- Цитологические основы законов Менделя

Содержание
- 2. Цитологические основы законов Менделя Цитологические основы законов Менделя базируются на: парности хромосом (парности генов, обусловливающих возможность
- 3. Цитологические основы дигибридного скрещивания Основные события при дигибридном скрещивании связаны с мейозом и половым процессом. Диплоидный
- 4. Если родители отличались по аллелям двух генов, в клетке присутствует четыре разных гена. Пусть один родитель
- 5. Цитологические основы третьего закона Менделя
- 6. Рассмотрим этот процесс на примере морских свинок. Чёрная окраска — B (доминантный признак) и коричневая окраска
- 7. Дигибридное скрещивание у морских свинок
- 8. Для того чтобы понять, каким образом будут комбинироваться при скрещивании двух гибридов первого поколения все возможные
- 9. Итак, каждая клетка соответствует 1/16 общего числа потомков, и если просуммировать отношение 9:3:3:1, то получим 16.
- 10. Цитологические основы расщепления признаков при дигибридном скрещивании
- 11. Полигибридное скрещивание Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании, т. е. когда родители
- 12. Хромосомная теория наследственности Дальнейшие исследования генетиков показали, что законы Менделя о независимом наследовании признаков при дигибридном
- 13. Эксперимент Т. Моргана Объект исследования: плодовая мушка дрозофила диплоидный набор = 8 хромосом А – серое
- 14. При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой, имеющей черное тело и зачаточные
- 15. Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с
- 16. Кроссинговер Гомологичные хромосомы могут перекрещиваться (кроссинговер или прекрест) и обмениваться гомологичными участками. В этом случае гены
- 17. Кроссинговер – обмен участками меж-ду гомологичными хро-мосомами в момент их временного сближения, т.е. конъюгации А а
- 19. если гены находятся в 1 хромосоме, они наследуются преимущественно вместе, образуя группу сцепления у зигот возникают
- 20. если анализирующему скрещиванию подвергались самки, закон сцепленного наследования нарушался у зигот возникают соотношения генотипов F1 АBab×ааbb=АBab
- 21. Р Х F 1 G АаВв аавв АВ Ав аВ ав ав АаВв Аавв ааВв аавв
- 22. Результаты скрещивание дрозофил: а) Полное сцепление без кроссинговера б) С частотой кроссинговера равной 17 %
- 23. Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности: 1. Гены располагаются в хромосомах; различные хромосомы
- 24. Генетическое определение пола У мужских и женских организмов все пары хромосом, кроме одной, одинаковы – аутосомы,
- 25. Классификация хромосом организма Хромосомы Аутосомы Половые хромосомы
- 26. Схема расщепления по признаку пола у дрозофилы Гомогаметный пол Гетерогаметный пол
- 28. Хромосомное определение пола
- 29. ХРОМОСОМНОЕ ОПРЕДЕЛЕНИЕ ПОЛА. Первый тип Мужской пол гетерогаметен – 50% гамет несут Х-хромосому, 50% гамет несут
- 30. Второй тип Cамки – ХY Самцы - ХХ Женский тип - гетерогаметен 50% гамет несут Х-хромосому,
- 31. Третий тип Мужской пол гетерогаметен 50% гамет несут Х-хромосому 50% гамет не имеют половой хромосомы Например,
- 32. Четвертый тип Самцы – ХХ Самки - Х0 Женский пол гетерогаметен 50% гамет несут Х-хромосому 50%
- 33. Гаплодиплоидный тип определения пола Широко распространен у пчел и муравьев. У этих организмов нет половых хромосом:
- 34. Сцепленное наследование Это явление заметил американский ученый Морган. Гены, находящиеся в половых хромосомах, называются сцепленными с
- 35. У человека Y-хромосома передается от отца к сыновьям, а Х-хромосома – к дочерям. Вторую хромосому дети
- 36. Сцепленное наследование У человека около 60 генов наследуются в связи с Х-хромосомой: Гемофилия Дальтонизм (цветовая слепота)
- 39. Скачать презентацию

Цитологические основы законов Менделя
Цитологические основы законов Менделя базируются на:
парности хромосом
Цитологические основы законов Менделя
Цитологические основы законов Менделя базируются на:
парности хромосом

Цитологические основы дигибридного скрещивания
Основные события при дигибридном скрещивании связаны
Цитологические основы дигибридного скрещивания
Основные события при дигибридном скрещивании связаны

Если родители отличались по аллелям двух генов, в клетке присутствует четыре
Если родители отличались по аллелям двух генов, в клетке присутствует четыре
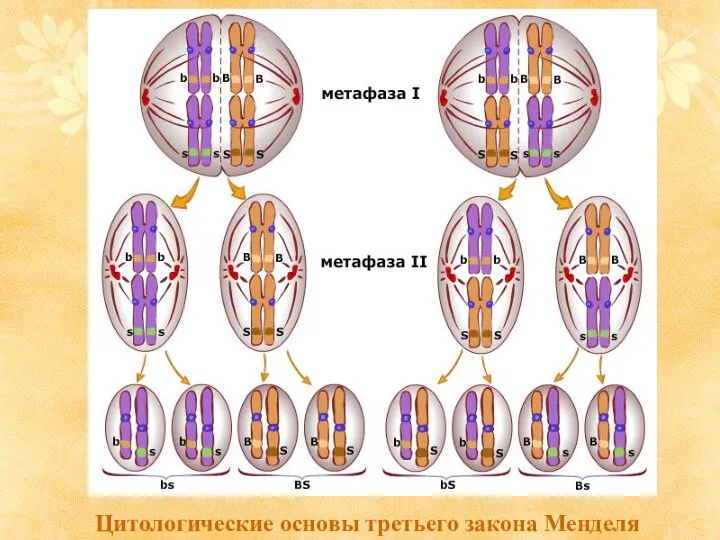
Цитологические основы третьего закона Менделя
Цитологические основы третьего закона Менделя

Рассмотрим этот процесс на примере морских свинок. Чёрная окраска — B
Рассмотрим этот процесс на примере морских свинок. Чёрная окраска — B
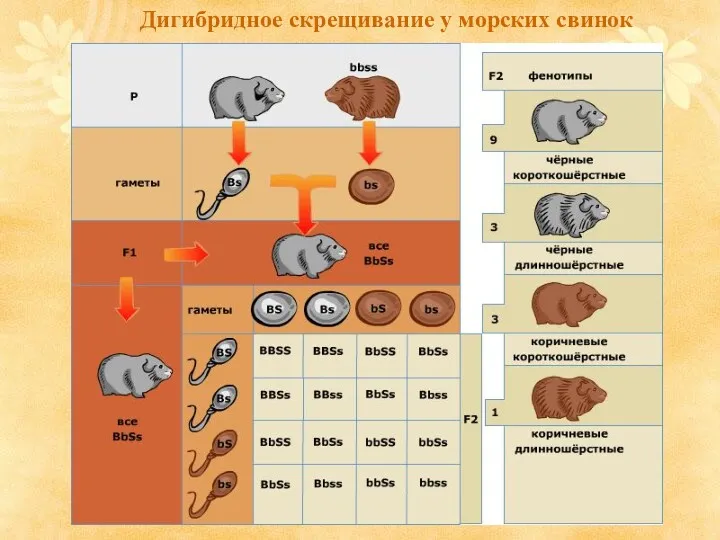
Дигибридное скрещивание у морских свинок
Дигибридное скрещивание у морских свинок

Для того чтобы понять, каким образом будут комбинироваться при скрещивании двух
Для того чтобы понять, каким образом будут комбинироваться при скрещивании двух

Итак, каждая клетка соответствует 1/16 общего числа потомков, и если
Итак, каждая клетка соответствует 1/16 общего числа потомков, и если
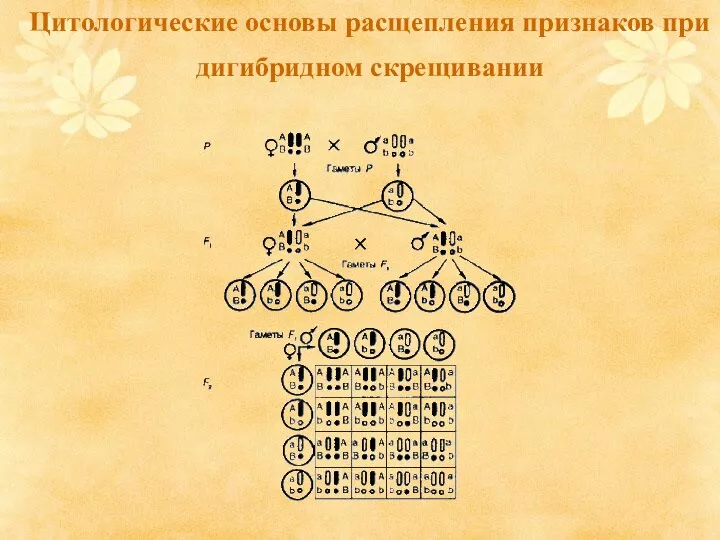
Цитологические основы расщепления признаков при дигибридном скрещивании
Цитологические основы расщепления признаков при дигибридном скрещивании

Полигибридное скрещивание
Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании,
Полигибридное скрещивание
Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании,

Хромосомная теория наследственности
Дальнейшие исследования генетиков показали, что законы Менделя о независимом
Хромосомная теория наследственности
Дальнейшие исследования генетиков показали, что законы Менделя о независимом
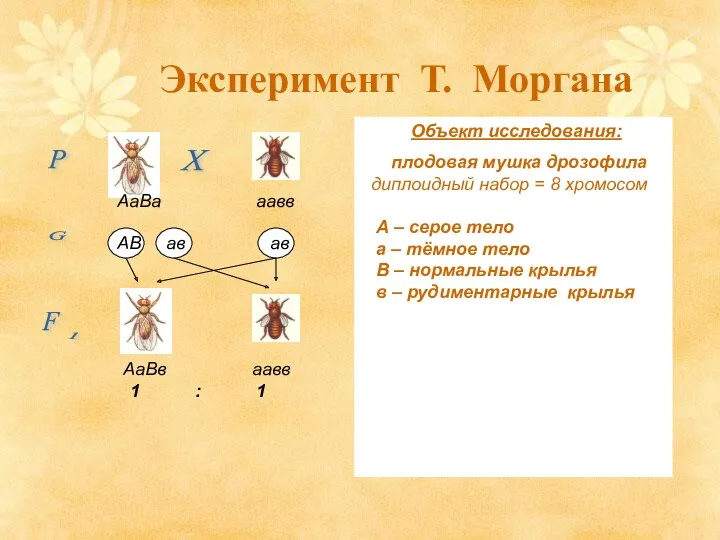
Эксперимент Т. Моргана
Объект исследования:
плодовая мушка дрозофила
Эксперимент Т. Моргана
Объект исследования:
плодовая мушка дрозофила

При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой,
При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой,

Полное сцепление наблюдается в том случае, если скрещиваются серый самец с
Полное сцепление наблюдается в том случае, если скрещиваются серый самец с

Кроссинговер
Гомологичные хромосомы могут перекрещиваться (кроссинговер или прекрест) и обмениваться гомологичными участками.
Кроссинговер
Гомологичные хромосомы могут перекрещиваться (кроссинговер или прекрест) и обмениваться гомологичными участками.
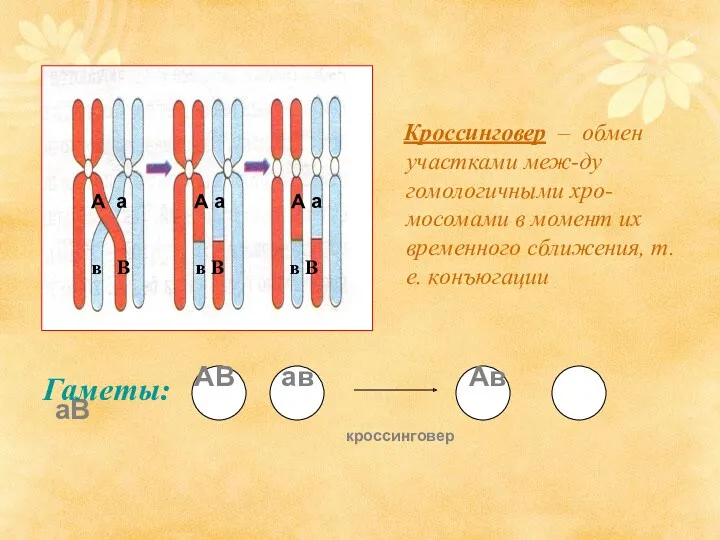
Кроссинговер – обмен участками меж-ду гомологичными хро-мосомами в момент их
Кроссинговер – обмен участками меж-ду гомологичными хро-мосомами в момент их

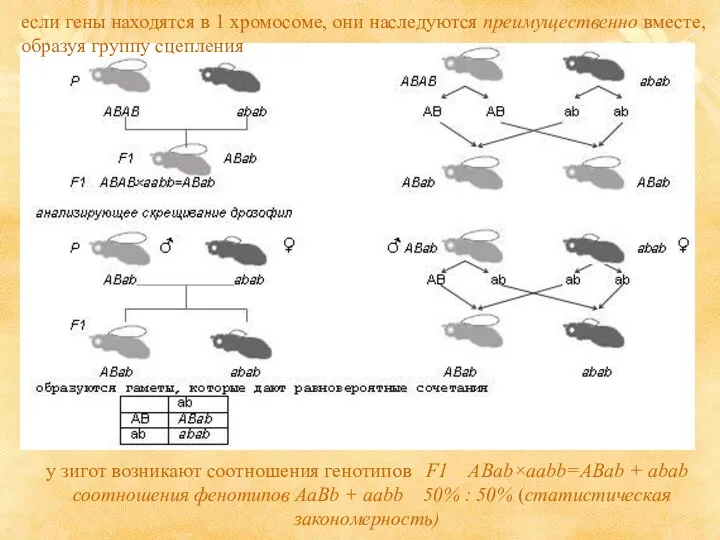
если гены находятся в 1 хромосоме, они наследуются преимущественно вместе, образуя
если гены находятся в 1 хромосоме, они наследуются преимущественно вместе, образуя
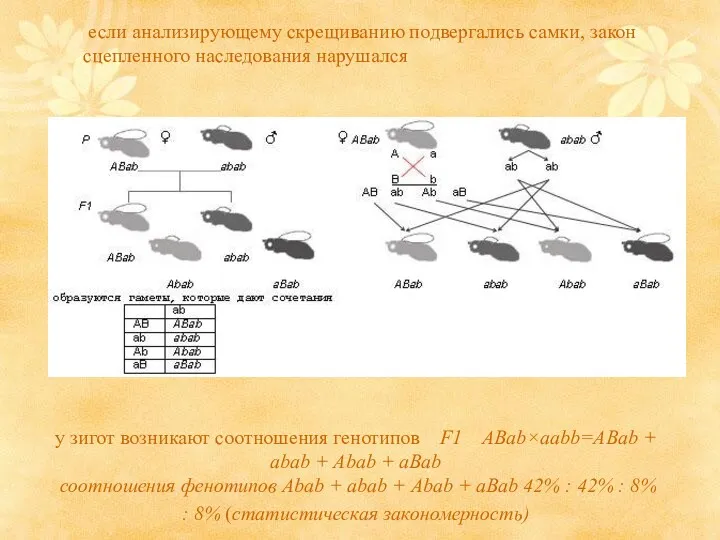
если анализирующему скрещиванию подвергались самки, закон сцепленного наследования нарушался
у зигот
если анализирующему скрещиванию подвергались самки, закон сцепленного наследования нарушался
у зигот

Р
Х
F
1
G
АаВв аавв
АВ Ав аВ
Р
Х
F
1
G
АаВв аавв
АВ Ав аВ

Результаты скрещивание дрозофил:
а) Полное сцепление без кроссинговера
б) С частотой кроссинговера равной
Результаты скрещивание дрозофил: а) Полное сцепление без кроссинговера б) С частотой кроссинговера равной

Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности:
1. Гены располагаются
Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности:
1. Гены располагаются

Генетическое определение пола
У мужских и женских организмов все пары хромосом,
Генетическое определение пола
У мужских и женских организмов все пары хромосом,
Классификация хромосом организма
Хромосомы
Аутосомы
Половые
хромосомы
Классификация хромосом организма
Хромосомы
Аутосомы
Половые
хромосомы
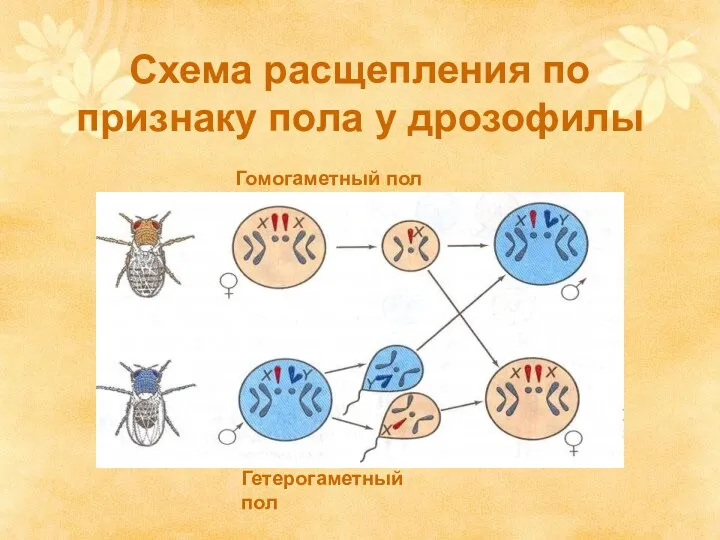
Схема расщепления по признаку пола у дрозофилы
Гомогаметный пол
Гетерогаметный пол
Схема расщепления по признаку пола у дрозофилы
Гомогаметный пол
Гетерогаметный пол

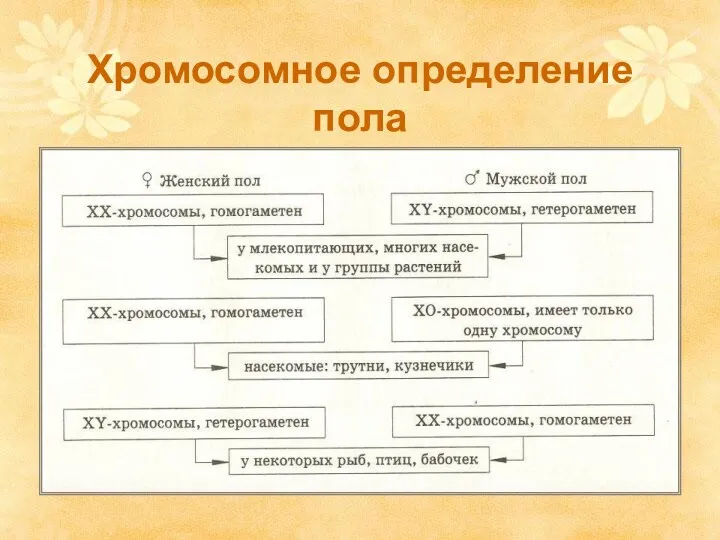
Хромосомное определение пола
Хромосомное определение пола

ХРОМОСОМНОЕ ОПРЕДЕЛЕНИЕ ПОЛА.
Первый тип
Мужской пол гетерогаметен –
50% гамет
ХРОМОСОМНОЕ ОПРЕДЕЛЕНИЕ ПОЛА.
Первый тип
Мужской пол гетерогаметен –
50% гамет

Второй тип
Cамки – ХY
Самцы - ХХ
Женский тип - гетерогаметен
50%
Второй тип
Cамки – ХY
Самцы - ХХ
Женский тип - гетерогаметен
50%

Третий тип
Мужской пол гетерогаметен
50% гамет несут
Х-хромосому
50% гамет
Третий тип
Мужской пол гетерогаметен
50% гамет несут
Х-хромосому
50% гамет

Четвертый тип
Самцы – ХХ
Самки - Х0
Женский пол гетерогаметен
50% гамет несут
Четвертый тип
Самцы – ХХ
Самки - Х0
Женский пол гетерогаметен
50% гамет несут

Гаплодиплоидный тип определения пола
Широко распространен у пчел и муравьев.
У
Гаплодиплоидный тип определения пола
Широко распространен у пчел и муравьев.
У

Сцепленное наследование
Это явление заметил американский ученый Морган. Гены, находящиеся в
Сцепленное наследование
Это явление заметил американский ученый Морган. Гены, находящиеся в

У человека Y-хромосома передается от отца к сыновьям, а Х-хромосома
У человека Y-хромосома передается от отца к сыновьям, а Х-хромосома

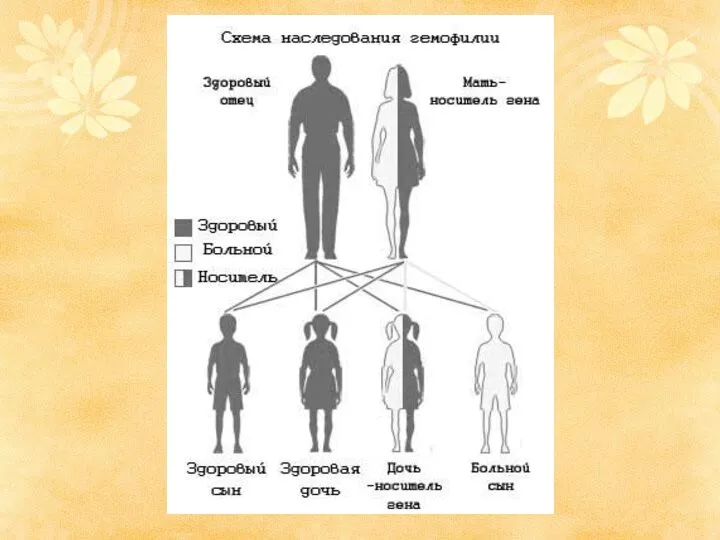
Сцепленное наследование
У человека около 60 генов наследуются в связи с Х-хромосомой:
Сцепленное наследование
У человека около 60 генов наследуются в связи с Х-хромосомой:
 Поле и его обитатели
Поле и его обитатели Биотические факторы среды
Биотические факторы среды Лесные дары. Ягоды
Лесные дары. Ягоды Опорно-двигательная система человека
Опорно-двигательная система человека Костная и мышечная системы. Скелет головы
Костная и мышечная системы. Скелет головы Антропогенез
Антропогенез Ботаника
Ботаника Кальцій у організмі людини
Кальцій у організмі людини Энергетический обмен. Метаболизм (обмен веществ)
Энергетический обмен. Метаболизм (обмен веществ) Фиксация молекулярного азота (азотфиксация, диазотрофия) микроорганизмами
Фиксация молекулярного азота (азотфиксация, диазотрофия) микроорганизмами Семенные растения
Семенные растения Структурные компоненты клетки и их физиологические функции
Структурные компоненты клетки и их физиологические функции Технология выращивания комнатных растений
Технология выращивания комнатных растений Орган зрения
Орган зрения Размножение и развитие животных. 3 класс
Размножение и развитие животных. 3 класс Биологические полимеры - нуклеиновые кислоты
Биологические полимеры - нуклеиновые кислоты Экскурсия по Лапландскому заповеднику
Экскурсия по Лапландскому заповеднику Презентация к внеклассному мероприятию Муравьи- общественные насекомые
Презентация к внеклассному мероприятию Муравьи- общественные насекомые презентация к уроку по теме: Цветок. Строение и значение цветка
презентация к уроку по теме: Цветок. Строение и значение цветка Задачи по генетике
Задачи по генетике Биотические факторы
Биотические факторы Слуховая сенсорная система
Слуховая сенсорная система Приспособленность организмов к определенной среде обитания
Приспособленность организмов к определенной среде обитания Генетична рівновага в популяціях і її порушення
Генетична рівновага в популяціях і її порушення Государственная итоговая аттестация обучающихся за основное общее образование по биологии в 2023 году
Государственная итоговая аттестация обучающихся за основное общее образование по биологии в 2023 году Лекарственные и съедобные растения, произрастающие в Якутии
Лекарственные и съедобные растения, произрастающие в Якутии Питание и пищеварение
Питание и пищеварение Красная книга растений
Красная книга растений