- Транспорт веществ через биологические мембраны. (Лекция 12)

Содержание
- 2. Элементарная живая система, способная к самостоятельному существованию, развитию и воспроизведению – это живая клетка, являющаяся основой
- 3. С точки зрения термодинамики клетка – открытая термодинамическая система. Это значит, что для поддержания постоянства своего
- 4. С другой стороны, клетка должна быть автономна по отношению к окружающей среде, т. е. вещество клетки
- 5. Единство автономности от окружающей среды и одновременно тесной связи с окружающей средой – необходимое условие функционирования
- 6. Поэтому важнейшее условие существования клетки (а следовательно - и жизни) – нормальное функционирование биологических мембран.
- 7. Биологическая мембрана – это функционально активная белково-липидная структура, образующая границу раздела между клеткой и окружающей средой
- 9. Суммарная масса внутриклеточных мембран достигает 2/3 общей массы обезвоженной клетки. В печени крысы массой около 6
- 10. Чем больше отношение суммарной площади мембран к объему клетки, тем выше интенсивность обменных процессов в клетке.
- 11. 1. Функции биологических мембран : Механическая – обеспечивает прочность и автономность клетки, внутриклеточных структур. Барьерно-транспортная -
- 12. Матричная – обеспечивает определенное взаимное расположение и ориентацию мембранных белков, их оптимальное взаимодействие (например, оптимальное взаимодействие
- 13. Генерация и проведение биопотенциалов. Рецепторная – некоторые белки, находящиеся в мембранах, являются рецепторами (молекулами, при помощи
- 14. Ферментативная – многие мембранные белки являются ферментами. Маркировка клетки - на мембране есть антигены, действующие как
- 15. 2. Структура биологических мембран Согласно современным представлениям, все клеточные и внутриклеточные мембраны устроены сходным образом: основу
- 16. В состав плазматической мембраны эукариотических клеток входят также полисахариды. Их короткие, сильно развлетвленные молекулы ковалентно связаны
- 17. Жидкостно-мозаичная модель биологической мембраны (Сингер и Николсон, 1972)
- 18. Основные виды мембранных липидов: 1) Фосфолипиды; 2) Сфингомиелины; 3) Гликолипиды; 4) Холестерол.
- 19. Преобладают среди них фосфолипиды, в молекуле которых условно выделяют головку, тело (шейку) и два хвоста. Вертикальный
- 20. Тело ФЛ образовано одним из многоатомных спиртов – глицерином или сфингозином. Соответственно, выделяют глицерофосфолипиды и сфингофосфолипиды.
- 21. Из-за большого количества гетероатомов головки обычно являются полярными. Через молекулу ортофосфорной кислоты они соединены с телом.
- 22. Жирные кислоты могут быть насыщенными или ненасыщенными. Обычно у двуцепочечных фософлипидов один из хвостов представлен насыщенной,
- 25. Основная особенность молекул фосфолипидов – их амфифильность. Полярная (заряженная) головка гидрофильна, неполярные жирнокислотные хвосты гидрофобны.
- 28. Бимолекулярный слой ФЛ, образующий биомембраны, в физиологических условиях представляет собой жидкий кристалл. В жидком кристалле сочетаются
- 29. Вязкость мембранных липидов во многом определяет свойства и поведение мембран. На вязкость липидов в бислое влияют
- 30. Чем длиннее и насыщеннее жирнокислотные цепи липидов, тем плотнее они могут быть упакованы. Плотная упаковка повышает
- 31. Вязкость биологических мембран составляет от 0,03 до 0,1 Па.с (т.е. в 30-100 раз больше, чем у
- 32. Виды движения молекул фосфолипидов: 1) Движение в пределах одного слоя (латеральная миграция); 2) Вращение вокруг собственной
- 33. Таким образом, в мембране обычной клетки млекопитающего липиды находятся в непрерывном движении и постоянно перемещаются. Среднее
- 34. Мембранные белки арзличаются: а) по положению в мембране (интегральные, полуинтегральные, поверхностные);
- 35. б) по функциям: 1. Структурные белки. 2. Транспортные белки (белки-переносчики, белки-каналы). 3. Белки, обеспечивающие непосредственное межклеточное
- 36. 3. Классификация процессов транспорта в биологических мембранах Прежде всего, принципиально различными являются: а) Транспорт низкомолекулярных веществ;
- 37. Транспорт низкомолекулярных соединений Пассивный транспорт 1) Перенос веществ осуществляется по преобладающему физико-химическому градиенту; 2) Без затраты
- 38. Понятие физико-химического градиента Градиентом физической величины называют скорость изменения этой величины в пространстве, т.е.
- 39. Например, а) градиент концентрации – б) градиент температуры – в) градиент электрического потенциала - и т.д.
- 40. Градиент – векторная величина. Вектор градиента направлен в сторону возрастания физической величины. Понятие градиента применимо к
- 41. В случае биологических мембран обычно используют среднее значение градиента. Например, в случае градиента концентрации: где С1
- 42. Виды пассивного транспорта: 1) Свободная диффузия липофильных веществ через фосфолипидный бислой. 2) Облегченная диффузия неэлектролитов. 3)
- 43. 4. Свободная диффузия липофильных (незаряженных) веществ через ФЛ-бислой Диффузия - это процесс переноса вещества (массы) из
- 44. Диффузия незаряженных частиц вызывается их концентрационным градиентом и направлена в сторону уменьшения этого градиента.
- 45. Диффузия постепенно уменьшает градиент концентрации до тех пор, пока не наступит состояние равновесия. Диффузия является пассивным
- 46. Для количественной характеристики диффузии используют физическую величину - поток вещества (Ф) : то есть поток вещества
- 47. Отношение потока вещества к площади, через которую он происходит, называется плотностью потока:
- 48. Уравнение диффузии (уравнение Фика) Знак «-» показывает, что поток направлен в сторону уменьшения концентрации (т.е. противоположную
- 49. D - коэффициент диффузии. (формула Стокса-Эйнштейна) Здесь R - универсальная газовая постоянная, Т - абсолютная температура;
- 50. Для биологической мембраны существенное значение имеет коэффициент распределения вещества между липидным слоем и водой. Поэтому уравнение
- 51. Если ввести коэффициент проницаемости то уравнение Фика приобретает вид:
- 52. Посредством простой диффузии через ФЛ-бислой проникают низкомолекулярные гидрофобные органические вещества (жирные кислоты, мочевина, эфиры, жирорастворимые витамины,
- 53. 5. Облегченная диффузия гидрофильных молекул Крупные гидрофильные молекулы (сахара, аминокислоты) перемещаются через мембраны с помощью специальных
- 54. Этот тип транспорта мембраны является одним из видов диффузии, поскольку транспортируемое вещество перемещается по градиенту концентрации.
- 56. Другой особенностью облегченной диффузии является феномен насыщения. Поток вещества, транспортируемого путём облегченной диффузии, растёт в зависимости
- 58. Кинетику облегченной диффузии отображает уравнение Михаэлиса-Ментен: KM – константа Михаэлиса (равна концентрации вещества вне клетки или
- 59. 6. Электродиффузия Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационного и электрического градиентов. Липидный
- 60. Движущей силой диффузии является не только разность концентрации ионов внутри и вне клетки, но также разность
- 61. Электрохимический потенциал определяет свободную энергию иона и учитывает все силы, способные побудить ион к движению.
- 62. Здесь: μ0- стандартный химический потенциал, который зависит от химической природы вещества и температуры, R - универсальная
- 63. Зависимость плотности потока ионов J от электрохимического градиента определяется уравнением Теорелла: где U - подвижность ионов,
- 64. Подставляя выражение для электрохимического потенциала в уравнение Теорелла, можно получить уравнение Нернста-Планка с учётом двух градиентов,
- 65. Ионные каналы мембраны представляют собой интегральные белки мембраны, которые образуют отверстия в мембране, заполненные водой. В
- 66. Существуют натриевые, калиевые, кальциевые и хлорные каналы. Каждый из них имеет так называемый селективный фильтр, который
- 67. Проницаемость ионных каналов может изменяться благодаря наличию ворот - определенных групп атомов в составе белков, формирующих
- 68. Механизмы регуляции положения ворот могут отличаться в различных каналах. Некоторые из них открываются при изменениях электрического
- 70. 7. Активный транспорт Существует несколько систем активного транспорта ионов в плазматической мембране (ионные насосы): 1) Натрий-калиевый
- 71. Натрий-калиевый насос существует в плазматических мембранах всех животных и растительных клеток. Он выкачивает ионы натрия из
- 72. Натрий-калиевый насос - один из интегральных белков мембраны. Он обладает энзимными свойствами и способен гидролизовать аденозинтрифосфорную
- 73. Молекула насоса существует в двух основных конформациях, взаимное преобразование которых стимулируется гидролизом ATФ. При повышении концентрации
- 74. При расщеплении натрий-калиевой АТФазой молекулы ATФ неорганический фосфат присоединяется к белку. Конформация натрий-калиевой АТФазы изменяется, три
- 75. Затем молекула неорганического фосфата отсоединяется от насоса-белка, и насос превращается в переносчик калия. В результате два
- 76. Один натрий-калиевый насос может перенести через мембрану 150- 600 ионов натрия в секунду. Следствием его работы
- 78. Скачать презентацию

Элементарная живая система, способная к самостоятельному существованию, развитию и воспроизведению –
Элементарная живая система, способная к самостоятельному существованию, развитию и воспроизведению –

С точки зрения термодинамики клетка – открытая термодинамическая система.
Это значит,
С точки зрения термодинамики клетка – открытая термодинамическая система.
Это значит,

С другой стороны, клетка должна быть автономна по отношению к окружающей
С другой стороны, клетка должна быть автономна по отношению к окружающей

Единство автономности от окружающей среды и одновременно тесной связи с окружающей
Единство автономности от окружающей среды и одновременно тесной связи с окружающей

Поэтому важнейшее условие существования клетки (а следовательно - и жизни) –

Биологическая мембрана – это функционально активная белково-липидная структура, образующая границу раздела
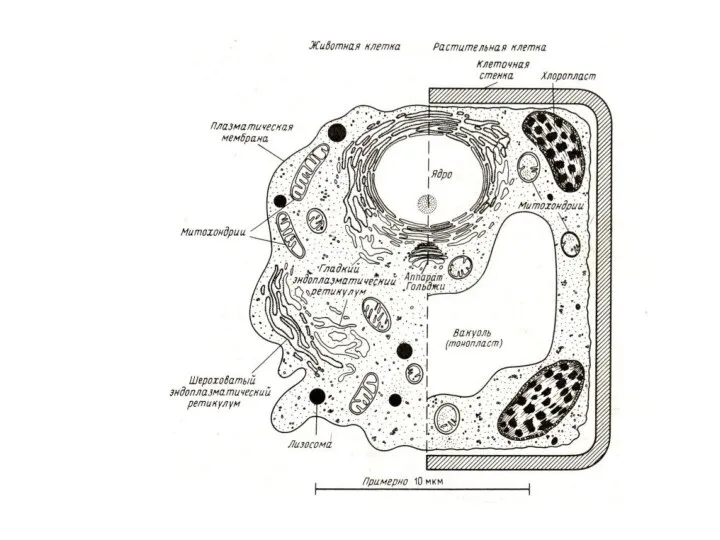

Суммарная масса внутриклеточных мембран достигает 2/3 общей массы обезвоженной клетки.
В печени
Суммарная масса внутриклеточных мембран достигает 2/3 общей массы обезвоженной клетки.
В печени

Чем больше отношение суммарной площади мембран к объему клетки, тем выше

1. Функции биологических мембран :
Механическая – обеспечивает прочность и автономность клетки,
1. Функции биологических мембран :
Механическая – обеспечивает прочность и автономность клетки,

Матричная – обеспечивает определенное взаимное расположение и ориентацию мембранных белков, их
Матричная – обеспечивает определенное взаимное расположение и ориентацию мембранных белков, их

Генерация и проведение биопотенциалов.
Рецепторная – некоторые белки, находящиеся в мембранах, являются
Генерация и проведение биопотенциалов.
Рецепторная – некоторые белки, находящиеся в мембранах, являются

Ферментативная – многие мембранные белки являются ферментами.
Маркировка клетки - на мембране
Ферментативная – многие мембранные белки являются ферментами.
Маркировка клетки - на мембране

2. Структура биологических мембран
Согласно современным представлениям, все клеточные и внутриклеточные мембраны
2. Структура биологических мембран
Согласно современным представлениям, все клеточные и внутриклеточные мембраны

В состав плазматической мембраны эукариотических клеток входят также полисахариды.
Их короткие,
В состав плазматической мембраны эукариотических клеток входят также полисахариды.
Их короткие,

Жидкостно-мозаичная модель биологической мембраны (Сингер и Николсон, 1972)
Жидкостно-мозаичная модель биологической мембраны (Сингер и Николсон, 1972)

Основные виды мембранных липидов:
1) Фосфолипиды;
2) Сфингомиелины;
3) Гликолипиды;
4) Холестерол.
Основные виды мембранных липидов:
1) Фосфолипиды;
2) Сфингомиелины;
3) Гликолипиды;
4) Холестерол.

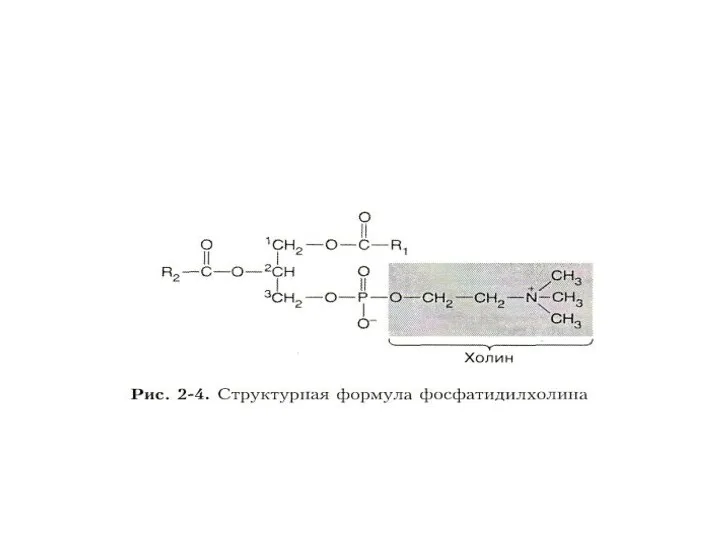
Преобладают среди них фосфолипиды, в молекуле которых условно выделяют головку, тело
Преобладают среди них фосфолипиды, в молекуле которых условно выделяют головку, тело

Тело ФЛ образовано одним из многоатомных спиртов – глицерином или сфингозином.
Тело ФЛ образовано одним из многоатомных спиртов – глицерином или сфингозином.

Из-за большого количества гетероатомов головки обычно являются полярными.
Через молекулу ортофосфорной
Из-за большого количества гетероатомов головки обычно являются полярными.
Через молекулу ортофосфорной

Жирные кислоты могут быть насыщенными или ненасыщенными.
Обычно у двуцепочечных фософлипидов
Жирные кислоты могут быть насыщенными или ненасыщенными.
Обычно у двуцепочечных фософлипидов
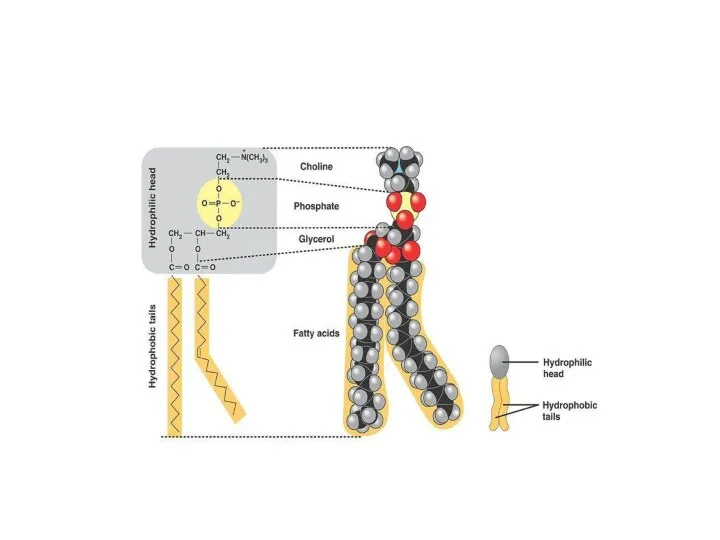

Основная особенность молекул фосфолипидов – их амфифильность.
Полярная (заряженная) головка гидрофильна, неполярные
Основная особенность молекул фосфолипидов – их амфифильность.
Полярная (заряженная) головка гидрофильна, неполярные

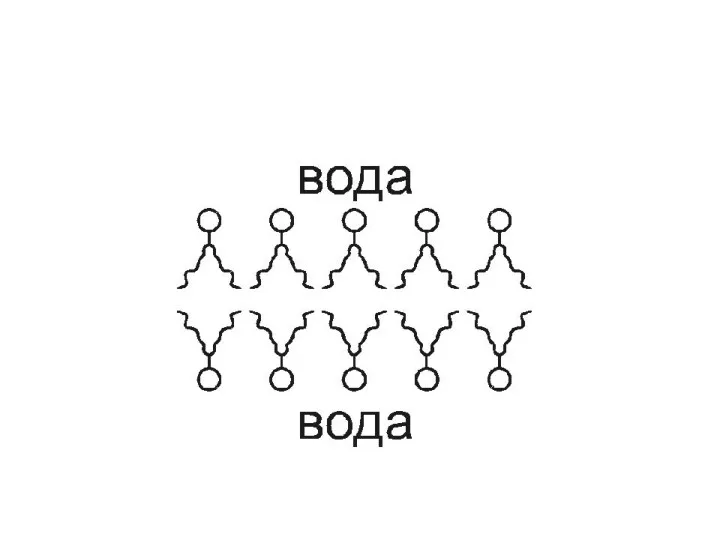

Бимолекулярный слой ФЛ, образующий биомембраны, в физиологических условиях представляет собой жидкий
Бимолекулярный слой ФЛ, образующий биомембраны, в физиологических условиях представляет собой жидкий

Вязкость мембранных липидов во многом определяет свойства и поведение мембран.
На вязкость
Вязкость мембранных липидов во многом определяет свойства и поведение мембран.
На вязкость

Чем длиннее и насыщеннее жирнокислотные цепи липидов, тем плотнее они могут
Чем длиннее и насыщеннее жирнокислотные цепи липидов, тем плотнее они могут

Вязкость биологических мембран составляет от 0,03 до 0,1 Па.с (т.е. в
Вязкость биологических мембран составляет от 0,03 до 0,1 Па.с (т.е. в

Виды движения молекул фосфолипидов:
1) Движение в пределах одного слоя (латеральная
Виды движения молекул фосфолипидов:
1) Движение в пределах одного слоя (латеральная

Таким образом, в мембране обычной клетки млекопитающего липиды находятся в непрерывном
Таким образом, в мембране обычной клетки млекопитающего липиды находятся в непрерывном
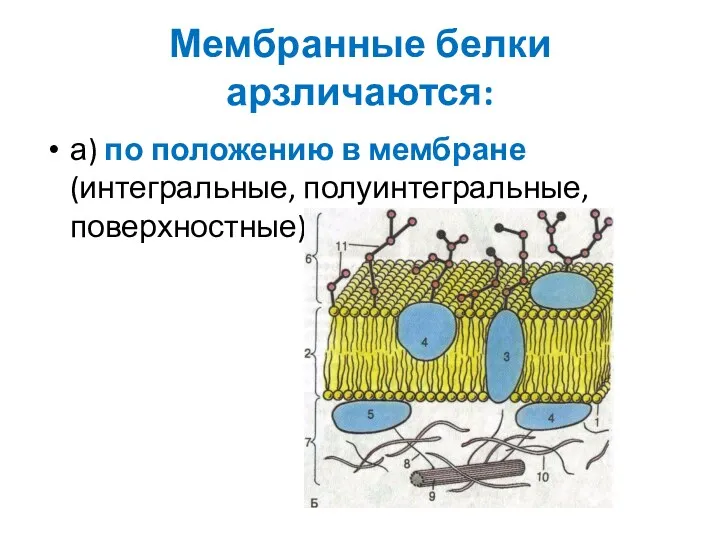
Мембранные белки арзличаются:
а) по положению в мембране (интегральные, полуинтегральные, поверхностные);
Мембранные белки арзличаются:
а) по положению в мембране (интегральные, полуинтегральные, поверхностные);

б) по функциям:
1. Структурные белки.
2. Транспортные белки (белки-переносчики, белки-каналы).
3. Белки, обеспечивающие
б) по функциям:
1. Структурные белки.
2. Транспортные белки (белки-переносчики, белки-каналы).
3. Белки, обеспечивающие

3. Классификация процессов транспорта в биологических мембранах
Прежде всего, принципиально различными являются:
3. Классификация процессов транспорта в биологических мембранах
Прежде всего, принципиально различными являются:

Транспорт низкомолекулярных соединений
Пассивный транспорт
1) Перенос веществ осуществляется по преобладающему физико-химическому градиенту;
2)
Транспорт низкомолекулярных соединений
Пассивный транспорт
1) Перенос веществ осуществляется по преобладающему физико-химическому градиенту;
2)

Понятие физико-химического градиента
Градиентом физической величины называют скорость изменения этой величины в
Понятие физико-химического градиента
Градиентом физической величины называют скорость изменения этой величины в

Например,
а) градиент концентрации –
б) градиент температуры –
в) градиент электрического
потенциала -
и
Например,
а) градиент концентрации –
б) градиент температуры –
в) градиент электрического
потенциала -
и

Градиент – векторная величина.
Вектор градиента направлен в сторону возрастания физической величины.
Понятие
Градиент – векторная величина.
Вектор градиента направлен в сторону возрастания физической величины.
Понятие

В случае биологических мембран обычно используют среднее значение градиента.
Например, в случае
В случае биологических мембран обычно используют среднее значение градиента.
Например, в случае

Виды пассивного транспорта:
1) Свободная диффузия липофильных веществ через фосфолипидный бислой.
2) Облегченная
Виды пассивного транспорта:
1) Свободная диффузия липофильных веществ через фосфолипидный бислой.
2) Облегченная

4. Свободная диффузия липофильных (незаряженных) веществ через ФЛ-бислой
Диффузия - это процесс
4. Свободная диффузия липофильных (незаряженных) веществ через ФЛ-бислой
Диффузия - это процесс

Диффузия незаряженных частиц вызывается их концентрационным градиентом и направлена в сторону

Диффузия постепенно уменьшает градиент концентрации до тех пор, пока не наступит
Диффузия постепенно уменьшает градиент концентрации до тех пор, пока не наступит

Для количественной характеристики диффузии используют физическую величину - поток вещества (Ф)
Для количественной характеристики диффузии используют физическую величину - поток вещества (Ф)

Отношение потока вещества к площади, через которую он происходит, называется плотностью
Отношение потока вещества к площади, через которую он происходит, называется плотностью

Уравнение диффузии (уравнение Фика)
Знак «-» показывает, что поток направлен в сторону
Уравнение диффузии (уравнение Фика)
Знак «-» показывает, что поток направлен в сторону

D - коэффициент диффузии.
(формула Стокса-Эйнштейна)
Здесь R - универсальная газовая постоянная,
Т
D - коэффициент диффузии.
(формула Стокса-Эйнштейна)
Здесь R - универсальная газовая постоянная,
Т

Для биологической мембраны существенное значение имеет коэффициент распределения вещества между липидным
Для биологической мембраны существенное значение имеет коэффициент распределения вещества между липидным

Если ввести коэффициент проницаемости
то уравнение Фика приобретает вид:
Если ввести коэффициент проницаемости
то уравнение Фика приобретает вид:

Посредством простой диффузии через ФЛ-бислой проникают низкомолекулярные гидрофобные органические вещества (жирные
Посредством простой диффузии через ФЛ-бислой проникают низкомолекулярные гидрофобные органические вещества (жирные

5. Облегченная диффузия гидрофильных молекул
Крупные гидрофильные молекулы (сахара, аминокислоты) перемещаются через
5. Облегченная диффузия гидрофильных молекул
Крупные гидрофильные молекулы (сахара, аминокислоты) перемещаются через

Этот тип транспорта мембраны является одним из видов диффузии, поскольку транспортируемое
Этот тип транспорта мембраны является одним из видов диффузии, поскольку транспортируемое
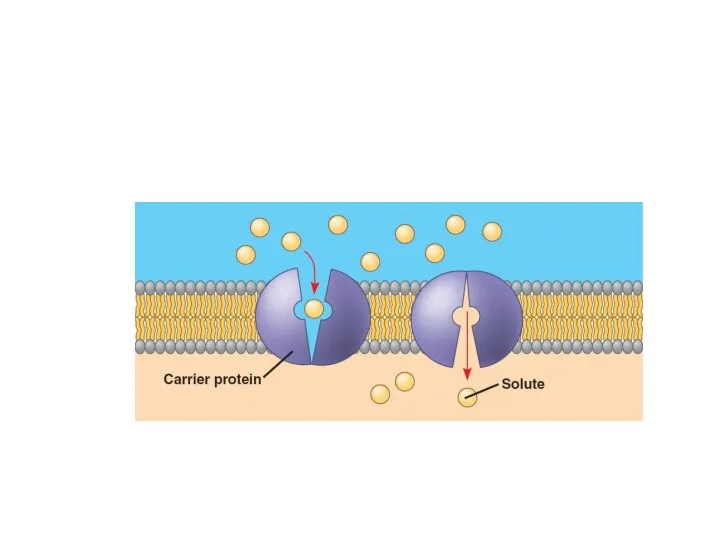

Другой особенностью облегченной диффузии является феномен насыщения.
Поток вещества, транспортируемого путём
Другой особенностью облегченной диффузии является феномен насыщения.
Поток вещества, транспортируемого путём
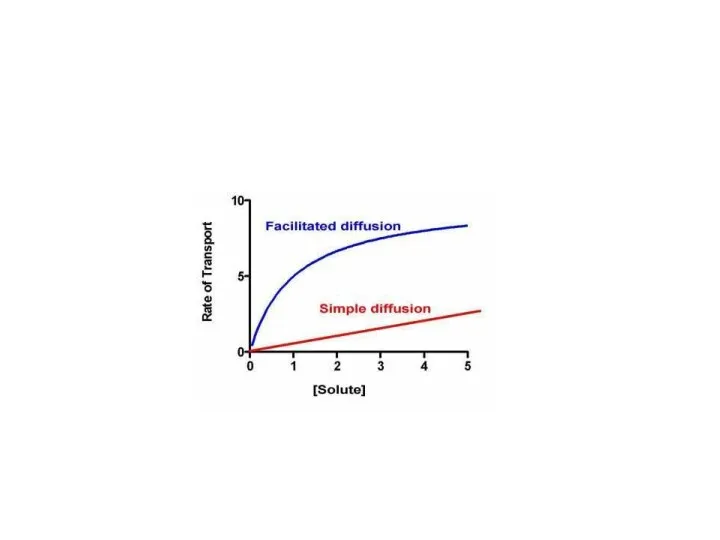

Кинетику облегченной диффузии отображает уравнение Михаэлиса-Ментен:
KM – константа Михаэлиса (равна концентрации
Кинетику облегченной диффузии отображает уравнение Михаэлиса-Ментен:
KM – константа Михаэлиса (равна концентрации

6. Электродиффузия
Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационного
6. Электродиффузия
Электродиффузия - диффузия электрически заряженных частиц (ионов) под влиянием концентрационного

Движущей силой диффузии является не только разность концентрации ионов внутри и
Движущей силой диффузии является не только разность концентрации ионов внутри и

Электрохимический потенциал определяет свободную энергию иона и учитывает все силы, способные
Электрохимический потенциал определяет свободную энергию иона и учитывает все силы, способные

Здесь: μ0- стандартный химический потенциал, который зависит от химической природы вещества
Здесь: μ0- стандартный химический потенциал, который зависит от химической природы вещества

Зависимость плотности потока ионов J от электрохимического градиента определяется уравнением Теорелла:
где
Зависимость плотности потока ионов J от электрохимического градиента определяется уравнением Теорелла:
где

Подставляя выражение для электрохимического потенциала в уравнение Теорелла, можно получить уравнение
Подставляя выражение для электрохимического потенциала в уравнение Теорелла, можно получить уравнение

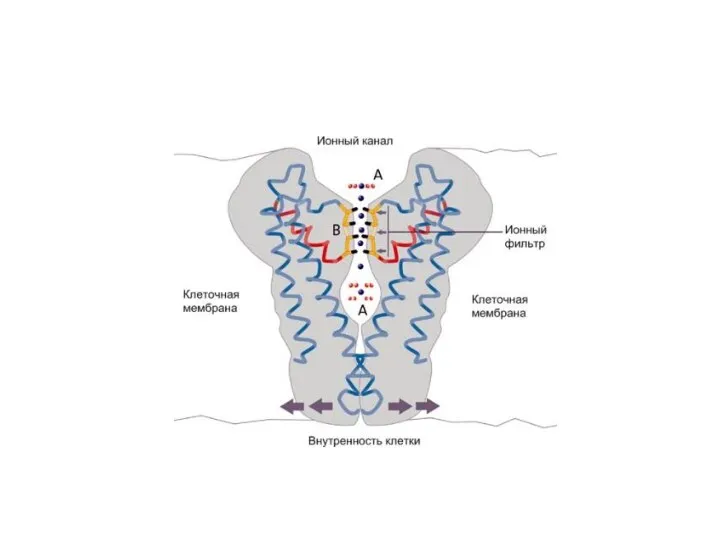
Ионные каналы мембраны представляют собой интегральные белки мембраны, которые образуют отверстия
Ионные каналы мембраны представляют собой интегральные белки мембраны, которые образуют отверстия

Существуют натриевые, калиевые, кальциевые и хлорные каналы.
Каждый из них имеет
Каждый из них имеет

Проницаемость ионных каналов может изменяться благодаря наличию ворот - определенных групп
Проницаемость ионных каналов может изменяться благодаря наличию ворот - определенных групп

Механизмы регуляции положения ворот могут отличаться в различных каналах.
Некоторые из
Механизмы регуляции положения ворот могут отличаться в различных каналах.
Некоторые из

7. Активный транспорт
Существует несколько систем активного транспорта ионов в плазматической мембране
7. Активный транспорт
Существует несколько систем активного транспорта ионов в плазматической мембране

Натрий-калиевый насос существует в плазматических мембранах всех животных и растительных клеток.
Натрий-калиевый насос существует в плазматических мембранах всех животных и растительных клеток.

Натрий-калиевый насос - один из интегральных белков мембраны.
Он обладает энзимными
Натрий-калиевый насос - один из интегральных белков мембраны.
Он обладает энзимными

Молекула насоса существует в двух основных конформациях, взаимное преобразование которых стимулируется
Молекула насоса существует в двух основных конформациях, взаимное преобразование которых стимулируется

При расщеплении натрий-калиевой АТФазой молекулы ATФ неорганический фосфат присоединяется к белку.
При расщеплении натрий-калиевой АТФазой молекулы ATФ неорганический фосфат присоединяется к белку.

Затем молекула неорганического фосфата отсоединяется от насоса-белка, и насос превращается в
Затем молекула неорганического фосфата отсоединяется от насоса-белка, и насос превращается в

Один натрий-калиевый насос может перенести через мембрану 150- 600 ионов натрия
Один натрий-калиевый насос может перенести через мембрану 150- 600 ионов натрия
 Царства живой природы
Царства живой природы Работа и сила мышц. Строение сократительного аппарата поперечно-полосатой мышечной ткани
Работа и сила мышц. Строение сократительного аппарата поперечно-полосатой мышечной ткани Органы дыхания
Органы дыхания Гены и аллели. Аллельные взаимодействия
Гены и аллели. Аллельные взаимодействия Dinoflagellates
Dinoflagellates Торговий Дім АКТИВ-HARVEST. Добрива для позакореневого живлення
Торговий Дім АКТИВ-HARVEST. Добрива для позакореневого живлення Отряд Стрекозы
Отряд Стрекозы Рефлекторный принцип деятельности ЦНС. Возбуждение и торможение. Функции нейронов и нейроглии. Физиология рецепторов
Рефлекторный принцип деятельности ЦНС. Возбуждение и торможение. Функции нейронов и нейроглии. Физиология рецепторов Создание, исследование и испытание доступного биовегетария
Создание, исследование и испытание доступного биовегетария Асимметрия полушарий головного мозга человека
Асимметрия полушарий головного мозга человека Закон Менделя
Закон Менделя презентация к уроку Органические молекулы - углеводы
презентация к уроку Органические молекулы - углеводы Составление тренировочных программ для мужчин различных типов сложения
Составление тренировочных программ для мужчин различных типов сложения Репчатый лук
Репчатый лук Презентация Организация исследовательской работы
Презентация Организация исследовательской работы Презентация к уроку Эволюция строения и функций органов и их систем
Презентация к уроку Эволюция строения и функций органов и их систем Опорно-двигательная система человека
Опорно-двигательная система человека Репродукція клітин
Репродукція клітин Protein Structure and Function
Protein Structure and Function Инструменты для груминга
Инструменты для груминга klasss
klasss Подцарство одноклеточные
Подцарство одноклеточные Осьминоги. Головоногие моллюски
Осьминоги. Головоногие моллюски Особенности размножения рыб
Особенности размножения рыб Урок-игра Кровь и кровообращение
Урок-игра Кровь и кровообращение Для начинающих улитководов
Для начинающих улитководов Класс Земноводные, или Амфибии (Amphibia)
Класс Земноводные, или Амфибии (Amphibia) Вид. Критерии вида
Вид. Критерии вида