Структурно-функциональные уровни организации наследственного материала. Молекулярные основы наследственности презентация
- Структурно-функциональные уровни организации наследственного материала. Молекулярные основы наследственности

Содержание
- 2. Термин «биология» впервые был предложен французским ученым Ж.Б. Ламарком в 1802 году. Этот термин состоит из
- 3. Фундаментальными свойствами жизни являются способность к самообновлению, самовоспроизведению, саморегуляции. На этих трех свойствах основаны все проявления
- 4. - раздражимость – способность давать ответную реакцию на действие факторов внешней среды; - размножение; - гомеостаз;
- 5. Уровни организации жизни Живая природа – целостная, но неоднородная система, которой свойственна иерархическая организация, элементы которой
- 6. Выделяют 6 уровней организации жизни. 1. Молекулярно – генетический уровень Элементарными структурами на данном уровне живого
- 7. 2. Клеточный уровень Элементарной структурой является клетка, а элементарным явлением – реакции клеточного обмена веществ. 3.
- 8. Элементарное специфическое явление: процесс реализации наследственной информации, закодированной в молекуле ДНК. Процесс реализации генотипа в фенотип.
- 9. 5. Биогеоценотический уровень Элементарной единицей является биогеоценоз – сообщество животных и растительных организмов, которые взаимодействуют с
- 10. Молекулярно-генетический уровень организации живых систем В наследственной структуре клетки и организма в целом выделяют три уровня
- 11. Генный уровень Наименьшей (элементарной) единицей наследственного материала является ген. Ген – это часть молекулы ДНК, имеющая
- 12. Хромосомный уровень Гены располагаются в хромосомах в линейном порядке. Каждая хромосома уникальна по набору входящих в
- 13. Геномный уровень Геном – совокупность всех генов, находящихся в гаплоидном наборе хромосом. * При оплодотворении два
- 14. Геномный уровень отличается высокой стабильностью. Он обеспечивает сложную систему взаимодействия генов. Результатом взаимодействия генов друг с
- 15. Ген как элементарная единица наследственной информации выполняет определенные функции и обладает определенными свойствами. Функции генов: хранение
- 16. Свойства генов: дискретность: один ген контролирует один признак; специфичность: каждый ген отвечает строго за определенный признак;
- 17. дозированность действия: один ген определяет одну дозу фенотипического проявления признака; способность к мутированию (изменению структуры); способность
- 18. Функциональная классификация генов Выделяют три группы генов: cтруктурные – контролируют развитие признаков путем синтеза соответствующих ферментов;
- 19. Гены в клетках прокариот и эукариот имеют определенные особенности строения. Принципиальное отличие: у прокариот ген имеет
- 20. У эукариот – прерывистую, он состоит из чередующихся участков – информативных (экзонов) и неинформативных участков, или
- 21. Экспрессия генов в биосинтезе белка В процессе синтеза белка условно выделяют три этапа: транскрипция; процессинг; трансляция.
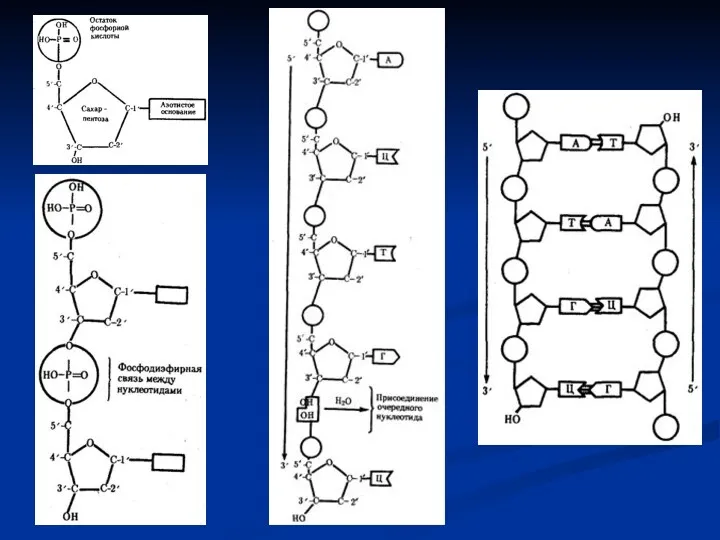
- 22. Биологической сущностью транскрипции является «переписывание» информации с молекулы ДНК на РНК, а химической – синтез молекулы
- 23. Синтез и-РНК в клетке всегда осуществляется от фосфатного конца к гидроксильному (5'→3‘). Поэтому матрицей для транскрипции
- 25. Транскрипция делится на три периода: инициация, элонгация, терминация.
- 26. Инициация транскрипции Синтез и-РНК осуществляется при помощи фермента РНК-полимеразы. *У прокариот имеется только одна РНК-полимераза, в
- 27. Фермент РНК-полимераза отыскивает в молекуле ДНК участок – промотор – и с помощью ряда белков –
- 28. Элонгация При участии фермента РНК-полимеразы происходит синтез молекулы РНК из свободных рибонуклеотидов, присоединившихся к матричной цепи
- 29. За 1 секунду между собой соединяются 50 нуклеотидов. Для клетки этой скорости недостаточно, поэтому инициация и
- 30. Терминация Происходит тогда, когда РНК-полимераза достигает терминатора – участка ДНК, где прекращается синтез молекулы и-РНК. В
- 31. Они называются палиндромами, тогда в и-РНК может получиться образование типа шпильки, не позволяющее РНК-полимеразе двигаться дальше.
- 32. Процессинг Процессинг включает целый ряд преобразований и-РНК, необходимых для ее нормального функционирования. 1. Образование колпачка, или
- 33. 3. Удаление части нуклеотидов на гидроксильном конце. 4. Присоединение на гидроксильном конце образования poli-А из 100-200
- 34. 5. Сплайсинг – процесс удаления интронов и соединения экзонов в молекулу РНК. Ядерная и-РНК называется юной,
- 35. 1 – лидирующий участок; АУГ - стартовый кодон; 2 – экзоны (их может быть много); 3
- 36. Трансляция Трансляция – это процесс синтеза полипептидных цепей, осуществляемый в рибосомах. В ходе трансляции происходит считывание
- 37. Инициация трансляции и-РНК своим кэпированным (фосфатным) концом отыскивает малую субъединицу рибосомы. Лидирующая последовательность соединяется с рибосомальной
- 38. К стартовому кодону присоединяется т-РНК, несущая аминокислоту метионин. Только после этого субъединицы рибосомы объединяются. Инициация заканчивается.
- 39. Элонгация Заключается в синтезе полипептида из аминокислот, которые доставляются в рибосому транспортными РНК. Сначала в аминоацильном
- 40. Затем аминокислоты, находящиеся в аминоацильном и пептидильном центрах соединяются при помощи специального фермента пептидной связью. *
- 41. В результате вторая т-РНК и соединенный с ней дипептид оказываются в петидильном центре, а в аминоацильный
- 42. Терминация Терминация наступает тогда, когда в аминоацильный центр поступает один из трех кодонов-терминаторов – УАА, УАГ,
- 43. Регуляция экспрессии генов Регуляция генной активности в клетках может происходить на всех этапах экспрессии – от
- 44. Кишечная палочка при попадании в среду, содержащую молочный сахар лактозу, вырабатывает фермент лактазу. Если же лактозы
- 45. Схема lac-оперона. Р – ген-регулятор; П – промотор; О – ген-оператор; Z, Y, A – структурные
- 46. Ген-регулятор кодирует синтез белка-репрессора, который в химическом отношении очень активен, в свободном состоянии не существует и
- 47. Когда в среде появляется лактоза, то репрессор связывается с ней, освобождая ген-оператор. РНК-полимераза присоединяется к промотору
- 48. Этот принцип регуляции называется принципом индукции. Индуктором в данном случае является молочный сахар – лактоза, ее
- 49. Возможен и другой принцип регуляции синтеза белка – принцип репрессии. В этом случае появление продуктов реакции
- 50. Принцип репрессии Исходно белок-репрессор находится в неактивной форме, поэтому он ни с чем не вступает в
- 51. Активация репрессора происходит только тогда, когда продуктов реакции накопится определенное количество (достаточно большое!). Нахождение репрессора в
- 52. Особенности регуляции генной активности у эукариот У эукариот принцип оперонной регуляции не обнаружен. Активность каждого гена
- 53. Один из таких участков находится перед промотором и называется препромоторным элементом; другие области лежат вдали от
- 54. Разновидности генов Наряду с функциональной классификацией генов существуют и другие их разновидности: псевдогены, онкогены и мобильные
- 55. Мобильные (прыгающие) гены – гены, не имеющие постоянной локализации не только в хромосоме, но и в
- 56. В последние десятилетия в генетике появилось еще одно новое понятие – «семейство генов», или «мультигенное семейство».
- 57. Организация генов мультигенных семейств может быть разной. Семейства актиновых и миозиновых генов разбросаны по всему геному.
- 59. Скачать презентацию

Термин «биология» впервые был предложен французским ученым Ж.Б. Ламарком в 1802
Термин «биология» впервые был предложен французским ученым Ж.Б. Ламарком в 1802

Фундаментальными свойствами жизни являются способность к самообновлению, самовоспроизведению, саморегуляции.
На этих трех
Фундаментальными свойствами жизни являются способность к самообновлению, самовоспроизведению, саморегуляции.
На этих трех

- раздражимость – способность давать ответную реакцию на действие факторов внешней
- раздражимость – способность давать ответную реакцию на действие факторов внешней

Уровни организации жизни
Живая природа – целостная, но неоднородная система, которой
Уровни организации жизни
Живая природа – целостная, но неоднородная система, которой

Выделяют 6 уровней организации жизни.
1. Молекулярно – генетический уровень
Элементарными структурами
Выделяют 6 уровней организации жизни.
1. Молекулярно – генетический уровень
Элементарными структурами

2. Клеточный уровень
Элементарной структурой является клетка, а элементарным явлением – реакции
2. Клеточный уровень
Элементарной структурой является клетка, а элементарным явлением – реакции

Элементарное специфическое явление: процесс реализации наследственной информации, закодированной в молекуле ДНК.
Элементарное специфическое явление: процесс реализации наследственной информации, закодированной в молекуле ДНК.

5. Биогеоценотический уровень
Элементарной единицей является биогеоценоз – сообщество животных и растительных
5. Биогеоценотический уровень
Элементарной единицей является биогеоценоз – сообщество животных и растительных

Молекулярно-генетический уровень организации живых систем
В наследственной структуре клетки и организма
Молекулярно-генетический уровень организации живых систем
В наследственной структуре клетки и организма

Генный уровень
Наименьшей (элементарной) единицей наследственного материала является ген.
Ген – это часть
Генный уровень
Наименьшей (элементарной) единицей наследственного материала является ген.
Ген – это часть

Хромосомный уровень
Гены располагаются в хромосомах в линейном порядке. Каждая хромосома уникальна
Хромосомный уровень
Гены располагаются в хромосомах в линейном порядке. Каждая хромосома уникальна

Геномный уровень
Геном – совокупность всех генов, находящихся в гаплоидном наборе хромосом.
Геномный уровень
Геном – совокупность всех генов, находящихся в гаплоидном наборе хромосом.

Геномный уровень отличается высокой стабильностью. Он обеспечивает сложную систему взаимодействия генов.

Ген как элементарная единица наследственной информации выполняет определенные функции и обладает

Свойства генов:
дискретность: один ген контролирует один признак;
специфичность: каждый ген
Свойства генов:
дискретность: один ген контролирует один признак;
специфичность: каждый ген

дозированность действия: один ген определяет одну дозу фенотипического проявления признака;
дозированность действия: один ген определяет одну дозу фенотипического проявления признака;

Функциональная классификация генов
Выделяют три группы генов:
cтруктурные – контролируют развитие признаков
Функциональная классификация генов
Выделяют три группы генов:
cтруктурные – контролируют развитие признаков

Гены в клетках прокариот и эукариот имеют определенные особенности строения.
Гены в клетках прокариот и эукариот имеют определенные особенности строения.

У эукариот – прерывистую, он состоит из чередующихся участков – информативных

Экспрессия генов в биосинтезе белка
В процессе синтеза белка условно выделяют три
Экспрессия генов в биосинтезе белка
В процессе синтеза белка условно выделяют три

Биологической сущностью транскрипции является «переписывание» информации с молекулы ДНК на
Биологической сущностью транскрипции является «переписывание» информации с молекулы ДНК на

Синтез и-РНК в клетке всегда осуществляется от фосфатного конца
Синтез и-РНК в клетке всегда осуществляется от фосфатного конца

Транскрипция делится
на три периода:
инициация,
элонгация,
терминация.
Транскрипция делится
на три периода:
инициация,
элонгация,
терминация.

Инициация транскрипции
Синтез и-РНК осуществляется при помощи фермента РНК-полимеразы. *У прокариот имеется
Инициация транскрипции
Синтез и-РНК осуществляется при помощи фермента РНК-полимеразы. *У прокариот имеется

Фермент РНК-полимераза отыскивает в молекуле ДНК участок – промотор – и
Фермент РНК-полимераза отыскивает в молекуле ДНК участок – промотор – и
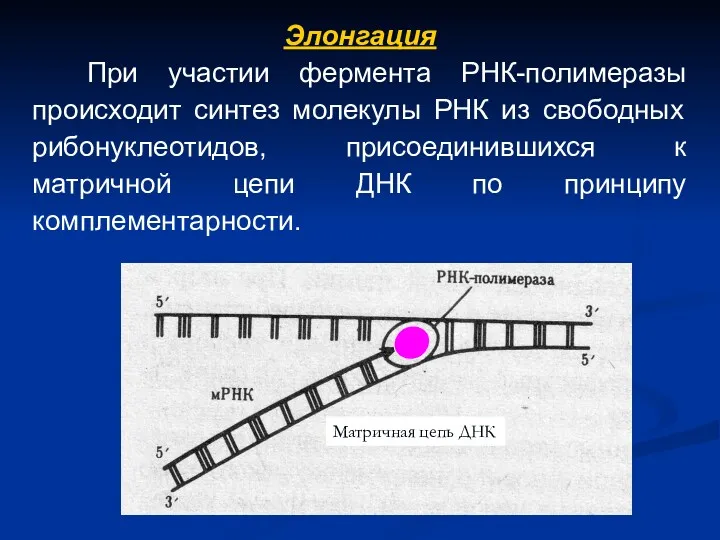
Элонгация
При участии фермента РНК-полимеразы происходит синтез молекулы РНК из свободных
Элонгация
При участии фермента РНК-полимеразы происходит синтез молекулы РНК из свободных

За 1 секунду между собой соединяются 50 нуклеотидов. Для клетки этой
За 1 секунду между собой соединяются 50 нуклеотидов. Для клетки этой

Терминация
Происходит тогда, когда РНК-полимераза достигает терминатора – участка ДНК, где прекращается
Терминация
Происходит тогда, когда РНК-полимераза достигает терминатора – участка ДНК, где прекращается
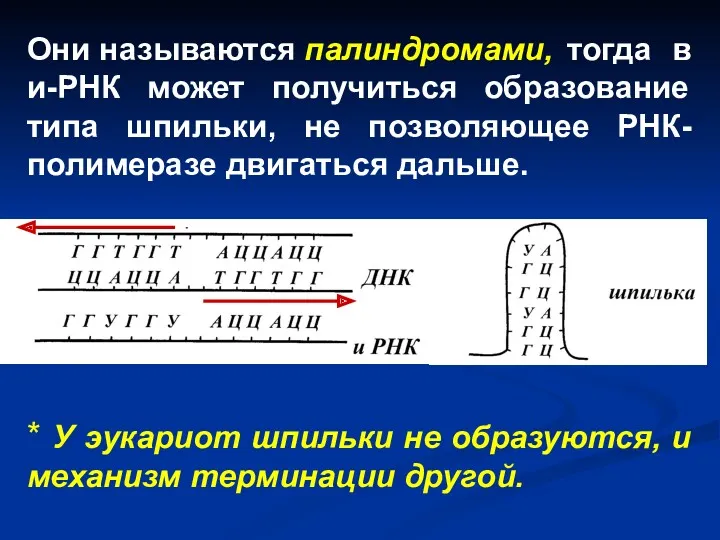
Они называются палиндромами, тогда в и-РНК может получиться образование типа шпильки, не
Они называются палиндромами, тогда в и-РНК может получиться образование типа шпильки, не

Процессинг
Процессинг включает целый ряд преобразований и-РНК, необходимых для ее нормального
Процессинг
Процессинг включает целый ряд преобразований и-РНК, необходимых для ее нормального

3. Удаление части нуклеотидов на гидроксильном конце.
4. Присоединение на гидроксильном конце
3. Удаление части нуклеотидов на гидроксильном конце.
4. Присоединение на гидроксильном конце
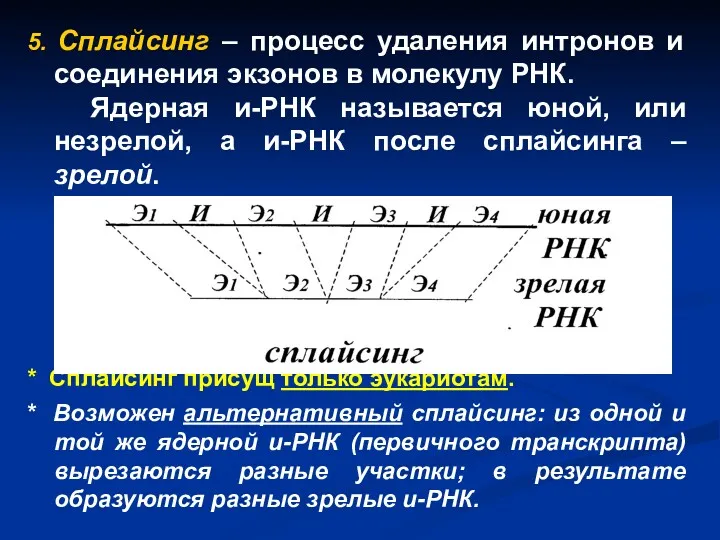
5. Сплайсинг – процесс удаления интронов и соединения экзонов в молекулу
5. Сплайсинг – процесс удаления интронов и соединения экзонов в молекулу

1 – лидирующий участок; АУГ - стартовый кодон; 2 –
1 – лидирующий участок; АУГ - стартовый кодон; 2 –

Трансляция
Трансляция – это процесс синтеза полипептидных цепей, осуществляемый в рибосомах.
Трансляция
Трансляция – это процесс синтеза полипептидных цепей, осуществляемый в рибосомах.
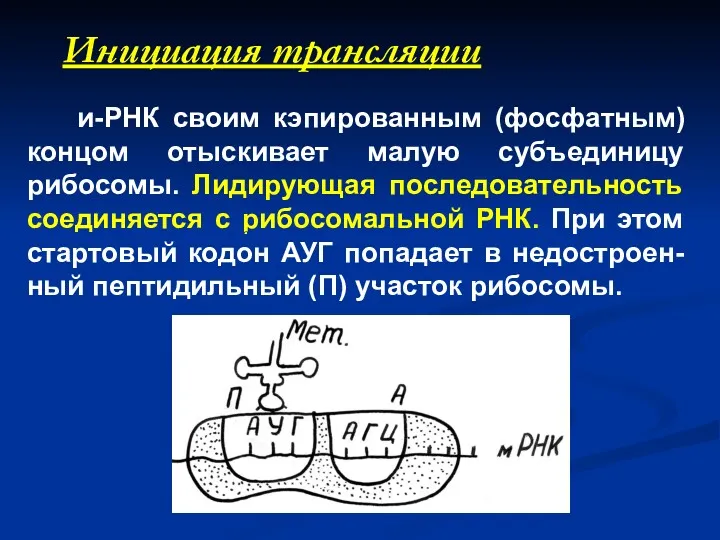
Инициация трансляции
и-РНК своим кэпированным (фосфатным) концом отыскивает малую субъединицу рибосомы.
Инициация трансляции
и-РНК своим кэпированным (фосфатным) концом отыскивает малую субъединицу рибосомы.
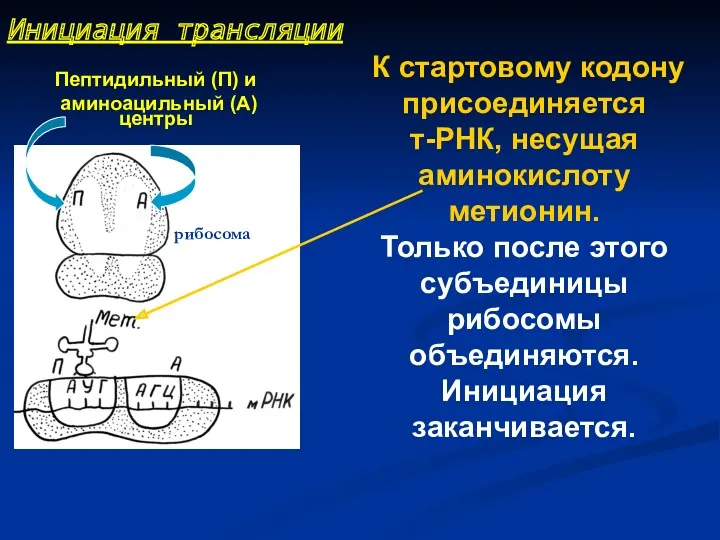
К стартовому кодону присоединяется т-РНК, несущая аминокислоту метионин.
Только после этого
Только после этого
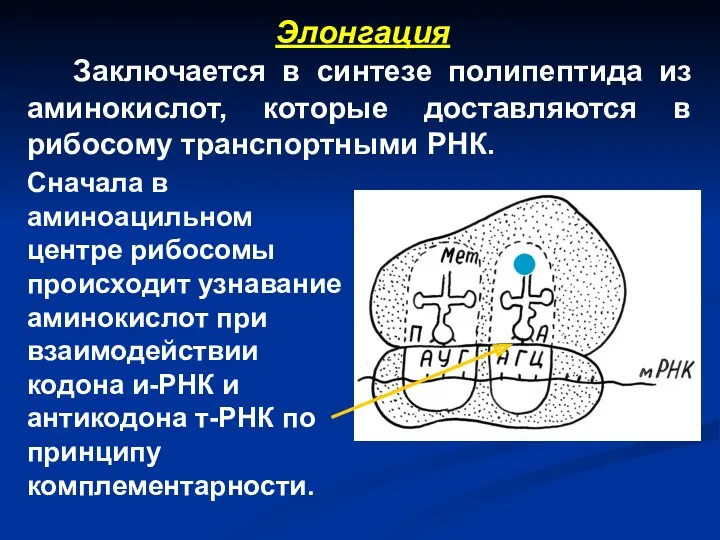
Элонгация
Заключается в синтезе полипептида из аминокислот, которые доставляются в рибосому
Элонгация
Заключается в синтезе полипептида из аминокислот, которые доставляются в рибосому
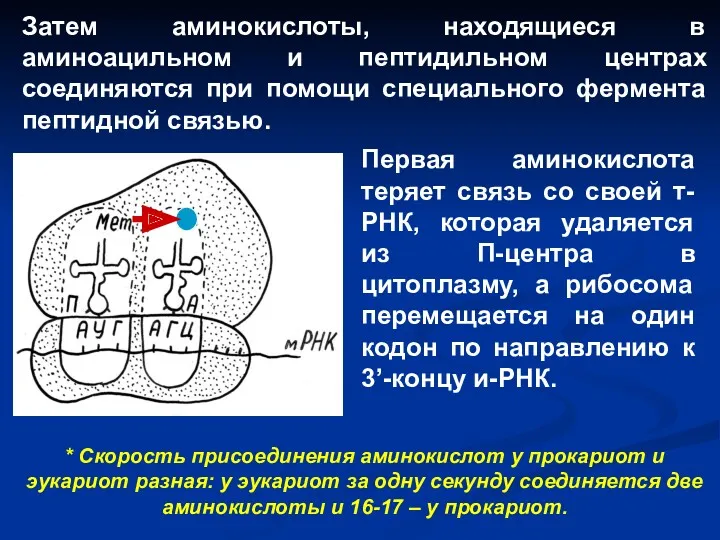
Затем аминокислоты, находящиеся в аминоацильном и пептидильном центрах соединяются при помощи
Затем аминокислоты, находящиеся в аминоацильном и пептидильном центрах соединяются при помощи

В результате вторая т-РНК и соединенный с ней дипептид оказываются в
В результате вторая т-РНК и соединенный с ней дипептид оказываются в
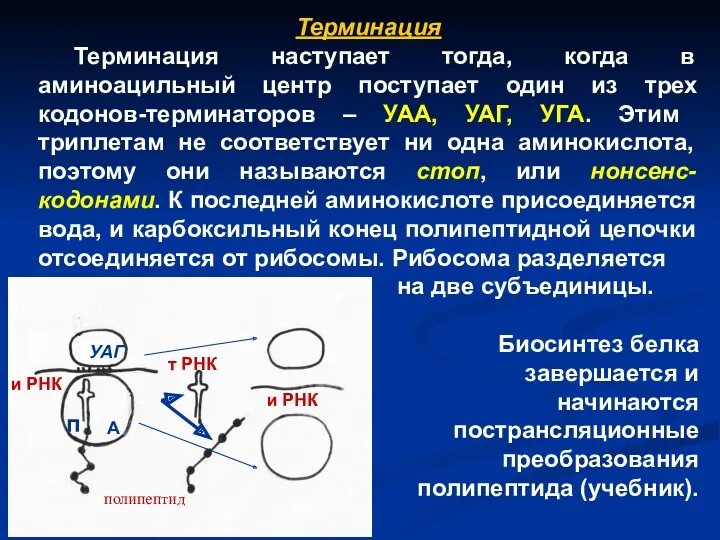
Терминация
Терминация наступает тогда, когда в аминоацильный центр поступает один из трех
Терминация
Терминация наступает тогда, когда в аминоацильный центр поступает один из трех

Регуляция экспрессии генов
Регуляция генной активности в клетках может происходить на всех
Регуляция экспрессии генов
Регуляция генной активности в клетках может происходить на всех

Кишечная палочка при попадании в среду, содержащую молочный сахар лактозу, вырабатывает
Кишечная палочка при попадании в среду, содержащую молочный сахар лактозу, вырабатывает

Схема lac-оперона. Р – ген-регулятор; П – промотор; О –
Схема lac-оперона. Р – ген-регулятор; П – промотор; О –
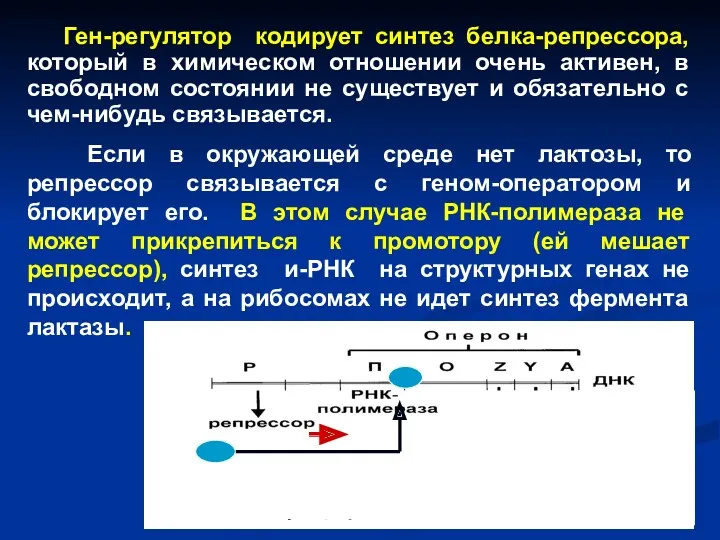
Ген-регулятор кодирует синтез белка-репрессора, который в химическом отношении очень активен, в
Ген-регулятор кодирует синтез белка-репрессора, который в химическом отношении очень активен, в
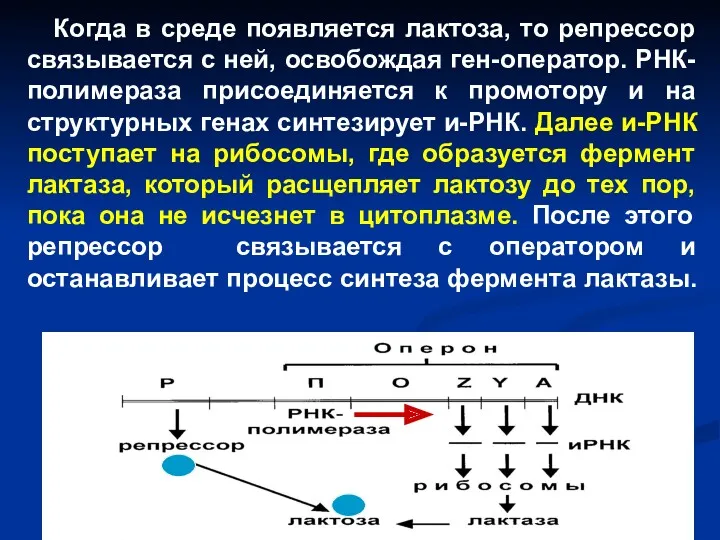
Когда в среде появляется лактоза, то репрессор связывается с ней,
Когда в среде появляется лактоза, то репрессор связывается с ней,
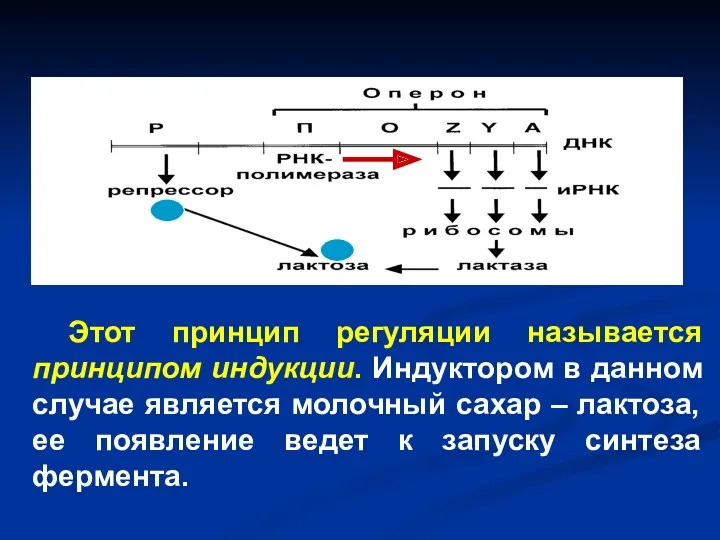
Этот принцип регуляции называется принципом индукции. Индуктором в данном случае является
Этот принцип регуляции называется принципом индукции. Индуктором в данном случае является

Возможен и другой принцип регуляции синтеза белка – принцип репрессии.
Возможен и другой принцип регуляции синтеза белка – принцип репрессии.

Принцип репрессии
Исходно белок-репрессор находится в неактивной форме, поэтому он
Принцип репрессии
Исходно белок-репрессор находится в неактивной форме, поэтому он
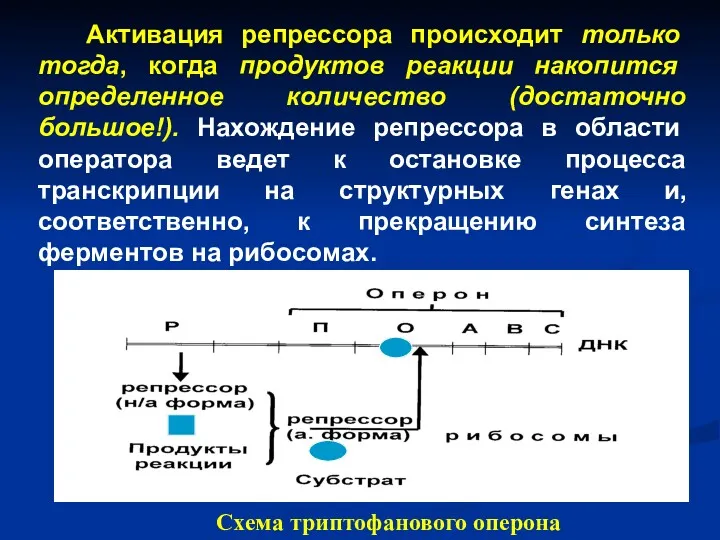
Активация репрессора происходит только тогда, когда продуктов реакции накопится определенное
Активация репрессора происходит только тогда, когда продуктов реакции накопится определенное

Особенности регуляции генной активности у эукариот
У эукариот принцип оперонной регуляции
Особенности регуляции генной активности у эукариот
У эукариот принцип оперонной регуляции

Один из таких участков находится перед промотором и называется препромоторным
Один из таких участков находится перед промотором и называется препромоторным

Разновидности генов
Наряду с функциональной классификацией генов существуют и другие их
Разновидности генов
Наряду с функциональной классификацией генов существуют и другие их

Мобильные (прыгающие) гены – гены, не имеющие постоянной локализации не
Мобильные (прыгающие) гены – гены, не имеющие постоянной локализации не

В последние десятилетия в генетике появилось еще одно новое понятие
В последние десятилетия в генетике появилось еще одно новое понятие
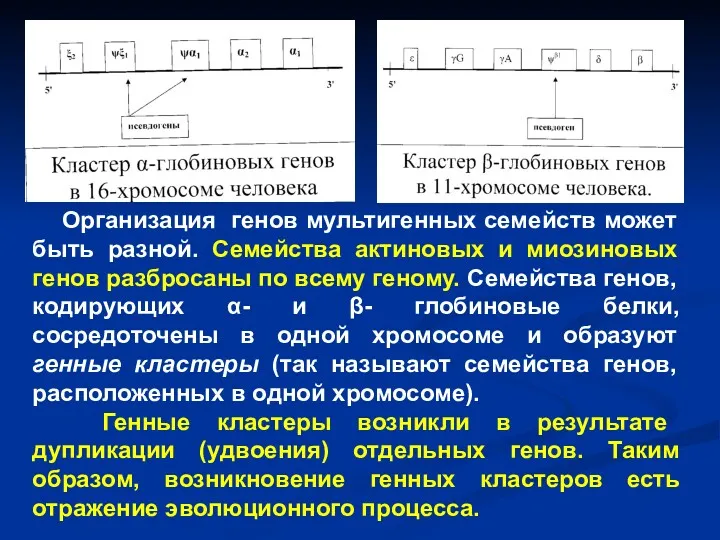
Организация генов мультигенных семейств может быть разной. Семейства актиновых и
Организация генов мультигенных семейств может быть разной. Семейства актиновых и
 Микробиологияның даму тарихы
Микробиологияның даму тарихы 5 класс, презентация с конспекту урока по биологии с использованием цифрового микроскопа на тему Строение клетки
5 класс, презентация с конспекту урока по биологии с использованием цифрового микроскопа на тему Строение клетки урок биологии в 8 классе Живая крепость
урок биологии в 8 классе Живая крепость Освоение суши растениями
Освоение суши растениями Понятие мутации
Понятие мутации Движение крови в организме
Движение крови в организме Кожа и ее производные
Кожа и ее производные Почему исчезли динозавры
Почему исчезли динозавры Эволюционное учение Чарльза Дарвина. Урок биологии в 11 классе
Эволюционное учение Чарльза Дарвина. Урок биологии в 11 классе Введение в биохимию. Строение и функции белков
Введение в биохимию. Строение и функции белков Роль марганца в организме человека
Роль марганца в организме человека Физиология продолговатого мозга и моста
Физиология продолговатого мозга и моста Предмет молекулярной биологии
Предмет молекулярной биологии Растения-хищники. 6 класс
Растения-хищники. 6 класс Якісний та кількісний склад комах-запилювачів рослин у межах Тячівського району Закарпаття
Якісний та кількісний склад комах-запилювачів рослин у межах Тячівського району Закарпаття Метаболизм клетки. Клеточное дыхание. Фотосинтез, хемосинтез
Метаболизм клетки. Клеточное дыхание. Фотосинтез, хемосинтез Презентация к уроку по биологии Цветок 6 класс
Презентация к уроку по биологии Цветок 6 класс Межвидовые взаимоотношения в мире микроорганизмов. (Лекция 6)
Межвидовые взаимоотношения в мире микроорганизмов. (Лекция 6) Отряд Чешуйчатые. Подотряд Ящерицы
Отряд Чешуйчатые. Подотряд Ящерицы Катаболизм. Цикл трикарбоновых кислот
Катаболизм. Цикл трикарбоновых кислот Пчелиный воск
Пчелиный воск Происхождение и начальные этапы развития жизни на Земле
Происхождение и начальные этапы развития жизни на Земле Спинной мозг. Строение
Спинной мозг. Строение Классификация витаминов
Классификация витаминов Обмен веществ и энергии
Обмен веществ и энергии Урок по ФГОС 5 класс Папоротникообразные
Урок по ФГОС 5 класс Папоротникообразные Охорона тварин
Охорона тварин презентация у уроку биологии Генетика человека
презентация у уроку биологии Генетика человека