- Сцепленное наследование

Содержание
- 2. В начале ХХ века исследователи пришли к выводу, что законы Менделя не всегда соблюдаются Под вопросом
- 3. Условия выполнения законов Менделя Моногенное наследование (элементарные признаки). Это означает, что за один признак отвечает один
- 4. Чистые гаметы - гаметы «чисты», т.е из пары аллелей в половых клетках присутствует один аллель. Выполняется
- 5. Соответствие законов Менделя поведению хромосом Г. Мендель считал, что признаки передаются из поколения в поколение как
- 6. К началу XX в. углубленное изучение поведения хромосом в ходе самовоспроизведения клеток, при созревании половых клеток,
- 7. Это привело немецкого цитолога и эмбриолога Бовери (1902—1907) и американского цитолога Сэттона (1902—1903) к утверждению тесной
- 8. Бовери и Сэттон заметили тесную взаимосвязь между менделевским наследованием признаков и поведением хромосом при образовании гамет
- 9. В частности, они отметили: хромосомы встречаются как гомологичные пары (Мендель считал, что наследственные факторы существуют в
- 10. хромосомы различных гомологичных пар группируются случайным образом при мейозе и распределяются в гаметы независимо от каждой
- 11. хромосомы сохраняют свою структуру, индивидуальность и генетический состав на протяжении жизненного цикла индивидуума (Мендель полагая, что
- 12. Стало очевидным, что основой Менделевских законов наследования является наличие генов в хромосомах и их поведение при
- 13. В 1906 году Бэтсон и Пеннет Стало ясно, что не для всех генов характерно независимое распределение
- 14. Наследование при полном (абсолютном) сцеплении генов Р: AaBb x aabb G: F1: AaBb, aabb А a
- 15. приступил к экспериментам в области генетики в 1909 г. Объектом исследований была избрана плодовая мушка -
- 16. Закон Моргана Первое скрещивание Анализирующее скрещивание Ожидалось получить 1:1:1:1
- 17. Закон Моргана Морган пришел к выводу, что гены обуславливающие развитие серой окраски тела и длинные крылья
- 19. Кроссинговер — обменом участками гомологичных хромосом в процессе их конъюгации в профазе мейоза I. Каждая из
- 21. Закон Моргана У самцов дрозофиллы кроссинговер не происходит 1:1
- 22. Закон Моргана Совместное наследование не аллельных генов – сцепление. Материальная основа сцепления – хромосома. Гены, локализованные
- 23. Профаза I
- 24. В зависимости от количества появившихся хиазм: одиночный кроссинговер - образуется только одна хиазма, что ведет к
- 25. Частота кроссинговера (расстояние между генами): число кроссоверных организмов = * 100% общее число потомков
- 26. Эта частота строго пропорциональна расстоянию между сцепленными генами и измеряется в морганидах 1 морганида соответствует 1%
- 27. Установлено, что вероятность кроссинговера между определенными сцепленными генами зависит от расстояния между ними в хромосоме Т.
- 28. Факторы, влияющие на частоту кроссинговера Температура – высокая и низкая температура повышает частоту Рентгеновские лучи –
- 29. Значение кроссинговера Кроссинговер — широко распространенное явление Он происходит практически у всех организмов, размножающихся половым путем
- 30. Хромосомная теория сцепления Моргана Гены, проявляющие сцепление, расположены в одной и той же хромосоме Гены расположены
- 31. Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну группу сцепления Количество групп сцепления
- 32. Работы Т. Моргана и его сотрудников подтвердили значение хромосом как основных носителей генов установили линейность расположения
- 33. Стадии профазы I на примере двух гомологичных хромосом На стадии лептотены начинают формироваться оси хромосом, в
- 34. Схема строения синаптонемного комплекса
- 35. Молекулярный механизм кроссинговера Двунитевые разрывы вносятся в ДНК с помощью белка SPO11 В местах разрывов образуются
- 37. Скачать презентацию

В начале ХХ века исследователи пришли к выводу, что законы Менделя
В начале ХХ века исследователи пришли к выводу, что законы Менделя

Условия выполнения законов Менделя
Моногенное наследование (элементарные признаки). Это означает, что за
Условия выполнения законов Менделя
Моногенное наследование (элементарные признаки). Это означает, что за

Чистые гаметы - гаметы «чисты», т.е из пары аллелей в половых
Чистые гаметы - гаметы «чисты», т.е из пары аллелей в половых

Соответствие законов Менделя поведению хромосом
Г. Мендель считал, что признаки передаются из
Соответствие законов Менделя поведению хромосом
Г. Мендель считал, что признаки передаются из

К началу XX в. углубленное изучение поведения хромосом
в ходе самовоспроизведения
К началу XX в. углубленное изучение поведения хромосом
в ходе самовоспроизведения

Это привело
немецкого цитолога и эмбриолога Бовери (1902—1907) и
американского цитолога
Это привело немецкого цитолога и эмбриолога Бовери (1902—1907) и американского цитолога

Бовери и Сэттон заметили тесную взаимосвязь между
менделевским наследованием признаков
и
Бовери и Сэттон заметили тесную взаимосвязь между
менделевским наследованием признаков
и

В частности, они отметили:
хромосомы встречаются как гомологичные пары (Мендель считал, что
В частности, они отметили:
хромосомы встречаются как гомологичные пары (Мендель считал, что

хромосомы различных гомологичных пар группируются случайным образом при мейозе и распределяются
хромосомы различных гомологичных пар группируются случайным образом при мейозе и распределяются

хромосомы сохраняют свою структуру, индивидуальность и генетический состав на протяжении жизненного
хромосомы сохраняют свою структуру, индивидуальность и генетический состав на протяжении жизненного

Стало очевидным, что основой Менделевских законов наследования является наличие генов в
Стало очевидным, что основой Менделевских законов наследования является наличие генов в
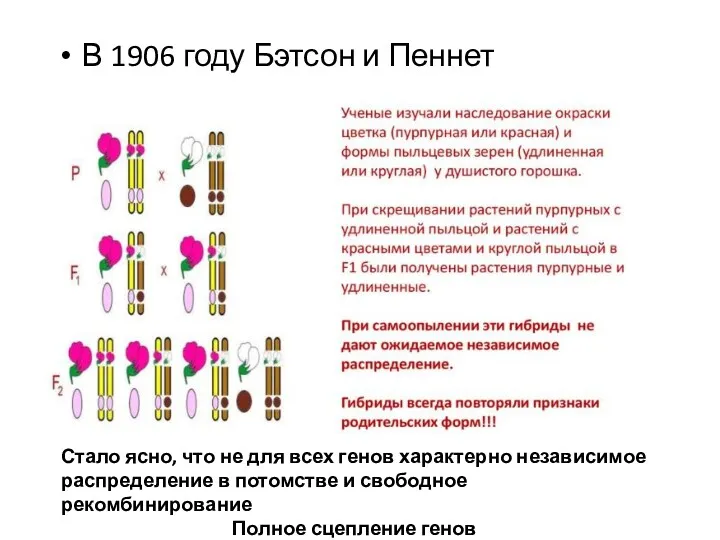
В 1906 году Бэтсон и Пеннет
Стало ясно, что не для всех
В 1906 году Бэтсон и Пеннет
Стало ясно, что не для всех
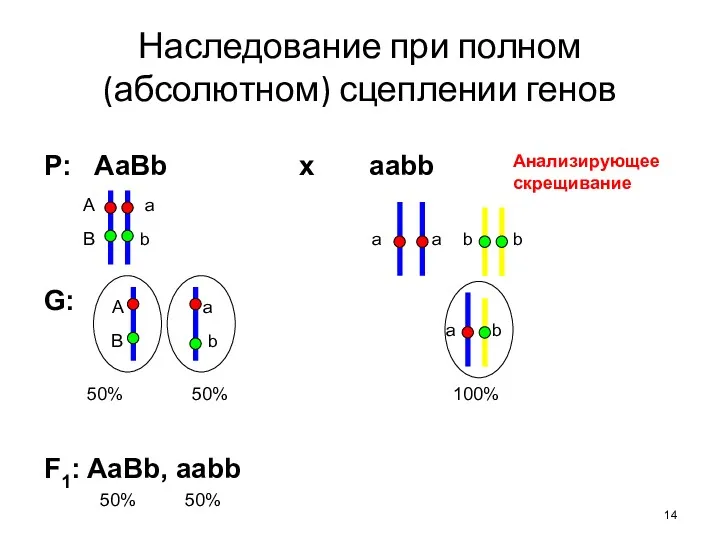
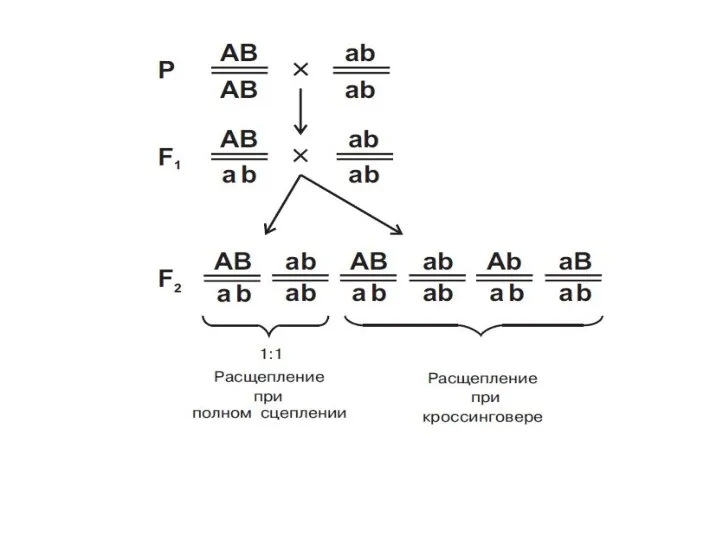
Наследование при полном (абсолютном) сцеплении генов
Р: AaBb x aabb
G:
F1: AaBb, aabb
А
Наследование при полном (абсолютном) сцеплении генов
Р: AaBb x aabb
G:
F1: AaBb, aabb
А

приступил к экспериментам в области генетики в 1909 г.
Объектом исследований
приступил к экспериментам в области генетики в 1909 г.
Объектом исследований
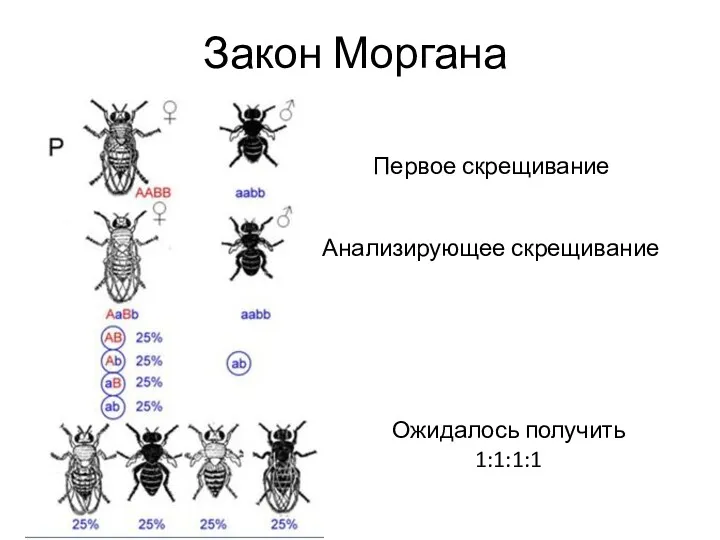
Закон Моргана
Первое скрещивание
Анализирующее скрещивание
Ожидалось получить
1:1:1:1
Закон Моргана
Первое скрещивание
Анализирующее скрещивание
Ожидалось получить
1:1:1:1

Закон Моргана
Морган пришел к выводу, что гены обуславливающие развитие серой окраски
Закон Моргана
Морган пришел к выводу, что гены обуславливающие развитие серой окраски
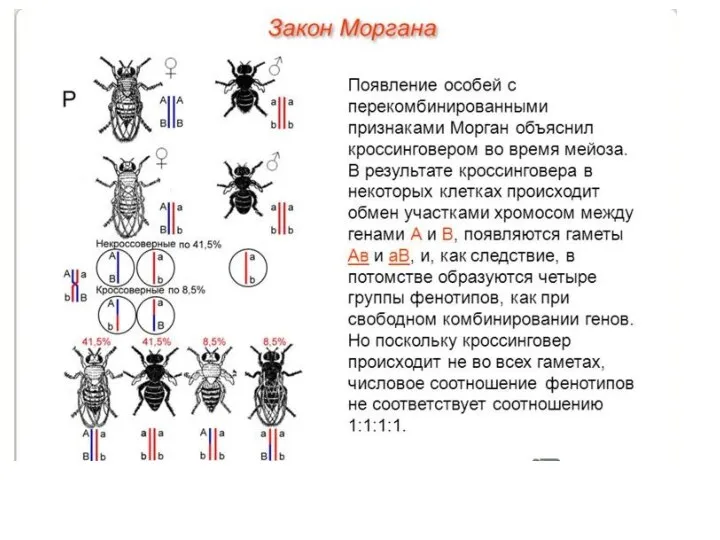
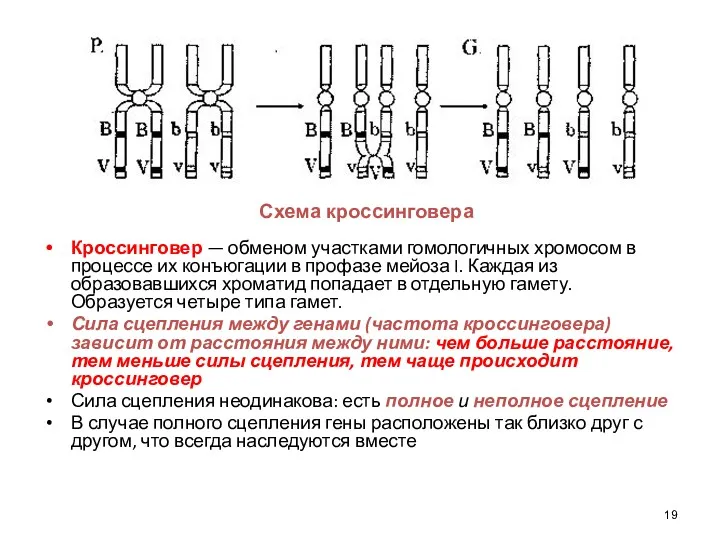
Кроссинговер — обменом участками гомологичных хромосом в процессе их конъюгации в
Кроссинговер — обменом участками гомологичных хромосом в процессе их конъюгации в
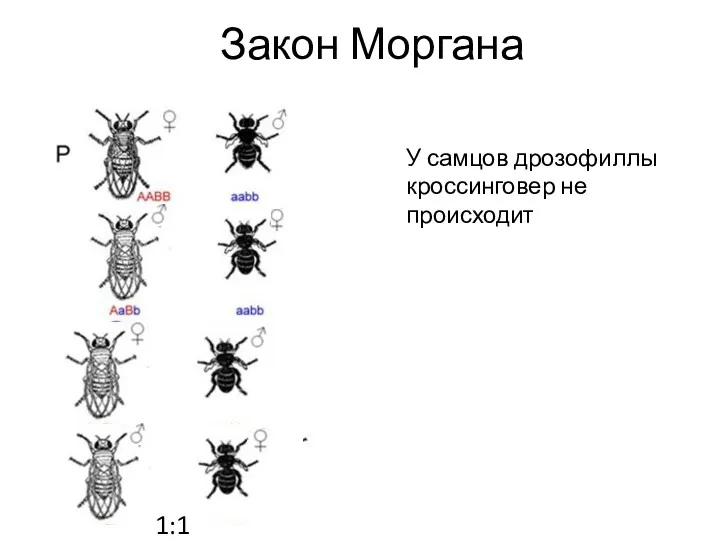
Закон Моргана
У самцов дрозофиллы кроссинговер не происходит
1:1
Закон Моргана
У самцов дрозофиллы кроссинговер не происходит
1:1

Закон Моргана
Совместное наследование не аллельных генов – сцепление.
Материальная основа сцепления –
Закон Моргана
Совместное наследование не аллельных генов – сцепление.
Материальная основа сцепления –
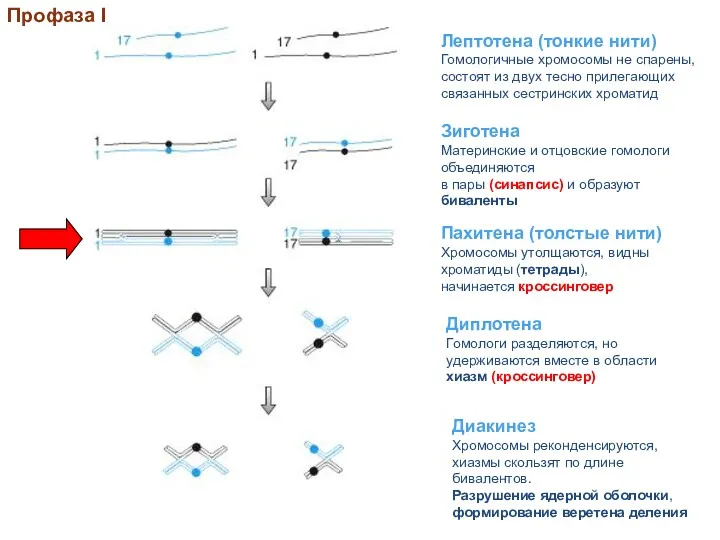
Профаза I
Профаза I

В зависимости от количества появившихся хиазм:
одиночный кроссинговер - образуется только
В зависимости от количества появившихся хиазм:
одиночный кроссинговер - образуется только

Частота кроссинговера (расстояние между генами):
число кроссоверных организмов
= * 100%
Частота кроссинговера (расстояние между генами):
число кроссоверных организмов
= * 100%

Эта частота строго пропорциональна расстоянию между сцепленными генами и измеряется в
Эта частота строго пропорциональна расстоянию между сцепленными генами и измеряется в

Установлено, что вероятность кроссинговера между определенными сцепленными генами зависит от расстояния
Установлено, что вероятность кроссинговера между определенными сцепленными генами зависит от расстояния

Факторы, влияющие на частоту кроссинговера
Температура – высокая и низкая температура повышает
Факторы, влияющие на частоту кроссинговера
Температура – высокая и низкая температура повышает

Значение кроссинговера
Кроссинговер — широко распространенное явление
Он происходит практически у всех организмов,
Значение кроссинговера
Кроссинговер — широко распространенное явление
Он происходит практически у всех организмов,

Хромосомная теория сцепления Моргана
Гены, проявляющие сцепление, расположены в одной и той
Хромосомная теория сцепления Моргана
Гены, проявляющие сцепление, расположены в одной и той

Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну
Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну

Работы Т. Моргана и его сотрудников подтвердили
значение хромосом как основных
Работы Т. Моргана и его сотрудников подтвердили
значение хромосом как основных
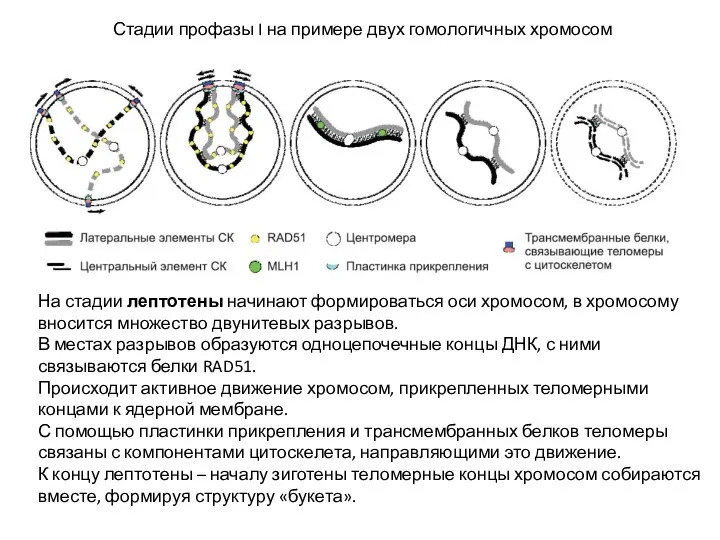
Стадии профазы I на примере двух гомологичных хромосом
На стадии лептотены начинают
Стадии профазы I на примере двух гомологичных хромосом
На стадии лептотены начинают
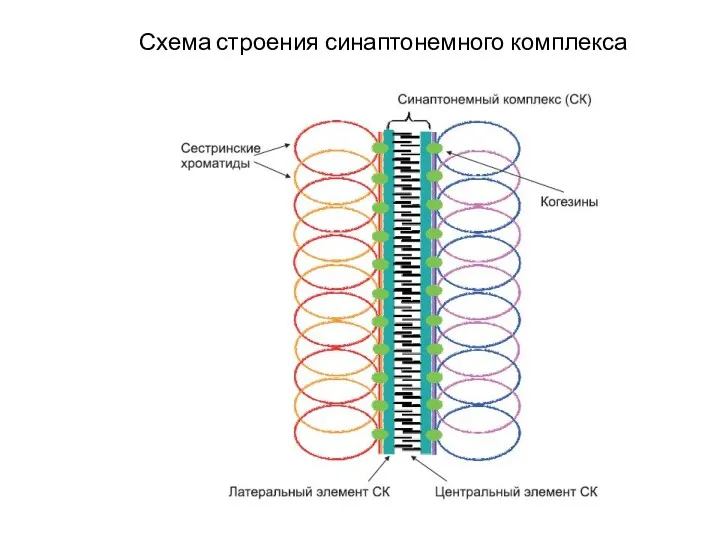
Схема строения синаптонемного комплекса
Схема строения синаптонемного комплекса
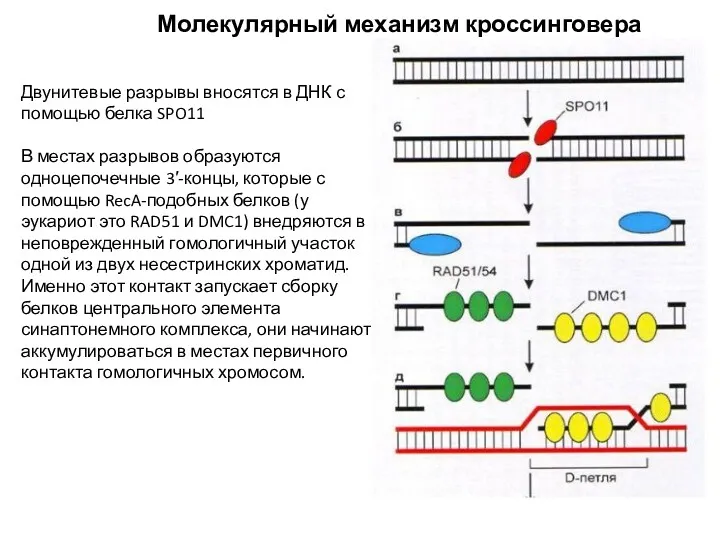
Молекулярный механизм кроссинговера
Двунитевые разрывы вносятся в ДНК с
помощью белка SPO11
В местах
Молекулярный механизм кроссинговера
Двунитевые разрывы вносятся в ДНК с
помощью белка SPO11
В местах
 Закономерности развития живой природы. Эволюционное учение
Закономерности развития живой природы. Эволюционное учение Останки мамонтов в России
Останки мамонтов в России Высшая нервная деятельность. Физиологические основы поведения человека
Высшая нервная деятельность. Физиологические основы поведения человека Опорно-двигательный аппарат человека
Опорно-двигательный аппарат человека Исследовательская работа по теме: Экологическая безопасность детских игрушек
Исследовательская работа по теме: Экологическая безопасность детских игрушек Строение и работа скелетных мышц. (8 класс)
Строение и работа скелетных мышц. (8 класс) Конституциональная антропология
Конституциональная антропология Introduction to artificial intelligence
Introduction to artificial intelligence Историческое развитие растительного мира
Историческое развитие растительного мира Развитие органического мира
Развитие органического мира Культивирование бурых водорослей
Культивирование бурых водорослей Клеточное строение стебля
Клеточное строение стебля Питание и пищеварение
Питание и пищеварение Многообразие организмов
Многообразие организмов Постэмбриональный период развития человека
Постэмбриональный период развития человека Филогенетический анализ и молекулярная эволюция. Лекция 5
Филогенетический анализ и молекулярная эволюция. Лекция 5 Обмен веществ и энергии. Роль белков, жиров и углеводов, их превращения в организме. Водно-солевой обмен. Терморегуляция
Обмен веществ и энергии. Роль белков, жиров и углеводов, их превращения в организме. Водно-солевой обмен. Терморегуляция Cell division
Cell division Клетка — элементарная структурная и функциональная единица живого
Клетка — элементарная структурная и функциональная единица живого Систематика растений
Систематика растений Экологическая дифференциация человечества
Экологическая дифференциация человечества Индивидуальное развитие организмов (онтогенез)
Индивидуальное развитие организмов (онтогенез) Лист. Внешнее и внутреннее строение
Лист. Внешнее и внутреннее строение Понятие биологического вида
Понятие биологического вида Оплодотворение. Этапы
Оплодотворение. Этапы Антропогенез – процесс возникновения и развития человека как биологического вида
Антропогенез – процесс возникновения и развития человека как биологического вида Микрофлора полости рта. Микробиология
Микрофлора полости рта. Микробиология Основы палеонтологии. Лекция 1
Основы палеонтологии. Лекция 1