- Возникновение жизненного цикла и онтогенеза metazoa

Содержание
- 2. Простейший онтогенез на клеточном уровне – метазойный клеточный цикл. Он включает в себя морфогенетический цикл –
- 3. Первое деление дробления яиц морского ежа: морфогенетический цикл роста-сокращения контактной зоны
- 4. Эволюция онтогенеза Protozoa связана с полимеризацией субклеточных органелл (энергид) и переходом к клональному самовоспроизведению При протозойном
- 5. Инфузории (Tetrahymena) – вершина эволюции онтогенеза полимерных Protozoa. Реснички становятся морфогенетическими аналогами клеток Metazoa, митоз –
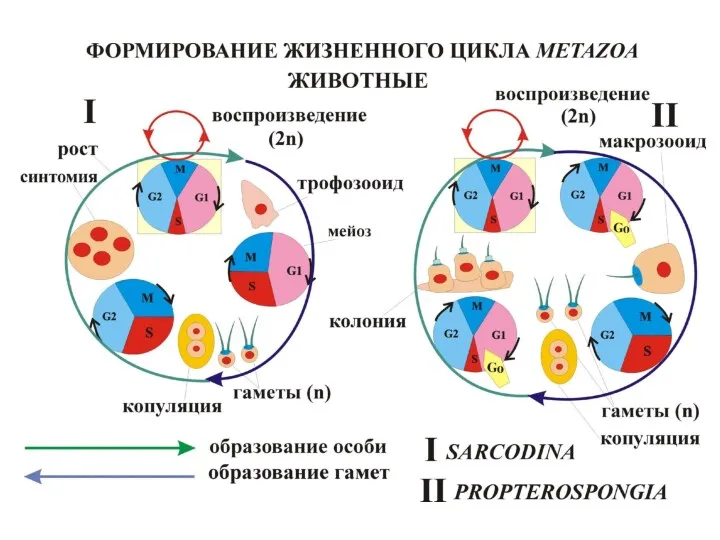
- 6. Варианты (индивидуальная изменчивость) чередования фаз жизненного цикла раковинных амеб Foraminifera: * - палинтомия, ** - мейоз,
- 8. НЕЗАВИСИМОСТЬ ЭВОЛЮЦИИ ОТ ФИЛОГЕНЕЗА У растений и социальных амеб многоклеточность возникает иначе, чем у животных, но
- 9. * - клетки в автоколебательном режиме секреции АМФ (гликоген почти весь израсходован) ** - клетки в
- 10. I – постоянная и слабая секреция цАМФ, II – режим автоколебаний, III – угасание секреции. C
- 11. Изменчивы число и размер агрегатов, и соотношение преспоровой и престеблевой части. Отбор в пользу более длинного
- 12. ВОЗНИКНОВЕНИЕ ОНТОГЕНЕЗА (ЖИЗНЕННОГО ЦИКЛА) METAZOA A – исходное чередование палинтомического (N) и монотомического (N+1) поколений Protozoa:
- 13. ЭВОЛЮЦИОННАЯ УСТОЙЧИВОСТЬ ОНТОГЕНЕЗА METAZOA: память об эволюционном происхождении сохраняется, несмотря на дивергенцию Двойная стрелка – монотомия,
- 14. ОНТОГЕНЕЗ, В ОТЛИЧИЕ ОТ МОРФОГЕНЕЗА, МОЖЕТ БЫТЬ (ИЛИ НЕ БЫТЬ) ПРЕДМЕТОМ ОТБОРА Светлая заливка – бластодерма,
- 15. Красные наконечники – «источник», синие стрелки – «сток» поверхности клетки, пунктирные стрелки – латеральный поток поверхности
- 16. Возникновение первичного эпителиального пласта (ЭП) Одиночные клетки движутся по недеформируемому субстрату (например, по стеклу) против градиента
- 17. Морфогенез ЭП возникает на основе его репарации – «заживления раны», минимизируя суммарную площадь свободной поверхности ЭП
- 18. Заживление ЭП подразумевает инверсию полярности выселяющихся из ЭП клеток. С этого шага начинается эпителио-мезенхимальный цикл (EMC)
- 19. Бластопор появляется из-за репарации места замыкания бластопора с неизбежной инверсией полярности клеток, выселяющихся из первичного ЭП.
- 20. ЭВОЛЮЦИЯ БЛАСТОПОРА НАЧИНАЕТСЯ С ЕГО ЗАМЫКАНИЯ. Движение клеток, заживляющее бластопор как рану в ЭП – типичный
- 21. КТ является зоной максимального перепада скорости движения клеток вдоль ЭП и давления внутри ЭП (теорема Бернулли).
- 22. Разделение стоков и источников поверхности КТ повторяет дифференцировку полюсов первичного ЭП с поправкой на новую топологию
- 23. В эволюции реализуются все «неконкурентные» варианты выделения стоков и источников поверхности КТ Один из секторов КТ
- 24. Эпиболия дорсального края бластопора при относительной неподвижности вентрального края перемещает бластопор на вентральную сторону, а «следом»
- 25. В пределе, при установлении связи движения мезодермы с формой яйца, получается щелевидный бластопор Ecdysozoa
- 26. КТ СТАНОВИТСЯ «ЦЕНТРОМ ИЗМЕНЧИВОСТИ» ИЗ-ЗА ОБРАТНОЙ СВЯЗИ ДВИЖЕНИЯ КЛЕТОК С ИЗМЕНЕНИЕМ ФОРМЫ КТ R1-R3 – радиусы
- 27. У морского ежа поток клеток через КТ стационарен в течение всей гаструляции Форма и размер зоны
- 28. Нестационарность потока клеток через КТ проявляется в виде осцилляций: например, при образовании ЗЩ у вьюна (серия
- 29. Гаструляция у ланцетника Ранняя гаструла ланцетника выглядит как изолированный ЭП, заворачивающийся внутрь для заживления «дыры» в
- 30. От ланцетника к анамниям прослеживается постепенное «слияние» AP оси c DV осью Карта зачатков у ланцетника
- 31. ЭВОЛЮЦИЯ БЛАСТОПОРА ХОРДОВЫХ: A – ланцетник, B – минога, C – амфибии и костистые рыбы, 1-3
- 32. Амниоты продолжают ретроградную эволюцию онтогенеза анамний. Изменчивость морфогенеза превращается в последовательность стадий онтогенеза. С появлением зародышевой
- 33. РЕТРОГРАДНАЯ ЭВОЛЮЦИЯ СИГНАЛЬНЫХ ПУТЕЙ В ЭМБРИОГНЕЗЕ ХОРДОВЫХ У Xenopus и Zebrafish β-катенин определяет место ДГБ У
- 35. Скачать презентацию
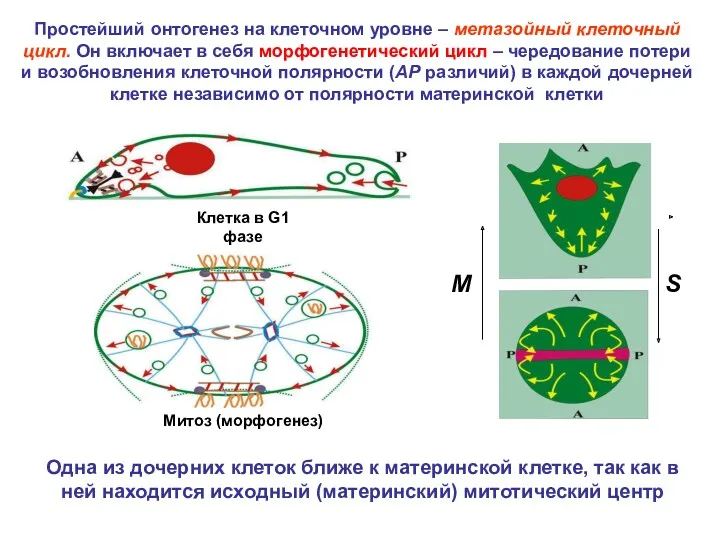
Простейший онтогенез на клеточном уровне – метазойный клеточный цикл. Он включает
Простейший онтогенез на клеточном уровне – метазойный клеточный цикл. Он включает

Первое деление дробления яиц морского ежа: морфогенетический цикл роста-сокращения контактной зоны
Первое деление дробления яиц морского ежа: морфогенетический цикл роста-сокращения контактной зоны
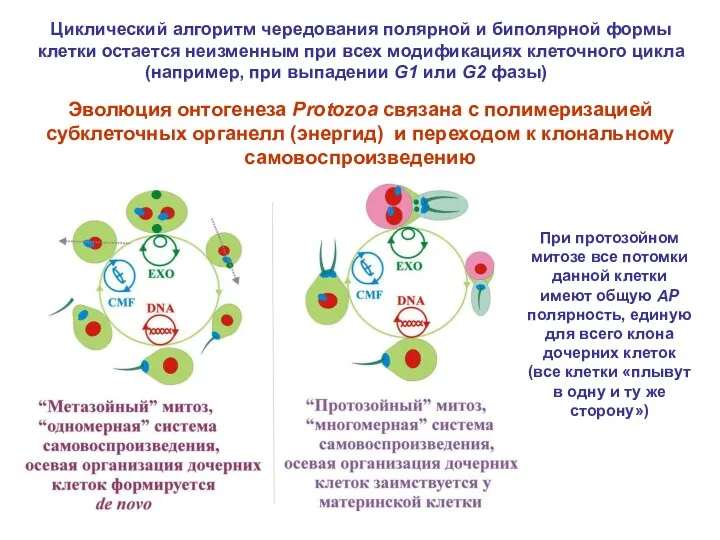
Эволюция онтогенеза Protozoa связана с полимеризацией субклеточных органелл (энергид) и переходом
Эволюция онтогенеза Protozoa связана с полимеризацией субклеточных органелл (энергид) и переходом
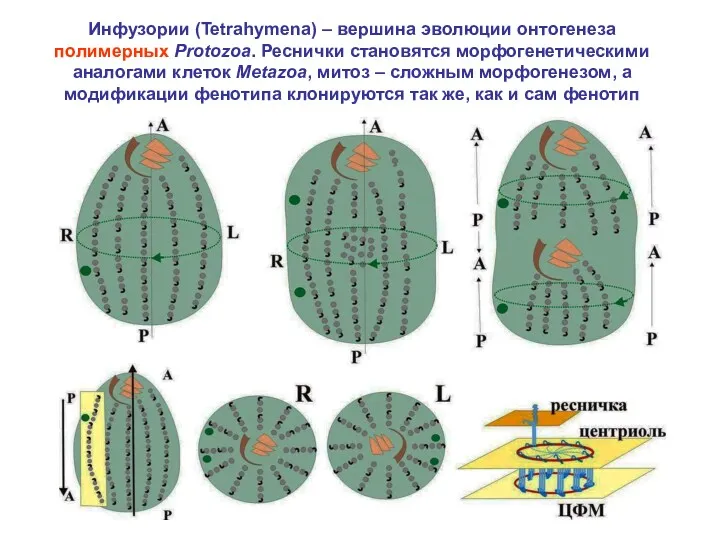
Инфузории (Tetrahymena) – вершина эволюции онтогенеза полимерных Protozoa. Реснички становятся морфогенетическими
Инфузории (Tetrahymena) – вершина эволюции онтогенеза полимерных Protozoa. Реснички становятся морфогенетическими
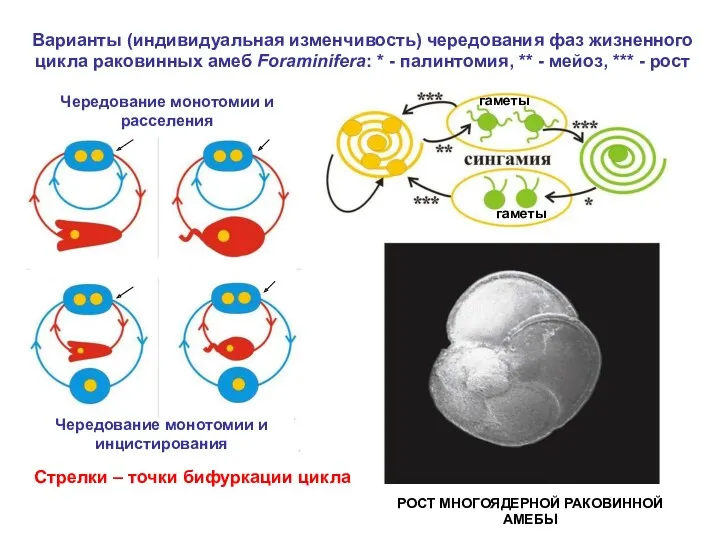
Варианты (индивидуальная изменчивость) чередования фаз жизненного цикла раковинных амеб Foraminifera: *
Варианты (индивидуальная изменчивость) чередования фаз жизненного цикла раковинных амеб Foraminifera: *
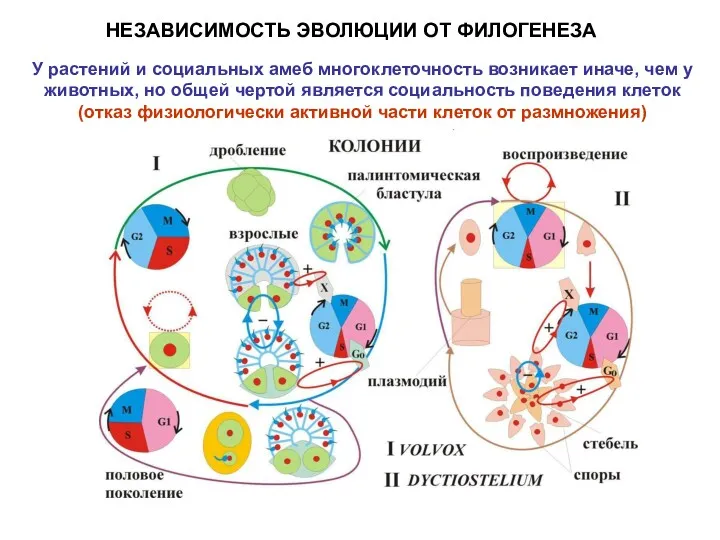
НЕЗАВИСИМОСТЬ ЭВОЛЮЦИИ ОТ ФИЛОГЕНЕЗА
У растений и социальных амеб многоклеточность возникает иначе,
НЕЗАВИСИМОСТЬ ЭВОЛЮЦИИ ОТ ФИЛОГЕНЕЗА
У растений и социальных амеб многоклеточность возникает иначе,
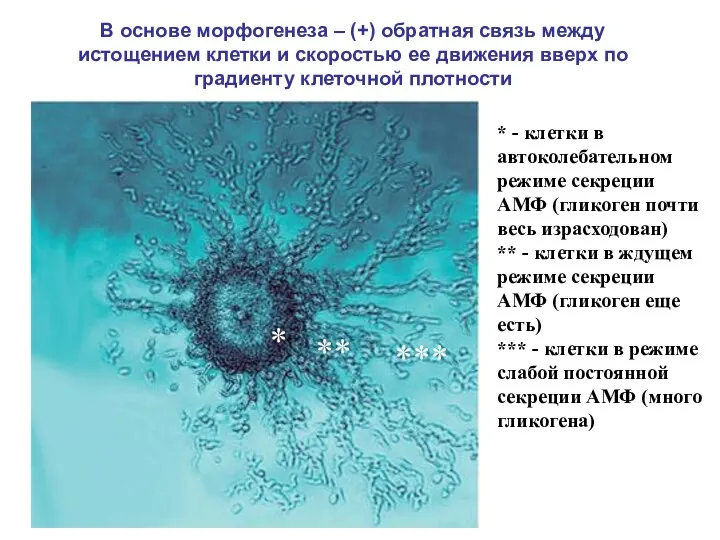
* - клетки в автоколебательном режиме секреции АМФ (гликоген почти весь
* - клетки в автоколебательном режиме секреции АМФ (гликоген почти весь
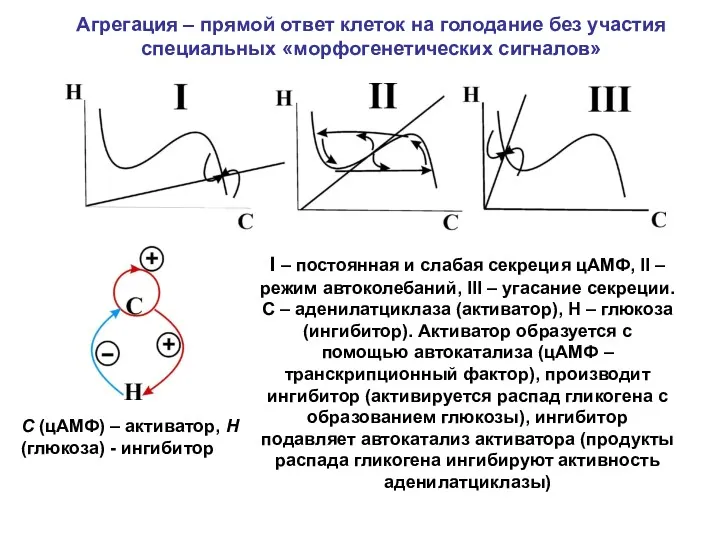
I – постоянная и слабая секреция цАМФ, II – режим автоколебаний,
I – постоянная и слабая секреция цАМФ, II – режим автоколебаний,
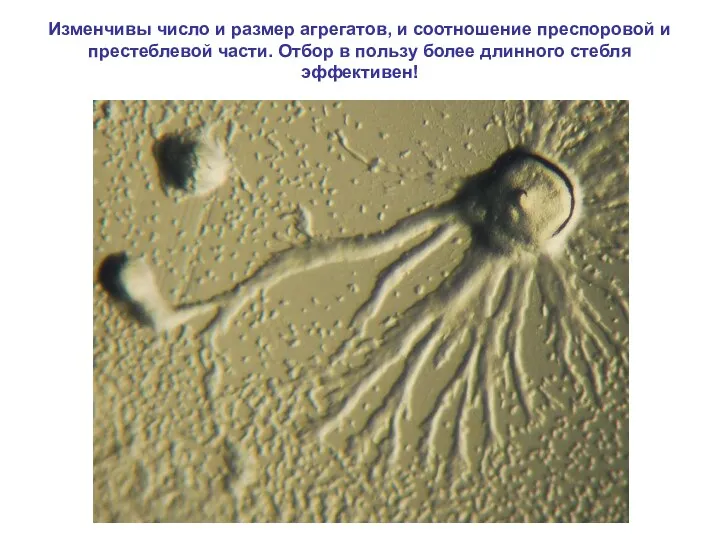
Изменчивы число и размер агрегатов, и соотношение преспоровой и престеблевой части.
Изменчивы число и размер агрегатов, и соотношение преспоровой и престеблевой части.
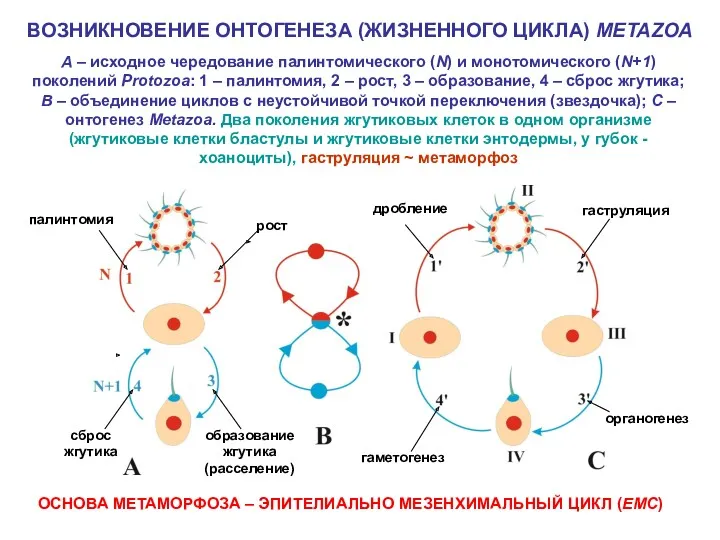
ВОЗНИКНОВЕНИЕ ОНТОГЕНЕЗА (ЖИЗНЕННОГО ЦИКЛА) METAZOA
A – исходное чередование палинтомического (N) и
ВОЗНИКНОВЕНИЕ ОНТОГЕНЕЗА (ЖИЗНЕННОГО ЦИКЛА) METAZOA
A – исходное чередование палинтомического (N) и
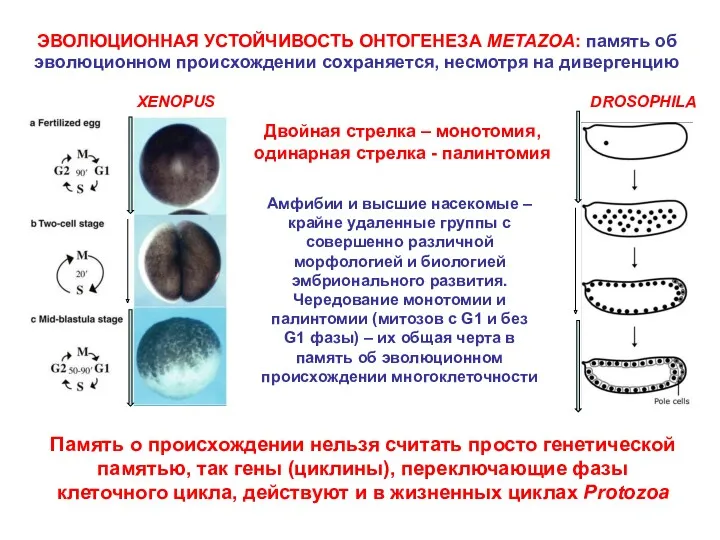
ЭВОЛЮЦИОННАЯ УСТОЙЧИВОСТЬ ОНТОГЕНЕЗА METAZOA: память об эволюционном происхождении сохраняется, несмотря на
ЭВОЛЮЦИОННАЯ УСТОЙЧИВОСТЬ ОНТОГЕНЕЗА METAZOA: память об эволюционном происхождении сохраняется, несмотря на
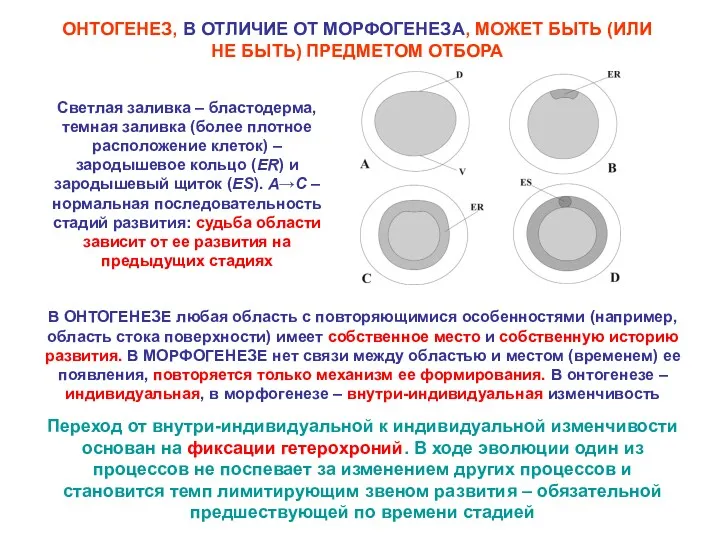
ОНТОГЕНЕЗ, В ОТЛИЧИЕ ОТ МОРФОГЕНЕЗА, МОЖЕТ БЫТЬ (ИЛИ НЕ БЫТЬ) ПРЕДМЕТОМ
ОНТОГЕНЕЗ, В ОТЛИЧИЕ ОТ МОРФОГЕНЕЗА, МОЖЕТ БЫТЬ (ИЛИ НЕ БЫТЬ) ПРЕДМЕТОМ

Красные наконечники – «источник», синие стрелки – «сток» поверхности клетки, пунктирные
Красные наконечники – «источник», синие стрелки – «сток» поверхности клетки, пунктирные
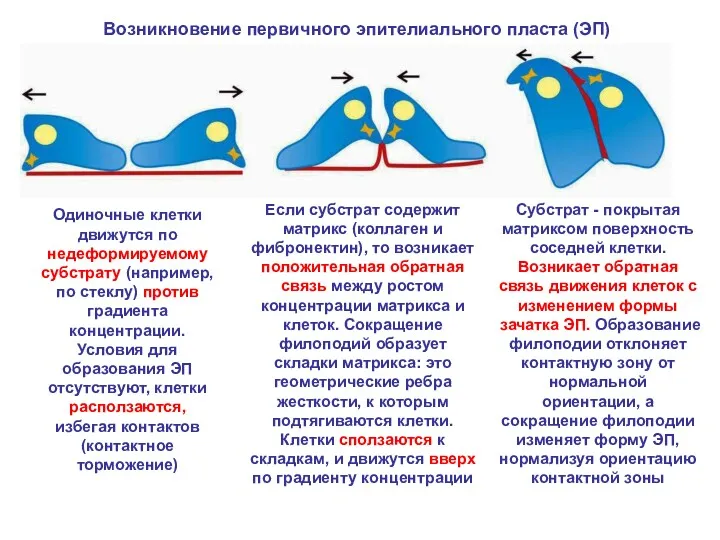
Возникновение первичного эпителиального пласта (ЭП)
Одиночные клетки движутся по недеформируемому субстрату (например,
Возникновение первичного эпителиального пласта (ЭП)
Одиночные клетки движутся по недеформируемому субстрату (например,
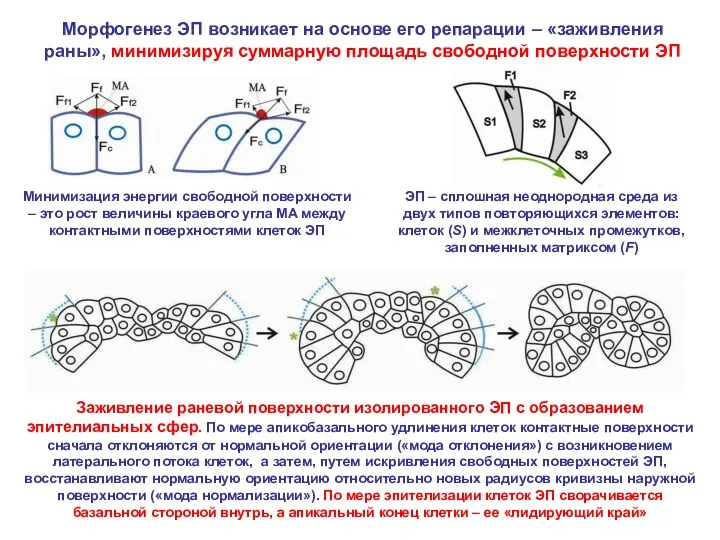
Морфогенез ЭП возникает на основе его репарации – «заживления раны», минимизируя
Морфогенез ЭП возникает на основе его репарации – «заживления раны», минимизируя
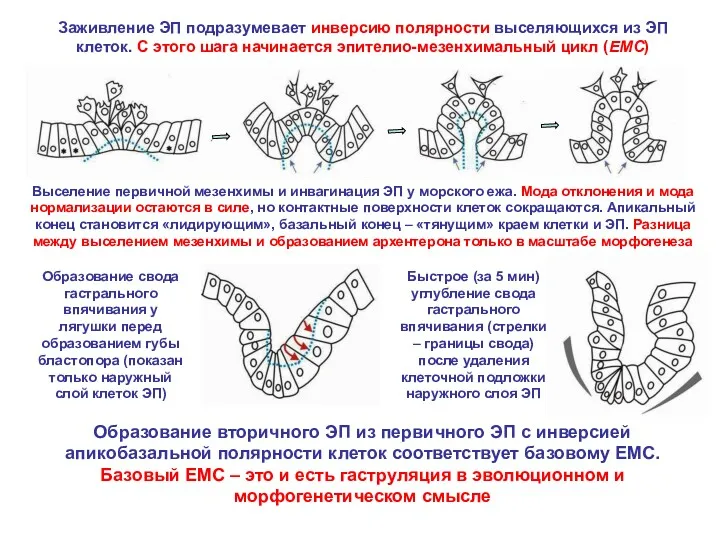
Заживление ЭП подразумевает инверсию полярности выселяющихся из ЭП клеток. С этого
Заживление ЭП подразумевает инверсию полярности выселяющихся из ЭП клеток. С этого

Бластопор появляется из-за репарации места замыкания бластопора с неизбежной инверсией полярности
Бластопор появляется из-за репарации места замыкания бластопора с неизбежной инверсией полярности
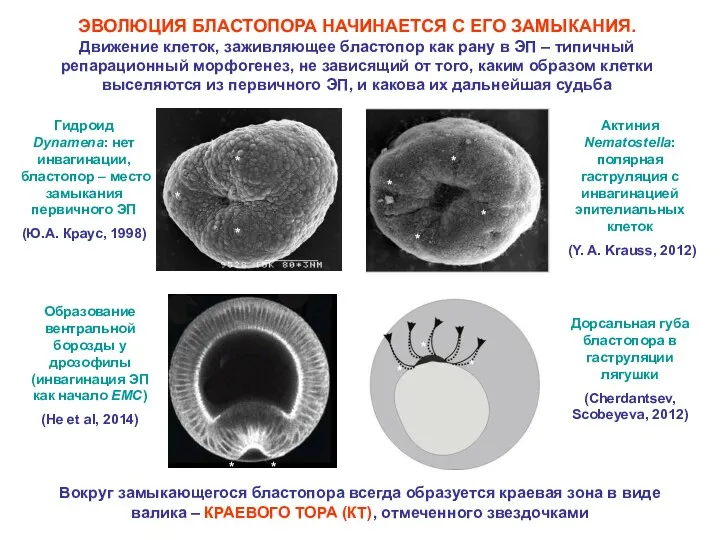
ЭВОЛЮЦИЯ БЛАСТОПОРА НАЧИНАЕТСЯ С ЕГО ЗАМЫКАНИЯ. Движение клеток, заживляющее бластопор как
ЭВОЛЮЦИЯ БЛАСТОПОРА НАЧИНАЕТСЯ С ЕГО ЗАМЫКАНИЯ. Движение клеток, заживляющее бластопор как
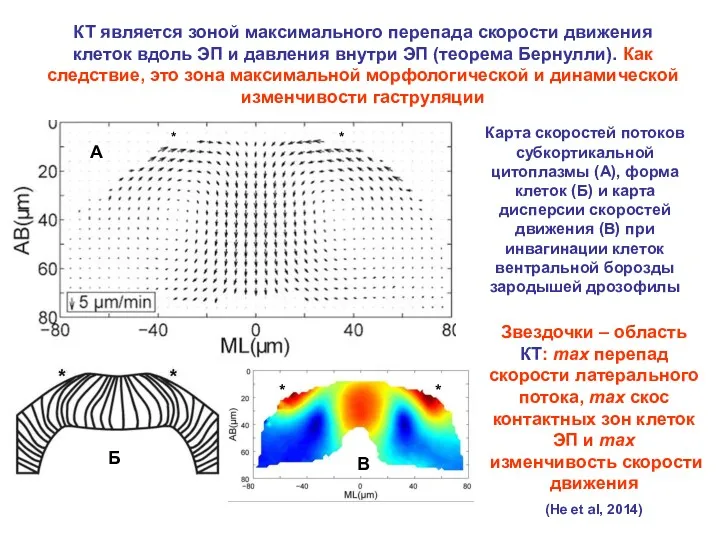
КТ является зоной максимального перепада скорости движения клеток вдоль ЭП и
КТ является зоной максимального перепада скорости движения клеток вдоль ЭП и

Разделение стоков и источников поверхности КТ повторяет дифференцировку полюсов первичного ЭП
Разделение стоков и источников поверхности КТ повторяет дифференцировку полюсов первичного ЭП
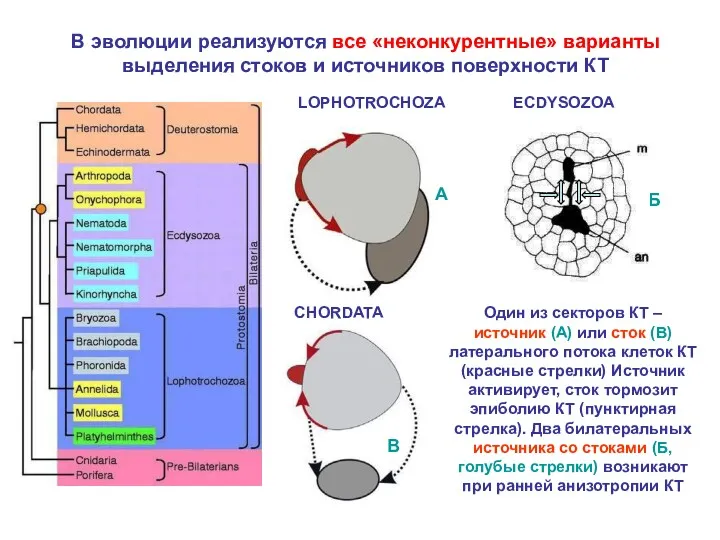
В эволюции реализуются все «неконкурентные» варианты выделения стоков и источников поверхности
В эволюции реализуются все «неконкурентные» варианты выделения стоков и источников поверхности
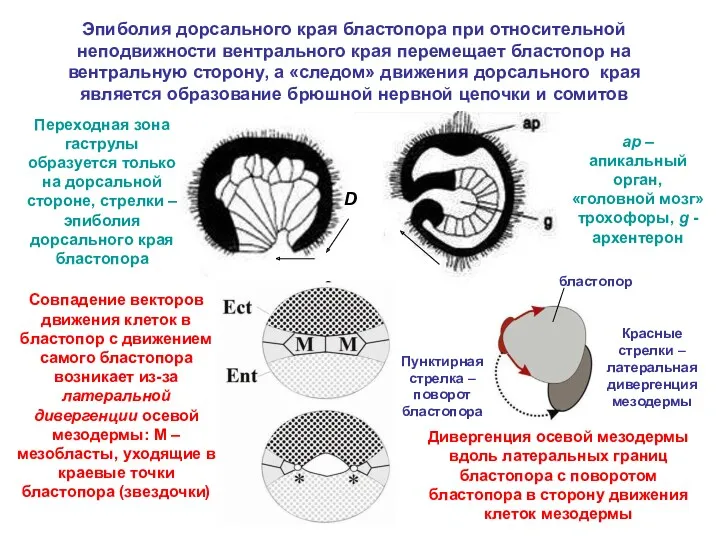
Эпиболия дорсального края бластопора при относительной неподвижности вентрального края перемещает бластопор
Эпиболия дорсального края бластопора при относительной неподвижности вентрального края перемещает бластопор
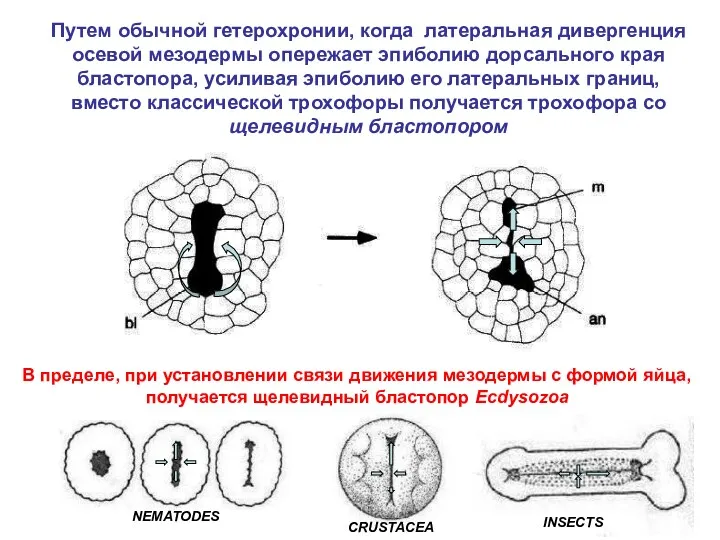
В пределе, при установлении связи движения мезодермы с формой яйца, получается
В пределе, при установлении связи движения мезодермы с формой яйца, получается
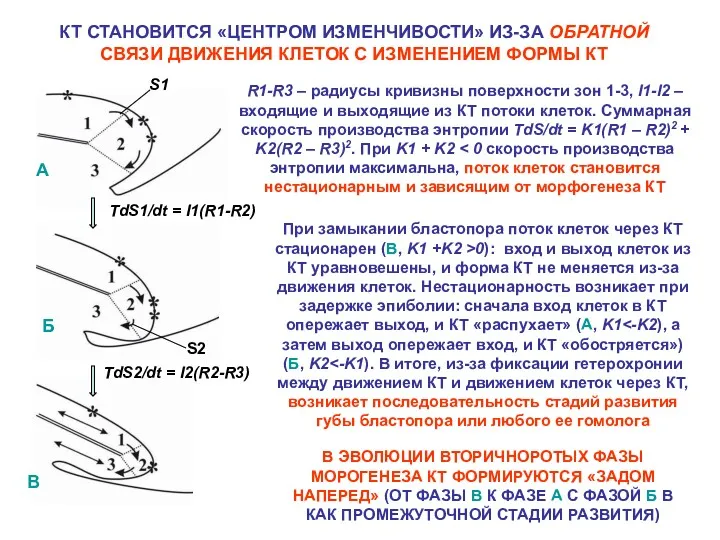
КТ СТАНОВИТСЯ «ЦЕНТРОМ ИЗМЕНЧИВОСТИ» ИЗ-ЗА ОБРАТНОЙ СВЯЗИ ДВИЖЕНИЯ КЛЕТОК С ИЗМЕНЕНИЕМ
КТ СТАНОВИТСЯ «ЦЕНТРОМ ИЗМЕНЧИВОСТИ» ИЗ-ЗА ОБРАТНОЙ СВЯЗИ ДВИЖЕНИЯ КЛЕТОК С ИЗМЕНЕНИЕМ
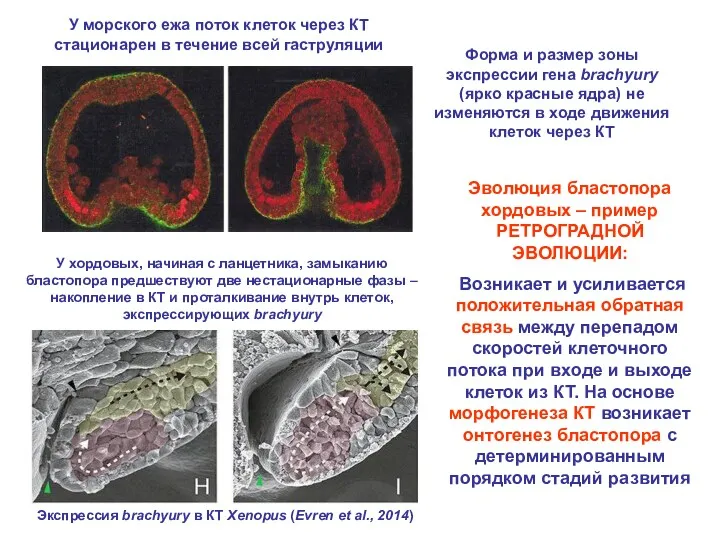
У морского ежа поток клеток через КТ стационарен в течение всей
У морского ежа поток клеток через КТ стационарен в течение всей
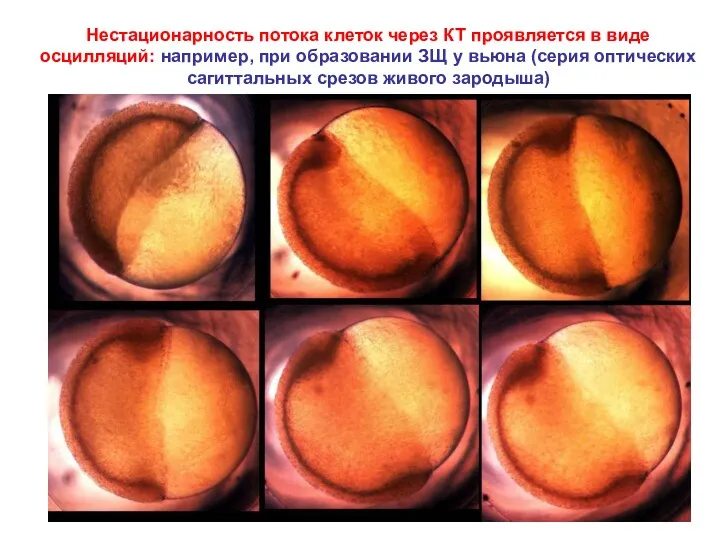
Нестационарность потока клеток через КТ проявляется в виде осцилляций: например, при
Нестационарность потока клеток через КТ проявляется в виде осцилляций: например, при
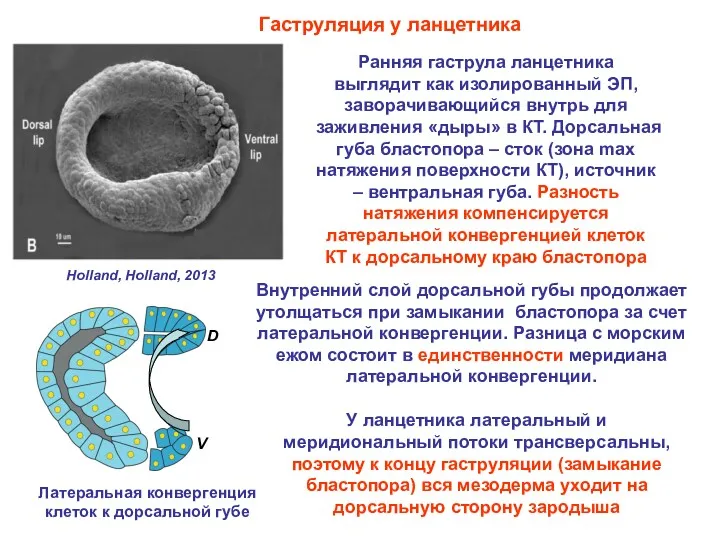
Гаструляция у ланцетника
Ранняя гаструла ланцетника выглядит как изолированный ЭП, заворачивающийся внутрь
Гаструляция у ланцетника
Ранняя гаструла ланцетника выглядит как изолированный ЭП, заворачивающийся внутрь
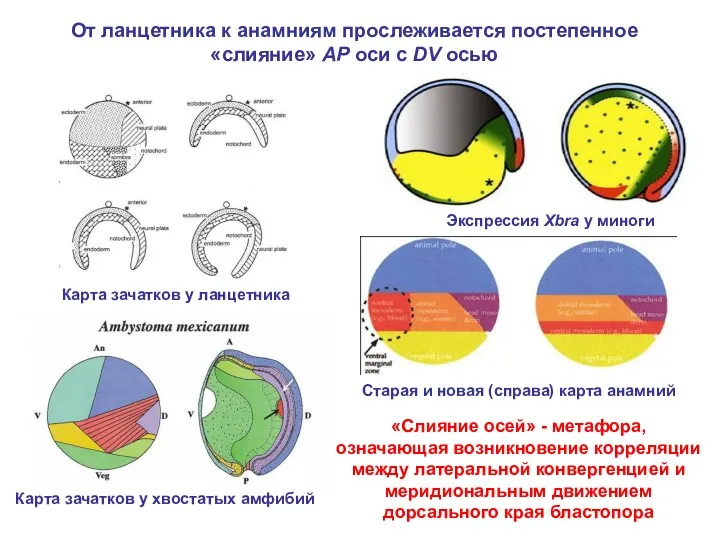
От ланцетника к анамниям прослеживается постепенное «слияние» AP оси c DV
От ланцетника к анамниям прослеживается постепенное «слияние» AP оси c DV

ЭВОЛЮЦИЯ БЛАСТОПОРА ХОРДОВЫХ: A – ланцетник, B – минога, C –
ЭВОЛЮЦИЯ БЛАСТОПОРА ХОРДОВЫХ: A – ланцетник, B – минога, C –
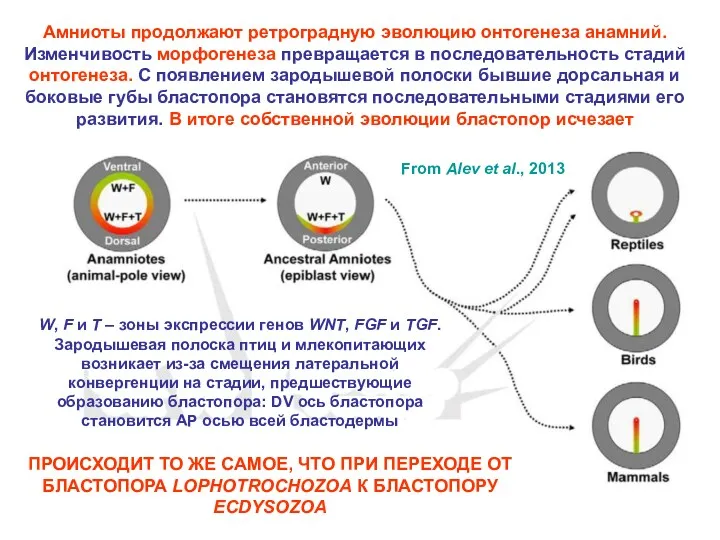
Амниоты продолжают ретроградную эволюцию онтогенеза анамний. Изменчивость морфогенеза превращается в последовательность
Амниоты продолжают ретроградную эволюцию онтогенеза анамний. Изменчивость морфогенеза превращается в последовательность

РЕТРОГРАДНАЯ ЭВОЛЮЦИЯ СИГНАЛЬНЫХ ПУТЕЙ В ЭМБРИОГНЕЗЕ ХОРДОВЫХ
У Xenopus и Zebrafish β-катенин
РЕТРОГРАДНАЯ ЭВОЛЮЦИЯ СИГНАЛЬНЫХ ПУТЕЙ В ЭМБРИОГНЕЗЕ ХОРДОВЫХ
У Xenopus и Zebrafish β-катенин
 Кити і дельфіни
Кити і дельфіни Встречаем птиц
Встречаем птиц Кожа
Кожа Плоские черви. (Лекция 4)
Плоские черви. (Лекция 4) Область, объект и предмет исследования. Понятийная база и концепции. Особенности биологических объектов
Область, объект и предмет исследования. Понятийная база и концепции. Особенности биологических объектов Опасные животные Нижегородской области
Опасные животные Нижегородской области Kingdom: animalia
Kingdom: animalia Особенности агрофитоценозов как антропогенно измененных НЭС
Особенности агрофитоценозов как антропогенно измененных НЭС Пчёлы. Продолжительность Жизни пчёл
Пчёлы. Продолжительность Жизни пчёл Жиры, Белки и Углеводы человека
Жиры, Белки и Углеводы человека Красноухая черепаха
Красноухая черепаха Структура биосферы
Структура биосферы Проект Геном человека - занятие по биохимии
Проект Геном человека - занятие по биохимии Биотические факторы
Биотические факторы Разнообразие животных. Урок 19
Разнообразие животных. Урок 19 Факторы, негативно влияющие на дыхательную систему
Факторы, негативно влияющие на дыхательную систему Разработка урока по биологии Углеводы и липиды 9 класс (проектная деятельность)
Разработка урока по биологии Углеводы и липиды 9 класс (проектная деятельность) Tissues. The histophysiology of the epithelial tissue
Tissues. The histophysiology of the epithelial tissue Ищите доктора в природе
Ищите доктора в природе Тип плоские черви. Класс цестоды
Тип плоские черви. Класс цестоды Дикі та свійські тварини
Дикі та свійські тварини ГИА 2017-18 по биологии. Результаты и подготовка
ГИА 2017-18 по биологии. Результаты и подготовка Спортивна фізіологія. Фізіологічна характеристика фізичних вправ. (Лекція 2)
Спортивна фізіологія. Фізіологічна характеристика фізичних вправ. (Лекція 2) Введение в физиологию. Развитие представления о рефлексе и роль отечественных ученых в создании рефлекторной теории
Введение в физиологию. Развитие представления о рефлексе и роль отечественных ученых в создании рефлекторной теории Генетические заболевания. Дальтонизм
Генетические заболевания. Дальтонизм Строение цветка. 6 класс
Строение цветка. 6 класс Виды молочного сырья и показатели его качества
Виды молочного сырья и показатели его качества Эндокринная система
Эндокринная система