- Вторичные посредники: производные липидов

Содержание
- 2. Состоит из 4-х субъединиц-доменов (тетрамер) Структура домена: сайт связывания с ИФ3 сайт сцепления типичный Са2+-канал (6
- 3. Nuclear patch-clamp electrophysiology. A: schematic of cell nucleus, illustrating that the outer membrane of the double-membrane
- 5. Typical single-channel current traces of X-InsP3R-1 in various cytoplasmic Ca2+ concentrations and saturating 10 µM InsP3.
- 7. Вторичные посредники: производные липидов ДАГ остается в мембране и активирует протеинкиназу С, которая фосфорилирует свои субстратные
- 8. Активация фосфолипазы C Норадреналин (NE) активирует цепи внутриклеточной сигнализации с участием внутриклеточных липидов в роли вторичных
- 9. Вторичные посредники: арахидоновая кислота синтез и производные ДАГ (отделен красным пунктиром) является источником еще одного вторичного
- 10. Вторичные посредники: арахидоновая кислота синтез и производные ДАГ является источником еще одного вторичного посредника липидного происхождения.
- 11. Вторичные посредники: арахидоновая кислота синтез и производные
- 12. Вторичные посредники: арахидоновая кислота ДАГ является источником еще одного вторичного посредника липидного происхождения. Некоторые G-белки активируют
- 13. Вторичные посредники: арахидоновая кислота Арахидоновая кислота активирует протеинкиназу С, которая в свою очередь фосфорилирует ионные каналы.
- 14. NO - вторичный посредник нежели необычный медиатор NO синтезируется NO-синтазой, которая в нейронах регулируется Са2+/калмодулином. NO
- 15. Каскад вторичного посредника NO When the NMDA receptor is activated, Ca2+ enters the cytosol, binds to
- 16. Вторичные посредники: СO Свойства моноокисида углерода (CO) схожи со свойствами NO. СО продуцируется в эндотелиальных клетках
- 17. Каскад вторичного посредника CO Heme has a highly conjugated porphyrin ring with an iron atom chelated
- 18. Еще вторичные посредники В сердце и поджелудочной железе рианодиновый рецептор активируется вторичным посредником - циклической АДФ-рибозой
- 19. Еще вторичные посредники D-serine is a co-agonist at the NMDA receptor. Type II astrocytes ensheath synapses
- 20. Некоторые киназы, активируемые через системы вторичных посредников Вторичные посредники регулируют нейронные функции, модулируя состояние внутриклеточных белков
- 21. Некоторые киназы: эффекты фосфорилирования Субстраты киназ и фосфатаз включают ферменты, лиганд-зависимые рецепторы, ионные каналы и структурные
- 22. Некоторые киназы: группы киназ и фосфатаз Протеин киназы и фосфатазы действуют на сериновые и треониновые остатки
- 23. Киназы, активируемые через системы вторичных посредников В мозге представлены тысячи протеинкиназ, но только малая их часть
- 24. цАМФ-зависимая протеин киназа (ПКА) (англ., cAMP-dependent protein kinase, PKA) активируется при увеличении концентрации цАМФ. Увеличение концентрации
- 25. цГМФ-зависимая протеин киназа (ПКГ) (англ., cGMP-dependent protein kinase, PKG) активируется при увеличении концентрации цГМФ. Увеличение концентрации
- 26. цАМФ(цГМФ)-зависимые протеин киназы (PKA, PKG)
- 27. Ca2+/калмодулин-зависимая протеинкиназа типа II (Ca2+/calmodulin-dependent protein kinase type II, CaMKII) регулируется комплексом Ca2+/калмодулин. Является серин/треониновой протеин
- 28. Ca2+/калмодулин-зависимая протеинкиназа типа II
- 29. Протеин киназа C (ПКС) (англ., protein kinase C, PKC) активируется в присутствии Ca2+, ДАГ и фосфолипида
- 30. Протеин киназа C (ПКС) (англ., protein kinase C, PKC)
- 31. Различные киназы, активируемые через системы вторичных посредников Domain structure of protein kinases. Protein kinases are encoded
- 32. Тирозинкиназы (tyrosine kinases) активируется либо внеклеточными сигналами, либо G-белками. Участвуют в каскадах, обеспечивающих рост клеток и
- 33. Рецепторная тирозинкиназа (receptor tyrosine kinase) Известно три рецептора рецепторной тирозин киназы: Рецептор Лиганд TrkA NGF (nerve
- 34. Белковая тирозинкиназа (protein tyrosine kinase) Существует два класса тирозиновых протеин киназ, которые фосфорилируют тирозиновые остатки белков-мишеней.
- 35. Тирозинкиназы (tyrosine kinases)
- 36. Митоген-активируемые протеин киназы (МАПК) (Mitogen-activated protein kinase, MAPK) Активируются как вторичными посредниками, так и при фосфорилировании
- 37. Митоген-активируемые протеин киназы (МАПК)
- 41. Фосфатазы (РР от англ., Protein Phosphatase) Выделяют сериновые/треониновые, тирозиновые и двойственные фосфатазы. Наиболее изученными являются сериновые/треониновые
- 42. Фосфатазы Все фосфатазы не характеризуются высокой субстратной специфичностью по сравнению с протеин киназами. PP1 дефосфорилирует широкий
- 43. Фосфатазы Domain structure of the catalytic subunits of some Ser/Thr phosphatases. The three major phosphoprotein phosphatases,
- 44. Сигнализация в ядро Вторичные посредники активируют синтез новой РНК и, соответственно, белков. Первым шагом для синтеза
- 45. Сигнализация в ядро Steps involved in transcription of DNA into RNA. Condensed chromatin (left) is decondensed
- 46. Сигнализация в ядро CREB (cAMP Response Element Binding protein) является повсеместным фактором транкрипции. CREB связывается со
- 48. Скачать презентацию
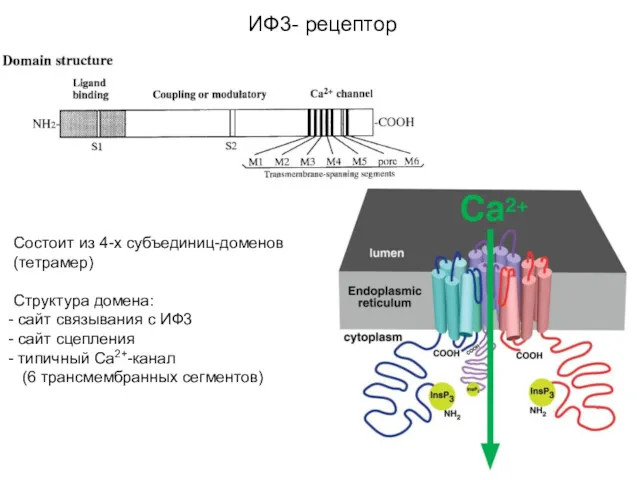
Состоит из 4-х субъединиц-доменов (тетрамер)
Структура домена:
сайт связывания с ИФ3
сайт
Состоит из 4-х субъединиц-доменов (тетрамер)
Структура домена:
сайт связывания с ИФ3
сайт
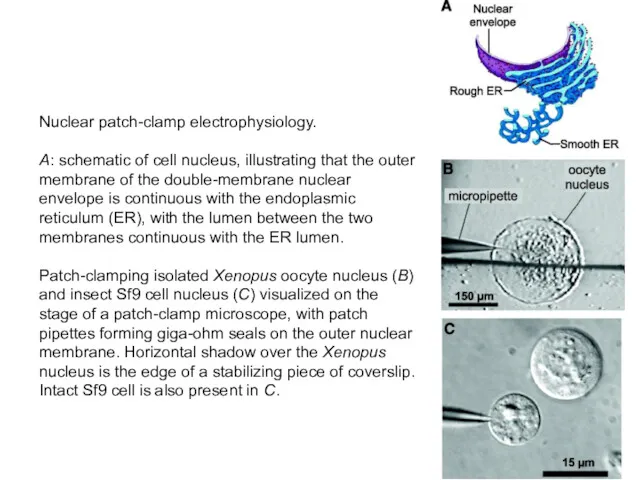
Nuclear patch-clamp electrophysiology.
A: schematic of cell nucleus, illustrating that the outer
Nuclear patch-clamp electrophysiology.
A: schematic of cell nucleus, illustrating that the outer

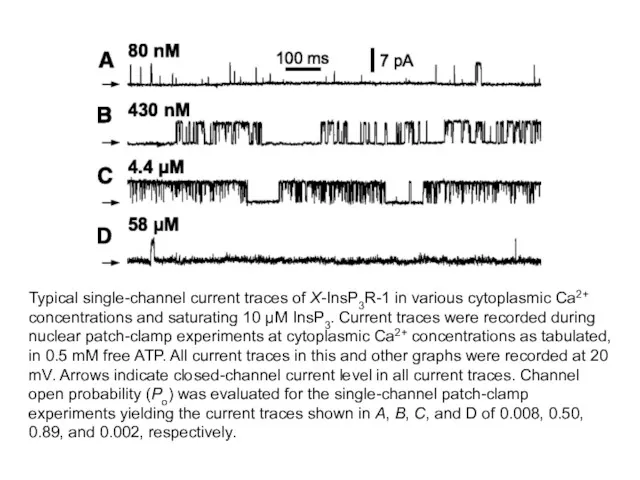
Typical single-channel current traces of X-InsP3R-1 in various cytoplasmic Ca2+ concentrations
Typical single-channel current traces of X-InsP3R-1 in various cytoplasmic Ca2+ concentrations

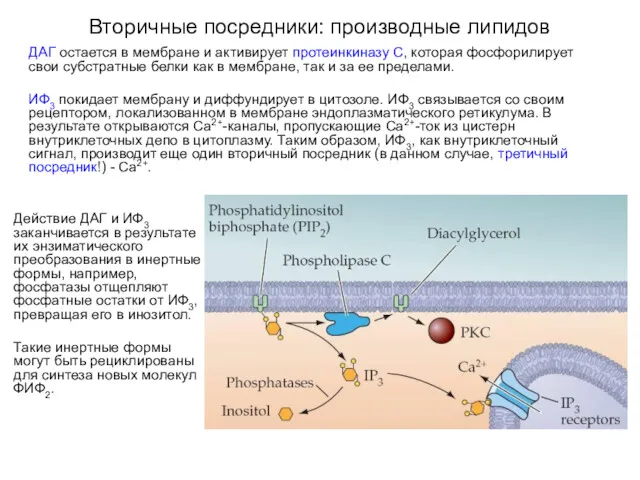
Вторичные посредники: производные липидов
ДАГ остается в мембране и активирует протеинкиназу С,
Вторичные посредники: производные липидов
ДАГ остается в мембране и активирует протеинкиназу С,
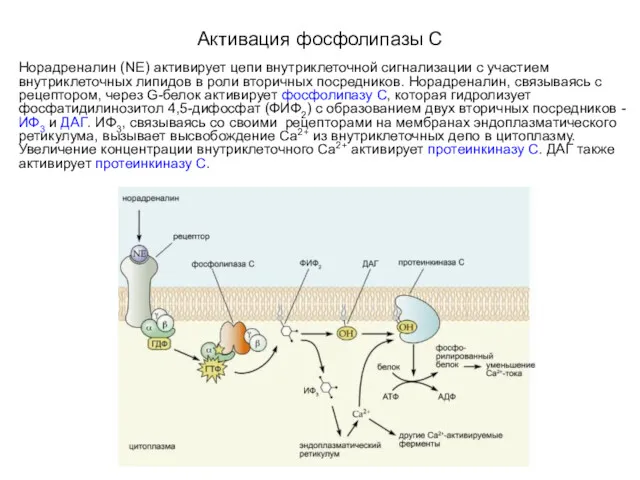
Активация фосфолипазы C
Норадреналин (NE) активирует цепи внутриклеточной сигнализации с участием
Активация фосфолипазы C
Норадреналин (NE) активирует цепи внутриклеточной сигнализации с участием
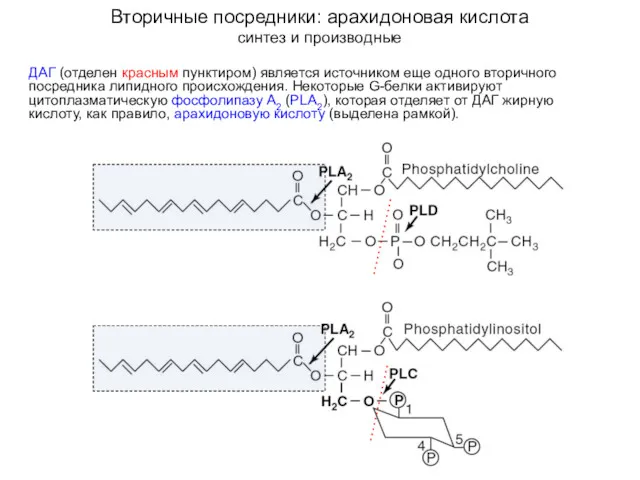
Вторичные посредники: арахидоновая кислота
синтез и производные
ДАГ (отделен красным пунктиром) является источником
Вторичные посредники: арахидоновая кислота
синтез и производные
ДАГ (отделен красным пунктиром) является источником
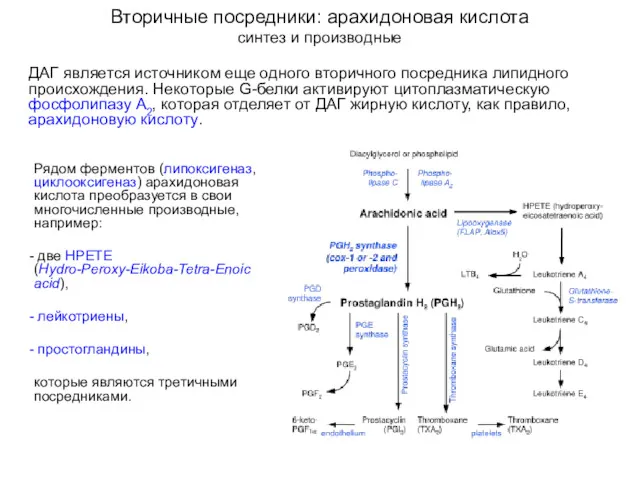
Вторичные посредники: арахидоновая кислота
синтез и производные
ДАГ является источником еще одного вторичного
Вторичные посредники: арахидоновая кислота
синтез и производные
ДАГ является источником еще одного вторичного
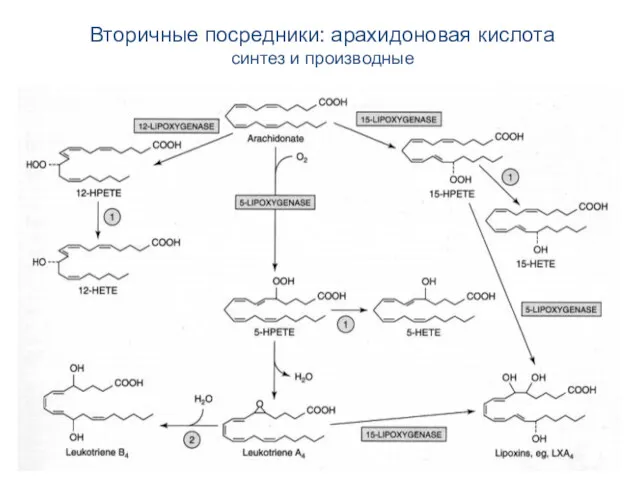
Вторичные посредники: арахидоновая кислота
синтез и производные
Вторичные посредники: арахидоновая кислота
синтез и производные
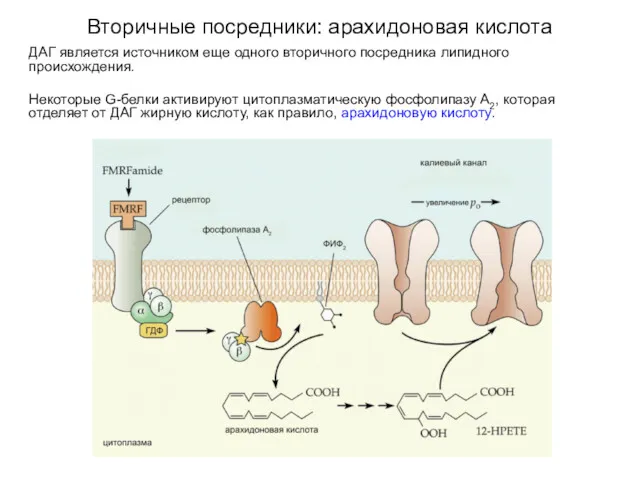
Вторичные посредники: арахидоновая кислота
ДАГ является источником еще одного вторичного посредника липидного
Вторичные посредники: арахидоновая кислота
ДАГ является источником еще одного вторичного посредника липидного
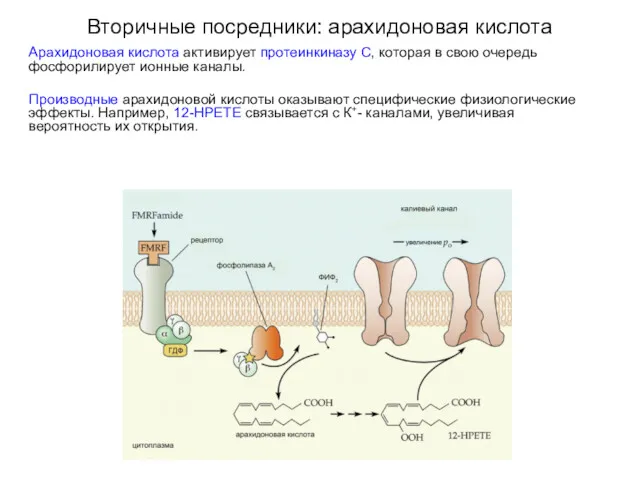
Вторичные посредники: арахидоновая кислота
Арахидоновая кислота активирует протеинкиназу С, которая в свою
Вторичные посредники: арахидоновая кислота
Арахидоновая кислота активирует протеинкиназу С, которая в свою
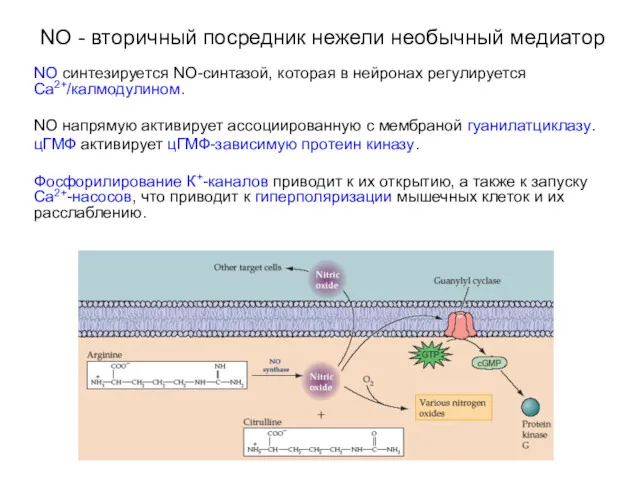
NO - вторичный посредник нежели необычный медиатор
NO синтезируется NO-синтазой, которая в
NO - вторичный посредник нежели необычный медиатор
NO синтезируется NO-синтазой, которая в
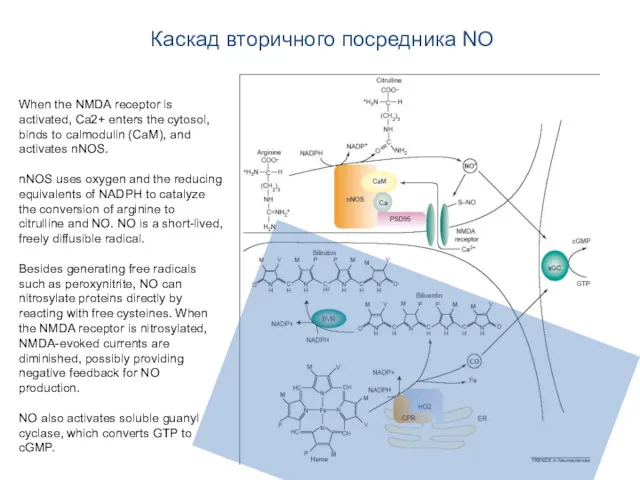
Каскад вторичного посредника NO
When the NMDA receptor is activated, Ca2+ enters
Каскад вторичного посредника NO
When the NMDA receptor is activated, Ca2+ enters

Вторичные посредники: СO
Свойства моноокисида углерода (CO) схожи со свойствами NO.
СО продуцируется
Вторичные посредники: СO
Свойства моноокисида углерода (CO) схожи со свойствами NO.
СО продуцируется
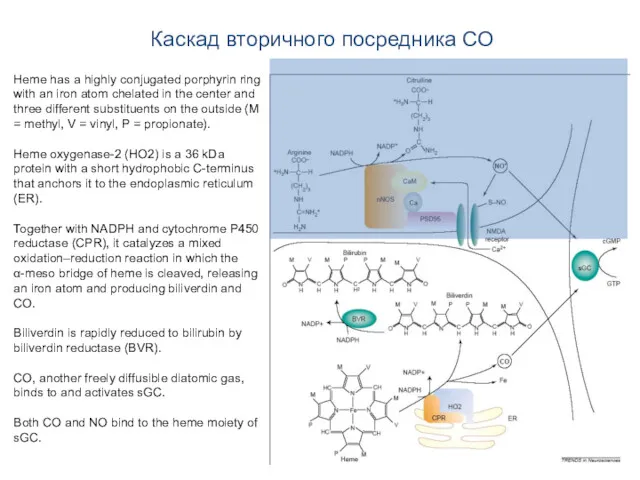
Каскад вторичного посредника CO
Heme has a highly conjugated porphyrin ring with
Каскад вторичного посредника CO
Heme has a highly conjugated porphyrin ring with
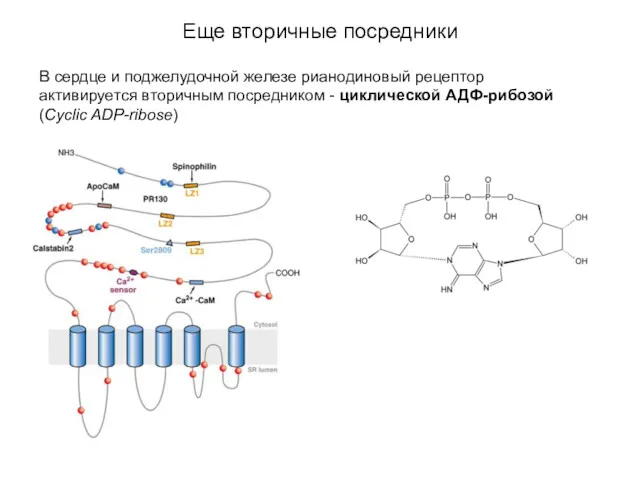
Еще вторичные посредники
В сердце и поджелудочной железе рианодиновый рецептор активируется вторичным
Еще вторичные посредники
В сердце и поджелудочной железе рианодиновый рецептор активируется вторичным
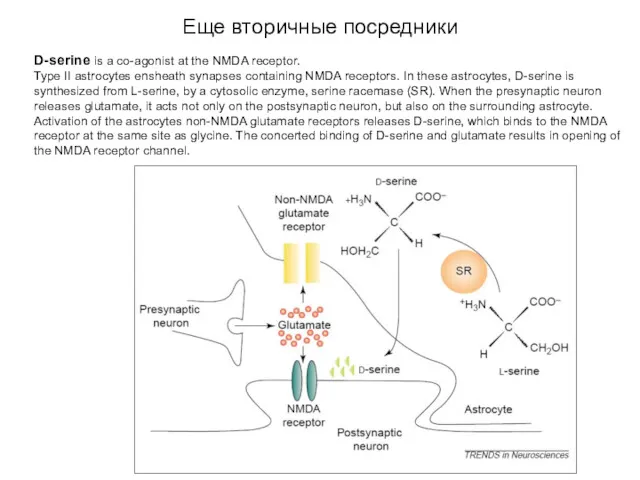
Еще вторичные посредники
D-serine is a co-agonist at the NMDA receptor.
Type
Еще вторичные посредники
D-serine is a co-agonist at the NMDA receptor.
Type
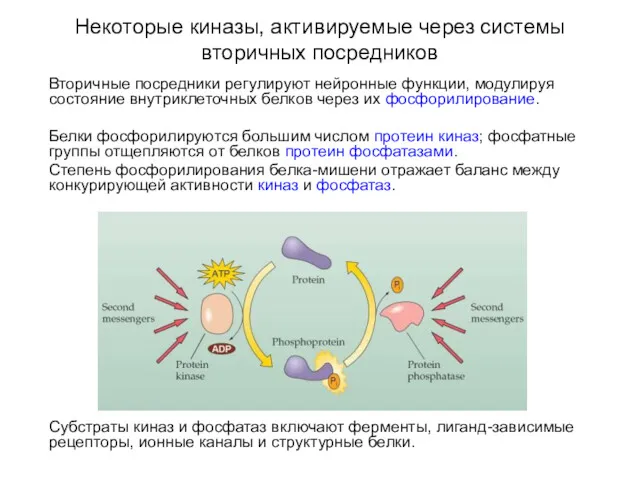
Некоторые киназы, активируемые через системы вторичных посредников
Вторичные посредники регулируют нейронные функции,
Некоторые киназы, активируемые через системы вторичных посредников
Вторичные посредники регулируют нейронные функции,

Некоторые киназы: эффекты фосфорилирования
Субстраты киназ и фосфатаз включают ферменты, лиганд-зависимые рецепторы,
Некоторые киназы: эффекты фосфорилирования
Субстраты киназ и фосфатаз включают ферменты, лиганд-зависимые рецепторы,

Некоторые киназы:
группы киназ и фосфатаз
Протеин киназы и фосфатазы действуют
на сериновые и
Некоторые киназы:
группы киназ и фосфатаз
Протеин киназы и фосфатазы действуют
на сериновые и
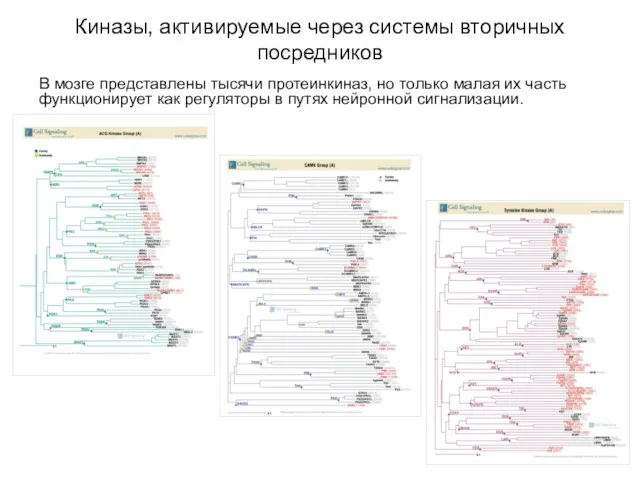
Киназы, активируемые через системы вторичных посредников
В мозге представлены тысячи протеинкиназ, но
Киназы, активируемые через системы вторичных посредников
В мозге представлены тысячи протеинкиназ, но
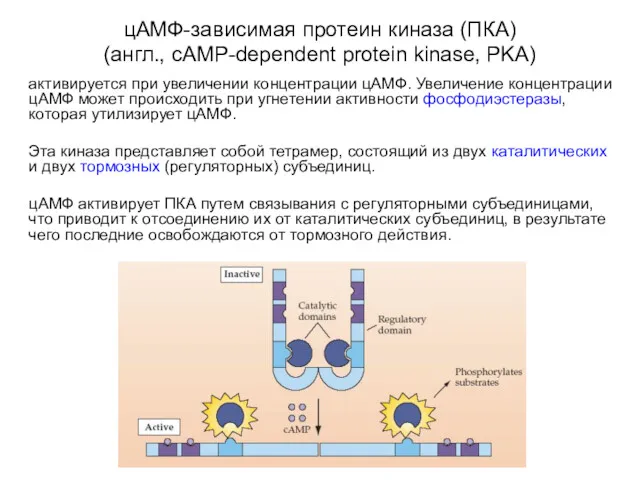
цАМФ-зависимая протеин киназа (ПКА)
(англ., cAMP-dependent protein kinase, PKA)
активируется при увеличении концентрации
цАМФ-зависимая протеин киназа (ПКА)
(англ., cAMP-dependent protein kinase, PKA)
активируется при увеличении концентрации

цГМФ-зависимая протеин киназа (ПКГ)
(англ., cGMP-dependent protein kinase, PKG)
активируется при увеличении концентрации
цГМФ-зависимая протеин киназа (ПКГ)
(англ., cGMP-dependent protein kinase, PKG)
активируется при увеличении концентрации
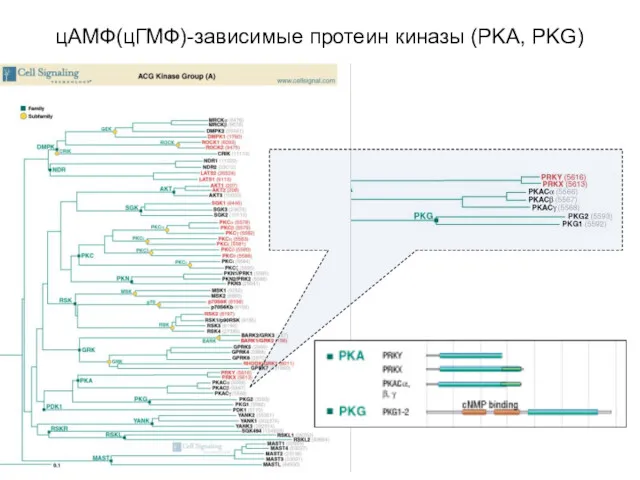
цАМФ(цГМФ)-зависимые протеин киназы (PKA, PKG)
цАМФ(цГМФ)-зависимые протеин киназы (PKA, PKG)

Ca2+/калмодулин-зависимая протеинкиназа типа II
(Ca2+/calmodulin-dependent protein kinase type II, CaMKII)
регулируется комплексом
Ca2+/калмодулин-зависимая протеинкиназа типа II
(Ca2+/calmodulin-dependent protein kinase type II, CaMKII)
регулируется комплексом
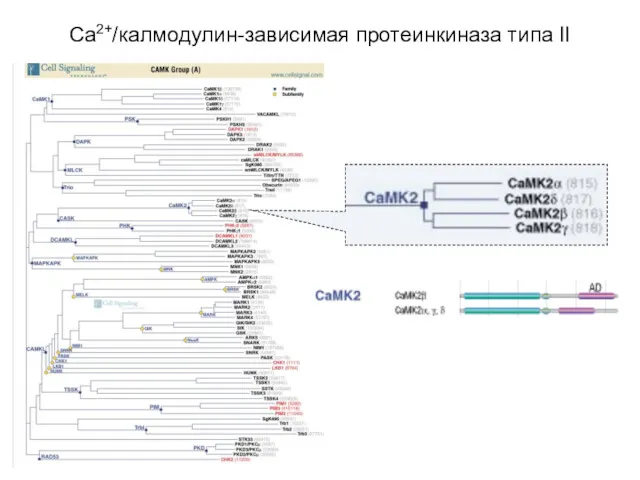
Ca2+/калмодулин-зависимая протеинкиназа типа II
Ca2+/калмодулин-зависимая протеинкиназа типа II
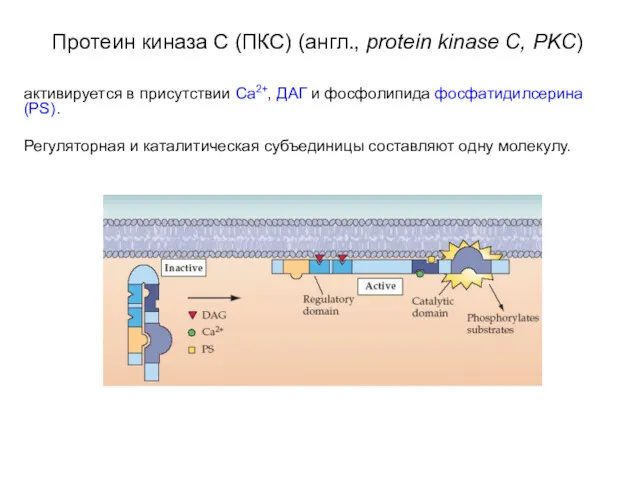
Протеин киназа C (ПКС) (англ., protein kinase C, PKC)
активируется в
Протеин киназа C (ПКС) (англ., protein kinase C, PKC)
активируется в
Протеин киназа C (ПКС) (англ., protein kinase C, PKC)
Протеин киназа C (ПКС) (англ., protein kinase C, PKC)
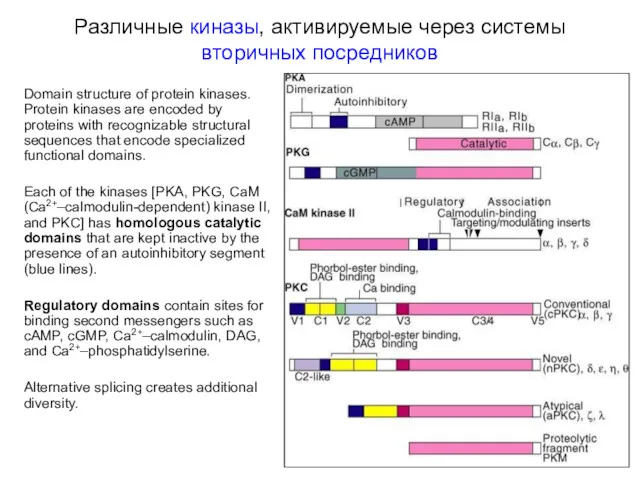
Различные киназы, активируемые через системы вторичных посредников
Domain structure of protein kinases.
Различные киназы, активируемые через системы вторичных посредников
Domain structure of protein kinases.
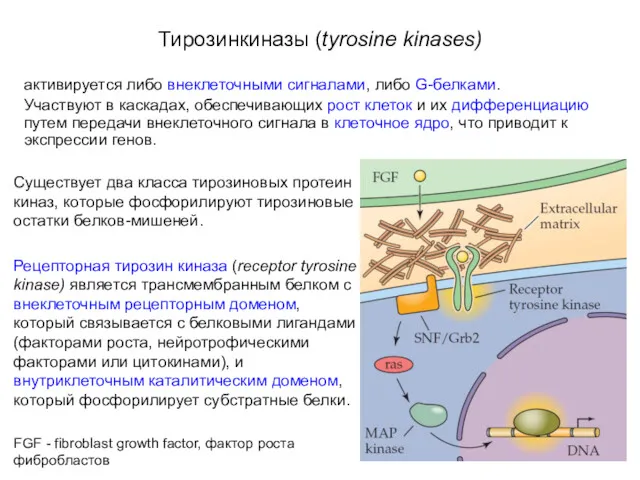
Тирозинкиназы (tyrosine kinases)
активируется либо внеклеточными сигналами, либо G-белками.
Участвуют в каскадах,
Тирозинкиназы (tyrosine kinases)
активируется либо внеклеточными сигналами, либо G-белками.
Участвуют в каскадах,
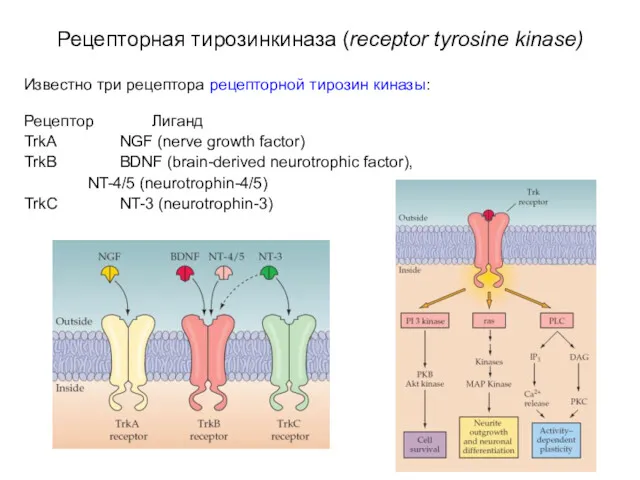
Рецепторная тирозинкиназа (receptor tyrosine kinase)
Известно три рецептора рецепторной тирозин киназы:
Рецептор
Рецепторная тирозинкиназа (receptor tyrosine kinase)
Известно три рецептора рецепторной тирозин киназы:
Рецептор
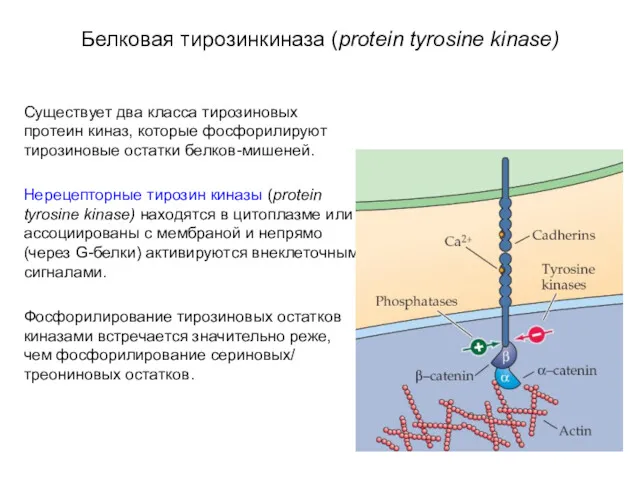
Белковая тирозинкиназа (protein tyrosine kinase)
Существует два класса тирозиновых протеин киназ,
Белковая тирозинкиназа (protein tyrosine kinase)
Существует два класса тирозиновых протеин киназ,
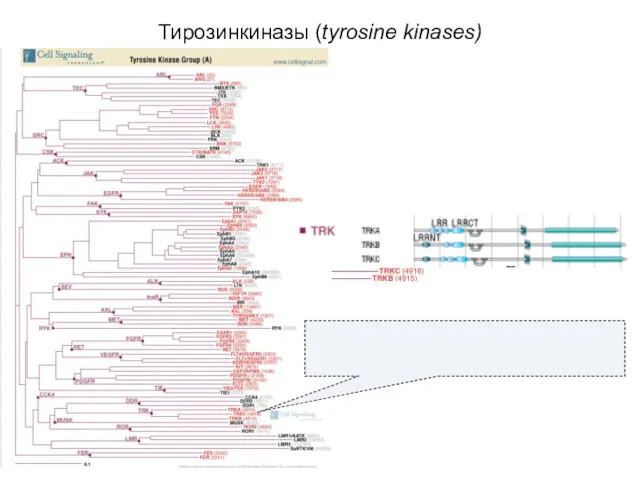
Тирозинкиназы (tyrosine kinases)
Тирозинкиназы (tyrosine kinases)
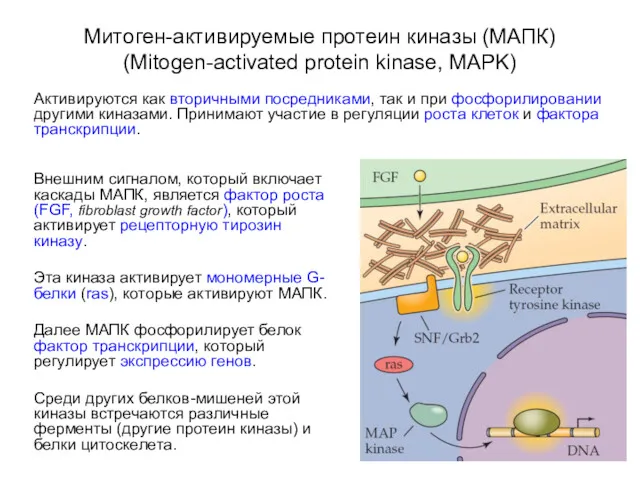
Митоген-активируемые протеин киназы (МАПК) (Mitogen-activated protein kinase, MAPK)
Активируются как вторичными
Митоген-активируемые протеин киназы (МАПК) (Mitogen-activated protein kinase, MAPK)
Активируются как вторичными
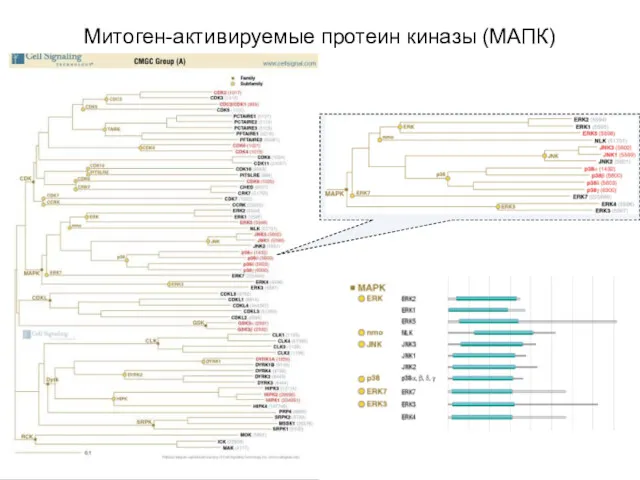
Митоген-активируемые протеин киназы (МАПК)
Митоген-активируемые протеин киназы (МАПК)




Фосфатазы (РР от англ., Protein Phosphatase)
Выделяют сериновые/треониновые, тирозиновые и двойственные фосфатазы.
Наиболее
Фосфатазы (РР от англ., Protein Phosphatase)
Выделяют сериновые/треониновые, тирозиновые и двойственные фосфатазы.
Наиболее

Фосфатазы
Все фосфатазы не характеризуются высокой субстратной специфичностью по сравнению с протеин
Фосфатазы
Все фосфатазы не характеризуются высокой субстратной специфичностью по сравнению с протеин

Фосфатазы
Domain structure of the catalytic subunits of some Ser/Thr phosphatases. The
Фосфатазы
Domain structure of the catalytic subunits of some Ser/Thr phosphatases. The

Сигнализация в ядро
Вторичные посредники активируют синтез новой РНК и, соответственно,
Сигнализация в ядро
Вторичные посредники активируют синтез новой РНК и, соответственно,
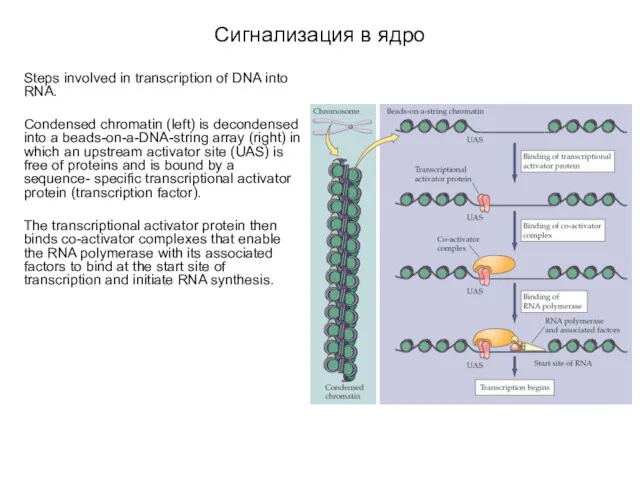
Сигнализация в ядро
Steps involved in transcription of DNA into RNA.
Condensed
Сигнализация в ядро
Steps involved in transcription of DNA into RNA.
Condensed
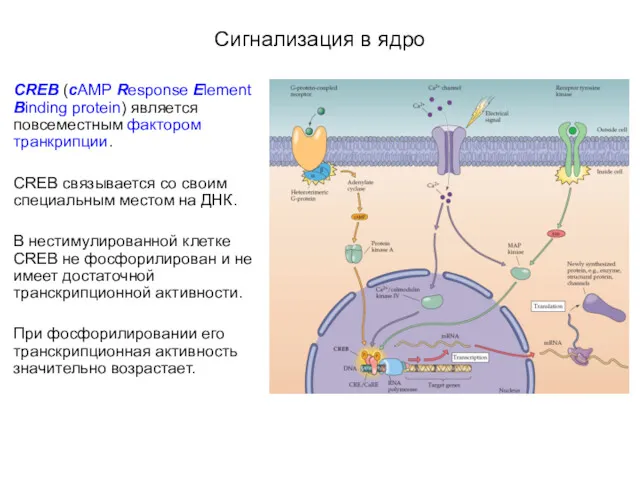
Сигнализация в ядро
CREB (cAMP Response Element Binding protein) является повсеместным фактором
Сигнализация в ядро
CREB (cAMP Response Element Binding protein) является повсеместным фактором
 Pallas' cat
Pallas' cat Исследовательская работа Влияние мобильного телефона на организм человека
Исследовательская работа Влияние мобильного телефона на организм человека Клетка. Строение клеток. Тест
Клетка. Строение клеток. Тест Кровообращение и лимфообращение
Кровообращение и лимфообращение Ягоды. Кроссворд
Ягоды. Кроссворд Декоративные качества плодов, стволов деревьев и кустарников
Декоративные качества плодов, стволов деревьев и кустарников Нервная система. Рефлекс. Инстинкт
Нервная система. Рефлекс. Инстинкт День птиц
День птиц Развитие кровеносной системы
Развитие кровеносной системы Механизмы влияния гуминовых препаратов на физиолого-биохимические процессы, происходящие в растениях
Механизмы влияния гуминовых препаратов на физиолого-биохимические процессы, происходящие в растениях Разнообразие животных
Разнообразие животных презентация к уроку Класс Птицы
презентация к уроку Класс Птицы Что такое адаптации? Какие бывают адаптации?
Что такое адаптации? Какие бывают адаптации? Молекулярно-генетические доказательства эволюции
Молекулярно-генетические доказательства эволюции Презентация к уроку биологии 7 класс животные Органы пищеварения
Презентация к уроку биологии 7 класс животные Органы пищеварения Простое вещество азот
Простое вещество азот тайна маленьких клеток
тайна маленьких клеток презентация Каскад водопадов на реке Шинок
презентация Каскад водопадов на реке Шинок В гости к осени. 2 класс
В гости к осени. 2 класс Ткани человека
Ткани человека Аквариум как химико-биологический объект исследования
Аквариум как химико-биологический объект исследования Презентация Вредные привычки. Законы РФ о курении
Презентация Вредные привычки. Законы РФ о курении Динозаврлар. Жалпы мәлімет
Динозаврлар. Жалпы мәлімет Тип Кольчатые черви
Тип Кольчатые черви Аптека под ногами
Аптека под ногами Генетическая инженерия в современной селекции и семеноводстве растений
Генетическая инженерия в современной селекции и семеноводстве растений 09_02_biologia
09_02_biologia Животные Арктических пустынь. Белый медведь
Животные Арктических пустынь. Белый медведь