Структурно-функциональная организация наследственного материала. Реализация генетической информации в признак презентация
- Структурно-функциональная организация наследственного материала. Реализация генетической информации в признак

Содержание
- 2. План: 1. Уровни организации наследственного материала. 2. Классификация генов. 3. Химическая и структурная организация гена. 4.
- 3. Материальная основа наследственности – нуклеиновые кислоты (генетический материал). Общие свойства генетического материала: Способность к самовоспроизведению. Способность
- 4. ИСТОРИЯ ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЙ ОБ ОРГАНИЗАЦИИ МАТЕРИАЛЬНОГО СУБСТРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ В 1865 Г. Мендель - высказал
- 5. ИСТОРИЯ ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЙ ОБ ОРГАНИЗАЦИИ МАТЕРИАЛЬНОГО СУБСТРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ 1953 - создание Дж. Уотсоном и
- 6. В 60-х гг. - произведена полная расшифровка генетического кода, установлено соответствие триплетов нуклеотидов в молекуле нуклеиновых
- 7. Сиквенирование-заводской процесс более1,5 млрд п.н. в месяц Сиквенс генома человека занял 9 месяцев 10 дней И
- 8. Уровни организации наследственного материала
- 9. Уровни организации генетического материала: 1. Генный. Ген – минимальная структурно-функциональная единица наследственности, которая кодирует определенный признак.
- 10. Генный уровень организации наследственного материала ген – это элементарная единица молекулярно-генетического уровня организации Ген – участок
- 11. Особенности структурной организации ДНК. Полимерная молекула, мономером которой является н у к л е о т
- 12. Азотистые основания Пуриновые — аденин, гуанин и пиримидиновые — тимин, цитозин
- 13. Нуклеиновые кислоты (НК) Полинуклеотидные цепи спирально закручены друг около друга и вместе вокруг воображаемой оси Диаметр
- 14. Нуклеиновые кислоты (НК) "правило Чаргаффа": в любом фрагменте ДНК содержание остатков гуанина всегда соответствует содержанию цитозина,
- 15. Названия нуклеотидов (А, Т, Г, Ц): Аденин – адениловый; гуанин – гуаниловый; цитозин – цитидиловый; тимин
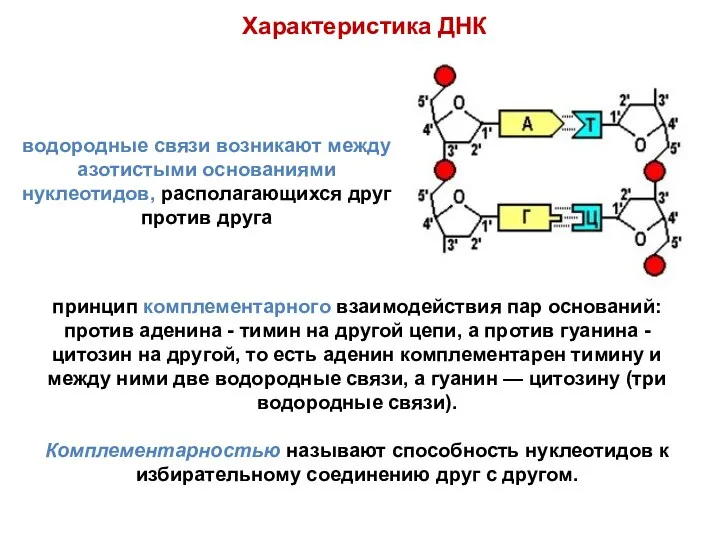
- 16. между 3'-углеродом остатка сахара одного нуклеотида и остатком фосфорной кислоты другого возникает фосфодиэфирная связь. Характеристика ДНК
- 17. водородные связи возникают между азотистыми основаниями нуклеотидов, располагающихся друг против друга принцип комплементарного взаимодействия пар оснований:
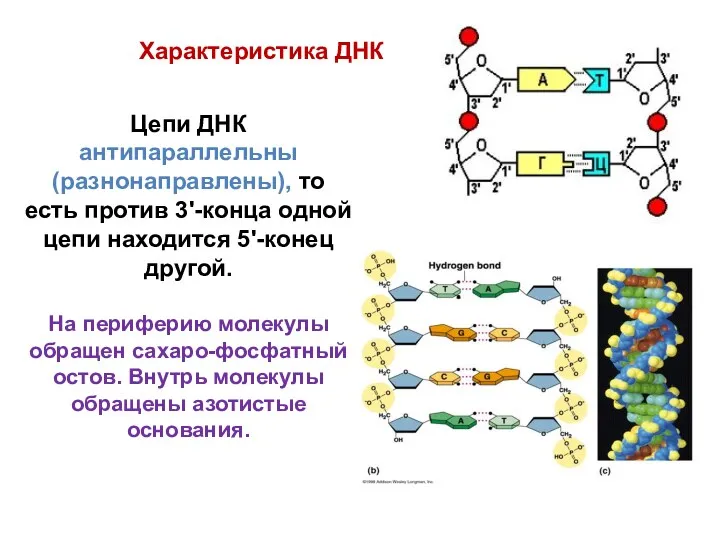
- 18. Цепи ДНК антипараллельны (разнонаправлены), то есть против 3'-конца одной цепи находится 5'-конец другой. На периферию молекулы
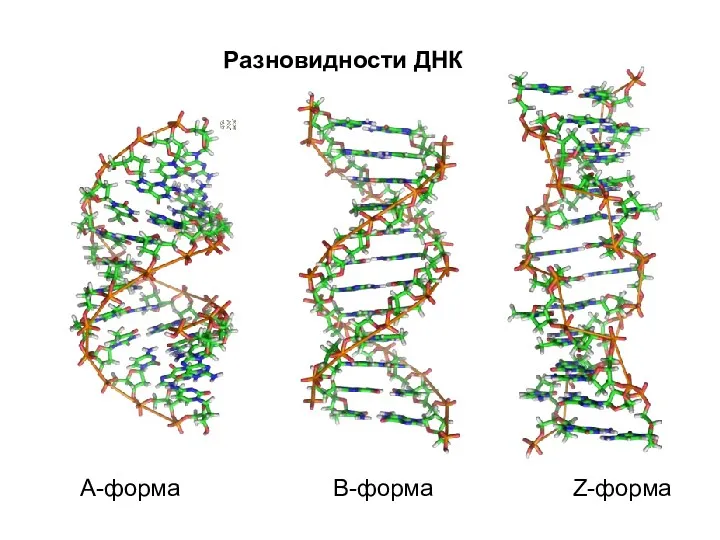
- 19. А-форма В-форма Z-форма Разновидности ДНК
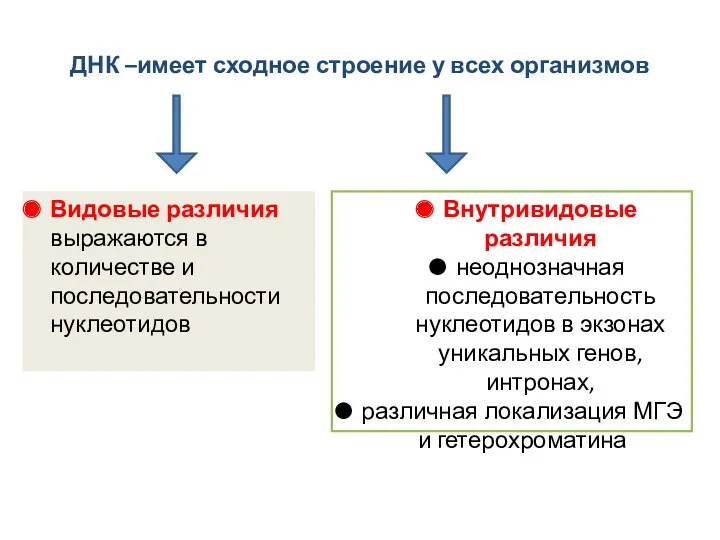
- 20. ДНК –имеет сходное строение у всех организмов Видовые различия выражаются в количестве и последовательности нуклеотидов Внутривидовые
- 21. ГЕНЕТИЧЕСКИЙ код и его свойства Особая система записи минокисло белка в виде нуклеотидов на молекуле ДНК
- 22. Каждая аминокислота полипептидной цепи кодируется сочетанием трех последовательно расположенных в цепи ДНК нуклеотидов, называемых ТРИПЛЕТАМИ или
- 23. Свойства генетического кода Специфичность - Каждый триплет способен кодировать только одну определенную аминокислоту. Универсальность - полное
- 24. Классификация генов 1. Структурные гены (3-5% ДНК): Уникальные (их продукты белки: ферменты, транспортные, строительные, рецепторные). много
- 25. Свойства генов: 1. Лабильность – способность генов мутировать – изменять нуклеотидную последовательность. Есть гены, мутации которых
- 26. Гены человека, встречающиеся в человеческих популяциях в разных молекулярных формах 1. Гены эритроцитарных антигенов систем: АВО:
- 27. Свойства генов: 4. Аллельное состояние – характерно для диплоидных организмов. 5. Специфичность – каждый ген контролирует
- 28. Пенентрантность гена – это вероятность проявления гена Пенентрантность гена выражают в % случаев его проявления к
- 29. Экспрессивность – степень выраженности гена (ген проявляется у всех гетерозигот, но в разной степени) Понятие экспрессивности
- 30. Строение генов про- и эукариот
- 31. Оперон прокариот Несколько структурных генов Ген-регулятор Ген-оператор P O S1S2S3Sn это непрерывная последовательность кодирующих нуклеотидов. Функциональная
- 32. Структура гена прокариот (ДНК-овых) Непрерывная последовательность кодирующих нуклеотидов ДНК АЦЦ-ГАТ-ТАТ-ЦЦА-АЦЦ…АТТ… ТРАНСКРИПЦИЯ иРНК УГГ-ЦУА-АУА-ГГУ-УГГ…УАА… ТРАНСЛЯЦИЯ ТРИ-ЛЕЙ-ИЛЕ-ГЛИ-ТРИ Порядок
- 33. Гены эукариот имеют мозаичную структуру: состоят из кодирующих (экзонов) и некодирующих (интронов) участков Например, ген фенилаланингидрооксилазы
- 34. Уникальные нуклеотидные последовательности представлены единичными копиями на геном, в них локализованы в основном структурные гены ключевых
- 35. Функциональная единица генома эукариот включает один структурный ген (транскрибируемая зона) и множество регуляторных участков ДНК (промотор
- 36. Структурная организация гена эукариот: Экзоны – нуклеотидные последовательности, кодирующие аминокислоты. Интроны – не кодирующие нуклеотидные последовательности
- 37. хромосомный уровень организации наследственного материала Хромосомы – это надмолекулярный комплекс ДНК и белков– ДНП ДНП состоит
- 38. хромосомы – ДНП ядра (в соматических клетках человека- 46, ХХ или 46, ХУ, в гаметах 23,
- 39. Основным компонентом интерфазного ядра является (окрашенное вещество). Впервые онХРОМАТИН описан ещё в 1880 году В. Флеммингом.
- 40. УРОВНИ КОМПАКТИЗАЦИИ ДНП. ИНТЕРФАЗНАЯ ХРОМОСОМА (три уровня: нуклеосомный, фибриллярный и хромонемный) Интерфазная хромосома – функционально активная
- 41. УРОВНИ КОМПАКТИЗАЦИИ ДНП: 1. НУКЛЕОСОМНЫЙ 4 класса гистоновых белков (из групп Н3, Н4, Н2А, Н2В) формируют
- 42. УРОВНИ КОМПАКТИЗАЦИИ ДНП: 2. ФИБРИЛЛЯРНЫЙ При образовании фибриллы нуклеосомы сближаются за счет разности зарядов «хвоста» гистона
- 43. УРОВНИ КОМПАКТИЗАЦИИ ДНП: 3. ХРОМОНЕМНЫЙ Образование вытянутых и компактных петель Каждая из петель содержит от 20000
- 44. УРОВНИ КОМПАКТИЗАЦИИ ДНП: 4. МЕТАФАЗНАЯ ХРОМОСОМА Метафазная хромосома – функционально не активна, максимально конденсирована, различима в
- 45. Длина деконденсированной ДНК от нескольких сантиметров до 1.2-2м. Длина митотической хромосомы измеряется микронами. В результате степень
- 46. В соматических клетках ♀♀ на периферии ядра глыбка хроматина – тельце Барра или Х-хроматин гетерохроматизированная одна
- 47. Геномный уровень организации наследственного материала Вся совокупность наследственного материала, заключенного в гаплоидном наборе хромосом данного вида
- 48. Геном прокариот (на примере кишечной палочки) В нуклеоиде 1 кольцевая хромосома (n) 2,5 тыс. генов в
- 49. длина ДНК в ядре клетки млекопитающего составляет примерно 2—5·109 пар нуклеотидов, т.е. в 1000 раз превосходит
- 50. Наследственный аппарат клеток человека состоит из внеядерного – митохондриального и ядерного
- 51. Геном митохондрий человека ДНК двуспиральная Кольцевая 16569 НП 37 генов митохондриальных белков 2 гена рРНК 22
- 52. Внеядерный аппарат Хромосомы митохондрий распределяются в митозе и мейозе неравномерно, обеспечивают цитоплазматическую наследственность, не подчиняющуюся законам
- 53. Ядерный (основной) аппарат клеток человека представлен хромосомами – ДНП ядра (в соматических клетках - 46, ХХ
- 54. Ядерный (основной) аппарат клеток человека В митозе и мейозе ядерные хромосомы сегрегируют к полюсам клетки, а
- 55. Кариотип 2n Шимпанзе – 48 Лошадь – 64 Свинья и кошка – 38 Собака – 78
- 56. 46, ХХ ♀ норма 46, ХУ ♂ Кариотип – совокупность данных о числе, размерах и структуре
- 57. Кариотипический анализ – определение кариотипа и идиограммы. Исследуют кариотип человека в окрашенных микропрепаратах клеток на стадии
- 59. КАРИОТИПИЧЕСКИЙ АНАЛИЗ С учетом морфологических параметров аутосомы человека классифицированы на 7 групп (А – 3 пары,
- 60. Методы дифференциального окрашивания хромосом позволяют индивидуализировать все хромосомы в кариотипе Методы дифференциального окрашивания хромосом основаны на
- 61. Световая микрофотография участка политенной хромосомы из клетки слюнной железы дрозофилы. Видна характерная картина распределения дисков. Эти
- 62. Кариотип Chironomus sp. Картирование хромосом представлено по системе Кейла [Keyl, Keyl, 1962] и Девай с соавторами
- 63. Идиограмма – графическое изображение хромосом с учетом их абсолютной и относительной длины, центромерного индекса, наличия вторичной
- 64. В зависимости от соотношения плеч – 3 типа хромосом Метацентрические Субметацентрические Акроцентрические Центромерный индекс (отношение длины
- 65. Теломеры на концах митотической хромосомы
- 66. 47, ХХХ ♀ 47, ХХУ ♂ патология 45, Х0 ♀ Синдром Шерешевского – Тернера – 45,
- 67. 47, ХХХ ♀ - 2 тельца Барра 47, ХХУ ♂ патология - 1 тельце Барра и
- 68. Синдром Патау (трисомия по 13 хр.) 47, 13 13 13 Хромосомы 18 трисомии синдром(с. Эдвардса) 47,
- 69. Основные этапы реализации генетической информации
- 70. результат взаимодействия наследственности и среды Формирование признака – Признак – свойство организма биохимического, физиологического или морфологического
- 71. ПРИЗНАК (на молекулярном уровне)
- 72. ЭКСПРЕССИЯ ГЕНОВ От лат. - EXPRESSIO – выражение, сила проявления. Процесс реализации генетической информации
- 73. Основные этапы экспрессии генов у ДНК-вых прокариот: 1 – транскрипция (синтез иРНК на матрице ДНК), 2
- 74. Основные этапы экспрессии генов у эукариот: 1 – транскрипция (синтез про-иРНК на матрице ДНК), 2 –
- 75. Основные этапы экспрессии генов эукариот ДНК про-иРНК иРНК белок (полипептид) Транскрипция (прямая) Процессинг, сплайсинг Трансляция Этапы
- 76. Регуляторы активности генов в претранскрипционный период Внутриклеточные pH ± Ионы ± Белки (гистоновые, негистоновые) ± Метаболиты
- 77. Функции регуляторов генной активности (в зоне действия) Освобождают ДНК от белков Деконденсация ДНП Ослабляют водородные связи
- 78. 2. Транскрипция начинается с присоединения РНК-полимеразы к «своему» промотору (ТАТА-блоку). происходит с одной полинуклеотидной цепи ДНК
- 79. РНК-полимераза – фермент транскрипции
- 80. Прямая транскрипция – синтез РНК на матрице ДНК ДНК РНК Белок (1958 год – центральная догма
- 81. 3. Процессинг про-иРНК: 1) Кэпирование – образование КЭП на 5’-конце про-иРНК. «Кэпирование» - присоединение к 5’-концу
- 83. Аномальный процессинг болезнь Альцгеймера
- 84. 4. Этап экспрессии генов Транспорт зрелой иРНК через ядерную мембрану
- 85. 5. Трансляция. Зрелая иРНК 5’-концом (КЭП) подходит к полисомам и протягивается через них транслируются кодон-инициатор и
- 86. Компоненты, необходимые для трансляции Зрелая иРНК Рибосомы (полисомы) Набор тРНК (~60) Набор аминокислот Набор ферментов
- 87. Строение рибосом Рибосомы (0,2 мкм ~ 60% белков, 40% - РНК (3 вида))
- 88. тРНК Имеет конфигурацию «клеверного листа», содержит 80 нукл., много минор. основ, поэтому петли: Антикодоновая (7 нукл.,
- 89. структура - тРНК вместе с аминокислотой называется аминоцил-тРНК.
- 90. Биосинтез на рибосоме Аминоцил т-РНК входит в рибосому, комплементарно связываясь с кодоном мРНК, происходит реакция, при
- 91. 6. Посттрансляционный этап – образование функционально активного белка: у эукариот «отрезается» метионин или триптофан, формируется вторичная,
- 93. Скачать презентацию

План:
1. Уровни организации наследственного материала.
2. Классификация генов.
3. Химическая и структурная
План: 1. Уровни организации наследственного материала. 2. Классификация генов. 3. Химическая и структурная
Материальная основа наследственности – нуклеиновые кислоты
(генетический материал).
Общие свойства генетического материала:
Способность
Материальная основа наследственности – нуклеиновые кислоты
(генетический материал).
Общие свойства генетического материала:
Способность

ИСТОРИЯ ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЙ ОБ ОРГАНИЗАЦИИ МАТЕРИАЛЬНОГО СУБСТРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
В
ИСТОРИЯ ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЙ ОБ ОРГАНИЗАЦИИ МАТЕРИАЛЬНОГО СУБСТРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
В

ИСТОРИЯ ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЙ ОБ ОРГАНИЗАЦИИ МАТЕРИАЛЬНОГО СУБСТРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
1953 -
ИСТОРИЯ ФОРМИРОВАНИЯ ПРЕДСТАВЛЕНИЙ ОБ ОРГАНИЗАЦИИ МАТЕРИАЛЬНОГО СУБСТРАТА НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ
1953 -

В 60-х гг. - произведена полная расшифровка генетического кода,
установлено соответствие
В 60-х гг. - произведена полная расшифровка генетического кода,
установлено соответствие

Сиквенирование-заводской процесс
более1,5 млрд п.н. в месяц
Сиквенс генома человека занял 9
более1,5 млрд п.н. в месяц
Сиквенс генома человека занял 9

Уровни организации наследственного материала
Уровни организации наследственного материала
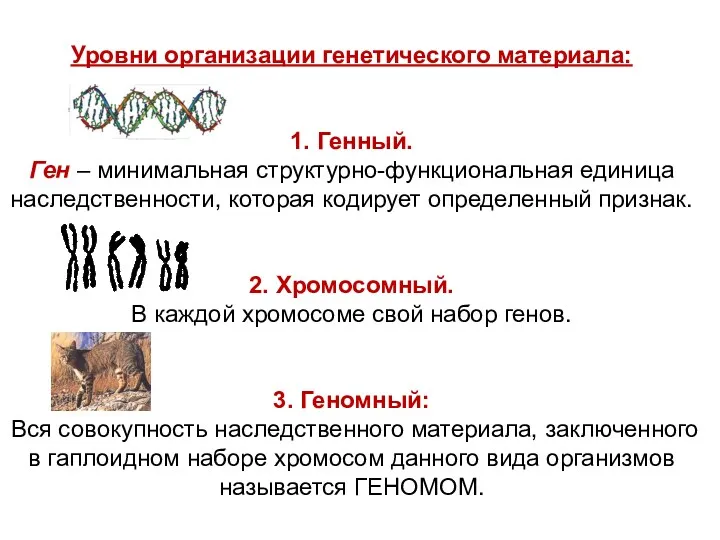
Уровни организации генетического материала:
1. Генный.
Ген – минимальная структурно-функциональная единица наследственности,
Уровни организации генетического материала:
1. Генный.
Ген – минимальная структурно-функциональная единица наследственности,

Генный уровень организации наследственного материала
ген – это элементарная единица молекулярно-генетического уровня
Генный уровень организации наследственного материала
ген – это элементарная единица молекулярно-генетического уровня
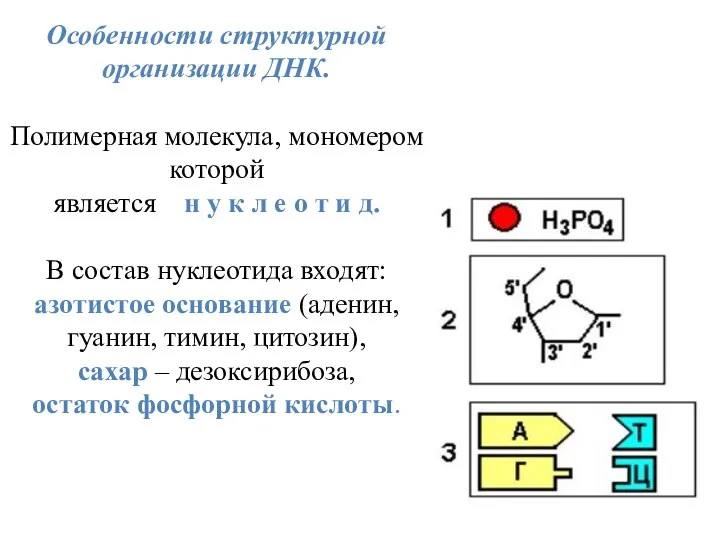
Особенности структурной организации ДНК.
Полимерная молекула, мономером которой
является н у к
Особенности структурной организации ДНК.
Полимерная молекула, мономером которой
является н у к
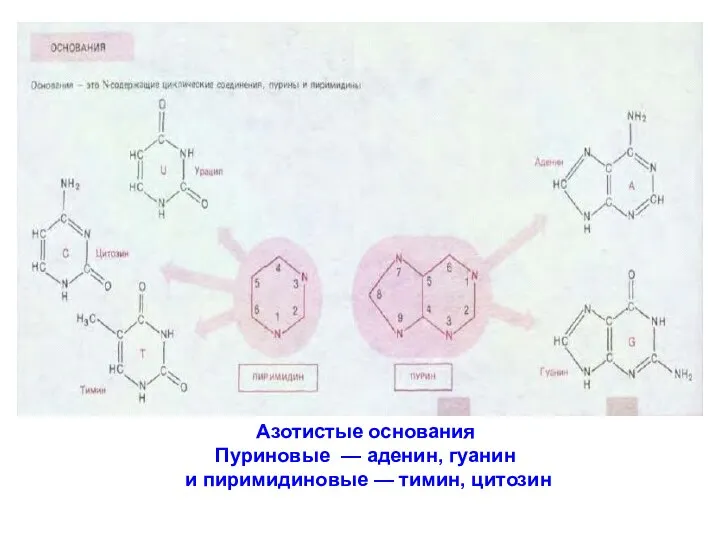
Азотистые основания
Пуриновые — аденин, гуанин
и пиримидиновые — тимин, цитозин
Азотистые основания
Пуриновые — аденин, гуанин
и пиримидиновые — тимин, цитозин
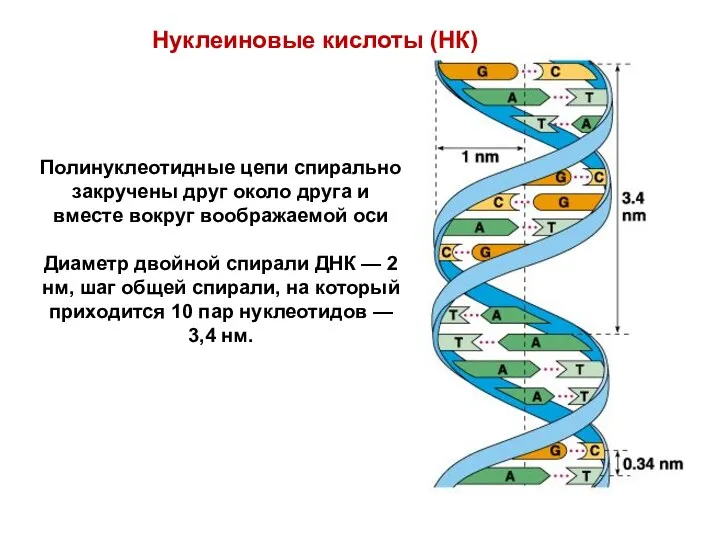
Нуклеиновые кислоты (НК)
Полинуклеотидные цепи спирально закручены друг около друга и вместе
Нуклеиновые кислоты (НК)
Полинуклеотидные цепи спирально закручены друг около друга и вместе
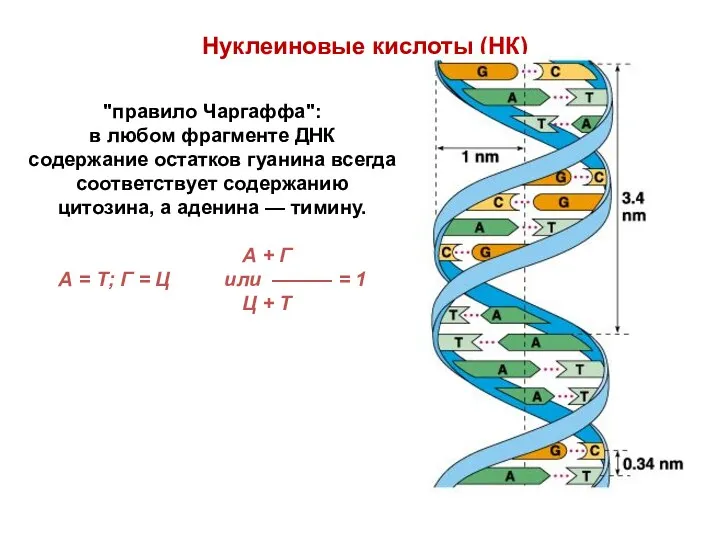
Нуклеиновые кислоты (НК)
"правило Чаргаффа":
в любом фрагменте ДНК содержание остатков гуанина всегда
Нуклеиновые кислоты (НК)
"правило Чаргаффа":
в любом фрагменте ДНК содержание остатков гуанина всегда
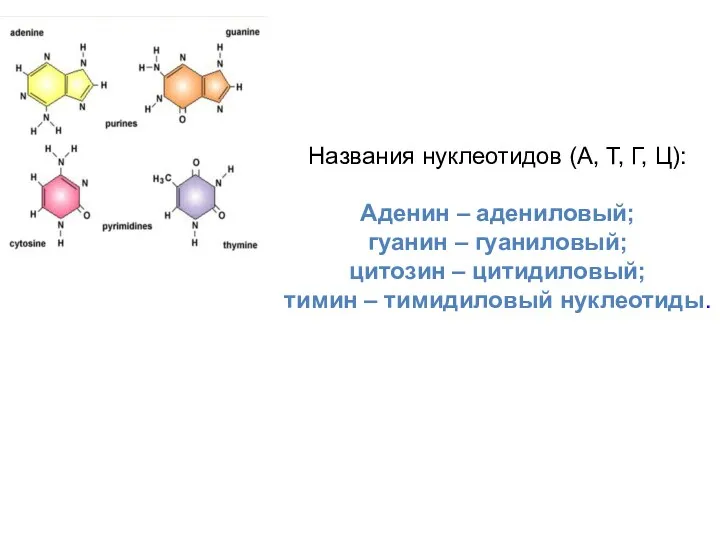
Названия нуклеотидов (А, Т, Г, Ц):
Аденин – адениловый;
гуанин – гуаниловый;
Названия нуклеотидов (А, Т, Г, Ц):
Аденин – адениловый;
гуанин – гуаниловый;
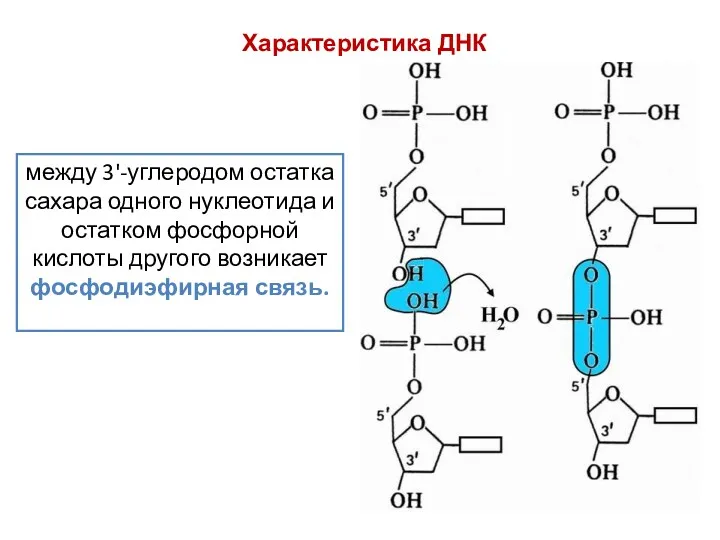
между 3'-углеродом остатка сахара одного нуклеотида и остатком фосфорной кислоты другого
между 3'-углеродом остатка сахара одного нуклеотида и остатком фосфорной кислоты другого
водородные связи возникают между азотистыми основаниями нуклеотидов, располагающихся друг против друга
водородные связи возникают между азотистыми основаниями нуклеотидов, располагающихся друг против друга
Цепи ДНК антипараллельны (разнонаправлены), то есть против 3'-конца одной цепи находится
Цепи ДНК антипараллельны (разнонаправлены), то есть против 3'-конца одной цепи находится
А-форма В-форма Z-форма
Разновидности ДНК
А-форма В-форма Z-форма
Разновидности ДНК
ДНК –имеет сходное строение у всех организмов
Видовые различия выражаются в количестве
ДНК –имеет сходное строение у всех организмов
Видовые различия выражаются в количестве

ГЕНЕТИЧЕСКИЙ код и его свойства
Особая система записи минокисло белка в
ГЕНЕТИЧЕСКИЙ код и его свойства
Особая система записи минокисло белка в
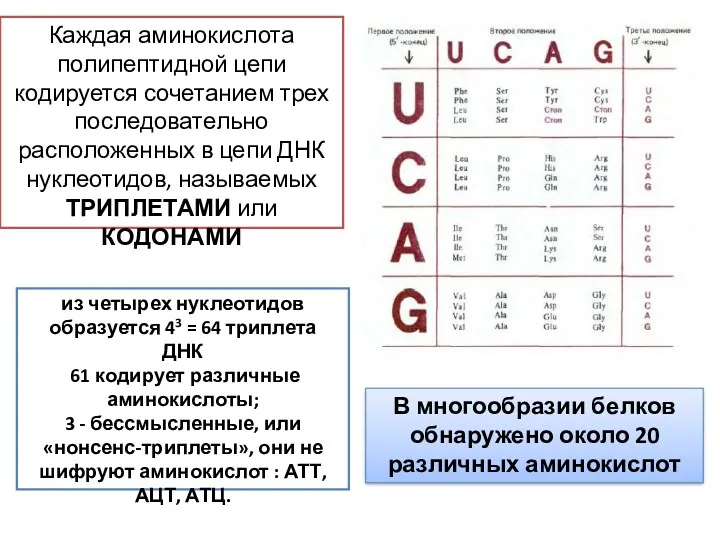
Каждая аминокислота полипептидной цепи кодируется сочетанием трех последовательно расположенных в цепи
Каждая аминокислота полипептидной цепи кодируется сочетанием трех последовательно расположенных в цепи

Свойства генетического кода
Специфичность -
Каждый триплет способен кодировать только одну определенную
Свойства генетического кода
Специфичность -
Каждый триплет способен кодировать только одну определенную

Классификация генов
1. Структурные гены (3-5% ДНК):
Уникальные (их продукты белки: ферменты, транспортные,
Классификация генов
1. Структурные гены (3-5% ДНК):
Уникальные (их продукты белки: ферменты, транспортные,

Свойства генов:
1. Лабильность – способность генов мутировать – изменять нуклеотидную последовательность.
Свойства генов:
1. Лабильность – способность генов мутировать – изменять нуклеотидную последовательность.

Гены человека, встречающиеся в человеческих популяциях в разных молекулярных формах
1. Гены
Гены человека, встречающиеся в человеческих популяциях в разных молекулярных формах
1. Гены

Свойства генов:
4. Аллельное состояние – характерно для диплоидных организмов.
5. Специфичность –
Свойства генов:
4. Аллельное состояние – характерно для диплоидных организмов.
5. Специфичность –
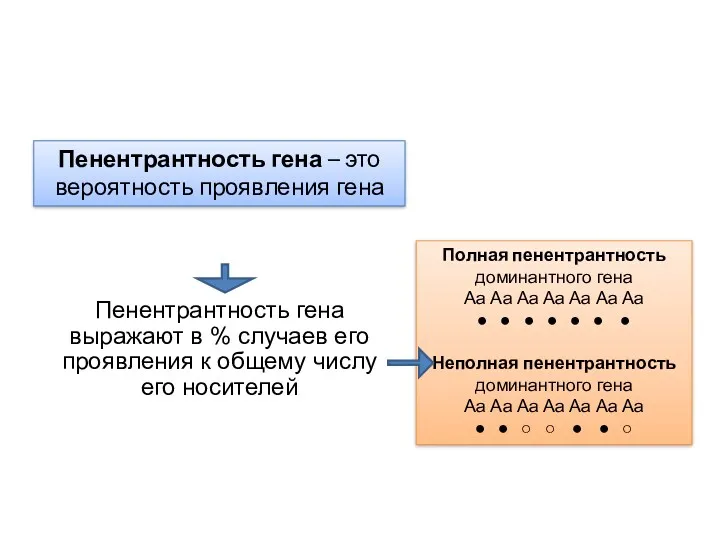
Пенентрантность гена – это вероятность проявления гена
Пенентрантность гена выражают в %
Пенентрантность гена – это вероятность проявления гена
Пенентрантность гена выражают в %

Экспрессивность – степень выраженности гена
(ген проявляется у всех гетерозигот, но
Экспрессивность – степень выраженности гена (ген проявляется у всех гетерозигот, но

Строение генов про- и эукариот
Строение генов про- и эукариот
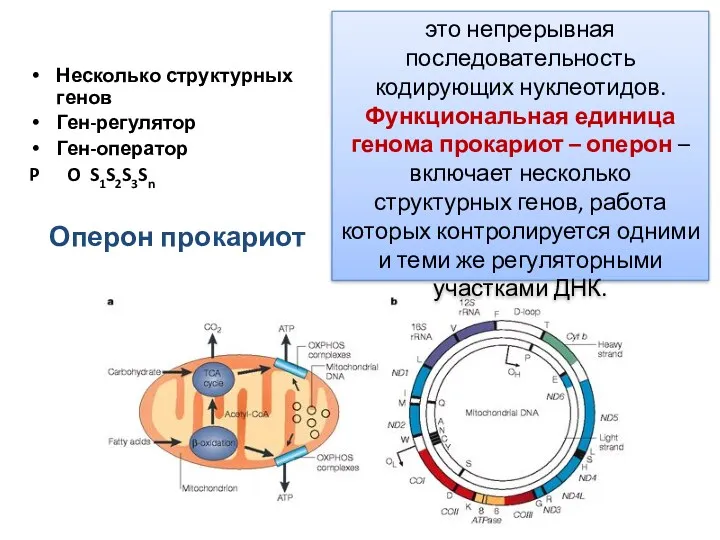
Оперон прокариот
Несколько структурных генов
Ген-регулятор
Ген-оператор
P O S1S2S3Sn
это непрерывная последовательность кодирующих нуклеотидов.
Функциональная
Оперон прокариот
Несколько структурных генов
Ген-регулятор
Ген-оператор
P O S1S2S3Sn
это непрерывная последовательность кодирующих нуклеотидов.
Функциональная

Структура гена прокариот (ДНК-овых)
Непрерывная последовательность кодирующих нуклеотидов
ДНК АЦЦ-ГАТ-ТАТ-ЦЦА-АЦЦ…АТТ…
ТРАНСКРИПЦИЯ
иРНК УГГ-ЦУА-АУА-ГГУ-УГГ…УАА…
ТРАНСЛЯЦИЯ
ТРИ-ЛЕЙ-ИЛЕ-ГЛИ-ТРИ
Порядок
Структура гена прокариот (ДНК-овых)
Непрерывная последовательность кодирующих нуклеотидов
ДНК АЦЦ-ГАТ-ТАТ-ЦЦА-АЦЦ…АТТ…
ТРАНСКРИПЦИЯ
иРНК УГГ-ЦУА-АУА-ГГУ-УГГ…УАА…
ТРАНСЛЯЦИЯ
ТРИ-ЛЕЙ-ИЛЕ-ГЛИ-ТРИ
Порядок

Гены эукариот
имеют мозаичную структуру: состоят из кодирующих (экзонов) и некодирующих
Гены эукариот
имеют мозаичную структуру: состоят из кодирующих (экзонов) и некодирующих

Уникальные нуклеотидные последовательности
представлены единичными копиями на геном, в них локализованы
Уникальные нуклеотидные последовательности
представлены единичными копиями на геном, в них локализованы
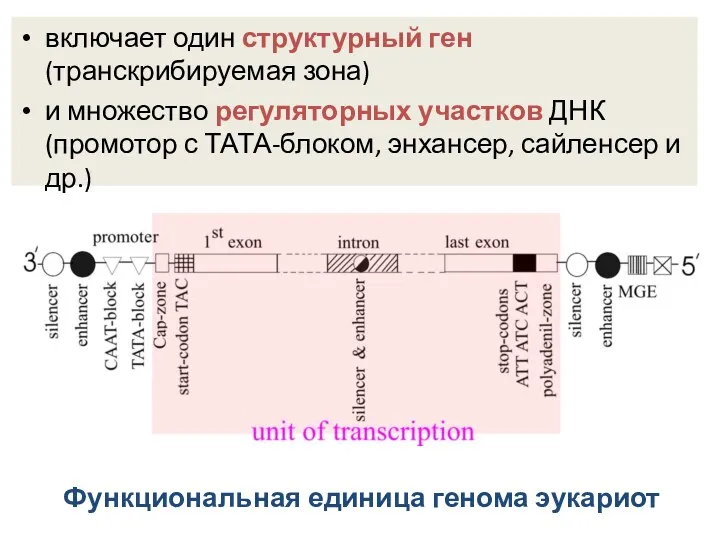
Функциональная единица генома эукариот
включает один структурный ген (транскрибируемая зона)
и
Функциональная единица генома эукариот
включает один структурный ген (транскрибируемая зона)
и

Структурная организация гена эукариот:
Экзоны – нуклеотидные последовательности, кодирующие аминокислоты.
Интроны – не
Структурная организация гена эукариот:
Экзоны – нуклеотидные последовательности, кодирующие аминокислоты.
Интроны – не

хромосомный уровень организации наследственного материала
Хромосомы
– это надмолекулярный комплекс ДНК и
хромосомный уровень организации наследственного материала
Хромосомы
– это надмолекулярный комплекс ДНК и
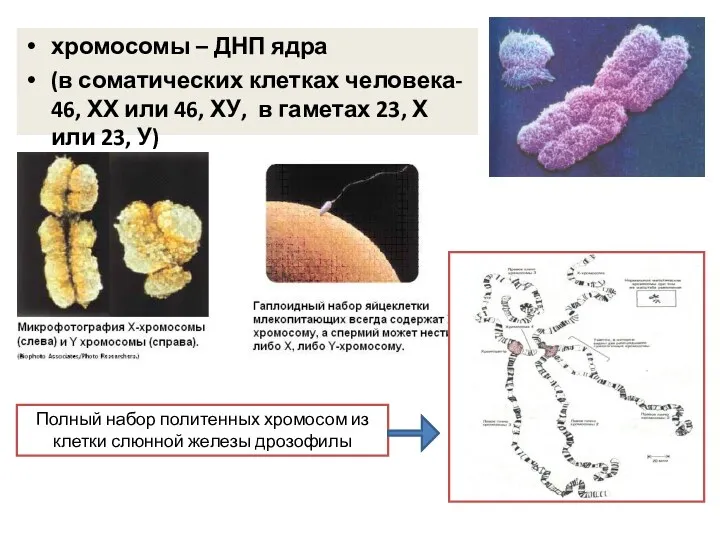
хромосомы – ДНП ядра
(в соматических клетках человека- 46, ХХ или
хромосомы – ДНП ядра
(в соматических клетках человека- 46, ХХ или
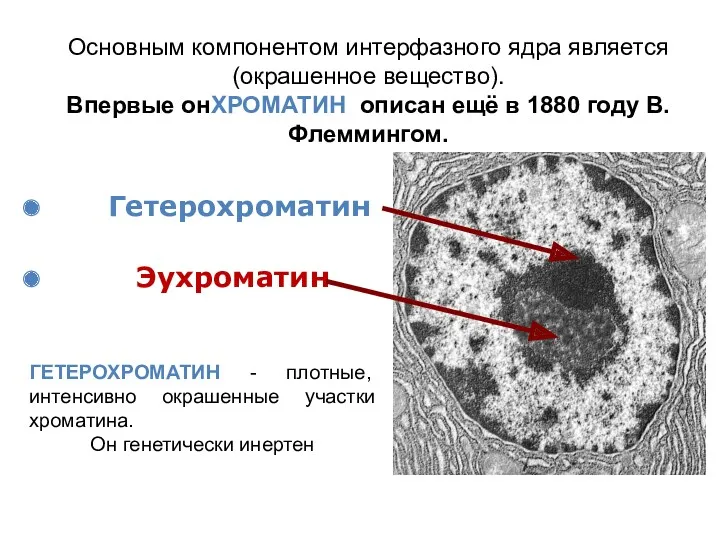
Основным компонентом интерфазного ядра является (окрашенное вещество).
Впервые онХРОМАТИН описан ещё в
Основным компонентом интерфазного ядра является (окрашенное вещество).
Впервые онХРОМАТИН описан ещё в

УРОВНИ КОМПАКТИЗАЦИИ ДНП.
ИНТЕРФАЗНАЯ ХРОМОСОМА
(три уровня: нуклеосомный, фибриллярный и хромонемный)
Интерфазная хромосома
УРОВНИ КОМПАКТИЗАЦИИ ДНП.
ИНТЕРФАЗНАЯ ХРОМОСОМА
(три уровня: нуклеосомный, фибриллярный и хромонемный)
Интерфазная хромосома
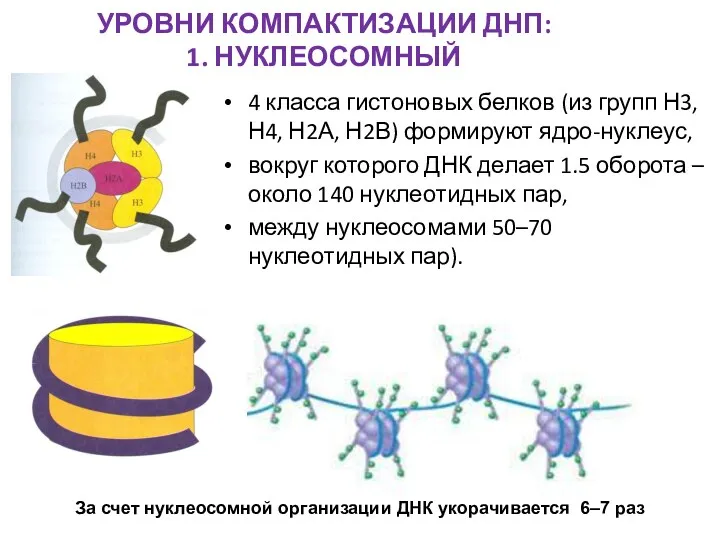
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
1. НУКЛЕОСОМНЫЙ
4 класса гистоновых белков (из групп Н3, Н4,
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
1. НУКЛЕОСОМНЫЙ
4 класса гистоновых белков (из групп Н3, Н4,
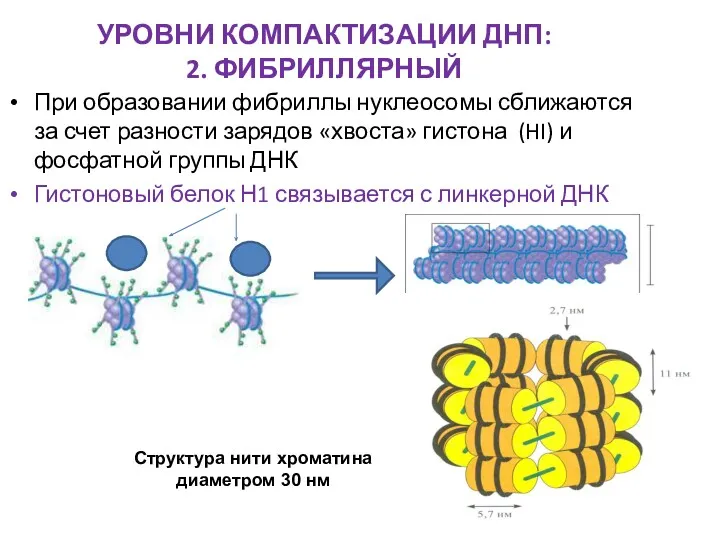
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
2. ФИБРИЛЛЯРНЫЙ
При образовании фибриллы нуклеосомы сближаются за счет разности
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
2. ФИБРИЛЛЯРНЫЙ
При образовании фибриллы нуклеосомы сближаются за счет разности
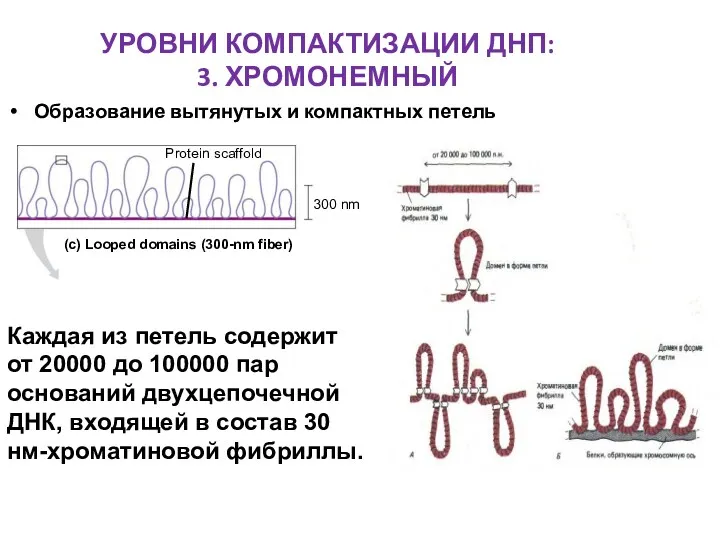
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
3. ХРОМОНЕМНЫЙ
Образование вытянутых и компактных петель
Каждая из петель содержит
от
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
3. ХРОМОНЕМНЫЙ
Образование вытянутых и компактных петель
Каждая из петель содержит от
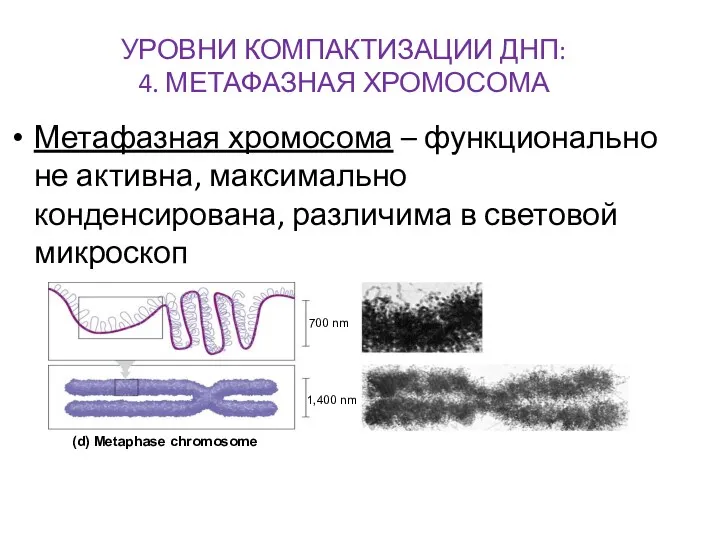
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
4. МЕТАФАЗНАЯ ХРОМОСОМА
Метафазная хромосома – функционально не активна, максимально
УРОВНИ КОМПАКТИЗАЦИИ ДНП:
4. МЕТАФАЗНАЯ ХРОМОСОМА
Метафазная хромосома – функционально не активна, максимально
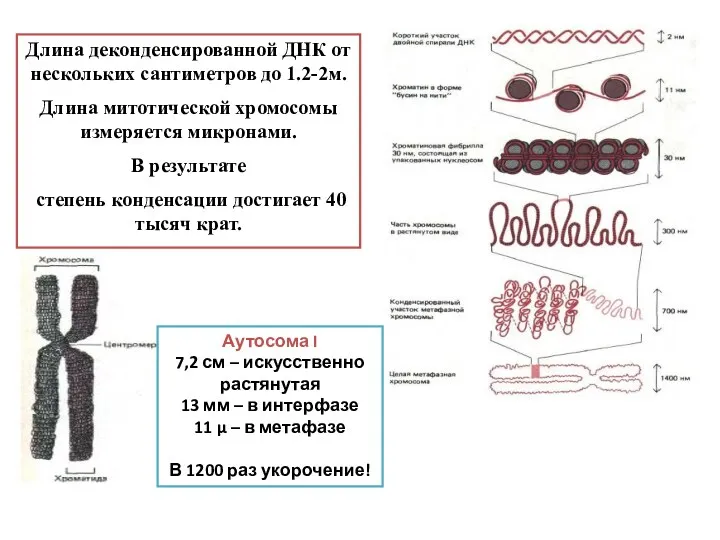
Длина деконденсированной ДНК от нескольких сантиметров до 1.2-2м.
Длина митотической хромосомы
Длина деконденсированной ДНК от нескольких сантиметров до 1.2-2м.
Длина митотической хромосомы
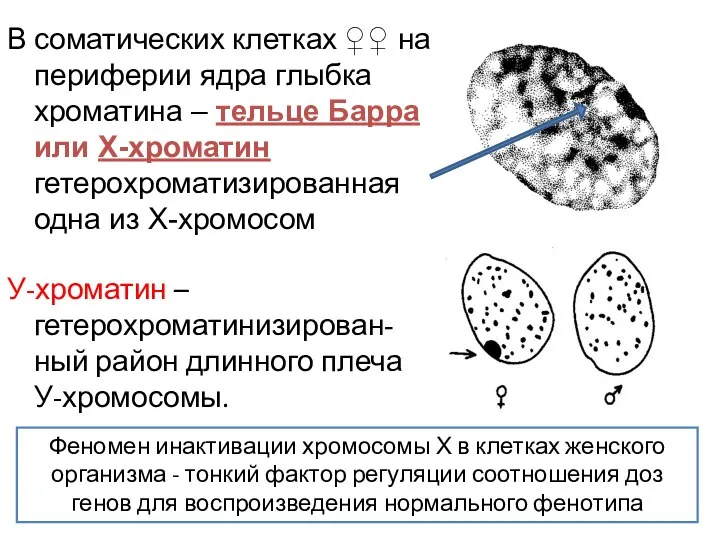
В соматических клетках ♀♀ на периферии ядра глыбка хроматина – тельце
В соматических клетках ♀♀ на периферии ядра глыбка хроматина – тельце

Геномный уровень организации наследственного материала
Вся совокупность наследственного материала, заключенного в гаплоидном
Геномный уровень организации наследственного материала
Вся совокупность наследственного материала, заключенного в гаплоидном
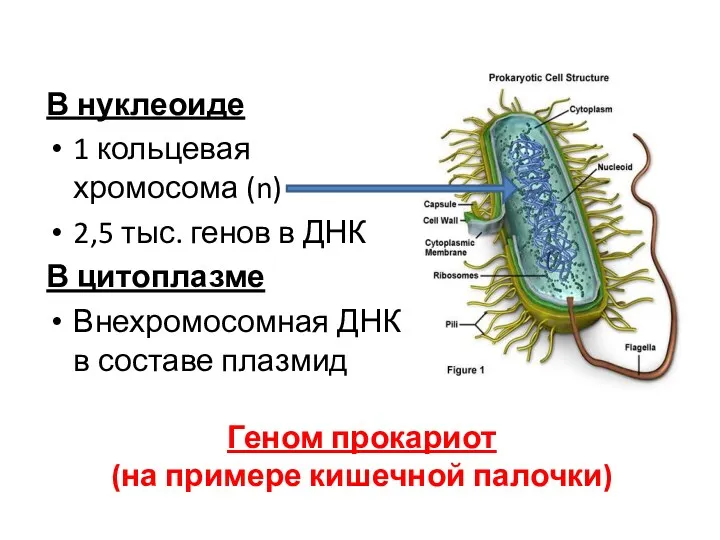
Геном прокариот
(на примере кишечной палочки)
В нуклеоиде
1 кольцевая хромосома (n)
2,5 тыс.
Геном прокариот
(на примере кишечной палочки)
В нуклеоиде
1 кольцевая хромосома (n)
2,5 тыс.

длина ДНК в ядре клетки млекопитающего составляет примерно 2—5·109 пар нуклеотидов,
длина ДНК в ядре клетки млекопитающего составляет примерно 2—5·109 пар нуклеотидов,

Наследственный аппарат клеток человека состоит из внеядерного – митохондриального и ядерного
Наследственный аппарат клеток человека состоит из внеядерного – митохондриального и ядерного
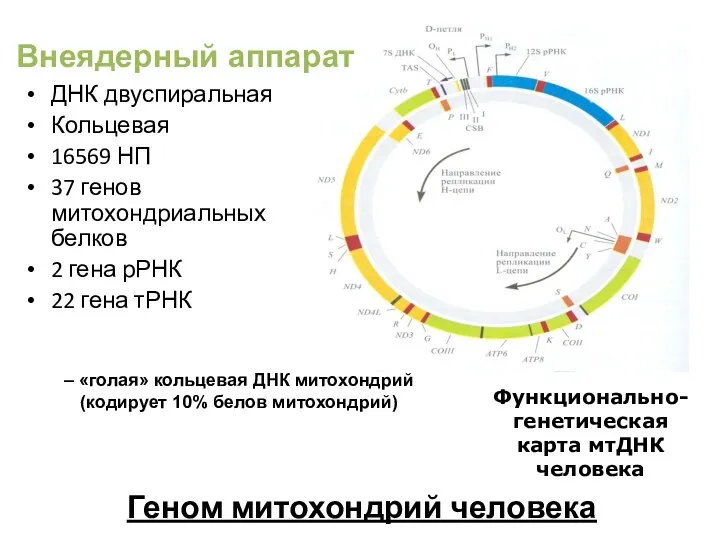
Геном митохондрий человека
ДНК двуспиральная
Кольцевая
16569 НП
37 генов митохондриальных белков
2 гена рРНК
22 гена
Геном митохондрий человека
ДНК двуспиральная
Кольцевая
16569 НП
37 генов митохондриальных белков
2 гена рРНК
22 гена

Внеядерный аппарат
Хромосомы митохондрий распределяются в митозе и мейозе неравномерно,
обеспечивают цитоплазматическую
Внеядерный аппарат
Хромосомы митохондрий распределяются в митозе и мейозе неравномерно,
обеспечивают цитоплазматическую
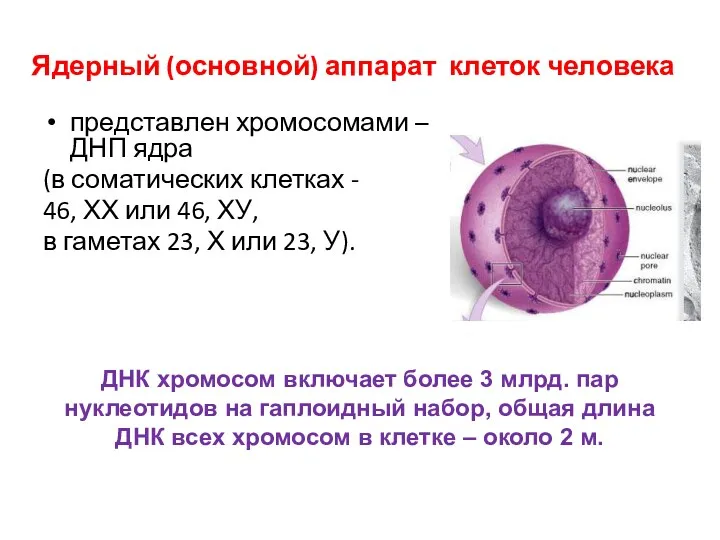
Ядерный (основной) аппарат клеток человека
представлен хромосомами – ДНП ядра
(в соматических
Ядерный (основной) аппарат клеток человека
представлен хромосомами – ДНП ядра
(в соматических

Ядерный (основной) аппарат клеток человека
В митозе и мейозе ядерные хромосомы сегрегируют
Ядерный (основной) аппарат клеток человека
В митозе и мейозе ядерные хромосомы сегрегируют

Кариотип 2n
Шимпанзе – 48
Лошадь – 64
Свинья и кошка – 38
Собака –
Кариотип 2n
Шимпанзе – 48
Лошадь – 64
Свинья и кошка – 38
Собака –
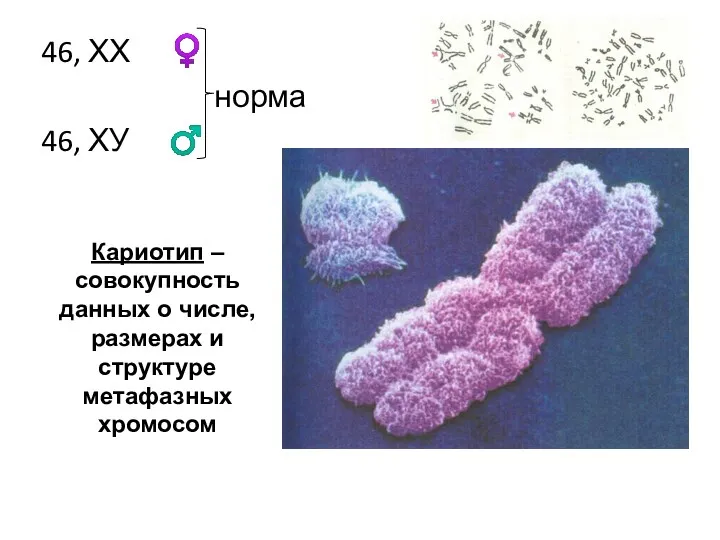
46, ХХ ♀
норма
46, ХУ ♂
Кариотип – совокупность данных о числе,
46, ХХ ♀
норма
46, ХУ ♂
Кариотип – совокупность данных о числе,
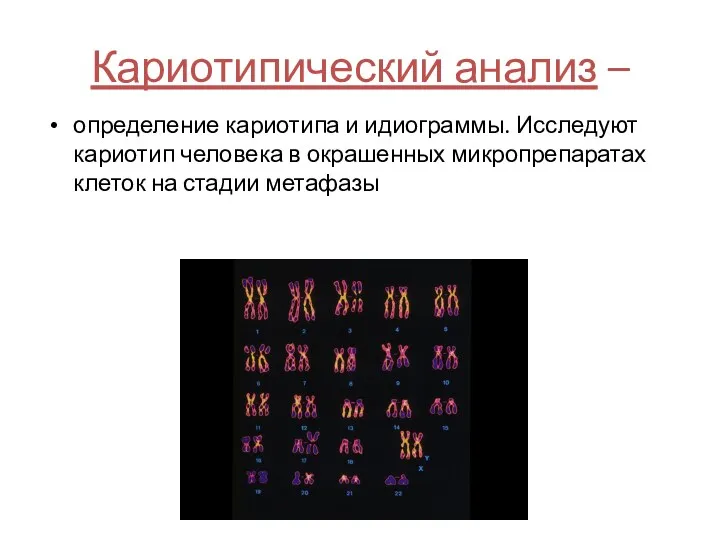
Кариотипический анализ –
определение кариотипа и идиограммы. Исследуют кариотип человека в
Кариотипический анализ –
определение кариотипа и идиограммы. Исследуют кариотип человека в


КАРИОТИПИЧЕСКИЙ АНАЛИЗ
С учетом морфологических параметров аутосомы человека классифицированы на 7 групп
КАРИОТИПИЧЕСКИЙ АНАЛИЗ
С учетом морфологических параметров аутосомы человека классифицированы на 7 групп

Методы дифференциального окрашивания хромосом
позволяют индивидуализировать все хромосомы в кариотипе
Методы
Методы дифференциального окрашивания хромосом
позволяют индивидуализировать все хромосомы в кариотипе
Методы

Световая микрофотография участка политенной хромосомы из клетки слюнной железы дрозофилы.
Видна
Световая микрофотография участка политенной хромосомы из клетки слюнной железы дрозофилы. Видна
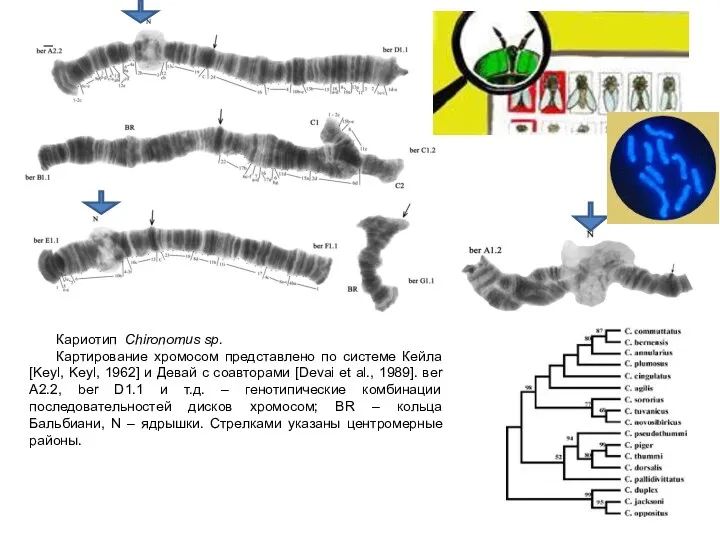
Кариотип Chironomus sp.
Картирование хромосом представлено по системе Кейла [Keyl, Keyl, 1962]
Кариотип Chironomus sp.
Картирование хромосом представлено по системе Кейла [Keyl, Keyl, 1962]

Идиограмма –
графическое изображение хромосом с учетом их абсолютной и относительной
Идиограмма – графическое изображение хромосом с учетом их абсолютной и относительной
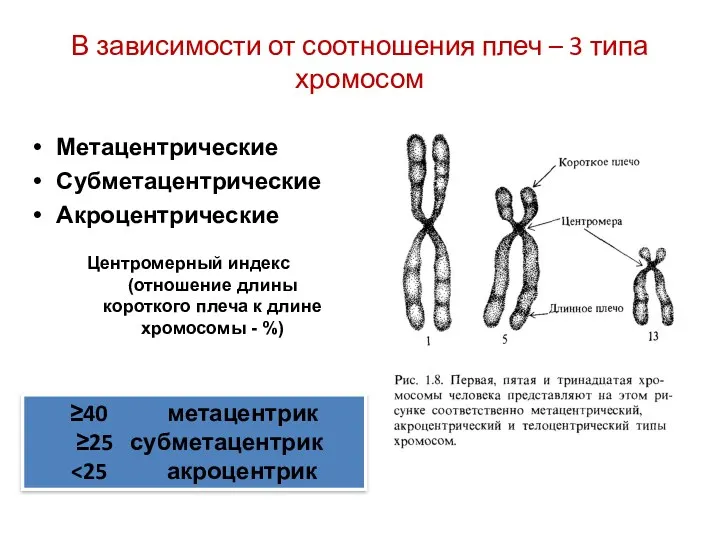
В зависимости от соотношения плеч – 3 типа хромосом
Метацентрические
Субметацентрические
Акроцентрические
Центромерный индекс (отношение
В зависимости от соотношения плеч – 3 типа хромосом
Метацентрические
Субметацентрические
Акроцентрические
Центромерный индекс (отношение
Теломеры на концах митотической хромосомы
Теломеры на концах митотической хромосомы
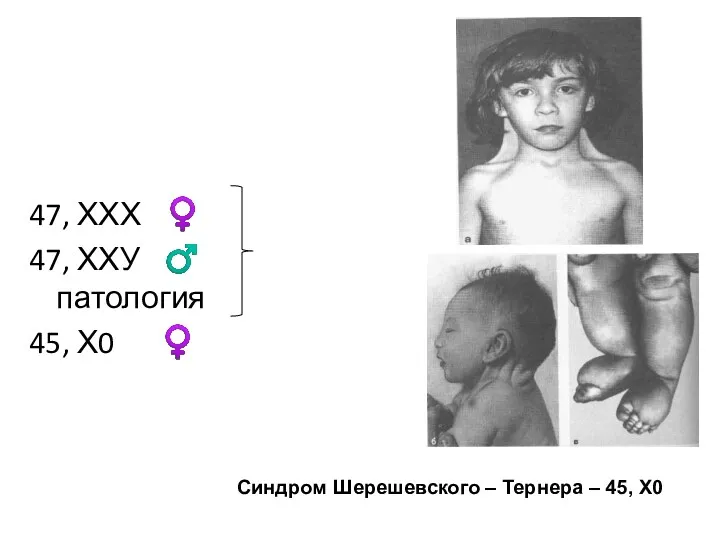
47, ХХХ ♀
47, ХХУ ♂ патология
45, Х0 ♀
Синдром Шерешевского – Тернера
47, ХХХ ♀
47, ХХУ ♂ патология
45, Х0 ♀
Синдром Шерешевского – Тернера
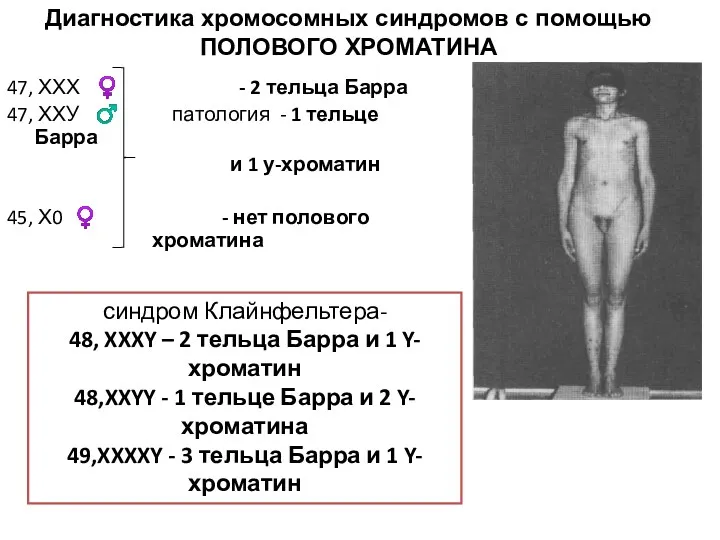
47, ХХХ ♀ - 2 тельца Барра
47, ХХУ ♂ патология -
47, ХХХ ♀ - 2 тельца Барра
47, ХХУ ♂ патология -
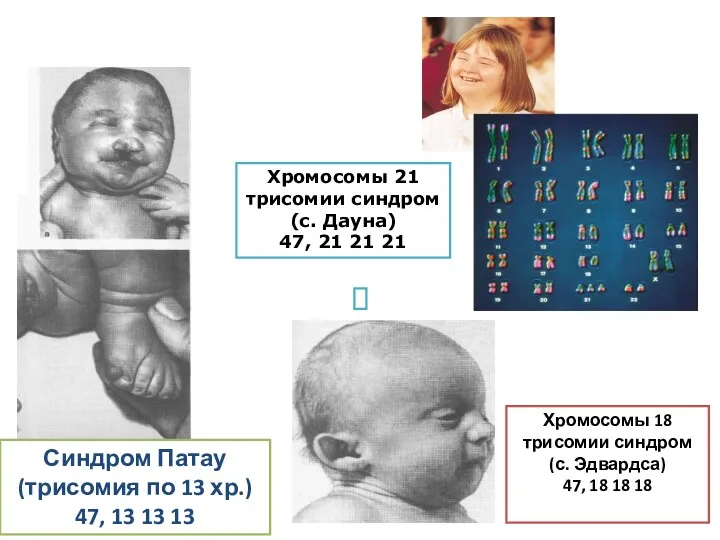
Синдром Патау
(трисомия по 13 хр.)
47, 13 13 13
Хромосомы 18 трисомии синдром(с.
Синдром Патау
(трисомия по 13 хр.)
47, 13 13 13
Хромосомы 18 трисомии синдром(с.

Основные этапы реализации генетической информации
Основные этапы реализации генетической информации

результат взаимодействия наследственности и среды
Формирование признака –
Признак – свойство организма
результат взаимодействия наследственности и среды
Формирование признака –
Признак – свойство организма

ПРИЗНАК
(на молекулярном уровне)
ПРИЗНАК
(на молекулярном уровне)

ЭКСПРЕССИЯ ГЕНОВ
От лат. - EXPRESSIO – выражение, сила проявления.
Процесс реализации генетической
ЭКСПРЕССИЯ ГЕНОВ
От лат. - EXPRESSIO – выражение, сила проявления.
Процесс реализации генетической
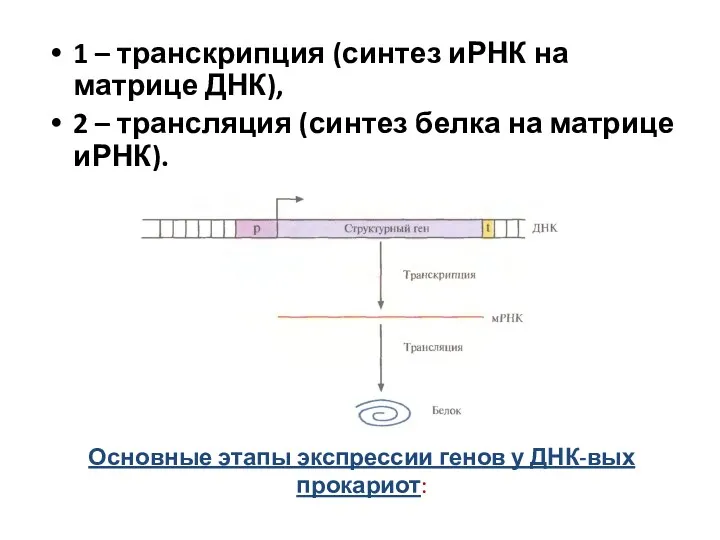
Основные этапы экспрессии генов у ДНК-вых прокариот:
1 – транскрипция (синтез
Основные этапы экспрессии генов у ДНК-вых прокариот:
1 – транскрипция (синтез
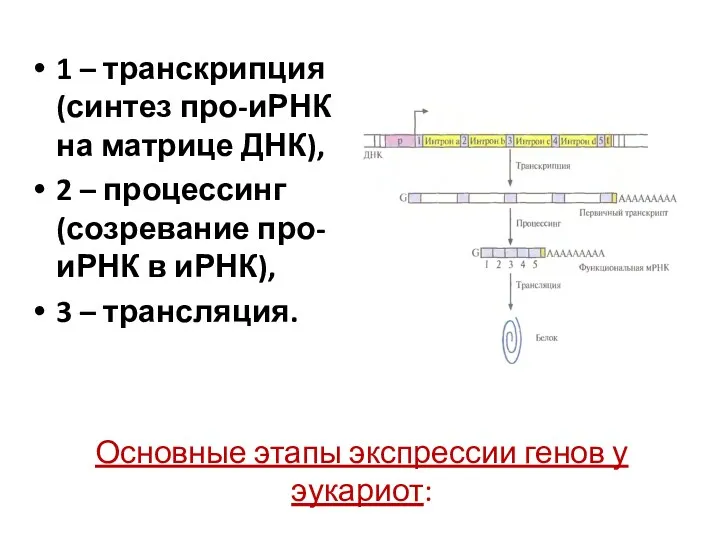
Основные этапы экспрессии генов у эукариот:
1 – транскрипция (синтез про-иРНК
Основные этапы экспрессии генов у эукариот:
1 – транскрипция (синтез про-иРНК
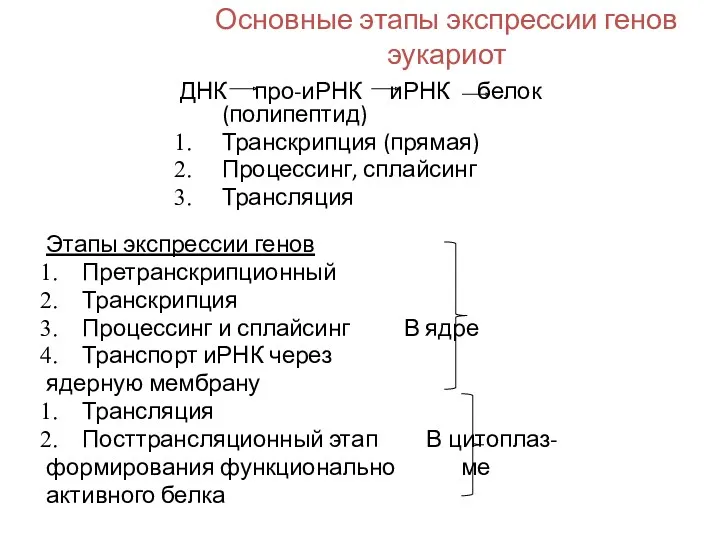
Основные этапы экспрессии генов эукариот
ДНК про-иРНК иРНК белок (полипептид)
Транскрипция (прямая)
Процессинг, сплайсинг
Трансляция
Этапы
Основные этапы экспрессии генов эукариот
ДНК про-иРНК иРНК белок (полипептид)
Транскрипция (прямая)
Процессинг, сплайсинг
Трансляция
Этапы
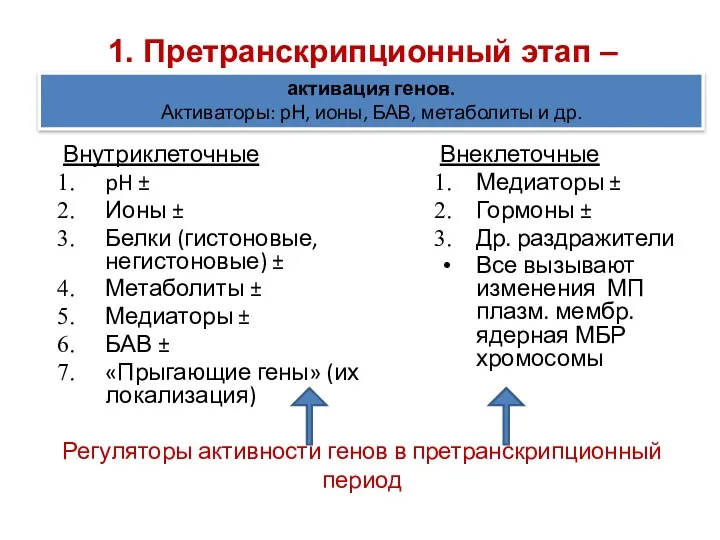
Регуляторы активности генов в претранскрипционный период
Внутриклеточные
pH ±
Ионы ±
Белки (гистоновые, негистоновые)
Регуляторы активности генов в претранскрипционный период
Внутриклеточные
pH ±
Ионы ±
Белки (гистоновые, негистоновые)

Функции регуляторов генной активности
(в зоне действия)
Освобождают ДНК от белков
Деконденсация ДНП
Ослабляют
Функции регуляторов генной активности
(в зоне действия)
Освобождают ДНК от белков
Деконденсация ДНП
Ослабляют
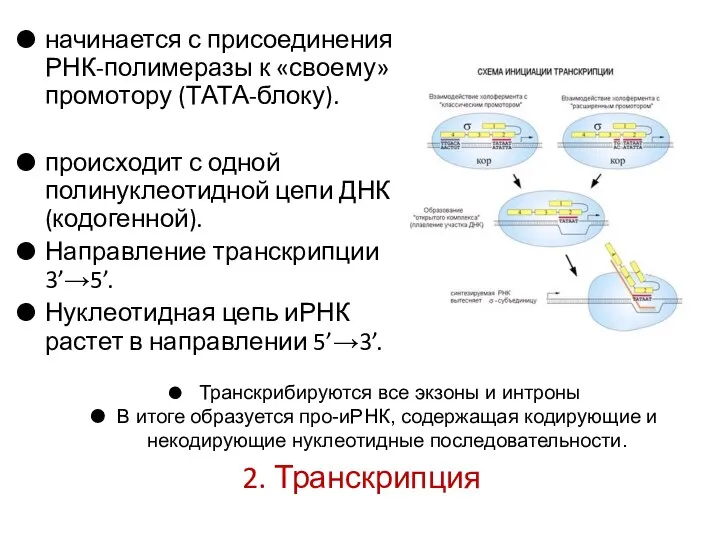
2. Транскрипция
начинается с присоединения РНК-полимеразы к «своему» промотору (ТАТА-блоку).
происходит
2. Транскрипция
начинается с присоединения РНК-полимеразы к «своему» промотору (ТАТА-блоку).
происходит
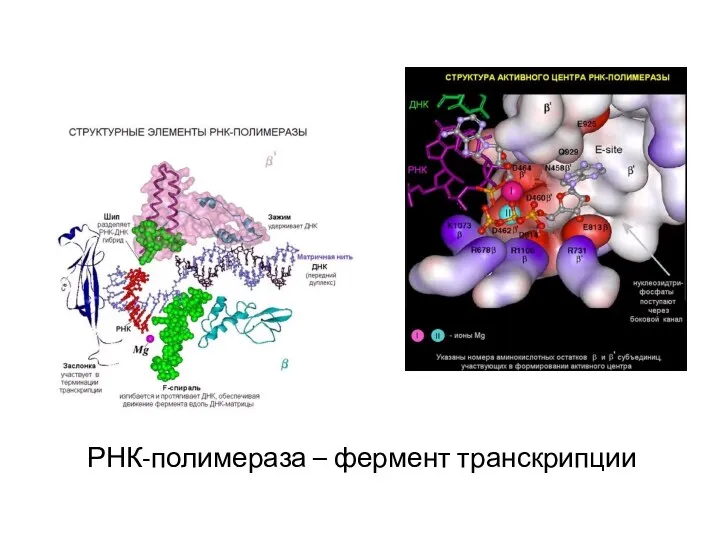
РНК-полимераза – фермент транскрипции
РНК-полимераза – фермент транскрипции

Прямая транскрипция – синтез РНК на матрице ДНК
ДНК РНК Белок (1958
Прямая транскрипция – синтез РНК на матрице ДНК
ДНК РНК Белок (1958

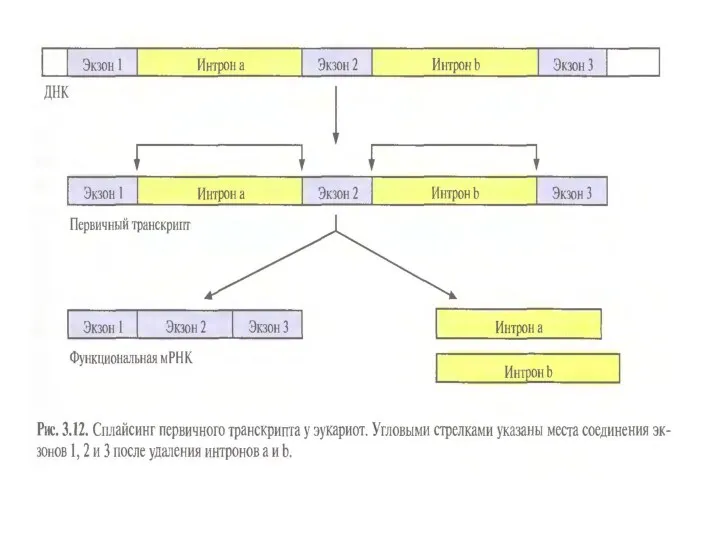
3. Процессинг про-иРНК:
1) Кэпирование – образование КЭП на 5’-конце про-иРНК.
3. Процессинг про-иРНК:
1) Кэпирование – образование КЭП на 5’-конце про-иРНК.

Аномальный процессинг болезнь Альцгеймера
Аномальный процессинг болезнь Альцгеймера

4. Этап экспрессии генов
Транспорт зрелой иРНК через ядерную мембрану
4. Этап экспрессии генов
Транспорт зрелой иРНК через ядерную мембрану
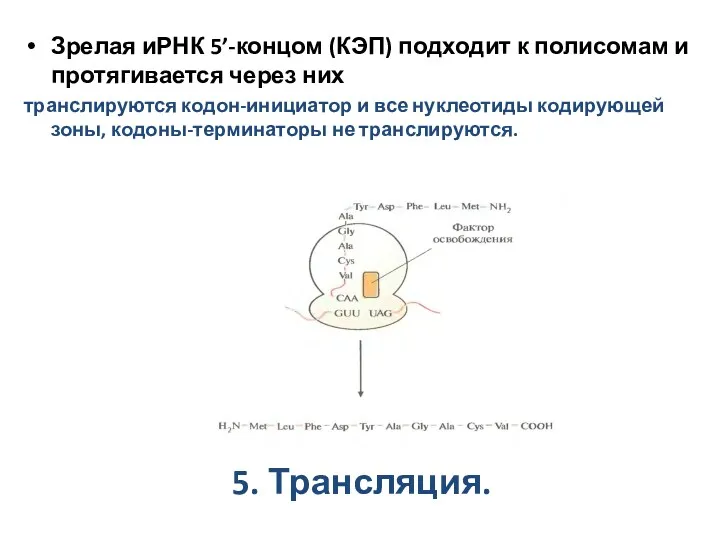
5. Трансляция.
Зрелая иРНК 5’-концом (КЭП) подходит к полисомам и протягивается
5. Трансляция.
Зрелая иРНК 5’-концом (КЭП) подходит к полисомам и протягивается

Компоненты, необходимые для трансляции
Зрелая иРНК
Рибосомы (полисомы)
Набор тРНК (~60)
Набор аминокислот
Набор ферментов
Компоненты, необходимые для трансляции
Зрелая иРНК
Рибосомы (полисомы)
Набор тРНК (~60)
Набор аминокислот
Набор ферментов
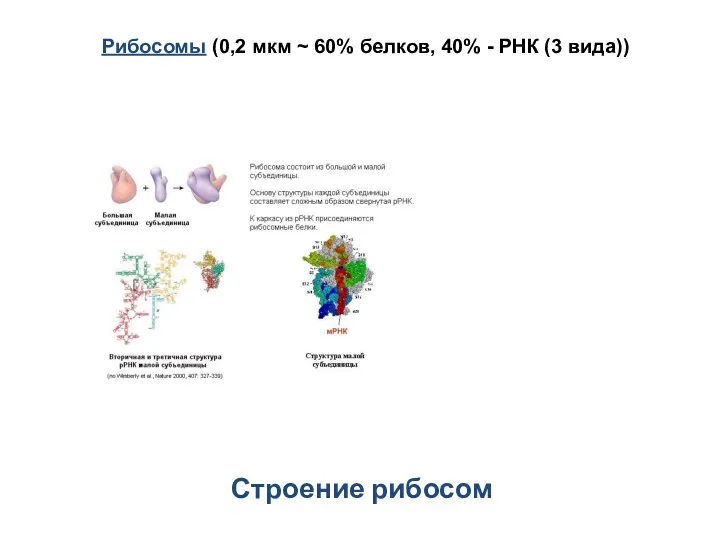
Строение рибосом
Рибосомы (0,2 мкм ~ 60% белков, 40% - РНК (3
Строение рибосом
Рибосомы (0,2 мкм ~ 60% белков, 40% - РНК (3
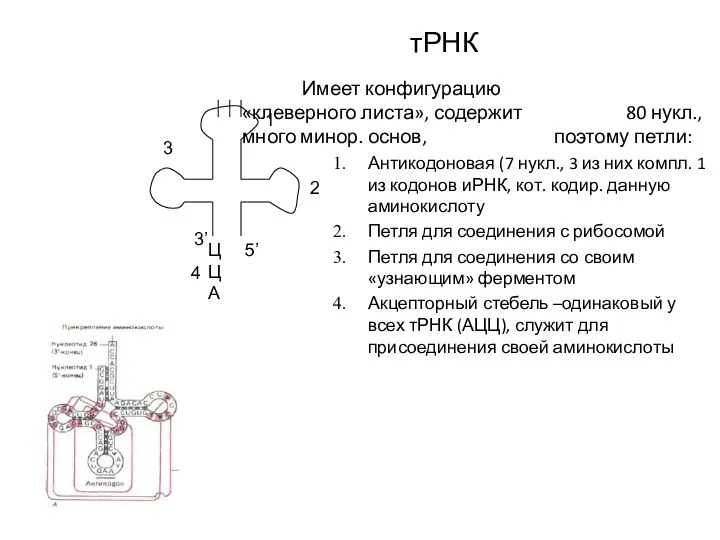
тРНК
Имеет конфигурацию «клеверного листа», содержит 80 нукл., много минор. основ, поэтому
тРНК
Имеет конфигурацию «клеверного листа», содержит 80 нукл., много минор. основ, поэтому
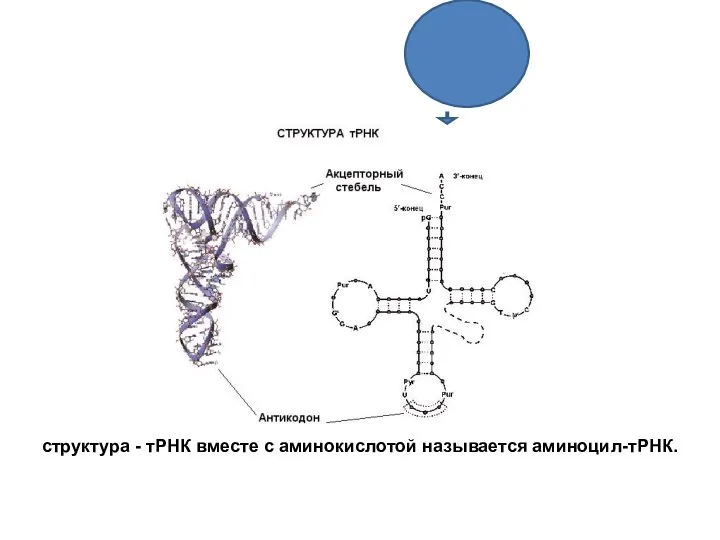
структура - тРНК вместе с аминокислотой называется аминоцил-тРНК.
структура - тРНК вместе с аминокислотой называется аминоцил-тРНК.
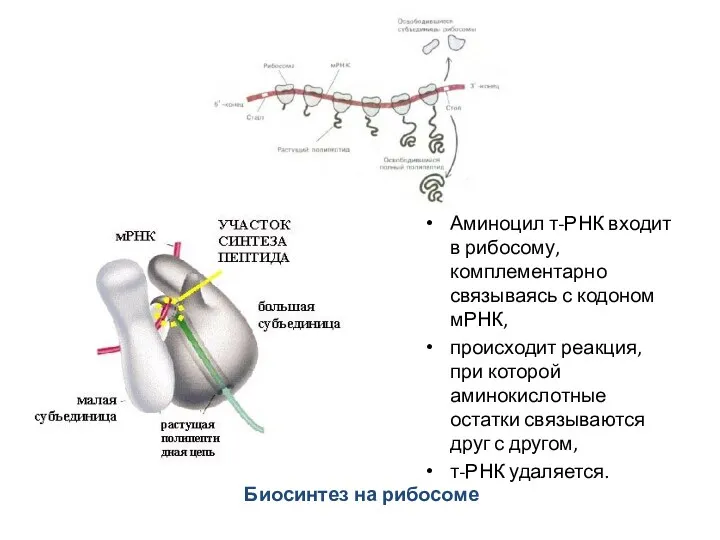
Биосинтез на рибосоме
Аминоцил т-РНК входит в рибосому, комплементарно связываясь с кодоном
Биосинтез на рибосоме
Аминоцил т-РНК входит в рибосому, комплементарно связываясь с кодоном
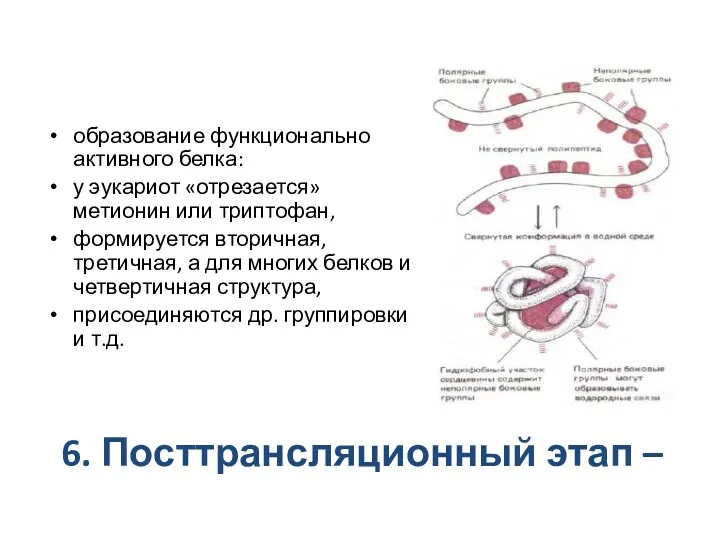
6. Посттрансляционный этап –
образование функционально активного белка:
у эукариот «отрезается»
6. Посттрансляционный этап –
образование функционально активного белка:
у эукариот «отрезается»
 Кәдімгі цикорий-Сichorium intybus l
Кәдімгі цикорий-Сichorium intybus l семейство розоцветные
семейство розоцветные Классификация животных и основные систематические группы
Классификация животных и основные систематические группы Көбею. Өсу және даму
Көбею. Өсу және даму Презентация по биологии для учащихся 8 класса на тему: Обмен веществ
Презентация по биологии для учащихся 8 класса на тему: Обмен веществ Кабинет биологии, фото
Кабинет биологии, фото Вегетативное размножение растений
Вегетативное размножение растений Екіншілік метаболиттер алу биотехнологиясы
Екіншілік метаболиттер алу биотехнологиясы Природний добір
Природний добір Мышцы конечностей
Мышцы конечностей Отдел Плауновидные Lycopodiophyta
Отдел Плауновидные Lycopodiophyta Технология возделывания яровой пшеницы
Технология возделывания яровой пшеницы Занятие элективного курса Основы биологии для 11профильного класса.
Занятие элективного курса Основы биологии для 11профильного класса. Животные степей
Животные степей Субтропические плоды
Субтропические плоды Экспрессия генов. Транскрипция, трансляция
Экспрессия генов. Транскрипция, трансляция Вода - источник жизни комнатных растений
Вода - источник жизни комнатных растений Анатомия и физиология женских половых органов
Анатомия и физиология женских половых органов Земледелие. Введение
Земледелие. Введение Пищеварение в кишечнике
Пищеварение в кишечнике Молодежный инновационный форум Воронежской области
Молодежный инновационный форум Воронежской области Вселенная микробов
Вселенная микробов Николай Иванович Вавилов
Николай Иванович Вавилов Биологическое и свободное окисление
Биологическое и свободное окисление Байгабилов Д., 2 курс
Байгабилов Д., 2 курс Природоохранные территории России. Баргузинский государственный биосферный заповедник
Природоохранные территории России. Баргузинский государственный биосферный заповедник Преддверно-улитковый нерв п. vestibulocochlearis (VIII пара)
Преддверно-улитковый нерв п. vestibulocochlearis (VIII пара) Ткани растений и их виды
Ткани растений и их виды