- Строение и функции белков. Ферменты

Содержание
- 2. Первичная структура белка Первичная структура белка – это последовательность расположения аминокислот в полипептидной цепи. Её определяют,
- 3. Вторичная структура белка Вторичная структура белка – способ упаковки очень длинной полипептидной цепи в α –
- 4. Третичная структура белка Третичная структура белка – трёхмерная пространственная ориентация полипептидной спирали или складчатой структуры в
- 5. Схема образования связей между радикалами аминокислот 1 – ионные связи, 2 – водородные связи, 3 –
- 6. Четвертичная структура белка Четвертичная структура белка – способ укладки в пространстве отдельных полипептидных цепей и формирование
- 7. Цветные реакции для обнаружения белков и аминокислот Для идентификации пептидов, белков и отдельных аминокислот используют так
- 8. Классификация белков Белки можно классифицировать: – по форме молекул (глобулярные и фибриллярные); – по молекулярной массе
- 9. Классификация простых белков Альбумины. Примерно 75-80% осмотического давления белков сыворотки крови приходится на альбумины; еще одна
- 10. Классификация сложных белков
- 11. Функции белков
- 12. Скорость биологически важных реакций органических соединений Общая скорость химической реакции определяется (лимитируется) скоростью ее наиболее медленной
- 13. Влияние катализатора на скорость реакции
- 14. Различают субстрат и действующее на него соединение – реагент (реакционная частица) . Субстрат - вещество, в
- 15. Строение фермента
- 16. Строение фермента
- 17. Абсолютная специфичность – фермент катализирует превращение только одного вещества. Например, расщепление мочевины уреазой. Групповая (относительная) специфичность
- 18. Механизм ферментативного катализа 1. Теория Фишера: «ключ-замок». 2. Теория Кошланда: «рука и перчатка». 3. Теория вынужденного
- 19. Механизм ферментативного катализа Теория «ключ-замок» Теория индуцированного взаимодействия активный центр субстрат фермент фермент-субстратный фермент продукт комплекс
- 20. Аллостерическая регуляция активности
- 21. Зависимость активности фермента от рН среды
- 22. Зависимость активности фермента от температуры фермент фермент после тепловой денатурации
- 23. Классификация ферментов Согласно современной классификации, выделяют шесть классов ферментов: оксидоредуктазы; трансферазы; гидролазы; лиазы; изомеразы; лигазы.
- 24. Оксидоредуктазы – катализируют окислительно-восстановительные реакции:
- 25. Коферменты дегидрогеназ НАД+ (НикотинамидАденинДинуклеотид), НАДФ содержат витамин РР ФАД (ФлавинАденинДинуклеотид), ФМН (ФлавинаденинМоноНуклеотид) содержат витамин В2 Коферменты
- 26. Трансферазы – катализируют реакции межмолекулярного переноса различных атомов, групп атомов и радикалов. Кофермент ацилтрансфераз HSКоА содержит
- 27. Гидролазы – ускоряют реакции гидролиза (при участии воды). Продукты реакции имеют более простое строение, чем субстрат.
- 28. Гликозидазы: амилаза, мальтаза, лактаза, сахараза Пептидазы: пепсин, трипсин, химотрипсин, эластаза
- 29. Лиазы – ферменты, катализирующие разрыв связей, а также обратимые реакции отщепления - присоединения различных групп от
- 30. Изомеразы – катализируют взаимопревращения структурных и простарнственных изомеров.
- 31. Лигазы (синтетазы) – катализируют синтез органических веществ из двух исходных молекул с использованием энергии распада АТФ
- 33. Скачать презентацию
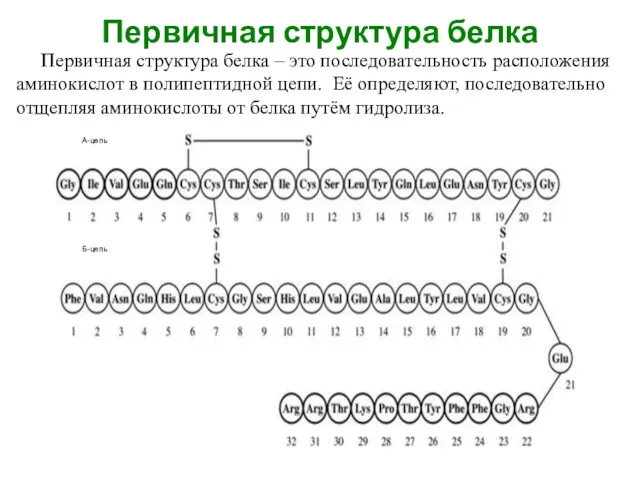
Первичная структура белка
Первичная структура белка – это последовательность расположения аминокислот в
Первичная структура белка
Первичная структура белка – это последовательность расположения аминокислот в
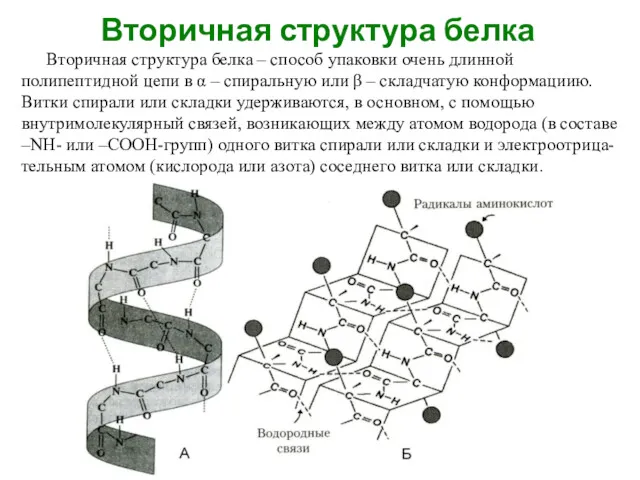
Вторичная структура белка
Вторичная структура белка – способ упаковки очень длинной полипептидной
Вторичная структура белка
Вторичная структура белка – способ упаковки очень длинной полипептидной

Третичная структура белка
Третичная структура белка – трёхмерная пространственная ориентация полипептидной спирали
Третичная структура белка
Третичная структура белка – трёхмерная пространственная ориентация полипептидной спирали
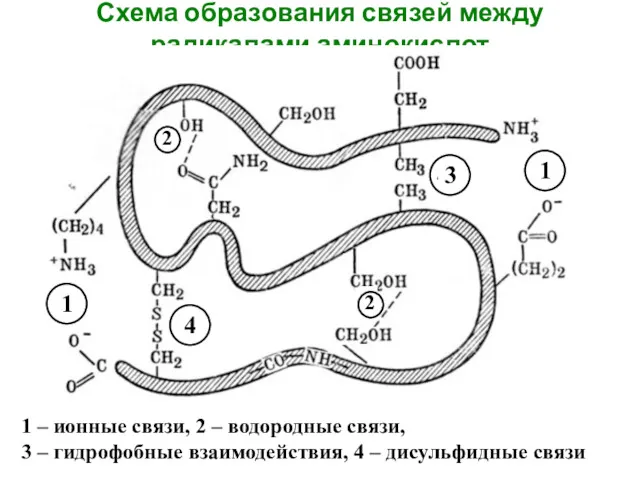
Схема образования связей между радикалами аминокислот
1 – ионные связи, 2 –
Схема образования связей между радикалами аминокислот
1 – ионные связи, 2 –
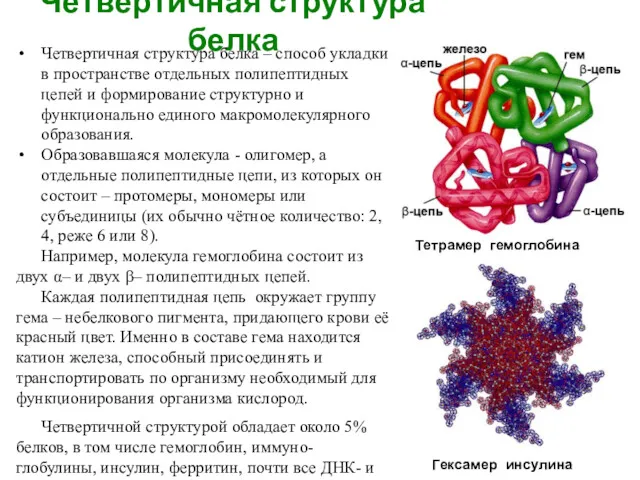
Четвертичная структура белка
Четвертичная структура белка – способ укладки в пространстве отдельных
Четвертичная структура белка
Четвертичная структура белка – способ укладки в пространстве отдельных

Цветные реакции для обнаружения белков и аминокислот
Для идентификации пептидов, белков и
Цветные реакции для обнаружения белков и аминокислот
Для идентификации пептидов, белков и

Классификация белков
Белки можно классифицировать:
– по форме молекул (глобулярные и фибриллярные);
–
Классификация белков
Белки можно классифицировать:
– по форме молекул (глобулярные и фибриллярные);
–

Классификация простых белков
Альбумины. Примерно 75-80% осмотического давления белков сыворотки крови приходится
Классификация простых белков
Альбумины. Примерно 75-80% осмотического давления белков сыворотки крови приходится
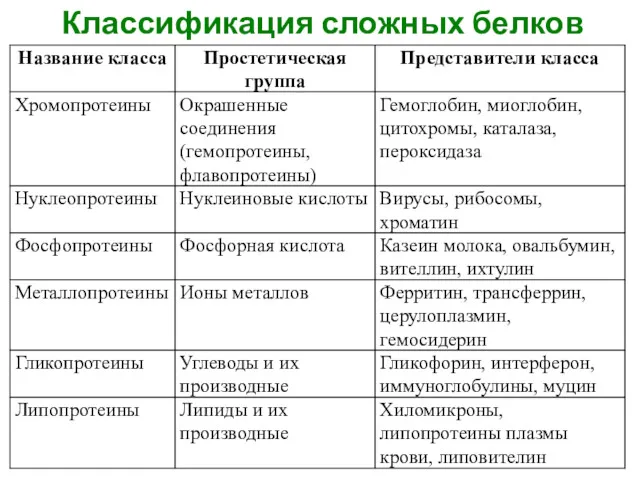
Классификация сложных белков
Классификация сложных белков

Функции белков
Функции белков

Скорость биологически важных реакций органических соединений
Общая скорость химической реакции определяется (лимитируется)
Скорость биологически важных реакций органических соединений
Общая скорость химической реакции определяется (лимитируется)
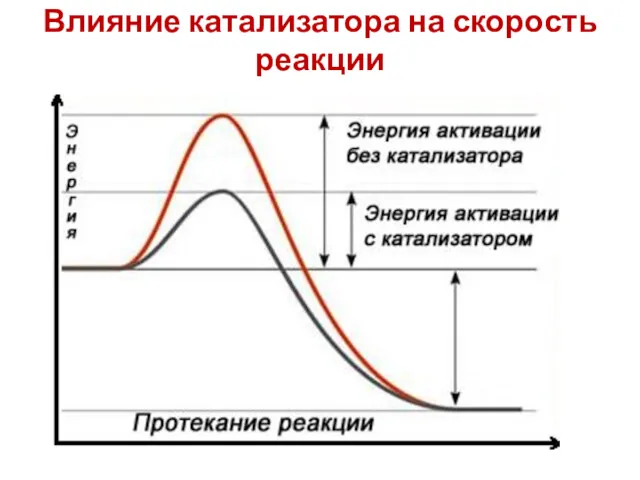
Влияние катализатора на скорость реакции
Влияние катализатора на скорость реакции

Различают субстрат и действующее на него соединение – реагент (реакционная частица)
Различают субстрат и действующее на него соединение – реагент (реакционная частица)
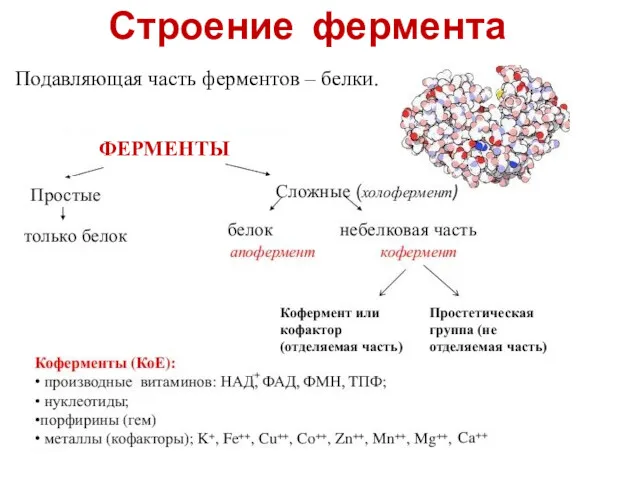
Строение фермента
Строение фермента
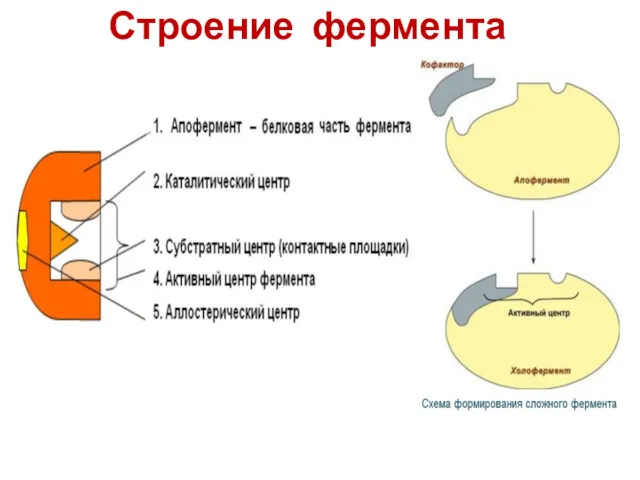
Строение фермента
Строение фермента

Абсолютная специфичность – фермент катализирует превращение только одного вещества. Например, расщепление мочевины уреазой.
Групповая
Абсолютная специфичность – фермент катализирует превращение только одного вещества. Например, расщепление мочевины уреазой.
Групповая
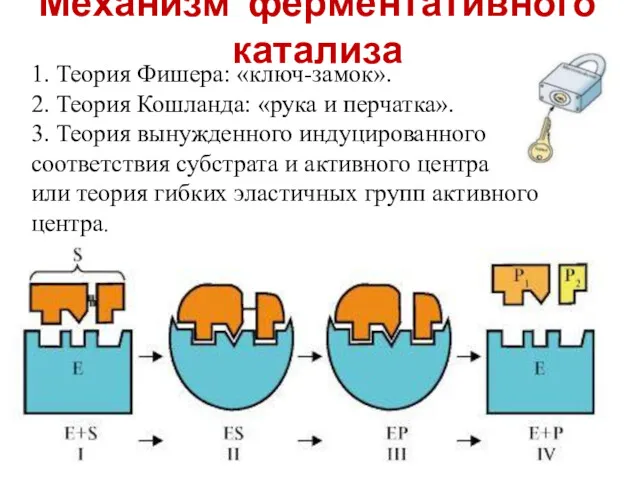
Механизм ферментативного катализа
1. Теория Фишера: «ключ-замок».
2. Теория Кошланда: «рука и перчатка».
3.
Механизм ферментативного катализа
1. Теория Фишера: «ключ-замок».
2. Теория Кошланда: «рука и перчатка».
3.
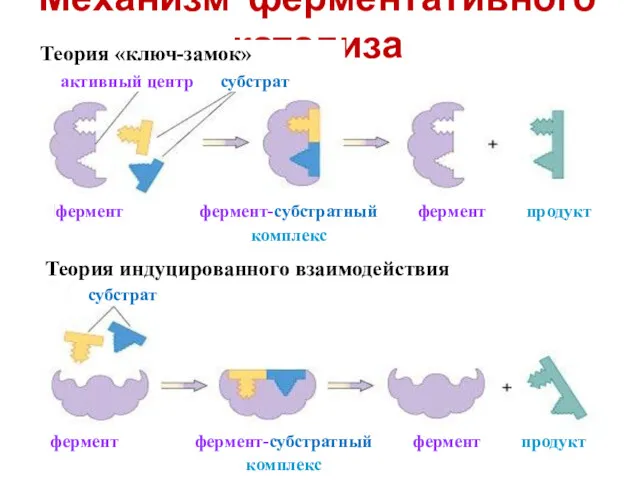
Механизм ферментативного катализа
Теория «ключ-замок»
Теория индуцированного взаимодействия
активный центр субстрат
фермент фермент-субстратный фермент
Механизм ферментативного катализа
Теория «ключ-замок»
Теория индуцированного взаимодействия
активный центр субстрат
фермент фермент-субстратный фермент
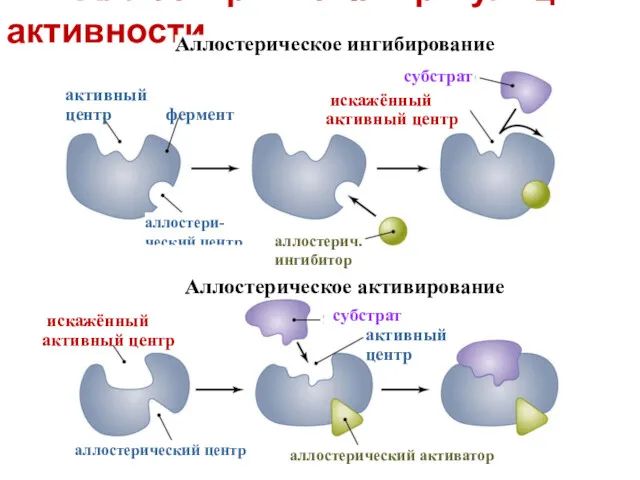
Аллостерическая регуляция активности
Аллостерическая регуляция активности
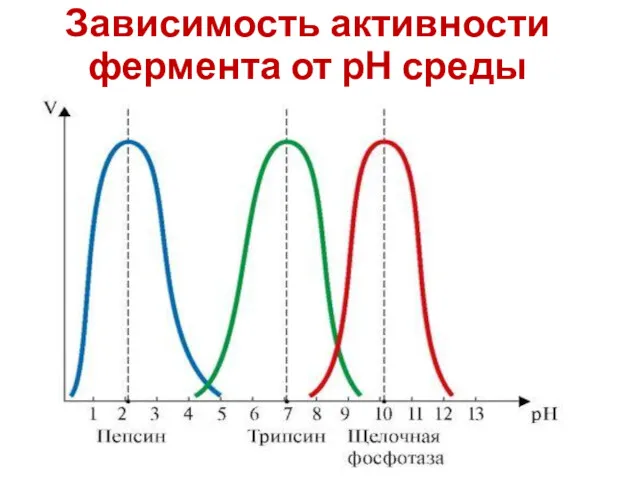
Зависимость активности фермента от рН среды
Зависимость активности фермента от рН среды
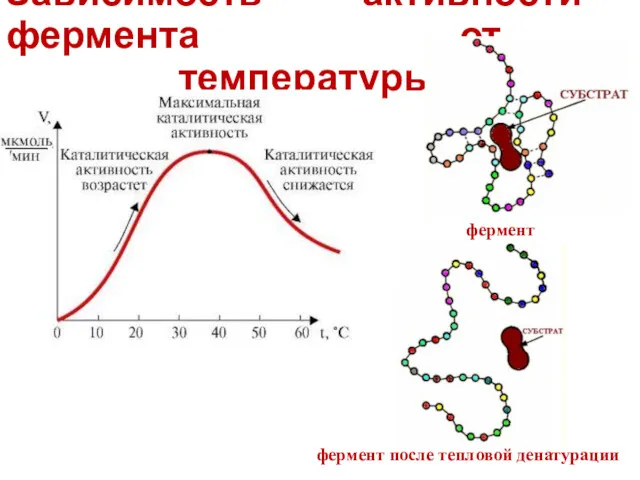
Зависимость активности фермента от температуры
фермент
фермент после тепловой денатурации
Зависимость активности фермента от температуры
фермент
фермент после тепловой денатурации

Классификация ферментов
Согласно современной классификации, выделяют шесть классов ферментов:
оксидоредуктазы;
трансферазы;
гидролазы;
лиазы;
изомеразы;
лигазы.
Классификация ферментов
Согласно современной классификации, выделяют шесть классов ферментов:
оксидоредуктазы;
трансферазы;
гидролазы;
лиазы;
изомеразы;
лигазы.
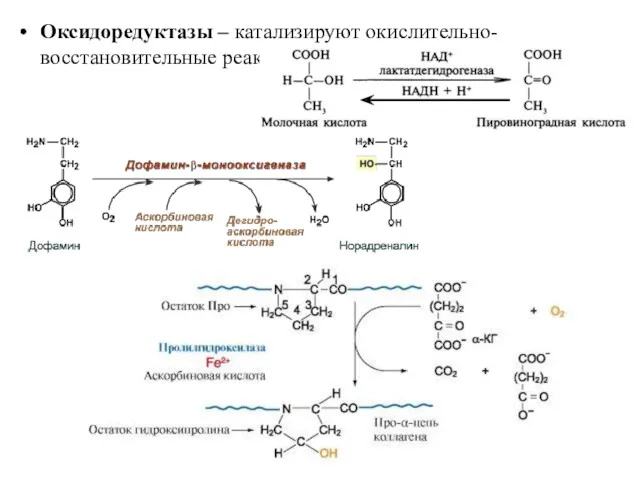
Оксидоредуктазы – катализируют окислительно-восстановительные реакции:
Оксидоредуктазы – катализируют окислительно-восстановительные реакции:
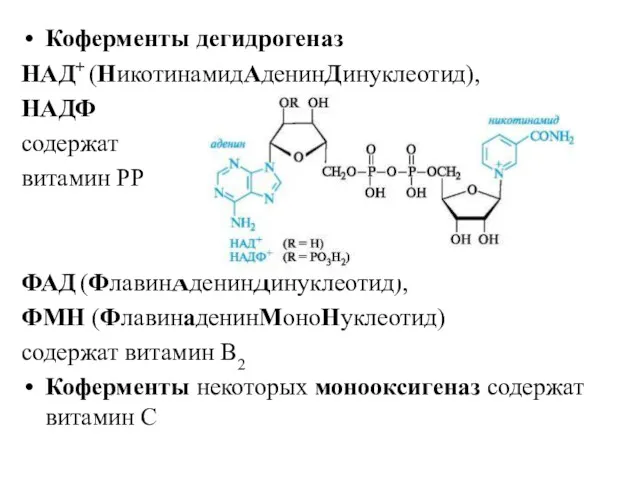
Коферменты дегидрогеназ
НАД+ (НикотинамидАденинДинуклеотид),
НАДФ
содержат
витамин РР
ФАД (ФлавинАденинДинуклеотид),
ФМН (ФлавинаденинМоноНуклеотид)
содержат витамин
Коферменты дегидрогеназ
НАД+ (НикотинамидАденинДинуклеотид),
НАДФ
содержат
витамин РР
ФАД (ФлавинАденинДинуклеотид),
ФМН (ФлавинаденинМоноНуклеотид)
содержат витамин
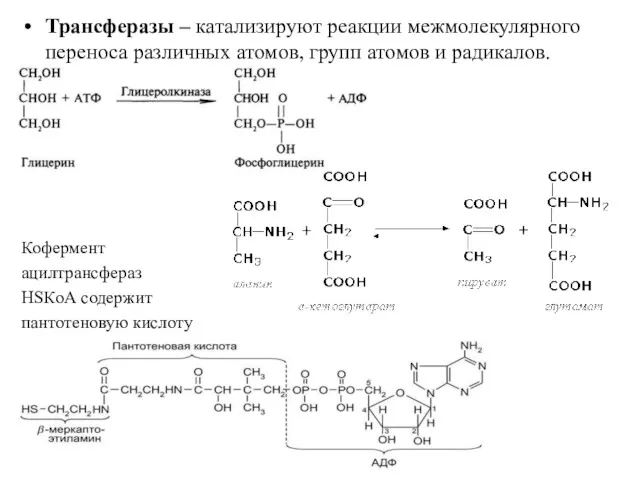
Трансферазы – катализируют реакции межмолекулярного переноса различных атомов, групп атомов и
Трансферазы – катализируют реакции межмолекулярного переноса различных атомов, групп атомов и
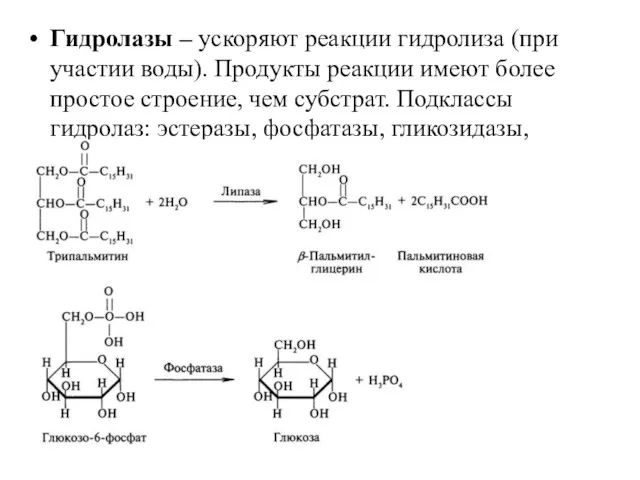
Гидролазы – ускоряют реакции гидролиза (при участии воды). Продукты реакции имеют
Гидролазы – ускоряют реакции гидролиза (при участии воды). Продукты реакции имеют
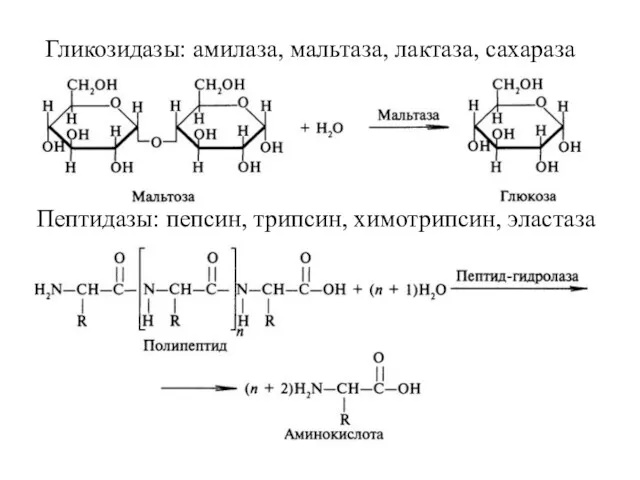
Гликозидазы: амилаза, мальтаза, лактаза, сахараза
Пептидазы: пепсин, трипсин, химотрипсин, эластаза
Гликозидазы: амилаза, мальтаза, лактаза, сахараза
Пептидазы: пепсин, трипсин, химотрипсин, эластаза
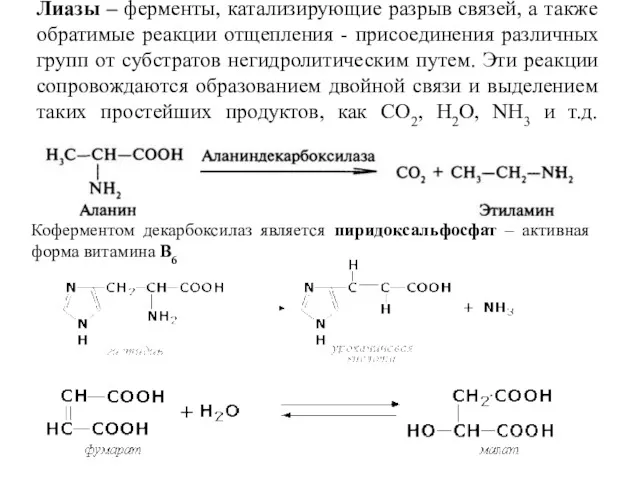
Лиазы – ферменты, катализирующие разрыв связей, а также обратимые реакции отщепления
Лиазы – ферменты, катализирующие разрыв связей, а также обратимые реакции отщепления
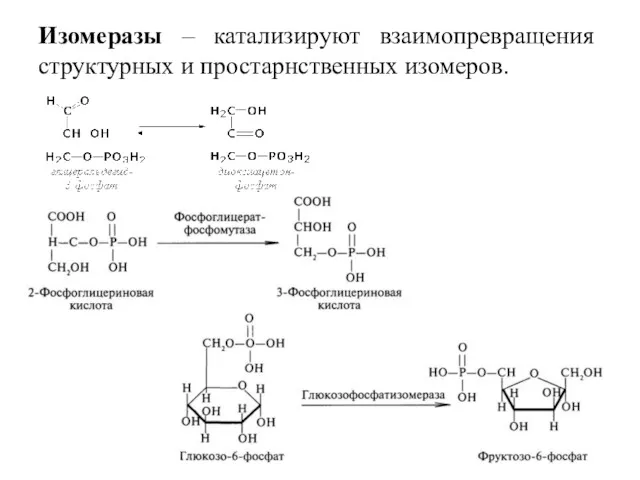
Изомеразы – катализируют взаимопревращения структурных и простарнственных изомеров.
Изомеразы – катализируют взаимопревращения структурных и простарнственных изомеров.

Лигазы (синтетазы) – катализируют синтез органических веществ из двух исходных молекул
Лигазы (синтетазы) – катализируют синтез органических веществ из двух исходных молекул
 Селекция и её достижения
Селекция и её достижения Живой мир Австралии
Живой мир Австралии Ведение в вирусологию
Ведение в вирусологию Red Wolf
Red Wolf Обмен белков-3
Обмен белков-3 Строение стебля
Строение стебля Морфология микроорганизмов. Прокариоты
Морфология микроорганизмов. Прокариоты Удивительные деревья
Удивительные деревья Системы органов. Опорно-двигательная. Строение кости. Скелет
Системы органов. Опорно-двигательная. Строение кости. Скелет Основные законы Менделя
Основные законы Менделя Красная книга
Красная книга Адаптация и здоровье.Психотипы человека и их склонность к возникновению заболеваний
Адаптация и здоровье.Психотипы человека и их склонность к возникновению заболеваний Класс Пресмыкающиеся, или рептилии
Класс Пресмыкающиеся, или рептилии Живая и неживая природа
Живая и неживая природа Слюнные железы
Слюнные железы Поленика арктическая и земляника садовая
Поленика арктическая и земляника садовая Цитотехнологія. Клітинна інженерія
Цитотехнологія. Клітинна інженерія Реализация системно-деятельностного подхода в рамках внеурочной деятельности
Реализация системно-деятельностного подхода в рамках внеурочной деятельности Зимующие птицы Башкирии
Зимующие птицы Башкирии Презентация по биологии Мхи, папоротники, хвощи и плауны для 6 класса.
Презентация по биологии Мхи, папоротники, хвощи и плауны для 6 класса. Отряд пресмыкающихся. Черепахи
Отряд пресмыкающихся. Черепахи Социальная психофизиология. Эволюционный путь генеза социальных отношений в сообществах живых организмов
Социальная психофизиология. Эволюционный путь генеза социальных отношений в сообществах живых организмов Что общего у разных растений
Что общего у разных растений Соотношение врожденного и приобретенного в поведении
Соотношение врожденного и приобретенного в поведении Dinoflagellates
Dinoflagellates Сцепленное наследование генов
Сцепленное наследование генов Потенциал действия
Потенциал действия Молекулярные основы наследственности
Молекулярные основы наследственности