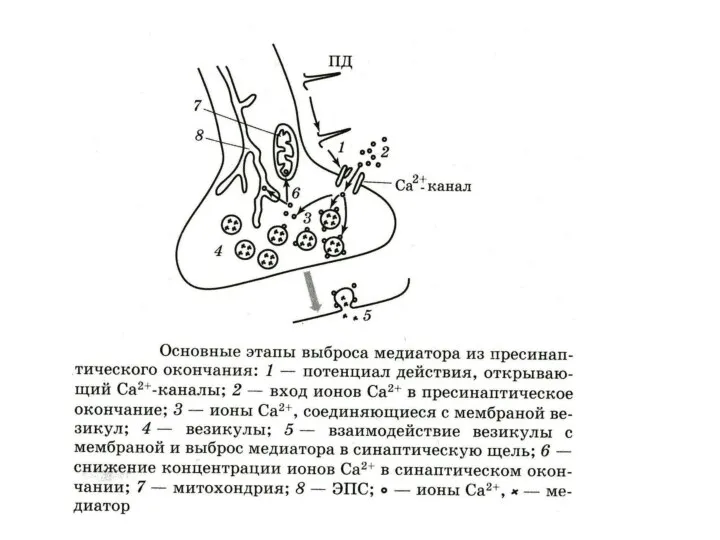
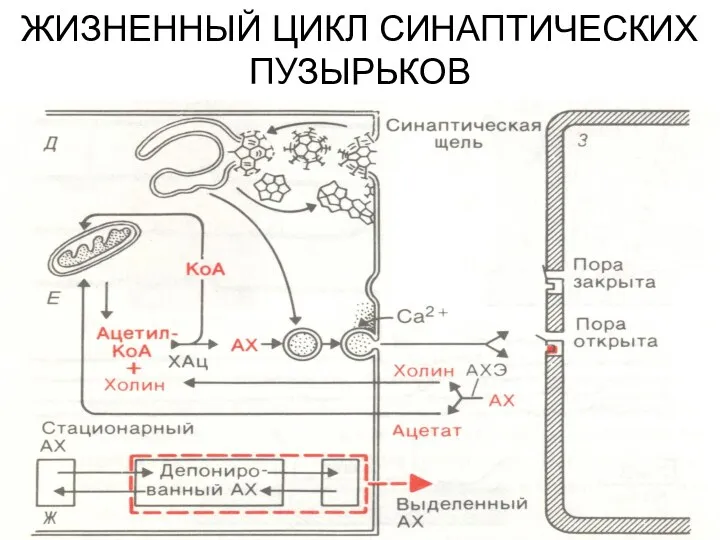
ЖИЗНЕННЫЙ ЦИКЛ СИНАПТИЧЕСКИХ ПУЗЫРЬКОВ
ЖИЗНЕННЫЙ ЦИКЛ СИНАПТИЧЕСКИХ ПУЗЫРЬКОВ





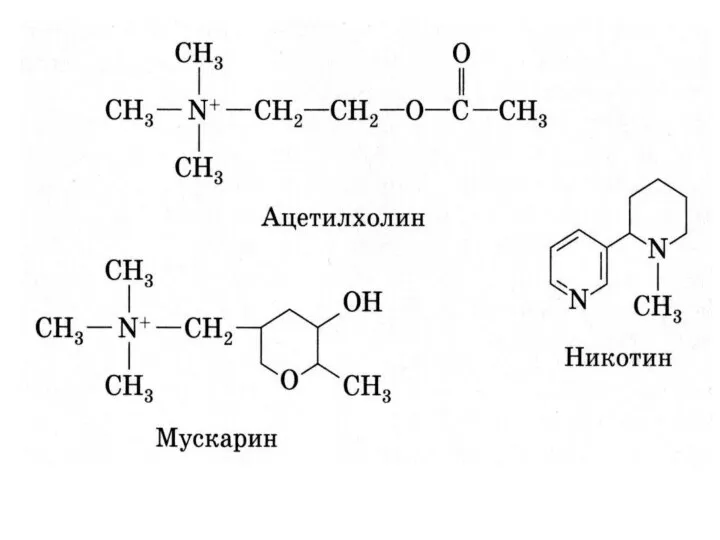
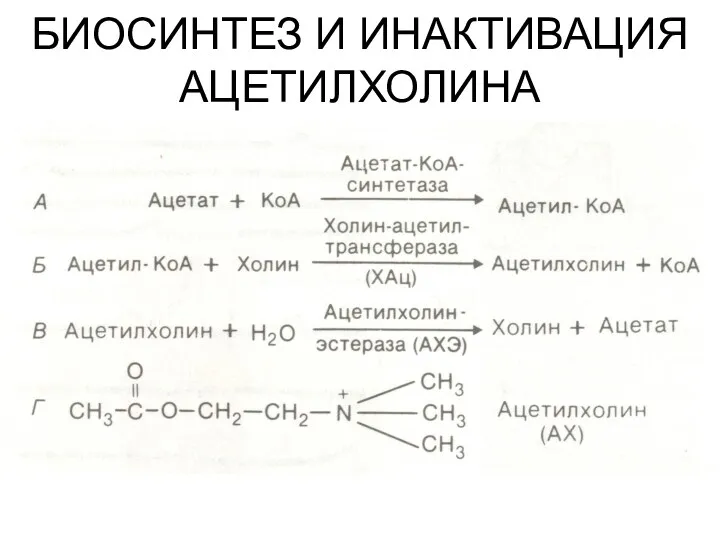
БИОСИНТЕЗ И ИНАКТИВАЦИЯ АЦЕТИЛХОЛИНА
БИОСИНТЕЗ И ИНАКТИВАЦИЯ АЦЕТИЛХОЛИНА
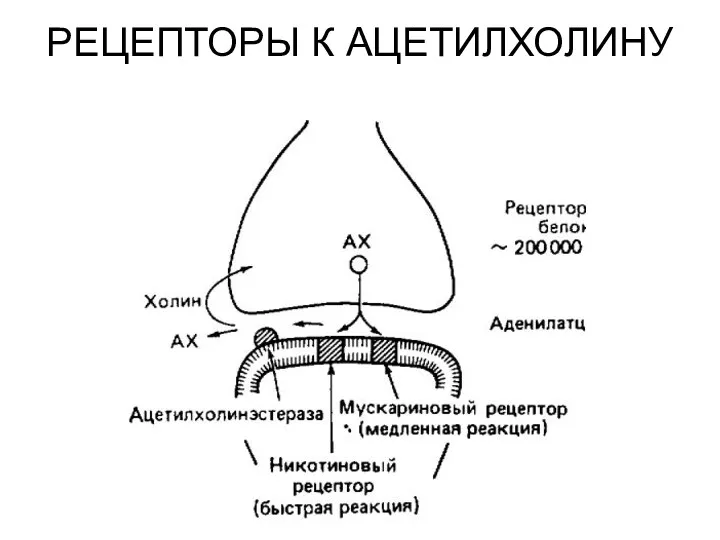
РЕЦЕПТОРЫ К АЦЕТИЛХОЛИНУ
РЕЦЕПТОРЫ К АЦЕТИЛХОЛИНУ

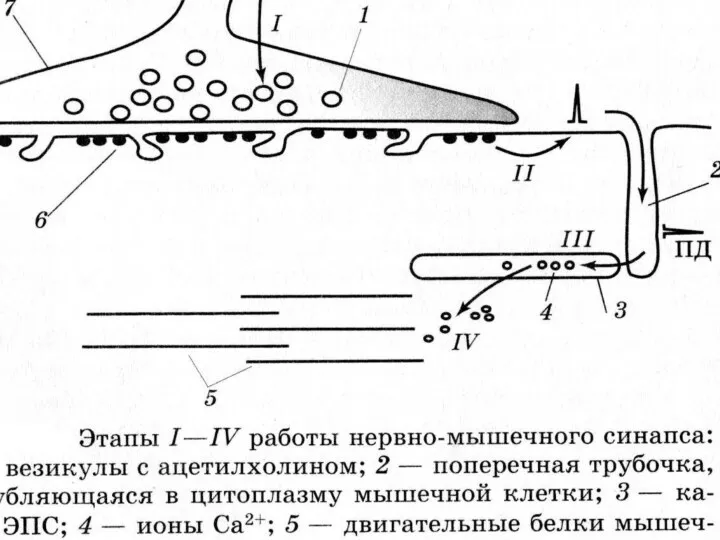
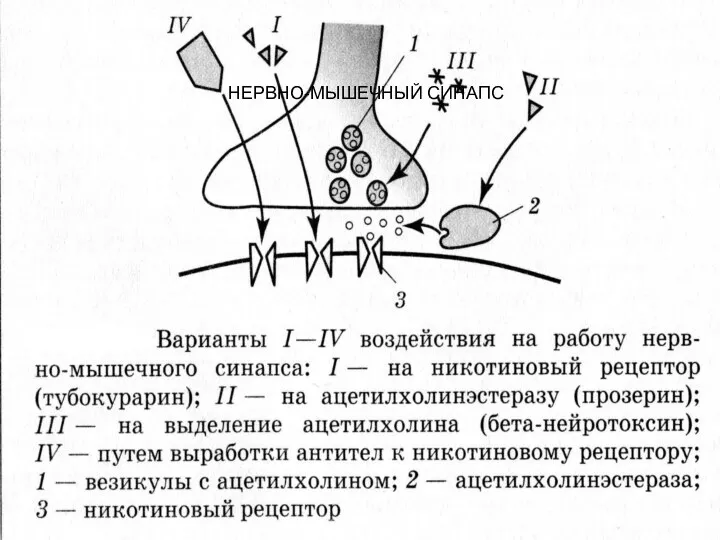
НЕРВНО-МЫШЕЧНЫЙ СИНАПС
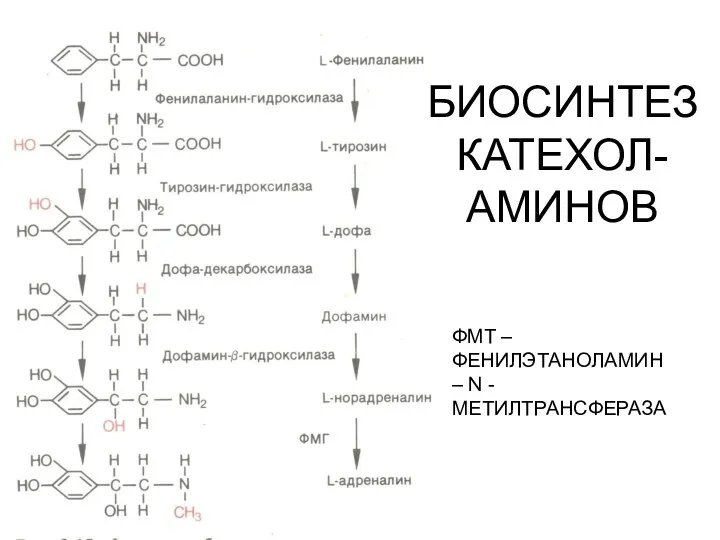
БИОСИНТЕЗ КАТЕХОЛ-АМИНОВ
ФМТ – ФЕНИЛЭТАНОЛАМИН – N -МЕТИЛТРАНСФЕРАЗА
БИОСИНТЕЗ КАТЕХОЛ-АМИНОВ
ФМТ – ФЕНИЛЭТАНОЛАМИН – N -МЕТИЛТРАНСФЕРАЗА



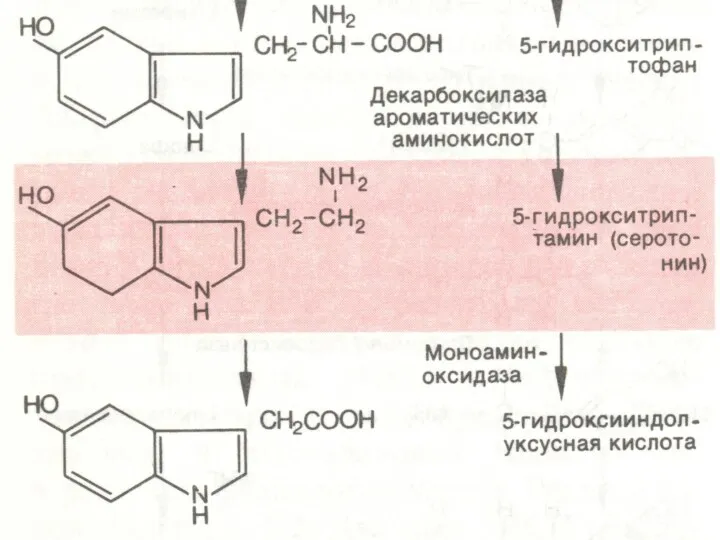
БИОСИНТЕЗ И ИНАКТИВАЦИЯ СЕРОТОНИНА
БИОСИНТЕЗ И ИНАКТИВАЦИЯ СЕРОТОНИНА


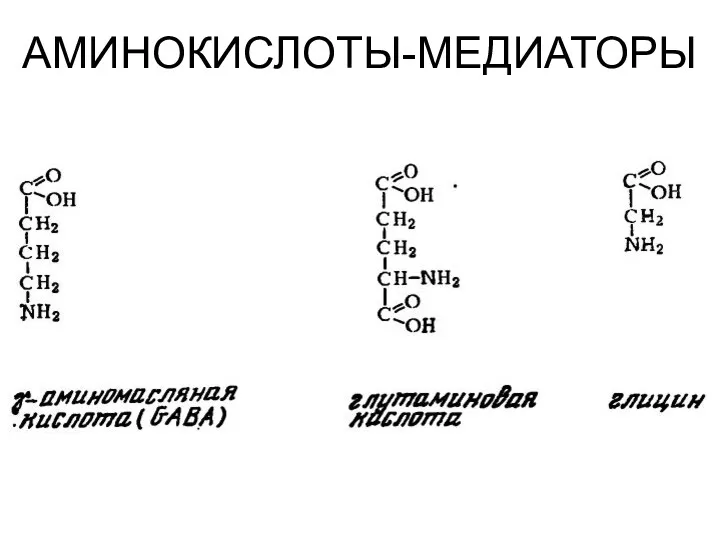
АМИНОКИСЛОТЫ-МЕДИАТОРЫ
АМИНОКИСЛОТЫ-МЕДИАТОРЫ

НЕЙРОПЕПТИДЫ
НЕЙРОПЕПТИДЫ

ХАРАКТЕРИСТИКА РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ В ЦНС
Все особенности распространения возбуждения в ЦНС объясняются
ХАРАКТЕРИСТИКА РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ В ЦНС
Все особенности распространения возбуждения в ЦНС объясняются

ПРОЦЕСС ТОРМОЖЕНИЯ В ЦНС
Торможение – активный процесс, возникающий в нервной системе
ПРОЦЕСС ТОРМОЖЕНИЯ В ЦНС
Торможение – активный процесс, возникающий в нервной системе
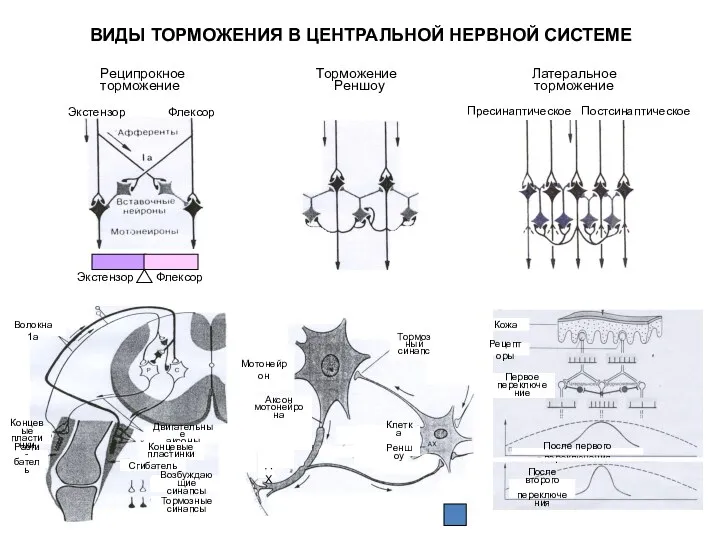
Стимул
ВИДЫ ТОРМОЖЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Стимул
ВИДЫ ТОРМОЖЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ

РОЛЬ ТОРМОЖЕНИЯ
1.Охранительная.
2.Важная роль в обработке поступающей в ЦНС информации.
3. Важный
РОЛЬ ТОРМОЖЕНИЯ
1.Охранительная.
2.Важная роль в обработке поступающей в ЦНС информации.
3. Важный

ОБЩАЯ ХАРАКТЕРИСТИКА МЫШЦ
Виды мышц. Поперечно-полосатые (скелетная, сердечная) и гладкие мышцы (составляют
ОБЩАЯ ХАРАКТЕРИСТИКА МЫШЦ
Виды мышц. Поперечно-полосатые (скелетная, сердечная) и гладкие мышцы (составляют

Структурно-функциональная характеристика мышечных волокон
Мышечное волокно – сократительная единица скелетной мышцы (диаметр
Структурно-функциональная характеристика мышечных волокон
Мышечное волокно – сократительная единица скелетной мышцы (диаметр
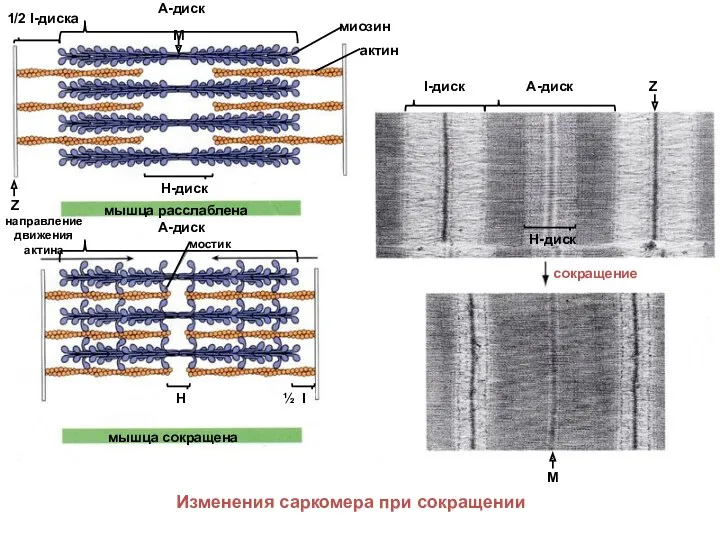
актин
миозин
А-диск
М
1/2 I-диска
Z
мышца расслаблена
мышца сокращена
Н-диск
А-диск
мостик
Н
½ I
направление движения актина
А-диск
I-диск
Z
М
Н-диск
сокращение
Изменения саркомера при сокращении
актин
миозин
А-диск
М
1/2 I-диска
Z
мышца расслаблена
мышца сокращена
Н-диск
А-диск
мостик
Н
½ I
направление движения актина
А-диск
I-диск
Z
М
Н-диск
сокращение
Изменения саркомера при сокращении
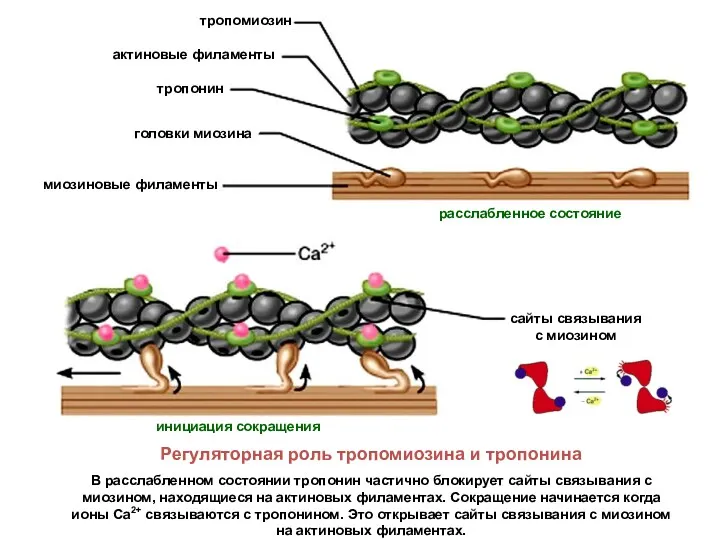
Регуляторная роль тропомиозина и тропонина
В расслабленном состоянии тропонин частично блокирует сайты
Регуляторная роль тропомиозина и тропонина
В расслабленном состоянии тропонин частично блокирует сайты
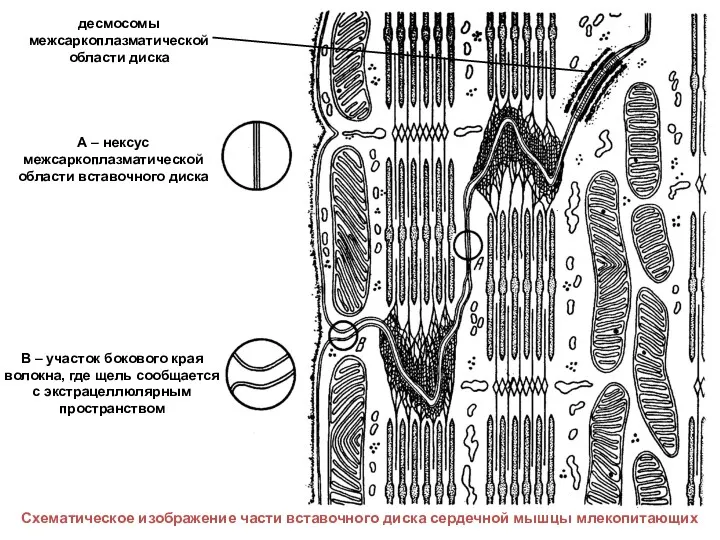
А – нексус межсаркоплазматической области вставочного диска
В – участок бокового края
А – нексус межсаркоплазматической области вставочного диска
В – участок бокового края
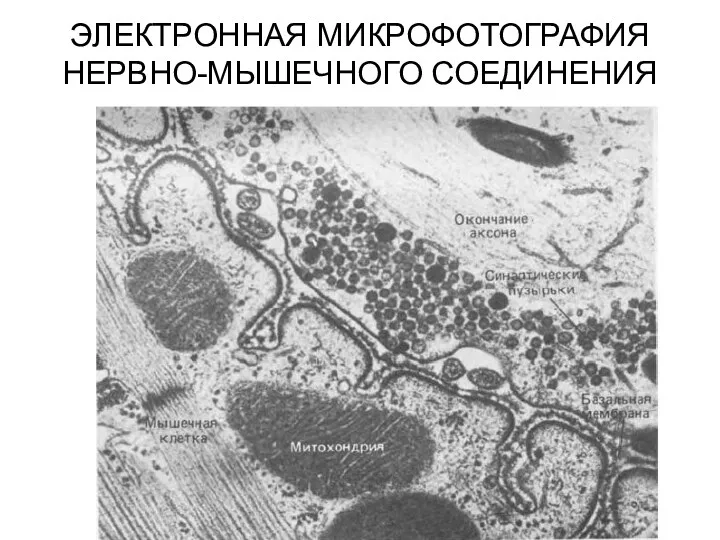
ЭЛЕКТРОННАЯ МИКРОФОТОГРАФИЯ НЕРВНО-МЫШЕЧНОГО СОЕДИНЕНИЯ
ЭЛЕКТРОННАЯ МИКРОФОТОГРАФИЯ НЕРВНО-МЫШЕЧНОГО СОЕДИНЕНИЯ

Теория мышечного сокращения и расслабления
Электромеханическое сопряжение – распространение ПД по сарколемме
Теория мышечного сокращения и расслабления
Электромеханическое сопряжение – распространение ПД по сарколемме

Двигательные единицы - совокупность мышечных волокон, иннервируемых разветвлениями аксона одного мотонейрона
Двигательные единицы - совокупность мышечных волокон, иннервируемых разветвлениями аксона одного мотонейрона

ТИПЫ СОКРАЩЕНИЙ
Изометрическое (изменяется тонус), изотоническое (изменяется длина) и ауксотоническое (изменяются длина
ТИПЫ СОКРАЩЕНИЙ
Изометрическое (изменяется тонус), изотоническое (изменяется длина) и ауксотоническое (изменяются длина

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Нервный центр – совокупность структур центральной нервной системы, координированная
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Нервный центр – совокупность структур центральной нервной системы, координированная
 Модификационная изменчивость
Модификационная изменчивость Методы исследования микроорганизмов и использование бактерий в биоиндикации
Методы исследования микроорганизмов и использование бактерий в биоиндикации Динамика содержания подвижного фосфора в серых лесных почвах Тюменской области
Динамика содержания подвижного фосфора в серых лесных почвах Тюменской области Митохондрия. Митохондрияның биологиялық рөлі
Митохондрия. Митохондрияның биологиялық рөлі Скелет. Строение, состав и соединение костей скелета человека
Скелет. Строение, состав и соединение костей скелета человека Вид. Популяция
Вид. Популяция Строение и функции кожи
Строение и функции кожи Ноосфера
Ноосфера Зелёная аптека. Экологическая игра
Зелёная аптека. Экологическая игра Анимированный кроссворд Строение клетки
Анимированный кроссворд Строение клетки Технологическое значение анатомических особенностей зерна
Технологическое значение анатомических особенностей зерна 20231016_prezentatsiya_2
20231016_prezentatsiya_2 Вирусы. Сходства вируса с живыми организмами. Размножение вирусов
Вирусы. Сходства вируса с живыми организмами. Размножение вирусов Биохимия печени. (Лекция 31)
Биохимия печени. (Лекция 31) Роль соли для организма человека
Роль соли для организма человека Моллюски
Моллюски Предмет и задачи микробиологии
Предмет и задачи микробиологии Неорганические вещества клетки (10 класс)
Неорганические вещества клетки (10 класс) Разнообразие животных. Урок 19
Разнообразие животных. Урок 19 Устный журнал: Зеленый наряд нашей планеты
Устный журнал: Зеленый наряд нашей планеты Уровни организации жизни
Уровни организации жизни Птицы из Красной книги Югры
Птицы из Красной книги Югры Өсімдіктер әлемі
Өсімдіктер әлемі Вопросы к викторине по охране природы
Вопросы к викторине по охране природы Личностно-ориентированная технология на уроках биологии
Личностно-ориентированная технология на уроках биологии Интегрированный урок биологии и химии с применением информационных технологий
Интегрированный урок биологии и химии с применением информационных технологий Забота о потомстве
Забота о потомстве Corynebacterium Erysipelothrix & Listeria
Corynebacterium Erysipelothrix & Listeria