- Структурная организация белковой молекулы. Лекция №1

Содержание
- 2. История кафедры биохимии Кафедра биологической химии возникла на основе кафедры физиологической химии Психоневрологического института в 1909
- 3. С1936 по 1946 г. кафедрой заведовал Александр Александрович Шмидт – крупнейший витаминолог страны. Им создан НИИ
- 4. С 1946 по 1948г. обязанности зав. кафедрой выполняла доц. Морозова Е.Н. С 1948 по 1967г. кафедру
- 5. С 1967 по 1986г. кафедру биохимии и биохимический отдел ЦНИЛ возглавлял проф. Соколовский Виктор Владимирович. В
- 6. Важным итогом этих исследований стала предложенная В.В. Соколовским концепция окислительно-восстановительного механизма неспецифической резистентности организма к действию
- 7. С 1987 по 2011г. кафедрой заведовал проф. Дадали Владимир Абдулович. В этот период расширены исследования в
- 8. С 2011 по 2014г. кафедрой руководил проф. Макаров Валерий Геннадиевич, занимающийся изучением механизмов лечебного действия природных
- 11. Типы химических связей, стабилизирующих структуру белка
- 12. Все аминокислоты по своим свойствам делятся на гидрофильные и гидрофобные. Последовательность аминокислот определена генетическим кодом в
- 13. Структура пептидной группы
- 14. Свойства пептидной группировки прочная ковалентная связь, разрушается только в процессе гидролиза четыре атома пептидной группировки лежат
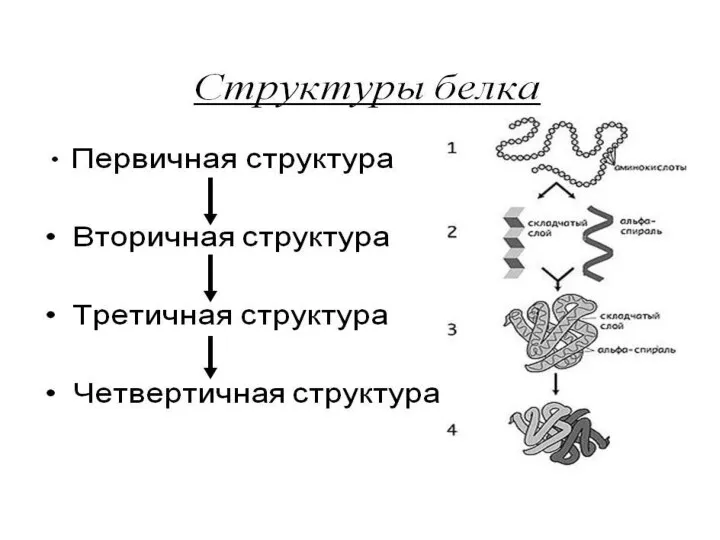
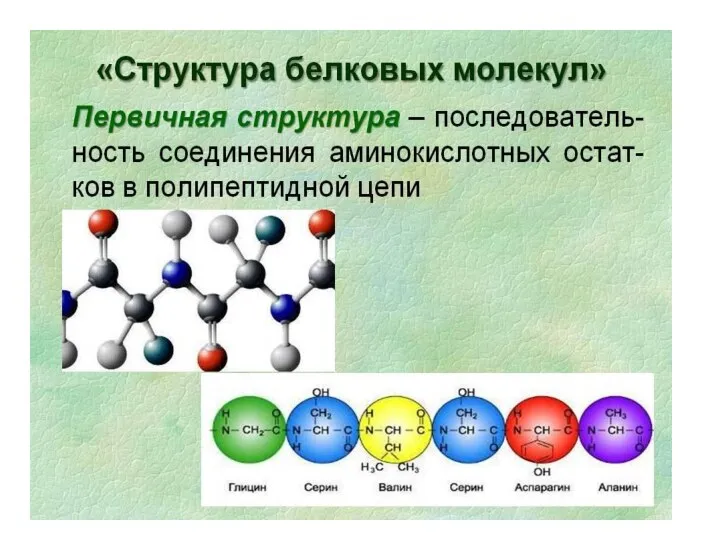
- 15. Роль первичной структуры белка определяет дальнейшую укладку белка в пространстве, т.е. вторичную, третичную и четвертичную структуру
- 16. При определении первичной структуры белков используются соединения, взаимодействующие с аминогруппой, с образованием окрашенного комплекса, например 2,4-динитрофторбензол
- 17. Вторичная структура белка α-спираль (модель Полинга-Кори) особенностей пептидной связи и удерживается водородными связями
- 18. За открытие вторичной структуры белка с использованием метода рентгеноструктурного анализа Л. Полинг получил нобелевскую премию (1954
- 20. α-Спираль правозакрученная по часовой стрелке вокруг оси (правый ход спирали обусловлен L-аминокислотами) Для каждого белка характерна
- 21. Факторы, нарушающие спирализацию включение пролина вызывает угол поворота цепи 135° заряженные радикалы аминокислот (электростатическое взаимодействие) объемные
- 22. Третичная структура белка в цитоплазме Ваша оценка "зачтено" (гидрофобные взаимодействия).
- 23. Строение коллагена Первичную структуру коллагена представляют в виде схемы ГЛИ – Х – У (30% -Гли,
- 25. Эластин Сшивки между остатками лизина трех или четырех пептидных цепей эластина образуют десмозин лиз лиз лиз
- 26. Роль третичной структуры Белок приобретает нативную форму и становится функционально активным за счет образования функциональных центров.
- 27. Шапероны Роль шаперонов первого типа -формирование нативной пространственной структуры белков или фолдинг. Они обеспечивают завершение формирования
- 28. Четвертичная структура белка , ионными.
- 29. Биологическая роль четвертичной структуры Комплекс белковых субъединиц в четвертичной структуре становится функционально активным Появление кооперативного эффекта
- 30. Кривая насыщения гемоглобина кислородом
- 31. Кривая насыщения гемоглобина и миоглобина кислородом
- 34. Функции белков Структурная Каталитическая (белки-ферменты) Регуляторная (гормоны белковой природы и регуляторные пептиды) Транспортная (альбумины и глобулины
- 36. Скачать презентацию

История кафедры биохимии
Кафедра биологической химии возникла на основе кафедры физиологической химии
История кафедры биохимии
Кафедра биологической химии возникла на основе кафедры физиологической химии

С1936 по 1946 г. кафедрой заведовал Александр Александрович Шмидт – крупнейший
С1936 по 1946 г. кафедрой заведовал Александр Александрович Шмидт – крупнейший

С 1946 по 1948г. обязанности зав. кафедрой выполняла доц. Морозова Е.Н.
С
С 1946 по 1948г. обязанности зав. кафедрой выполняла доц. Морозова Е.Н.
С

С 1967 по 1986г. кафедру биохимии и биохимический отдел ЦНИЛ возглавлял
С 1967 по 1986г. кафедру биохимии и биохимический отдел ЦНИЛ возглавлял

Важным итогом этих исследований стала предложенная В.В. Соколовским концепция окислительно-восстановительного механизма
Важным итогом этих исследований стала предложенная В.В. Соколовским концепция окислительно-восстановительного механизма

С 1987 по 2011г. кафедрой заведовал проф. Дадали Владимир Абдулович. В
С 1987 по 2011г. кафедрой заведовал проф. Дадали Владимир Абдулович. В

С 2011 по 2014г. кафедрой руководил проф. Макаров Валерий Геннадиевич, занимающийся
С 2011 по 2014г. кафедрой руководил проф. Макаров Валерий Геннадиевич, занимающийся
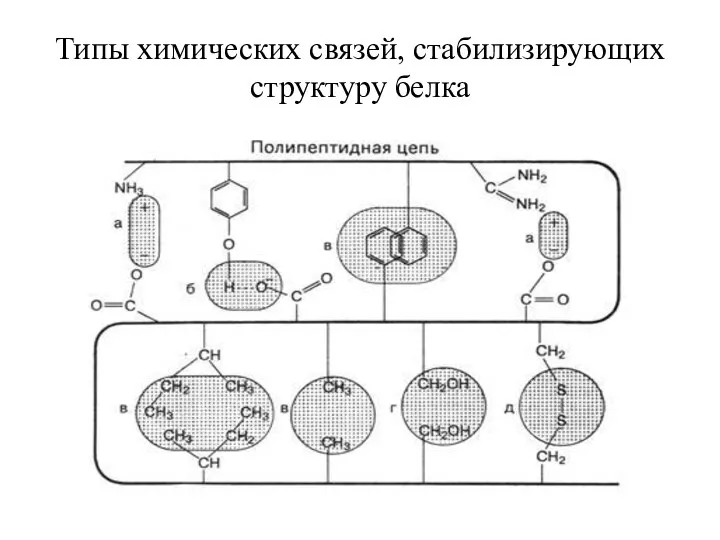
Типы химических связей, стабилизирующих структуру белка
Типы химических связей, стабилизирующих структуру белка

Все аминокислоты по своим свойствам делятся на гидрофильные и гидрофобные.
Последовательность аминокислот
Все аминокислоты по своим свойствам делятся на гидрофильные и гидрофобные.
Последовательность аминокислот
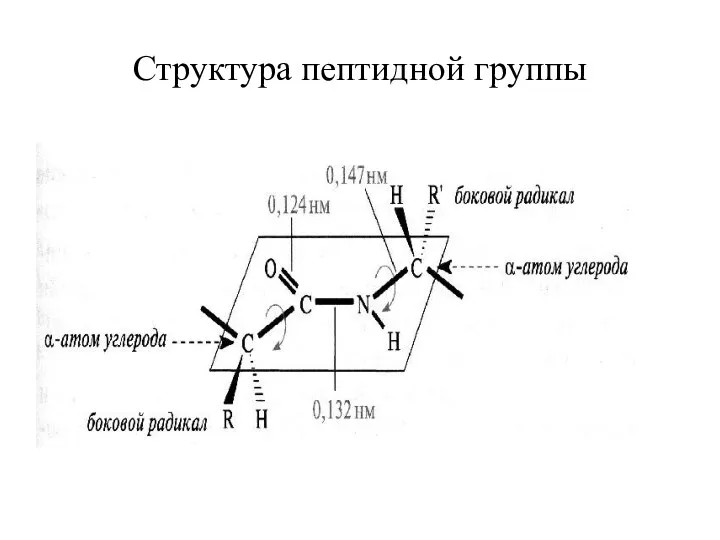
Структура пептидной группы
Структура пептидной группы

Свойства пептидной группировки
прочная ковалентная связь, разрушается только в процессе гидролиза
четыре атома
Свойства пептидной группировки
прочная ковалентная связь, разрушается только в процессе гидролиза
четыре атома

Роль первичной структуры белка
определяет дальнейшую укладку белка в пространстве, т.е. вторичную,
Роль первичной структуры белка
определяет дальнейшую укладку белка в пространстве, т.е. вторичную,

При определении первичной структуры белков используются соединения, взаимодействующие с аминогруппой, с
При определении первичной структуры белков используются соединения, взаимодействующие с аминогруппой, с

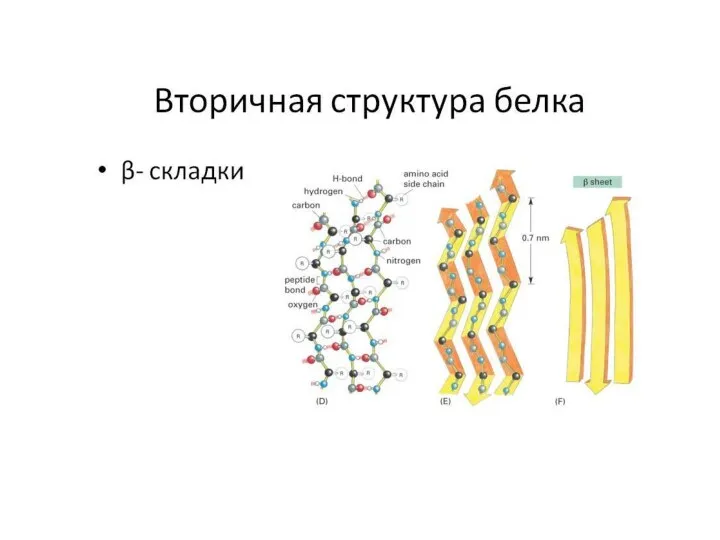
Вторичная структура белка
α-спираль (модель Полинга-Кори)
особенностей пептидной связи и удерживается водородными связями
Вторичная структура белка
α-спираль (модель Полинга-Кори)
особенностей пептидной связи и удерживается водородными связями

За открытие вторичной структуры белка с использованием метода рентгеноструктурного анализа
Л.
За открытие вторичной структуры белка с использованием метода рентгеноструктурного анализа
Л.

α-Спираль правозакрученная по часовой стрелке вокруг оси (правый ход спирали обусловлен
α-Спираль правозакрученная по часовой стрелке вокруг оси (правый ход спирали обусловлен

Факторы, нарушающие спирализацию
включение пролина вызывает угол поворота цепи 135°
заряженные радикалы аминокислот
Факторы, нарушающие спирализацию
включение пролина вызывает угол поворота цепи 135°
заряженные радикалы аминокислот
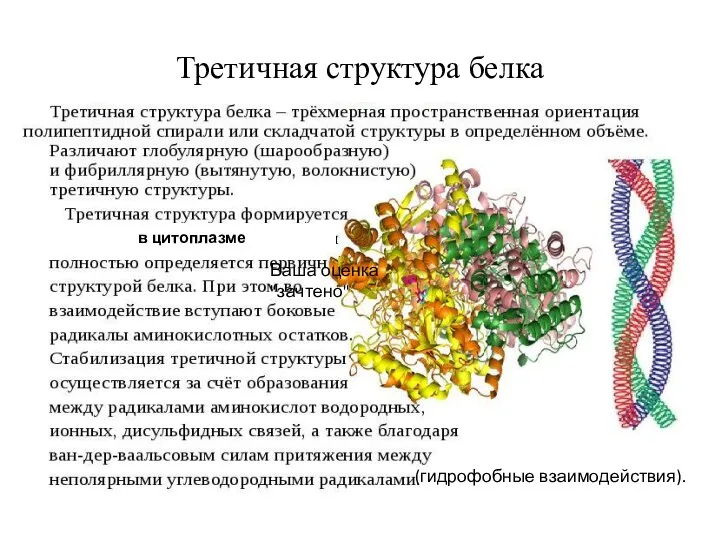
Третичная структура белка
в цитоплазме
Ваша оценка "зачтено"
(гидрофобные взаимодействия).
Третичная структура белка
в цитоплазме
Ваша оценка "зачтено"
(гидрофобные взаимодействия).

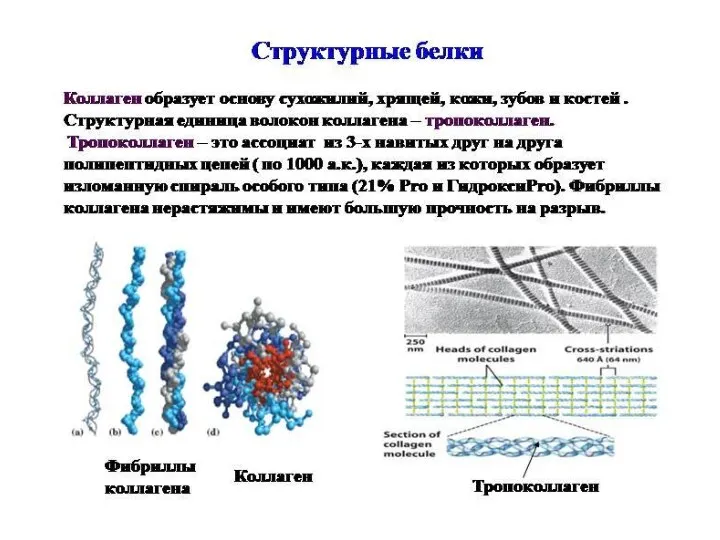
Строение коллагена
Первичную структуру коллагена представляют в виде схемы ГЛИ – Х
Строение коллагена
Первичную структуру коллагена представляют в виде схемы ГЛИ – Х
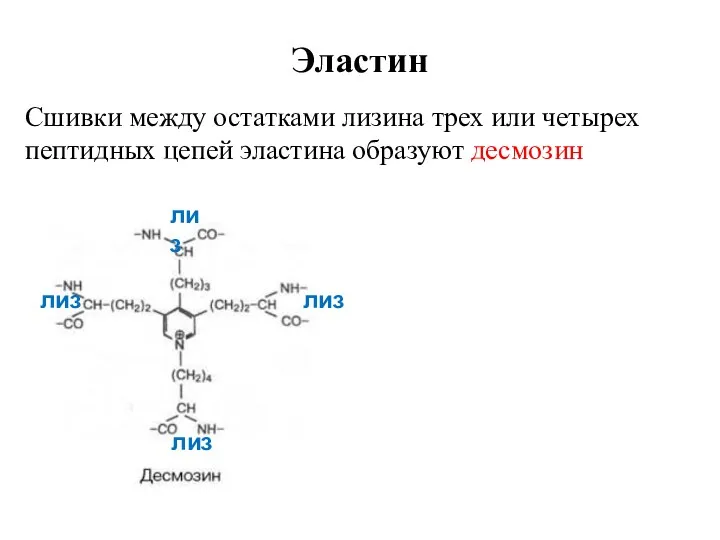
Эластин
Сшивки между остатками лизина трех или четырех пептидных цепей эластина образуют
Эластин
Сшивки между остатками лизина трех или четырех пептидных цепей эластина образуют

Роль третичной структуры
Белок приобретает нативную форму и становится функционально активным за
Роль третичной структуры
Белок приобретает нативную форму и становится функционально активным за

Шапероны
Роль шаперонов первого типа -формирование нативной пространственной структуры белков или фолдинг.
Шапероны
Роль шаперонов первого типа -формирование нативной пространственной структуры белков или фолдинг.

Четвертичная структура белка
, ионными.
Четвертичная структура белка
, ионными.

Биологическая роль четвертичной структуры
Комплекс белковых субъединиц в четвертичной структуре становится функционально
Биологическая роль четвертичной структуры
Комплекс белковых субъединиц в четвертичной структуре становится функционально
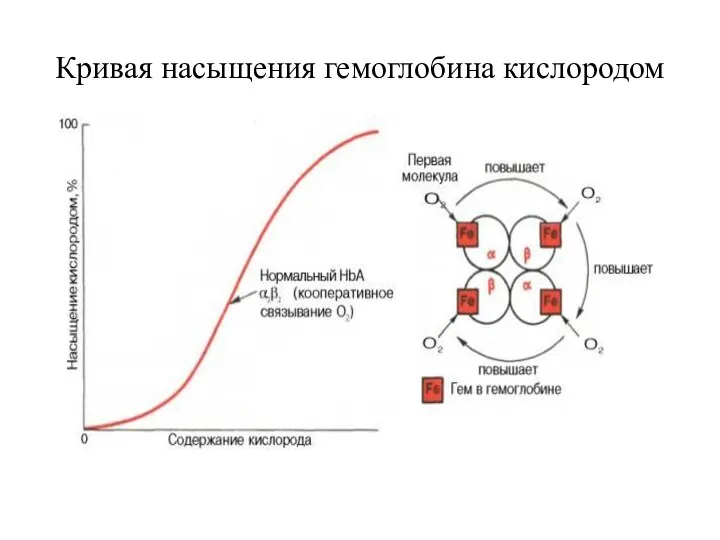
Кривая насыщения гемоглобина кислородом
Кривая насыщения гемоглобина кислородом
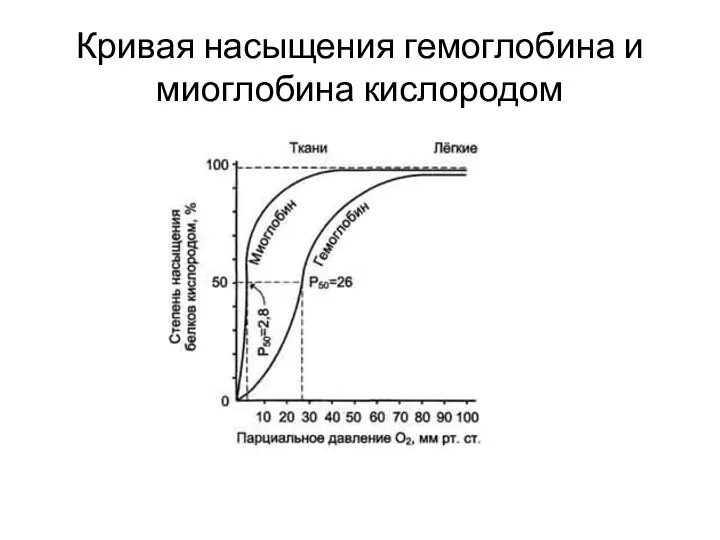
Кривая насыщения гемоглобина и миоглобина кислородом
Кривая насыщения гемоглобина и миоглобина кислородом



Функции белков
Структурная
Каталитическая (белки-ферменты)
Регуляторная (гормоны белковой природы и регуляторные пептиды)
Транспортная (альбумины
Функции белков
Структурная
Каталитическая (белки-ферменты)
Регуляторная (гормоны белковой природы и регуляторные пептиды)
Транспортная (альбумины
 Викторина по биологии
Викторина по биологии Культивирование красных водорослей
Культивирование красных водорослей Рослини. Тварини. Гриби. Бактерії. Віруси
Рослини. Тварини. Гриби. Бактерії. Віруси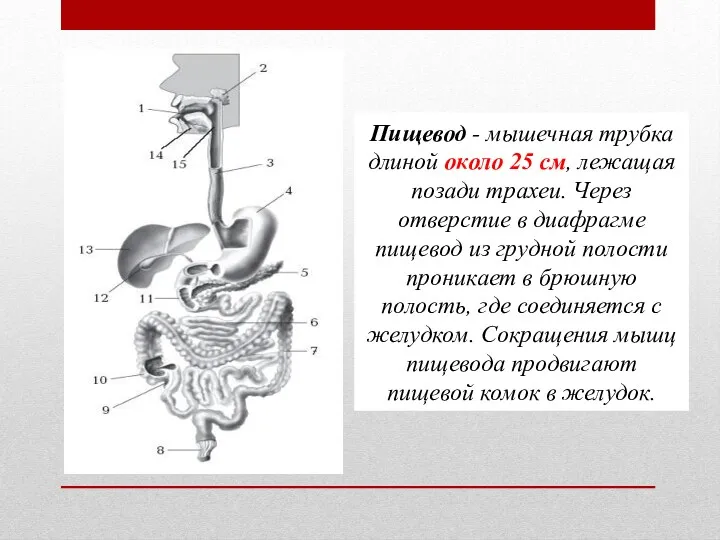 Устройство пищевода
Устройство пищевода Механизмы регуляции пролиферативных процессов клетках
Механизмы регуляции пролиферативных процессов клетках Свойства живого вещества
Свойства живого вещества Пагін. Його будова
Пагін. Його будова Физиология центральной нервной системы. Лекция № 8
Физиология центральной нервной системы. Лекция № 8 Химический состав клетки. Тема урока №1, №2
Химический состав клетки. Тема урока №1, №2 Белоктың трансляциясы
Белоктың трансляциясы Тип хордовые. Общая характеристика. Подтип бесчерепные. Ланцетник
Тип хордовые. Общая характеристика. Подтип бесчерепные. Ланцетник Генетика. Становление и роль в современном естествознании
Генетика. Становление и роль в современном естествознании Устройство речевого аппарата
Устройство речевого аппарата Плоские черви. (Лекция 4)
Плоские черви. (Лекция 4) Молекулярная генетика. Нуклеиновые кислоты
Молекулярная генетика. Нуклеиновые кислоты Тип Губки
Тип Губки Клонирование в бактериальных геномах
Клонирование в бактериальных геномах Индивидуальное развитие организмов. Биогенетический закон
Индивидуальное развитие организмов. Биогенетический закон Сердечно-сосудистая система рыб
Сердечно-сосудистая система рыб Знаешь ли ты деревья
Знаешь ли ты деревья Способы деления бактериальных клеток и жизненные циклы бактерий
Способы деления бактериальных клеток и жизненные циклы бактерий Условия прорастания семян
Условия прорастания семян Физиология носа и околоносовых пазух
Физиология носа и околоносовых пазух Слава воде. Организм человека и вода
Слава воде. Организм человека и вода Мышцы туловища. Мышцы головы и шеи
Мышцы туловища. Мышцы головы и шеи Морфология и структура вирусов
Морфология и структура вирусов Соматический и автономный (вегетативный) отделы нервной системы
Соматический и автономный (вегетативный) отделы нервной системы Арктический, или полярный волк
Арктический, или полярный волк