- Соотношение микроэволюционных и макроэволюционных процессов

Содержание
- 2. 1. Материалом для эволюции служат, как правило, очень мелкие, но дискретные изменения наследственности - мутации. Мутационная
- 3. 4. Эволюция носит дивергентный характер, т.е. один таксон может стать предком нескольких дочерних таксонов, но каждый
- 4. 7. Обмен аллелями, "поток генов" возможен лишь внутри вида. Если мутация имеет положительную селективную ценность на
- 5. 10. Любой реальный, а не сборный таксон имеет однокорневое, монофилетическое происхождение (следствие из п. 4). Монофилетическое
- 6. Главная черта СТЭ, иногда называемой неодарвинизмом, в том, что она представляет собой вероятностную концепцию, ибо и
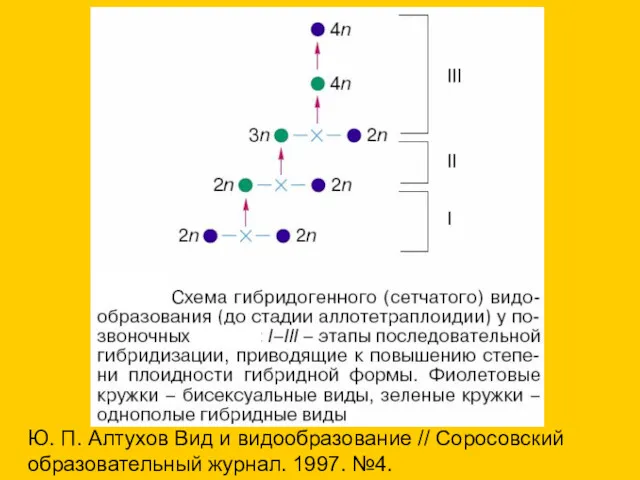
- 7. «В эволюции ряда групп тесно связаны межвидовая гибридизация, стадия бесполого размножения (партеногенез или гиногенез) и полиплоидия»...
- 8. 1. При межвидовой гибридизации образуется диплоидная форма, переходящая к однополому размножению. 2. При скрещивании этих гибридных
- 9. Появление тетраплоидов благодаря формированию нередуцированных гамет
- 10. Ю. П. Алтухов Вид и видообразование // Соросовский образовательный журнал. 1997. №4.
- 11. Тибетская жаба (Индия) Bufo zamdensis с триплоидным кариотипом (n=33) самка самец
- 12. Предварительно выровненный гибридный мейоз Stock et. al., 2006
- 13. Видообразование через реорганизацию закрытой системы генома Carson H. The Genetics of Speciation at the Diploid Level
- 14. Britten R., Davidson E. Repetitive and Non-repetitive DNA Sequences and a Speculation on the Origin of
- 15. Оказалось, что элементарные эволюционные факторы (волны жизни, миграция, изоляция, эффект основателя, дрейф генов, эффект бутылочного горла
- 16. Микроэволюция – совокупность эволюционных процессов, протекающих в популяциях вида и приводящих к изменению генофонда этих популяций
- 17. Микроэволюция – совокупность эволюционных процессов, протекающих в популяциях вида и приводящих к изменению генофонда этих популяций
- 18. Традиционно считается, что микроэволюция – процесс видообразования, макроэволюция – процесс образования более крупных таксонов. Микро- и
- 19. Соответствующие термины — микро- и макроэволюция — были предложены в 1927 Ю.А.Филипченко Б. Ренш (1959) -
- 20. Р.Гольдшмидт «Эволюция», 1940 (R.Goldschmidt Evolution. New Haven, 1940. 436 p.) Микроэволюционные процессы обусловлены микромутациями. Макроэволюционные изменения
- 21. Дж. Симпсон «Темпы и формы эволюции» Единство факторов и механизмов микро- и макроэволюции. Гипотеза «квантовой эволюции».
- 22. Формы эволюции (по Дж. Симпсону) Simpson, 1944, 1953 Видообразование (адаптивная радиация таксона на несколько внутри одной
- 23. Направление видообразования. Направление (линия Р→Q) представляет собой результирующий вектор последовательных видообразовательных изменений в данном направлении (из
- 24. Различные формы видообразования
- 25. Филетическая эволюция а - направленное движение популяции, хорошо приспособленной к устойчивой зоне; б - направленная специализация
- 26. Филетическое направление в линии плейстоценовых европейских слонов Elephas planifrons — Е. meridionalis. Ламеллярный индекс служит мерой
- 27. Сравнение скорости эволюционных изменений и величины популяции при квантовой и обычной филетической эволюции (Simpson, 1953) появление
- 28. Квантовая эволюция на примере семейства Equidae (переход от питания листьями деревьев к питанию травой)
- 29. Возникновение Stylinodontinae от Conoryctinae путем квантовой эволюции (Patterson, 1949)
- 30. Теория главных направлений эволюции (А.Н. Северцов, 1939) Ароморфоз - повышение уровня организации (арогенез). Идиоадаптация - выработка
- 31. снижение экологической валентности таксона повышение экологической валентности таксона
- 32. Теория главных направлений эволюции (И.И. Шмальгаузен , 1939) Ароморфоз - повышение уровня организации (арогенез, эпиморфоз). Идиоадаптация
- 33. 1) алломорфоз, или смена среды, при котором одни связи организма со средой заменяются другими, более или
- 34. Теломорфоз назикобатрахус крот айолот кольчатая червяга
- 35. Гиперморфоз
- 36. Катаморфоз
- 37. Гипоморфоз аксолотль протей амфиума сирен
- 38. Гипотеза прерывистого равновесия (Eldredge, Gould, 1972, 1977) 1) традиционно эволюцию рассматривают как состояние медленного и равномерного
- 39. В. Эволюция путем обычного анагенеза, направленность задается благодаря внутривидовым изменениям и видообразование просто увеличивает число линий,
- 40. (Eldredge and Gould, 1972) Эволюционная тенденция в кладе В - нет предпочтительного направления видообразования, тенденция определяется
- 41. Стив Гулд (Steve Gould, 1980, 1982) Существует коренное различие между микро- и макроэволюцией. Попытка объяснить макроэволюцию
- 42. Проблема соотношений микро- и макроэволюции остается остро дискуссионной... сальтационная концепция рассматривает микро- и макроэволюцию как качественно
- 43. В. Грант «Эволюционный процесс» Макроэволюция подразумевает изменения гораздо большего масштаба, чем те, которые происходят при микроэволюции
- 44. Системная концепция макроэволюции - интегратизм (В.А.Энгельгардт ) Макроэволюция - системно организованный процесс, организация которого выражается в
- 45. «Напомним возражение — весьма обоснованное,— которое выдвигают телеологи против механистической теории эволюции: если эволюция направляется случайными
- 46. Доклад С.Г.Инге-Вечтомова на рабочем совещании "Происхождение и эволюция живых систем"
- 47. Простые мутации, замены пар нуклеотидов не являются движущей силой эволюции (нестыковка по времени). «...трудно исправить часы,
- 48. «...все ныне сущее возникло за счет комбинаторики исходно возникшего небольшого числа неких инвариантных доменов, которые далее
- 49. Единство механизмов микро- и макроэволюции (экологические механизмы) Эволюционная судьба каждого вида и их взаимодействие подчинены общей
- 51. Скачать презентацию

1. Материалом для эволюции служат, как правило, очень мелкие, но дискретные
1. Материалом для эволюции служат, как правило, очень мелкие, но дискретные

4. Эволюция носит дивергентный характер, т.е. один таксон может стать предком
4. Эволюция носит дивергентный характер, т.е. один таксон может стать предком

7. Обмен аллелями, "поток генов" возможен лишь внутри вида. Если мутация
7. Обмен аллелями, "поток генов" возможен лишь внутри вида. Если мутация

10. Любой реальный, а не сборный таксон имеет однокорневое, монофилетическое происхождение
10. Любой реальный, а не сборный таксон имеет однокорневое, монофилетическое происхождение

Главная черта СТЭ, иногда называемой неодарвинизмом, в том, что она представляет
Главная черта СТЭ, иногда называемой неодарвинизмом, в том, что она представляет

«В эволюции ряда групп тесно связаны межвидовая гибридизация, стадия бесполого размножения
«В эволюции ряда групп тесно связаны межвидовая гибридизация, стадия бесполого размножения

1. При межвидовой гибридизации образуется диплоидная форма, переходящая к однополому размножению.
2.
1. При межвидовой гибридизации образуется диплоидная форма, переходящая к однополому размножению.
2.
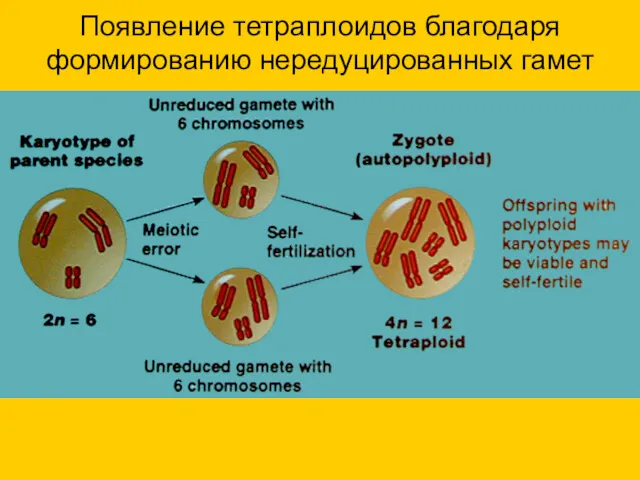
Появление тетраплоидов благодаря формированию нередуцированных гамет
Появление тетраплоидов благодаря формированию нередуцированных гамет
Ю. П. Алтухов Вид и видообразование // Соросовский образовательный журнал. 1997.
Ю. П. Алтухов Вид и видообразование // Соросовский образовательный журнал. 1997.

Тибетская жаба (Индия) Bufo zamdensis с триплоидным кариотипом (n=33)
самка
самец
Тибетская жаба (Индия) Bufo zamdensis с триплоидным кариотипом (n=33)
самка
самец
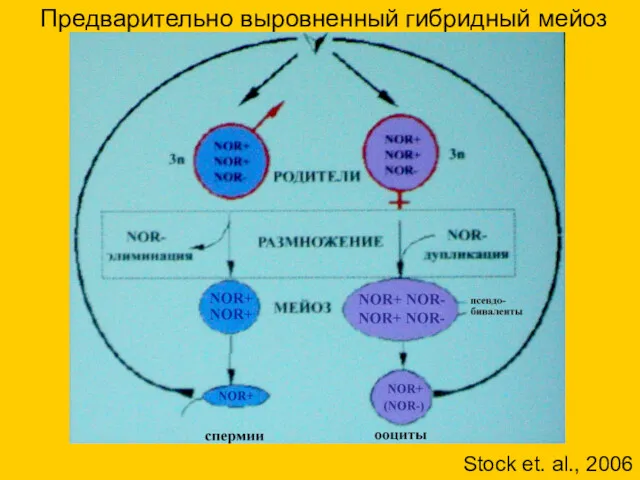
Предварительно выровненный гибридный мейоз
Stock et. al., 2006
Предварительно выровненный гибридный мейоз
Stock et. al., 2006
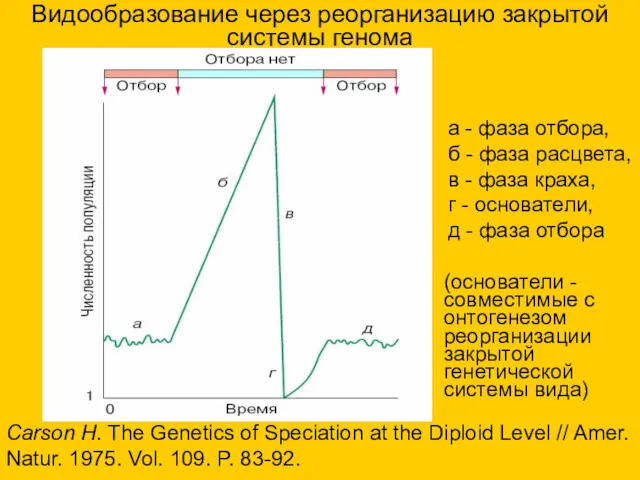
Видообразование через реорганизацию закрытой системы генома
Carson H. The Genetics of Speciation
Видообразование через реорганизацию закрытой системы генома
Carson H. The Genetics of Speciation
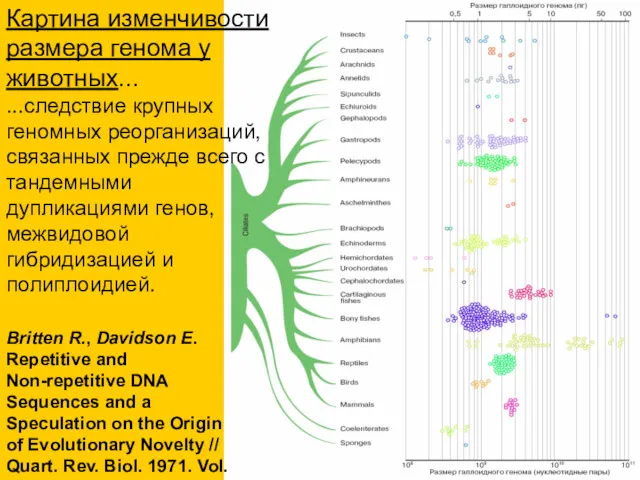
Britten R., Davidson E. Repetitive and Non-repetitive DNA Sequences and a
Britten R., Davidson E. Repetitive and Non-repetitive DNA Sequences and a

Оказалось, что элементарные эволюционные факторы (волны жизни, миграция, изоляция, эффект основателя,
Оказалось, что элементарные эволюционные факторы (волны жизни, миграция, изоляция, эффект основателя,

Микроэволюция – совокупность эволюционных процессов, протекающих в популяциях вида и приводящих
Микроэволюция – совокупность эволюционных процессов, протекающих в популяциях вида и приводящих

Микроэволюция – совокупность эволюционных процессов, протекающих в популяциях вида и приводящих
Микроэволюция – совокупность эволюционных процессов, протекающих в популяциях вида и приводящих

Традиционно считается, что микроэволюция – процесс видообразования, макроэволюция – процесс образования
Традиционно считается, что микроэволюция – процесс видообразования, макроэволюция – процесс образования

Соответствующие термины — микро- и макроэволюция — были предложены в 1927
Соответствующие термины — микро- и макроэволюция — были предложены в 1927

Р.Гольдшмидт «Эволюция», 1940
(R.Goldschmidt Evolution. New Haven, 1940. 436 p.)
Микроэволюционные процессы
Р.Гольдшмидт «Эволюция», 1940
(R.Goldschmidt Evolution. New Haven, 1940. 436 p.)
Микроэволюционные процессы

Дж. Симпсон
«Темпы и формы эволюции»
Единство факторов и механизмов микро- и
Дж. Симпсон
«Темпы и формы эволюции»
Единство факторов и механизмов микро- и

Формы эволюции (по Дж. Симпсону) Simpson, 1944, 1953
Видообразование (адаптивная радиация таксона
Формы эволюции (по Дж. Симпсону) Simpson, 1944, 1953
Видообразование (адаптивная радиация таксона
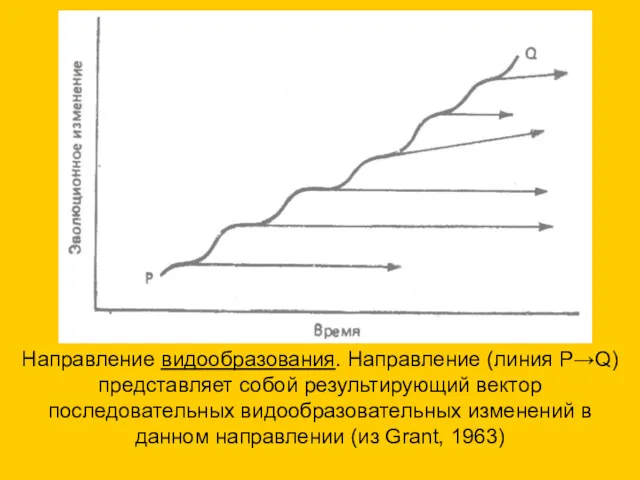
Направление видообразования. Направление (линия Р→Q) представляет собой результирующий вектор последовательных видообразовательных
Направление видообразования. Направление (линия Р→Q) представляет собой результирующий вектор последовательных видообразовательных
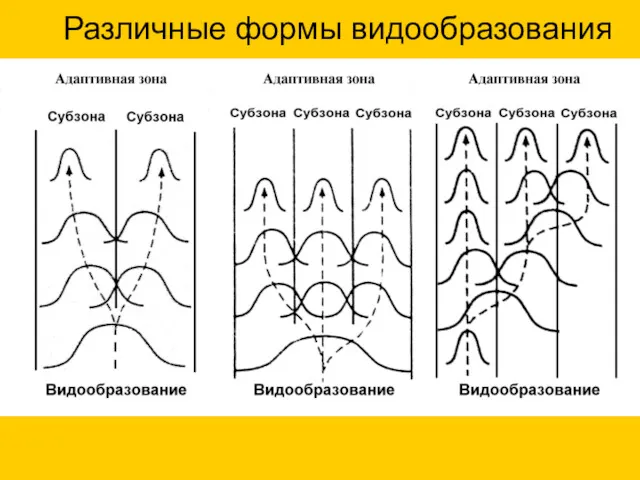
Различные формы видообразования
Различные формы видообразования
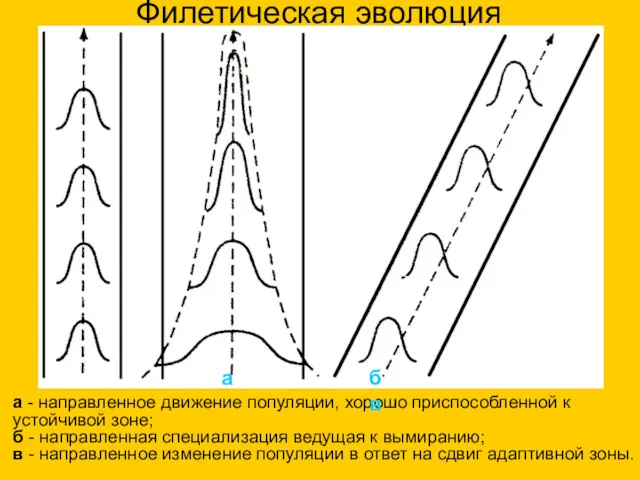
Филетическая эволюция
а - направленное движение популяции, хорошо приспособленной к устойчивой зоне;
Филетическая эволюция
а - направленное движение популяции, хорошо приспособленной к устойчивой зоне;
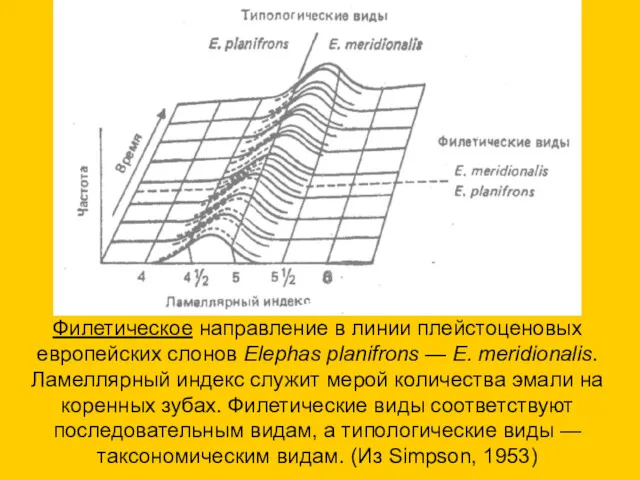
Филетическое направление в линии плейстоценовых европейских слонов Elephas planifrons — Е.
Филетическое направление в линии плейстоценовых европейских слонов Elephas planifrons — Е.
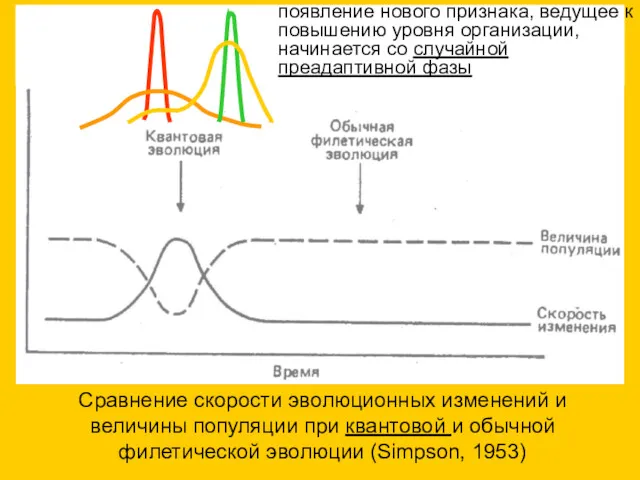
Сравнение скорости эволюционных изменений и величины популяции при квантовой и обычной
Сравнение скорости эволюционных изменений и величины популяции при квантовой и обычной
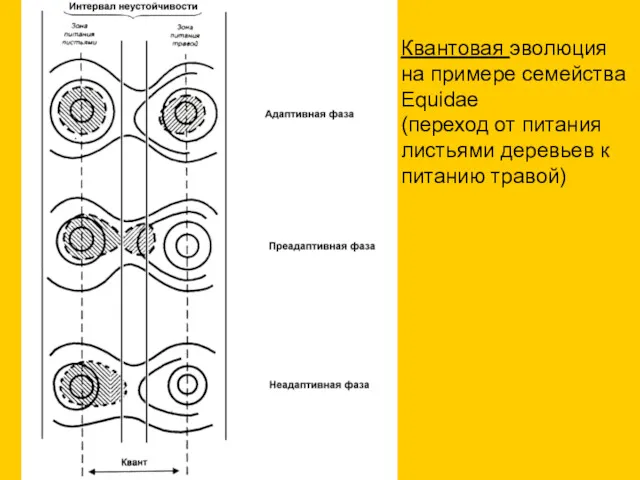
Квантовая эволюция на примере семейства Equidae
(переход от питания листьями деревьев к
Квантовая эволюция на примере семейства Equidae
(переход от питания листьями деревьев к
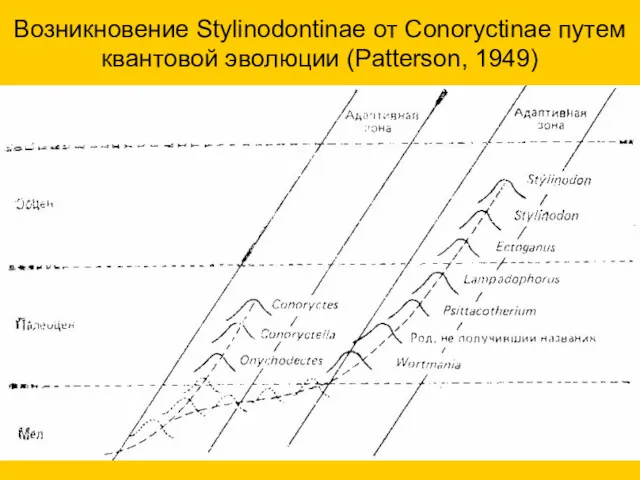
Возникновение Stylinodontinae от Conoryctinae путем квантовой эволюции (Patterson, 1949)
Возникновение Stylinodontinae от Conoryctinae путем квантовой эволюции (Patterson, 1949)

Теория главных направлений эволюции (А.Н. Северцов, 1939)
Ароморфоз - повышение уровня организации
Теория главных направлений эволюции (А.Н. Северцов, 1939)
Ароморфоз - повышение уровня организации
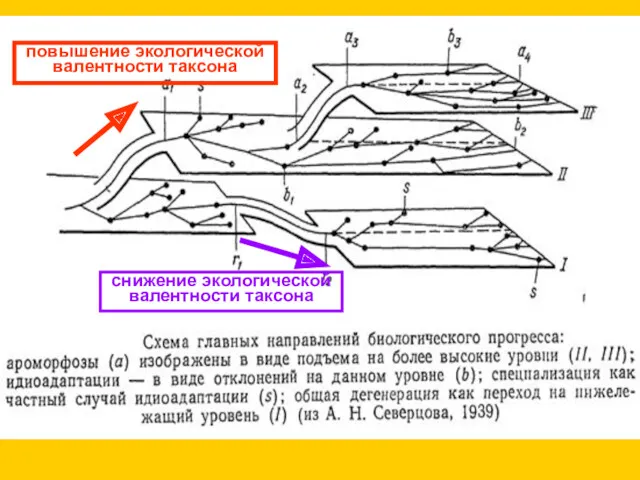
снижение экологической
валентности таксона
повышение экологической
валентности таксона
снижение экологической
валентности таксона
повышение экологической
валентности таксона

Теория главных направлений эволюции (И.И. Шмальгаузен , 1939)
Ароморфоз - повышение уровня
Теория главных направлений эволюции (И.И. Шмальгаузен , 1939)
Ароморфоз - повышение уровня

1) алломорфоз, или смена среды, при котором одни связи организма со
1) алломорфоз, или смена среды, при котором одни связи организма со

Теломорфоз
назикобатрахус
крот
айолот
кольчатая червяга
Теломорфоз
назикобатрахус
крот
айолот
кольчатая червяга

Гиперморфоз
Гиперморфоз
Катаморфоз
Катаморфоз

Гипоморфоз
аксолотль
протей
амфиума
сирен
Гипоморфоз
аксолотль
протей
амфиума
сирен

Гипотеза прерывистого равновесия (Eldredge, Gould, 1972, 1977)
1) традиционно эволюцию рассматривают как
Гипотеза прерывистого равновесия (Eldredge, Gould, 1972, 1977)
1) традиционно эволюцию рассматривают как
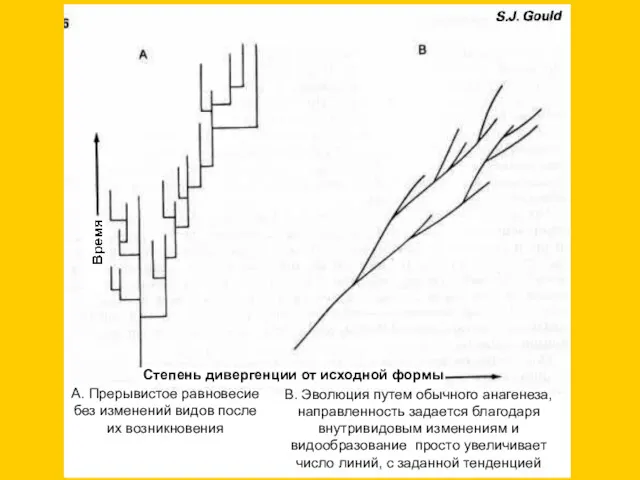
В. Эволюция путем обычного анагенеза, направленность задается благодаря внутривидовым изменениям и
В. Эволюция путем обычного анагенеза, направленность задается благодаря внутривидовым изменениям и
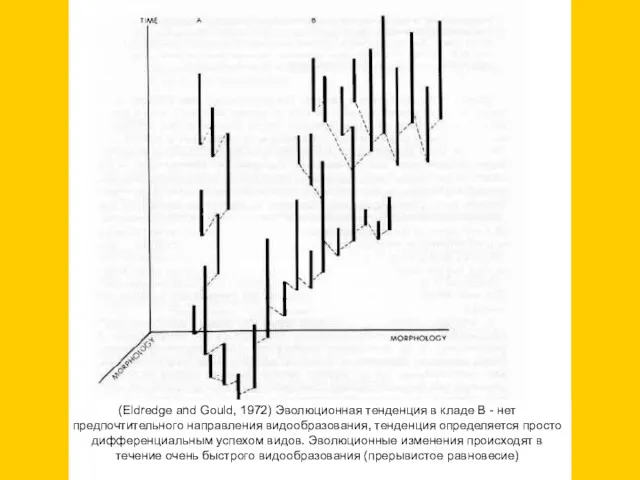
(Eldredge and Gould, 1972) Эволюционная тенденция в кладе В - нет
(Eldredge and Gould, 1972) Эволюционная тенденция в кладе В - нет

Стив Гулд
(Steve Gould, 1980, 1982)
Существует коренное различие между микро- и
Стив Гулд
(Steve Gould, 1980, 1982)
Существует коренное различие между микро- и

Проблема соотношений микро- и макроэволюции остается остро дискуссионной...
сальтационная концепция рассматривает
Проблема соотношений микро- и макроэволюции остается остро дискуссионной... сальтационная концепция рассматривает

В. Грант
«Эволюционный процесс»
Макроэволюция подразумевает изменения гораздо большего масштаба, чем
В. Грант
«Эволюционный процесс»
Макроэволюция подразумевает изменения гораздо большего масштаба, чем

Системная концепция макроэволюции
- интегратизм (В.А.Энгельгардт )
Макроэволюция - системно организованный процесс, организация которого
Системная концепция макроэволюции - интегратизм (В.А.Энгельгардт ) Макроэволюция - системно организованный процесс, организация которого

«Напомним возражение — весьма обоснованное,— которое выдвигают телеологи против механистической теории
«Напомним возражение — весьма обоснованное,— которое выдвигают телеологи против механистической теории
Доклад С.Г.Инге-Вечтомова на рабочем совещании "Происхождение и эволюция живых систем"
Доклад С.Г.Инге-Вечтомова на рабочем совещании "Происхождение и эволюция живых систем"

Простые мутации, замены пар нуклеотидов не являются движущей силой эволюции (нестыковка
Простые мутации, замены пар нуклеотидов не являются движущей силой эволюции (нестыковка

«...все ныне сущее возникло за счет комбинаторики исходно возникшего небольшого числа
«...все ныне сущее возникло за счет комбинаторики исходно возникшего небольшого числа

Единство механизмов микро- и макроэволюции (экологические механизмы)
Эволюционная судьба каждого вида
Единство механизмов микро- и макроэволюции (экологические механизмы)
Эволюционная судьба каждого вида
 Ещё немного о белках
Ещё немного о белках Загадка человека. Наследственность. Урок 3
Загадка человека. Наследственность. Урок 3 Тип Членистоногие. Класс Паукообразные
Тип Членистоногие. Класс Паукообразные Кольчатые черви
Кольчатые черви Урок по теме Семейства класса двудольных растений
Урок по теме Семейства класса двудольных растений Гуморальная регуляция. Эндокринная система
Гуморальная регуляция. Эндокринная система Строение клеток живых организмов. Лабораторная работа
Строение клеток живых организмов. Лабораторная работа Мінез-құлық қалыптасуының жалпы принцептері. ЖЖІӘ туралы түсінік. Туа біткен және жүре пайда болған іс әрекеттерінің түрлері
Мінез-құлық қалыптасуының жалпы принцептері. ЖЖІӘ туралы түсінік. Туа біткен және жүре пайда болған іс әрекеттерінің түрлері Биоактивные пептиды
Биоактивные пептиды Нейромедиаторы. Критерии нейромедиатора
Нейромедиаторы. Критерии нейромедиатора Структурно-функциональные особенности биокатализа
Структурно-функциональные особенности биокатализа песчаная эфа
песчаная эфа Минеральное питание растений. Ассимиляция элементов
Минеральное питание растений. Ассимиляция элементов Цикличность поведения женских особей
Цикличность поведения женских особей Анатомия домашних животных
Анатомия домашних животных Понятие об организме. Основные части организма
Понятие об организме. Основные части организма Biological effects of nitric oxide and its role in cell signaling
Biological effects of nitric oxide and its role in cell signaling Формы размножения организмов. Бесполое размножение
Формы размножения организмов. Бесполое размножение Класс Пресмыкающиеся
Класс Пресмыкающиеся Многообразие насекомых
Многообразие насекомых Барыня тыква
Барыня тыква Филогенетический анализ и молекулярная эволюция. Лекция 5
Филогенетический анализ и молекулярная эволюция. Лекция 5 Черепно-мозговые нервы
Черепно-мозговые нервы Биофизика гормональных рецепторов
Биофизика гормональных рецепторов Подготовка к Всероссийской проверочной работе по биологии. 5 класс
Подготовка к Всероссийской проверочной работе по биологии. 5 класс Наука биология и условия жизни организмов
Наука биология и условия жизни организмов Нуклеїнові кислоти (ДНК та РНК)
Нуклеїнові кислоти (ДНК та РНК) Биомембраны. Пассивный и активный транспорт веществ
Биомембраны. Пассивный и активный транспорт веществ