- Слуховий і вестибулярний аналізатори

Содержание
- 2. Звук Звукові сигнали можна уявити як коливальні рухи повітря з різною частотою і силою. Вони збуджують
- 3. Схема відділів вуха Вухо людини здатне сприймати звук при коливанні повітря в діапазоні від 16 до
- 4. Зовнішнє вухо Зовнішнє вухо вловлює і проводить звукові коливання до барабанної перетинки.
- 5. Схема поширення звукової хвилі Звукові коливання сприймаються барабанною перетинкою і за допомогою системи кісточок передаються на
- 6. Среднє вухо Слухові кісточки: молоточок, коваделко і стремінце передають і підсилюють коливання звукової хвилі на мембрані
- 8. Внутрішнє вухо Внутрішнє вухо включає присінок, півколові канали і завитку. Завитка це кістковий спіральний канал, який
- 9. Кортіїв орган На базилярній мембрані розташовується кортіїв орган. Рецепторні клітини слуху відносяться до вторинночутливих клітин двох
- 10. Волоскові клітини Внутрішні волоскові клітини утворюють до 90% волокон слухового нерва і 10% зовнішні волоскові клітини.
- 11. Вестибулярна і базилярна мембрани поділяють кістковий канал завитки на три канали: Верхній - scala vestibuli, він
- 12. Ендо- и перилімфа внутрішнього вуха Простір середніх сходів заповнений ендолімфою. Простір верхнього і нижнього каналів заповнений
- 13. Механізм передачі коливань ендолімфи на рецепторні клітини кортіївого органу Хвиля, яка виникла в ендолімфі приводить в
- 14. Амплітудний максимум Основний механізм розрізнення висоти тонів обумовлений тим, що хвиля коливань молекул повітря, передаючись на
- 15. Частотний (амплітудний) максимум Схеми механізмів передачі звукової хвилі по структурам кортіївого органу: А - поширення звукової
- 16. Визначення гучності Діапазон амплітуди коливань ендолімфи пов'язаний з амплітудою коливання мембран. В результаті з ростом амплітуди
- 17. Сильний звук і реакція м'язів середнього вуха Сильний звук може викликати пошкодження барабанної перетинки. Тому для
- 18. Предвокалізаційний рефлекс Вберегти вухо людини від пошкоджуючої дії гучних звуків може допомогти предвокалізацйний рефлекс. Коли людина
- 20. Аналіз звукових коливань в ЦНС Закодована інформація міститься в звуковому стимулі у вигляді нейронного збудження, пройшовши
- 21. Обробка звукової інформації в ЦНС Дорсальний кохлеарний тракт переходить на протилежну сторону і закінчується в ядрі
- 22. Центри кори При одночасному збудженні нейронів підкіркових центрів і скроневих відділів кори формуються процеси, що забезпечують
- 23. Центри кори та їх зв’язки Від скроневої слухової ділянки кори відходять і низхідні шляхи до всіх
- 24. Слухова орієнтація в просторі Слухова орієнтація в просторі можлива лише при бінауральному слусі. Враховуючи те, що
- 25. Людина і тварини мають просторовий слух, тобто здатні визначати положення джерела звуку в просторі. Якщо джерело
- 26. Вестибулярний аналізатор Вестибулярний апарат - орган рівноваги, що дозволяє визначати положення і переміщення тіла в просторі.
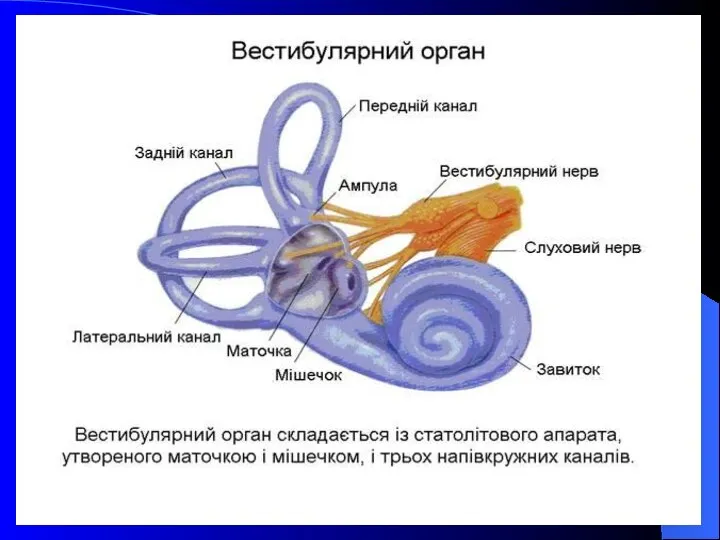
- 27. Вестибулярний орган складається з: Статолітового апарату Півколових каналів Маточки і мішечка Завитки Завиткового нерва Вестибулярного нерва
- 29. 1) отоліти; 2) отолітова мембрана; 3) волоски рецепторних клітин; 4) рецепторні клітини; 5) опорні клітини; 6)
- 30. Механізм збуждення рецепторних клітин Рецепторна клітина має пейсмекерні властивості (200 імп/с). Рух ендолімфи зрушує стероцилії в
- 31. Маточка і мішечок Рецептори маточки і мішечка контролюють прямолінійні рухи людини. Подразнюються вони при зміні швидкості
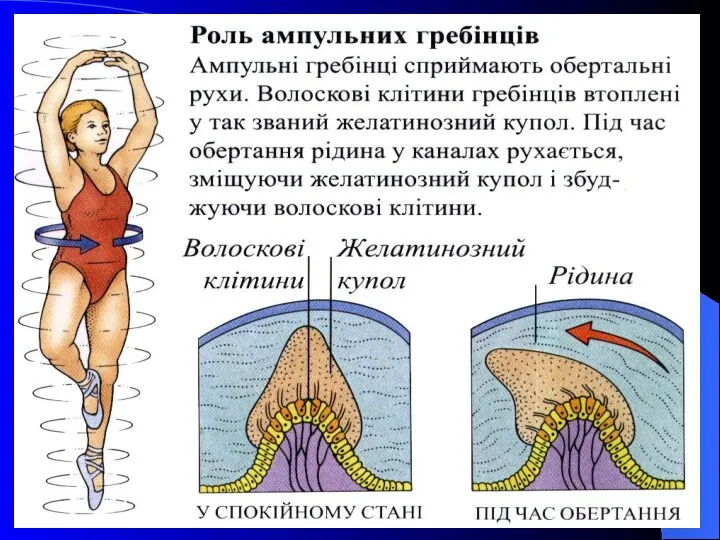
- 32. Півколові канали Природним стимулом для рецепторів півколових каналів є кутові прискорення. Війки їх знаходяться в желатиноподібній
- 33. Півколові канали розташовані у трьох взаємно перпендикулярних площинах. На одному зі своїх кінців кожен півколовий канал
- 34. Рецептори маточки реагують на зміну швидкості горизонтального руху, а мішечка - вертикального (ліфтні рефлекси) Механізм виникнення
- 37. Скачать презентацию

Звук
Звукові сигнали можна уявити як коливальні рухи повітря з різною частотою
Звук
Звукові сигнали можна уявити як коливальні рухи повітря з різною частотою
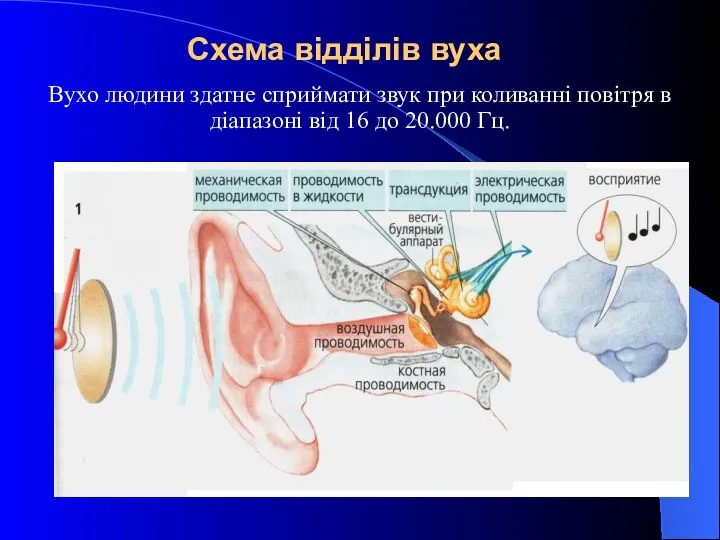
Схема відділів вуха
Вухо людини здатне сприймати звук при коливанні повітря в
Схема відділів вуха
Вухо людини здатне сприймати звук при коливанні повітря в
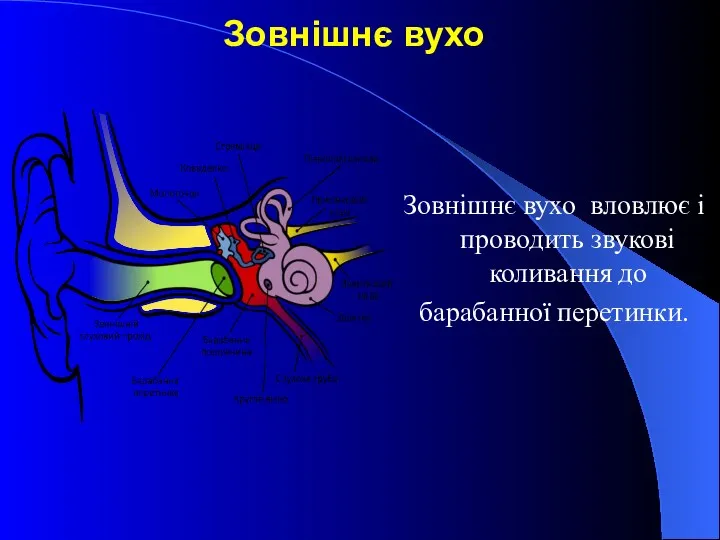
Зовнішнє вухо
Зовнішнє вухо вловлює і проводить звукові коливання до
барабанної перетинки.
Зовнішнє вухо
Зовнішнє вухо вловлює і проводить звукові коливання до
барабанної перетинки.
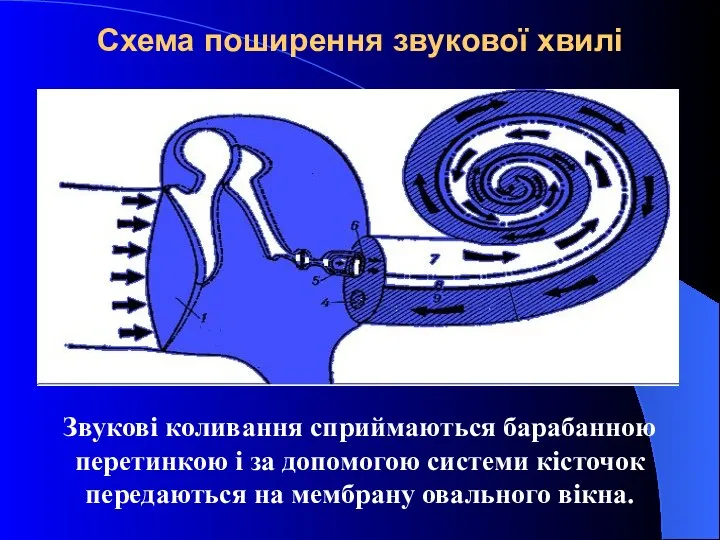
Схема поширення звукової хвилі
Звукові коливання сприймаються барабанною перетинкою і за допомогою
Схема поширення звукової хвилі
Звукові коливання сприймаються барабанною перетинкою і за допомогою

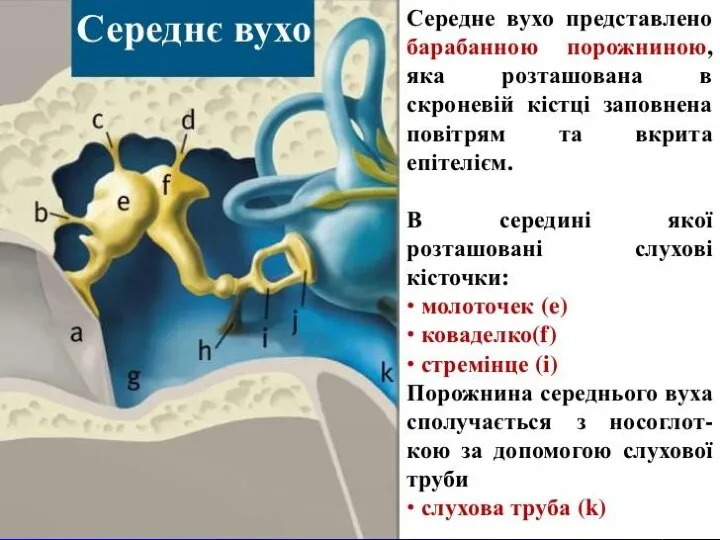
Среднє вухо
Слухові кісточки: молоточок, коваделко і стремінце передають і підсилюють коливання
Среднє вухо
Слухові кісточки: молоточок, коваделко і стремінце передають і підсилюють коливання

Внутрішнє вухо
Внутрішнє вухо включає присінок, півколові канали і завитку.
Завитка це кістковий
Внутрішнє вухо
Внутрішнє вухо включає присінок, півколові канали і завитку.
Завитка це кістковий
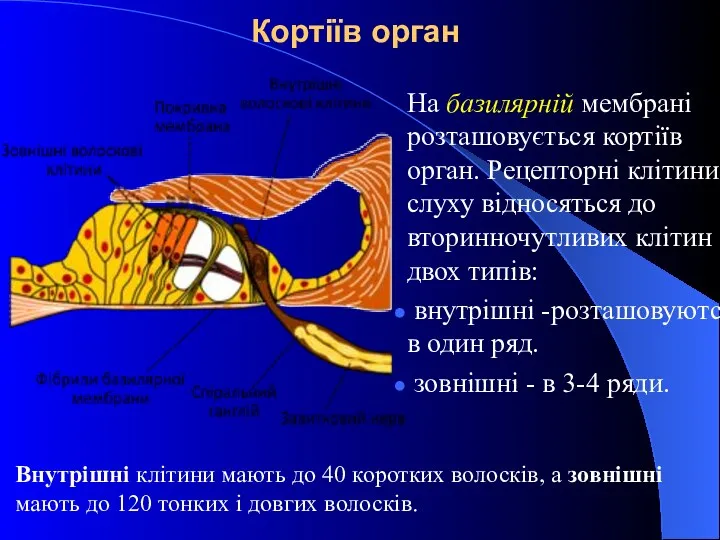
Кортіїв орган
На базилярній мембрані розташовується кортіїв орган. Рецепторні клітини слуху відносяться
Кортіїв орган
На базилярній мембрані розташовується кортіїв орган. Рецепторні клітини слуху відносяться

Волоскові клітини
Внутрішні волоскові клітини утворюють до 90% волокон слухового нерва і
Волоскові клітини
Внутрішні волоскові клітини утворюють до 90% волокон слухового нерва і

Вестибулярна і базилярна мембрани поділяють кістковий канал завитки на три канали:
Верхній
Вестибулярна і базилярна мембрани поділяють кістковий канал завитки на три канали:
Верхній

Ендо- и перилімфа внутрішнього вуха
Простір середніх сходів заповнений ендолімфою. Простір верхнього
Ендо- и перилімфа внутрішнього вуха
Простір середніх сходів заповнений ендолімфою. Простір верхнього

Механізм передачі коливань ендолімфи на рецепторні клітини кортіївого органу
Хвиля, яка виникла
Механізм передачі коливань ендолімфи на рецепторні клітини кортіївого органу
Хвиля, яка виникла

Амплітудний максимум
Основний механізм розрізнення висоти тонів обумовлений тим, що хвиля коливань
Амплітудний максимум
Основний механізм розрізнення висоти тонів обумовлений тим, що хвиля коливань
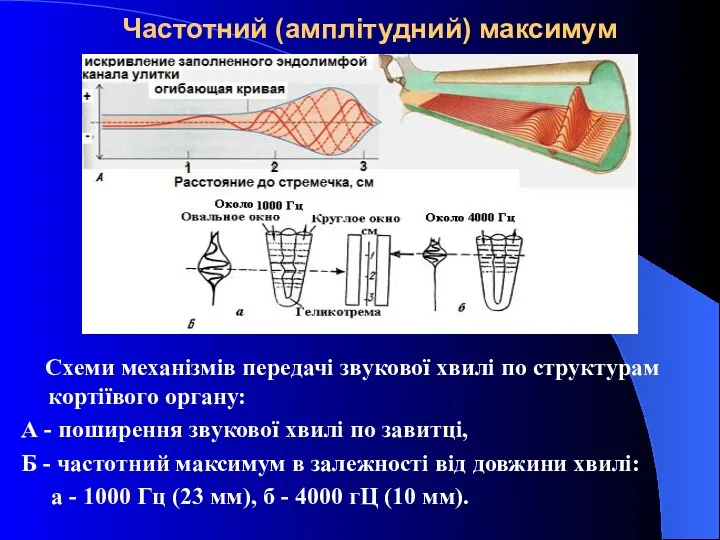
Частотний (амплітудний) максимум
Схеми механізмів передачі звукової хвилі по структурам кортіївого
Частотний (амплітудний) максимум
Схеми механізмів передачі звукової хвилі по структурам кортіївого

Визначення гучності
Діапазон амплітуди коливань ендолімфи пов'язаний з амплітудою коливання мембран. В
Визначення гучності
Діапазон амплітуди коливань ендолімфи пов'язаний з амплітудою коливання мембран. В

Сильний звук і реакція м'язів середнього вуха
Сильний звук може викликати пошкодження
Сильний звук і реакція м'язів середнього вуха
Сильний звук може викликати пошкодження

Предвокалізаційний рефлекс
Вберегти вухо людини від пошкоджуючої дії гучних звуків може допомогти
Предвокалізаційний рефлекс
Вберегти вухо людини від пошкоджуючої дії гучних звуків може допомогти

Аналіз звукових коливань в ЦНС
Закодована інформація міститься в звуковому стимулі у
Аналіз звукових коливань в ЦНС
Закодована інформація міститься в звуковому стимулі у

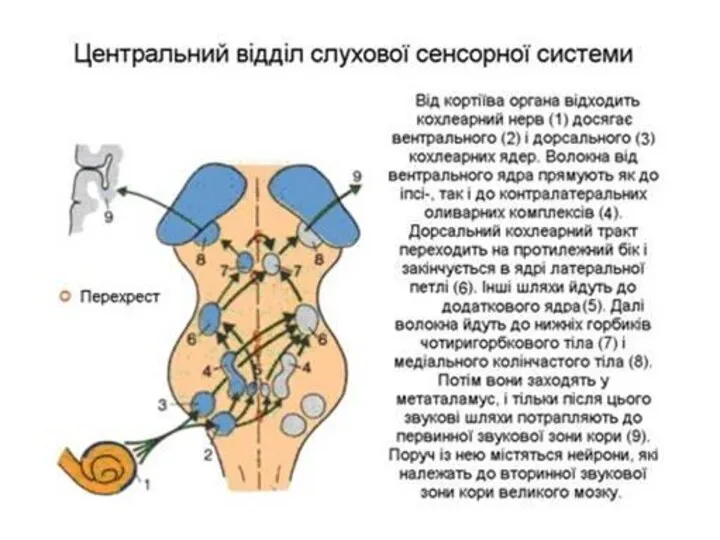
Обробка звукової інформації в ЦНС
Дорсальний кохлеарний тракт переходить на протилежну
Обробка звукової інформації в ЦНС
Дорсальний кохлеарний тракт переходить на протилежну

Центри кори
При одночасному збудженні нейронів підкіркових центрів і скроневих відділів кори
Центри кори
При одночасному збудженні нейронів підкіркових центрів і скроневих відділів кори

Центри кори та їх зв’язки
Від скроневої слухової ділянки кори відходять і
Центри кори та їх зв’язки
Від скроневої слухової ділянки кори відходять і
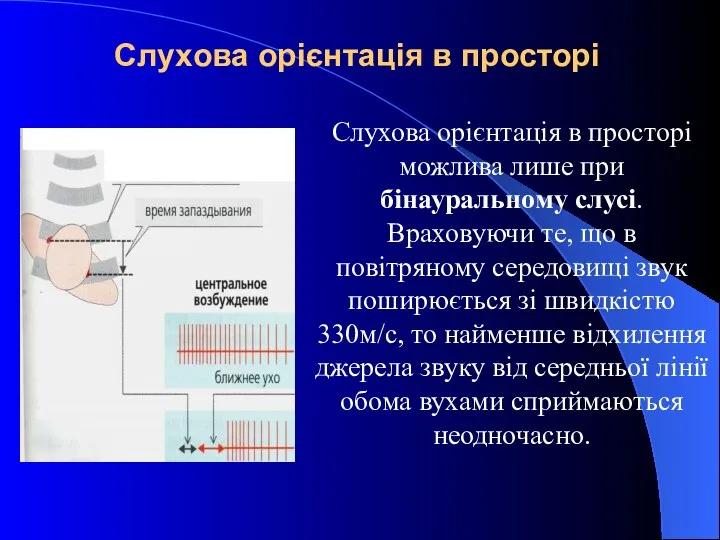
Слухова орієнтація в просторі
Слухова орієнтація в просторі можлива лише при бінауральному
Слухова орієнтація в просторі
Слухова орієнтація в просторі можлива лише при бінауральному

Людина і тварини мають просторовий слух, тобто здатні визначати положення джерела
Людина і тварини мають просторовий слух, тобто здатні визначати положення джерела

Вестибулярний аналізатор
Вестибулярний апарат - орган рівноваги, що дозволяє визначати положення і
Вестибулярний аналізатор
Вестибулярний апарат - орган рівноваги, що дозволяє визначати положення і

Вестибулярний орган
складається з:
Статолітового апарату
Півколових каналів
Маточки і мішечка
Завитки
Завиткового нерва
Вестибулярного нерва
Вестибулярний орган
складається з:
Статолітового апарату
Півколових каналів
Маточки і мішечка
Завитки
Завиткового нерва
Вестибулярного нерва
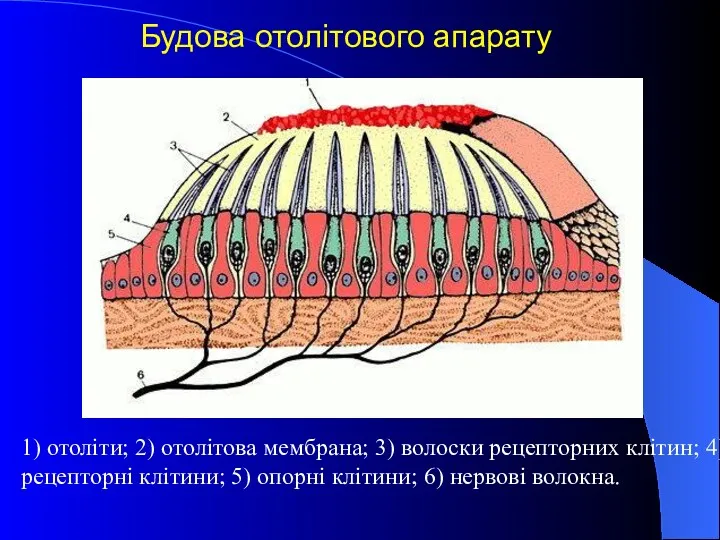
1) отоліти; 2) отолітова мембрана; 3) волоски рецепторних клітин; 4) рецепторні
1) отоліти; 2) отолітова мембрана; 3) волоски рецепторних клітин; 4) рецепторні
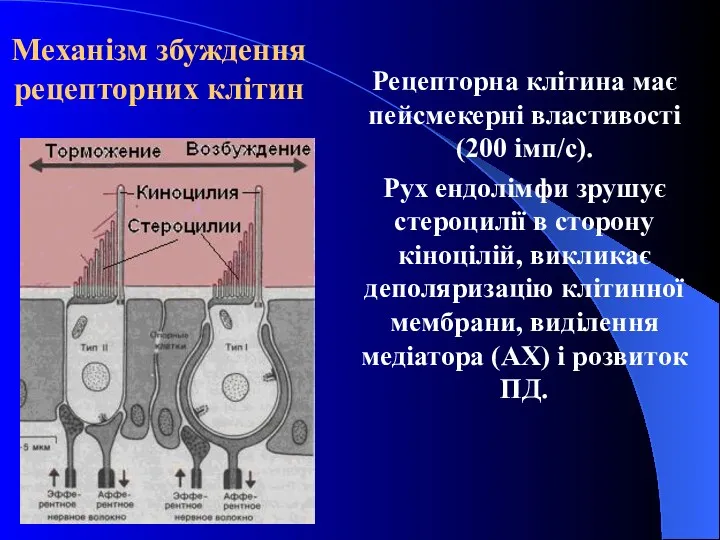
Механізм збуждення рецепторних клітин
Рецепторна клітина має пейсмекерні властивості (200 імп/с).
Рух ендолімфи
Механізм збуждення рецепторних клітин
Рецепторна клітина має пейсмекерні властивості (200 імп/с).
Рух ендолімфи

Маточка і мішечок
Рецептори маточки і мішечка контролюють прямолінійні рухи людини. Подразнюються
Маточка і мішечок
Рецептори маточки і мішечка контролюють прямолінійні рухи людини. Подразнюються

Півколові канали
Природним стимулом для рецепторів півколових каналів є кутові прискорення. Війки
Півколові канали
Природним стимулом для рецепторів півколових каналів є кутові прискорення. Війки

Півколові канали розташовані у трьох взаємно перпендикулярних площинах. На одному зі
Півколові канали розташовані у трьох взаємно перпендикулярних площинах. На одному зі
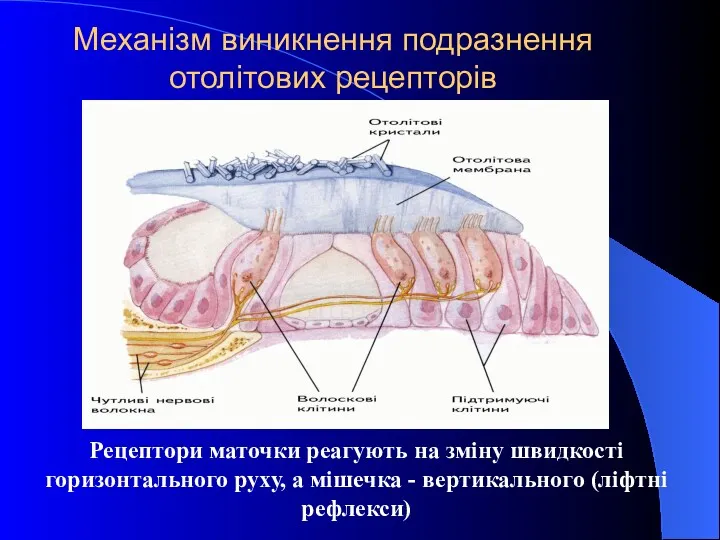
Рецептори маточки реагують на зміну швидкості горизонтального руху, а мішечка -
Рецептори маточки реагують на зміну швидкості горизонтального руху, а мішечка -
 Популяционно-видовой уровень организации жизни
Популяционно-видовой уровень организации жизни зрительный анализатор2
зрительный анализатор2 Класс Земноводные, или Амфибии (Amphibia)
Класс Земноводные, или Амфибии (Amphibia) Доказательства происхождения животных
Доказательства происхождения животных Епітеліальні тканини
Епітеліальні тканини Мастер-класс. Структурно-логические технологиии(применение на уроках биологии схем,таблиц, разноуровневых заданий).
Мастер-класс. Структурно-логические технологиии(применение на уроках биологии схем,таблиц, разноуровневых заданий). Мир микробов
Мир микробов 27 февраля – Международный день белого медведя
27 февраля – Международный день белого медведя Паразитизм. Хищничество
Паразитизм. Хищничество Кости черепа
Кости черепа Формовое садоводство
Формовое садоводство Токсикология биологических вредных веществ
Токсикология биологических вредных веществ Раздел вторичнополостные. Надтип полимерные. Тип кольчатые черви
Раздел вторичнополостные. Надтип полимерные. Тип кольчатые черви Сумчатые млекопитающие
Сумчатые млекопитающие Плоды и ягоды. Семечковые плоды
Плоды и ягоды. Семечковые плоды Трансгенные организмы
Трансгенные организмы Обмін БЖВ
Обмін БЖВ Скелет конечностей
Скелет конечностей Кислотно-щелочное состояние организма (КЩС)
Кислотно-щелочное состояние организма (КЩС) Презентация к КВН Зеленые друзья 6 класс
Презентация к КВН Зеленые друзья 6 класс Vitamins. Classes of Vitamins
Vitamins. Classes of Vitamins Хвойные деревья. Кустарники. Лиственные деревья
Хвойные деревья. Кустарники. Лиственные деревья Журавлеподібні
Журавлеподібні Morphometrical and histological comparative analyses of omnivorous birds gastrointestinal tract
Morphometrical and histological comparative analyses of omnivorous birds gastrointestinal tract Обрезка плодовых деревьев и кустарников
Обрезка плодовых деревьев и кустарников Ткани организмов. Культивирование тканей
Ткани организмов. Культивирование тканей Methods in behavioral genetics
Methods in behavioral genetics Царства живой природы
Царства живой природы