- Введение в молекулярную биологию

Содержание
- 2. Введение в молекулярную биологию К.б.н., доц. Казанская Елена Анатольевна ГОУВПО СЕВЕРО-ЗАПАДНЫЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ ИМ. И.И.
- 3. План: Белок – субстрат жизни. Строение и функции белков. Нуклеиновые кислоты. Некоторые особенности организации наследственного аппарата
- 4. «Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их
- 5. Строение и функции белков Белки́ (протеи́ны) — высокомолекулярные органические вещества, состоящие из соединённых в цепочку пептидной
- 6. Первичная структура белка - это последовательность расположения аминокислотных остатков в полипептидной цепи. Аминокислоты соединяются в полипептид
- 7. Вторичная структура белка - это упорядоченное строение полипептидных цепей, обусловленное водородными связями между группами С=О и
- 8. Третичная структура белка - это пространственная конформация полипептида, имеющего вторичную структуру, и обусловленная взаимодействиями между радикалами.
- 9. Четвертичная структура белка - это агрегация двух или большего числа полипептидных цепей, имеющих третичную структуру, в
- 10. Функции белков Ферментативная: гидролазы, каталазы, полимеразы, синтетазы и др. ферменты. Регуляторная: белки-репрессоры и белки-активаторы транскрипции. Рецепторная:
- 11. 2. Нуклеиновые кислоты Структура нуклеотида Нуклеиновые кислоты – это линейные полимеры, состоящие из последовательно расположенных структурных
- 12. Пурины: аденин (А) и гуанин (Г) - содержат два гетероцикла. Пиримидины: тимин (Т), цитозин (Ц) и
- 13. Субстратом для построения цепи нуклеиновой кислоты являются нуклеозидтрифосфаты. Соединение нуклеотидов происходит путем взаимодействия фосфата одного нуклеотида
- 14. Нуклеиновые кислоты Дезоксирибонуклеиновая кислота (ДНК) Рибонуклеиновая кислота (РНК)
- 15. ДНК. Дезоксирибонуклеотид – состоит из остатков трех веществ: азотистого основания, дезоксирибозы (пентозы), фосфорной кислоты. Пиримидиновые азотистые
- 16. ДНК. Канонические пары оснований: Аденин (А) – Тимин (Т) образуют 2 водородные связи Гуанин (Г) –
- 17. Модель строения ДНК, предложенная Уотсоном и Криком (1953) ДНК. Двуцепочечная молекула Скелетная основа полинуклеотидных цепей содержит
- 18. При исследовании синтетических молекул ДНК было показано, что ДНК может принимать различные конформации. Наиболее часто встречается
- 19. РНК. Рибонуклеотид – состоит из остатков трех веществ: 1) азотистого основания, 2) рибозы (пентозы) 3) фосфорной
- 20. Транспортные РНК содержат обычно 76 нуклеотидов (около 10% от общего содержания РНК в клетке). Функции т-РНК:
- 21. 3. Некоторые особенности организации наследственного аппарата у прокариот, эукариот и вирусов.
- 22. Особенности организации наследственного аппарата прокариот. Нуклеоид (генофор) - кольцевая двуспиральная правозакрученная молекула ДНК, которая свернута во
- 23. 1. Объем=106 пары нуклеотидов 2. Молекула ДНК 3. ДНК не связана с гистонами 4. Большую часть
- 24. Особенности организации наследственного аппарата вирусов. Способы увеличения генетической информации: 1) двукратное считывание одной и той же
- 25. Гены, кодирующие рРНК и тРНК в геноме вирусов обычно отсутствуют. Геном вируса включает: структурные гены, которые
- 26. Вирусные РНК в зависимости от выполняемых функций подразделяются на две группы. 1. плюс-нити РНК (+РНК, позитивный
- 27. Рамка считывания Gag кодирует полипептид, содержащий белок капсида; pol — ферменты, ответственные за цикл размножения вируса,
- 28. 4. Реализация генетической информации в клетке.
- 29. Центральная догма молекулярной биологии Центральная догма* молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической
- 30. Способы передачи информации общие ДНК→ДНК (репликация) ДНК→РНК (транскрипция) РНК→белок (трансляция) специальные РНК→ДНК (обратная транскрипция) Способы передачи
- 31. Матричные процессы репликация транскрипция трансляция репарация Матричные процессы Характеристика: Наличие матрицы Участие ферментов Затраты энергии Три
- 32. Репликация ДНК Репликация ДНК– процесс образования идентичных копий ДНК, осуществляемый комплексом ферментов и структурных белков. Репликация
- 33. ДНК-полимеразы – осуществляют синтез новых цепей ДНК Ферменты репликации топоизомераза геликаза (хеликаза) праймаза (РНК-полимераза) эндонуклеаза ДНК-лигаза
- 34. ДНК- полимераза праймаза Затравка РНК Удаление затравки Свойства ДНК-полимераз
- 35. Репликация ДНК. Разъединение двух цепей ДНК у эукариот начинается одновременно в нескольких участках (у прокариот в
- 36. Родительская ДНК Ранняя стадия репликации Репликативные вилки Движение репликативных вилок Слияние репликативных вилок Поздняя стадия репликации
- 37. Единица репликации – репликон. Репликон – молекула ДНК или ее участок, способные к автономной репликации. Инициация
- 38. Этапы репликации 1. Инициация (начало синтеза дочерних цепей ДНК) раскручивание ДНК разрыв водородных связей формирование репликативной
- 39. Этапы репликациии 2. Элонгация (удлинение дочерних цепей ДНК) рост цепи (синтез ДНК) синтез РНК-затравки (праймера) замена
- 40. Этапы репликации 3. Терминация (завершение синтеза дочерних цепей ДНК) вырезание РНК-затравки (праймера) синтез цепи ДНК на
- 41. Взаимное пространственное соответствие структур двух молекул или их частей, позволяющее формировать функционально целостный комплекс за счёт
- 42. Транскрипция Транскрипция осуществляется при помощи фермента РНК-полимеразы. Транскрипция – биосинтез одноцепочечной молекулы РНК на матрице ДНК.
- 43. Единица транскрипции – транскриптон. Схема транскриптона у прокариот Схема транскриптона у эукариот Интроны – неинформативные участки
- 44. Сложная инициация транскрипции у эукариот (пример) соединение элемента перед промотором (ТАТА-бокса) с ТАТА-фактором (ТF) «узнавание» РНК-полимеразой
- 45. Транскрипция 1. Инициация 2. Элонгация 3. Терминация - расплетание ДНК на участке 10-20 нуклеотидов - формирование
- 46. Посттранскрипционные изменения (процессинг) Процессинг (созревание) мРНК – совокупность биохимических реакций, в результате которых происходят структурные и
- 47. Генетический код Генетический код – это способ записи генетической информации о структуре белков (полипептидов) посредством последовательности
- 48. Генетический код Единица генетического кода – кодон (триплет нуклеотидов). Триплет или кодон - последовательность из трех
- 49. Свойства генетического кода: помехоустойчивость – отношение числа консервативных замен к числу радикальных замен. универсальность - принцип
- 50. Трансляция (синтез белка) Состоит из двух стадий: Активирование аминокислоты. Присоединение аминокислоты к tРНК (аминоацилирование). Подготовительный этап
- 51. Трансляция (синтез белка) 1. Инициация – сборка активной рибосомы -связывание мРНК с малой субъединицей рибосомы -стартовый
- 52. 2. Элонгация – рост пептидной цепи -связывание новой тРНК с аминоацильным центром -замыкание пептидной связи между
- 53. УАА (UAA), УАГ (UAG), УГА (UGA) – стоп-кодоны 3. Терминация – прерывание трансляции Трансляция (синтез белка)
- 54. Передача генетической информации ретровируса. Обратная транскрипция.
- 55. Передача генетической информации ретровируса. Обратная транскрипция ВИЧ Онковирусы +РНК Синтез ДНК на матрице РНК - процесс
- 56. Открытие явления обратной транскрипции сыграло большую роль в развитии генной инженерии, микробиологии. С помощью ревертаз получают
- 57. Репарация -процесс устранения повреждений нуклеотидной последовательности ДНК. Восстановления структуры ДНК осуществляются особыми ферментативными системами клетки (ферменты
- 58. “Узнавание” поврежденного участка ДНК (фермент эндонуклеаза). 2. Надрезание одной цепи ДНК в месте повреждения (фермент эндонуклеаза).
- 59. Фоторепарация (световая) Механизм: Под влияние УФ-лучей образуются димеры пиримидиновых оснований. Свет активирует фермент фотолиазу, которая узнает
- 61. Скачать презентацию

Введение в
молекулярную биологию
К.б.н., доц. Казанская Елена Анатольевна
ГОУВПО СЕВЕРО-ЗАПАДНЫЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
ИМ.
Введение в
молекулярную биологию
К.б.н., доц. Казанская Елена Анатольевна
ГОУВПО СЕВЕРО-ЗАПАДНЫЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
ИМ.

План:
Белок – субстрат жизни. Строение и функции белков.
Нуклеиновые кислоты.
Некоторые особенности организации
План:
Белок – субстрат жизни. Строение и функции белков.
Нуклеиновые кислоты.
Некоторые особенности организации

«Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен
«Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен

Строение и функции белков
Белки́ (протеи́ны) — высокомолекулярные органические вещества, состоящие из соединённых
Строение и функции белков
Белки́ (протеи́ны) — высокомолекулярные органические вещества, состоящие из соединённых
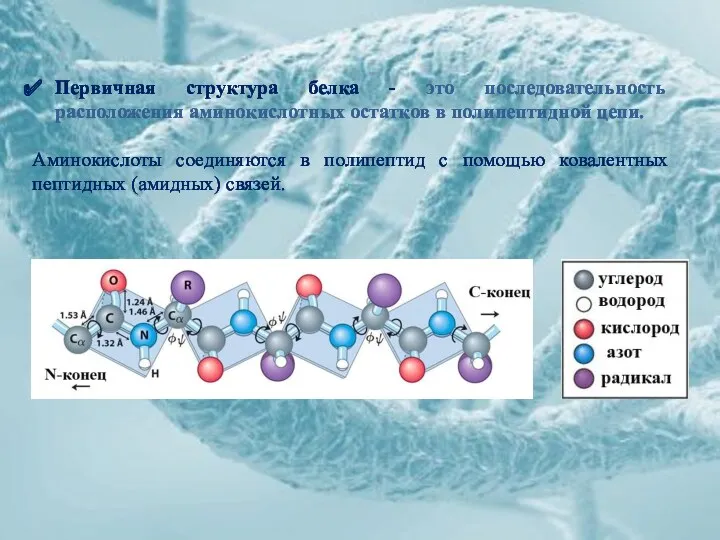
Первичная структура белка - это последовательность расположения аминокислотных остатков в полипептидной
Первичная структура белка - это последовательность расположения аминокислотных остатков в полипептидной
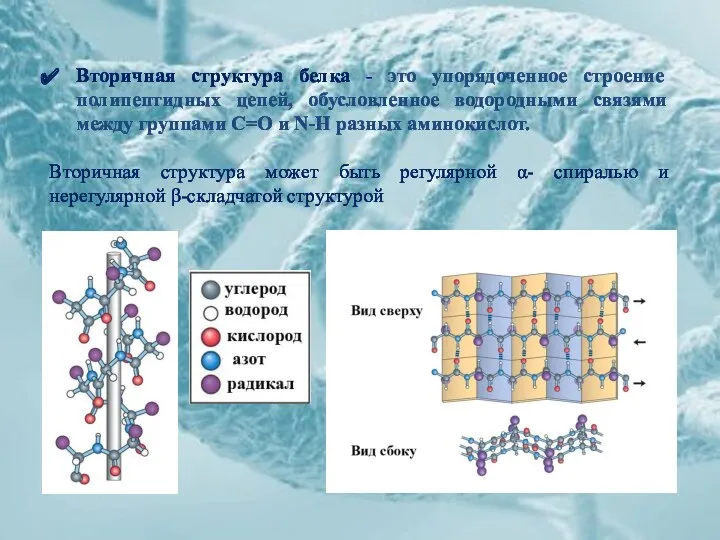
Вторичная структура белка - это упорядоченное строение полипептидных цепей, обусловленное водородными
Вторичная структура белка - это упорядоченное строение полипептидных цепей, обусловленное водородными
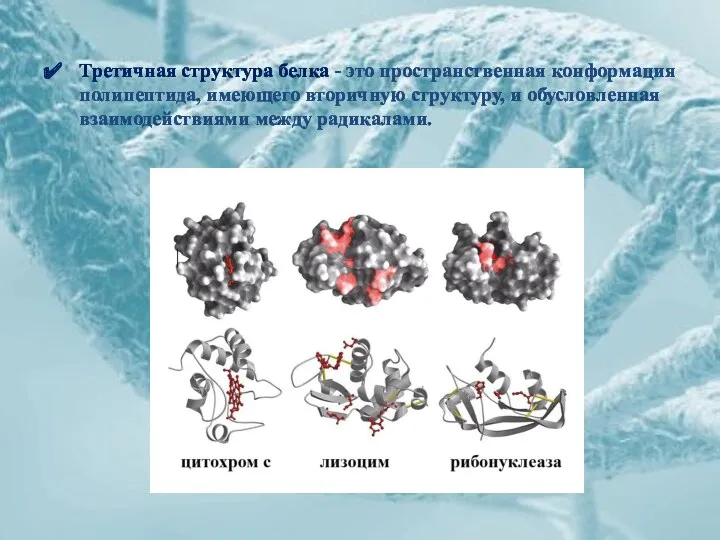
Третичная структура белка - это пространственная конформация полипептида, имеющего вторичную структуру,
Третичная структура белка - это пространственная конформация полипептида, имеющего вторичную структуру,
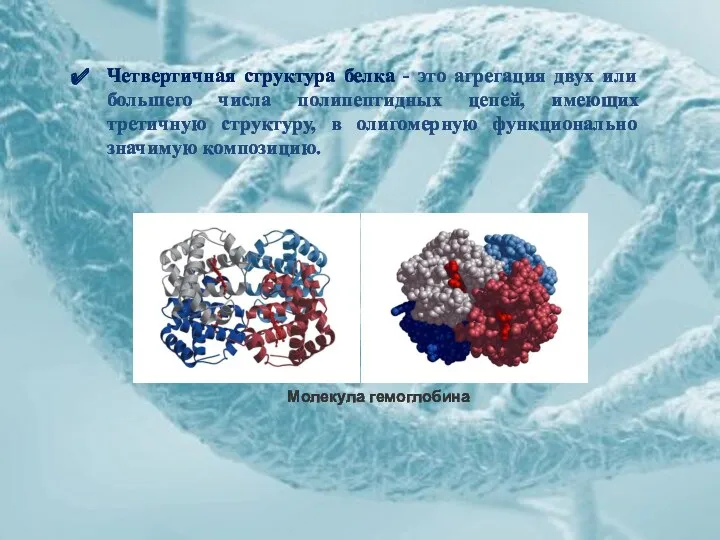
Четвертичная структура белка - это агрегация двух или большего числа полипептидных
Четвертичная структура белка - это агрегация двух или большего числа полипептидных

Функции белков
Ферментативная: гидролазы, каталазы, полимеразы, синтетазы и др. ферменты.
Регуляторная: белки-репрессоры и
Функции белков
Ферментативная: гидролазы, каталазы, полимеразы, синтетазы и др. ферменты.
Регуляторная: белки-репрессоры и
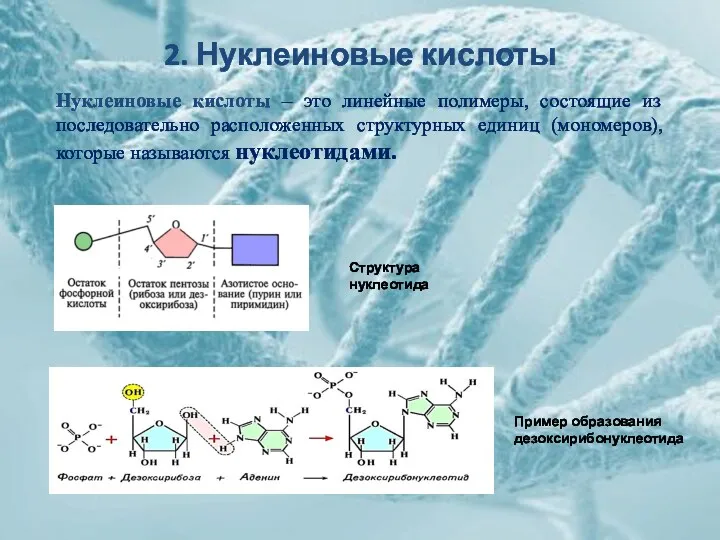
2. Нуклеиновые кислоты
Структура нуклеотида
Нуклеиновые кислоты – это линейные полимеры, состоящие из
2. Нуклеиновые кислоты
Структура нуклеотида
Нуклеиновые кислоты – это линейные полимеры, состоящие из
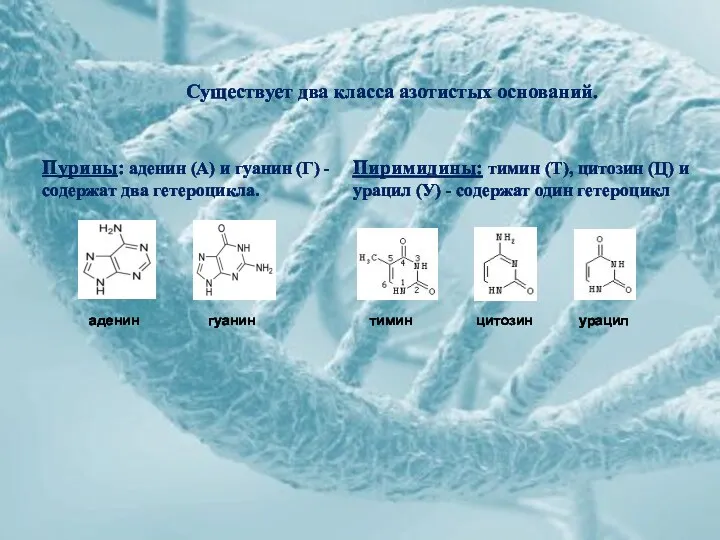
Пурины: аденин (А) и гуанин (Г) - содержат два гетероцикла.
Пиримидины:
Пурины: аденин (А) и гуанин (Г) - содержат два гетероцикла.
Пиримидины:
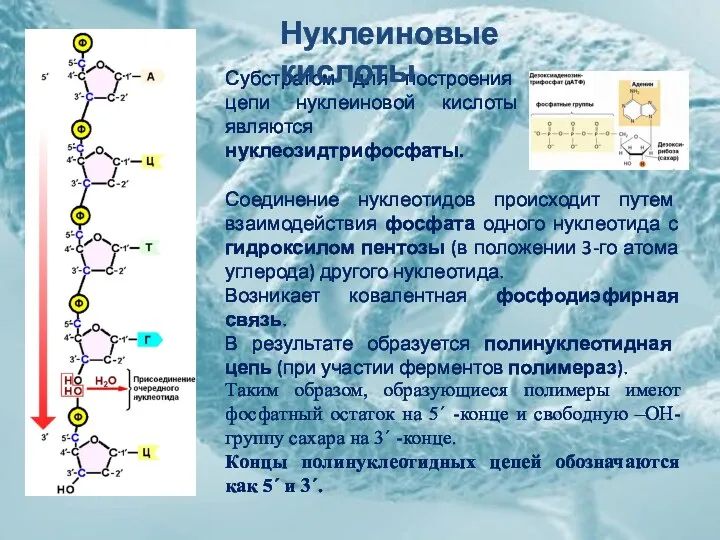
Субстратом для построения цепи нуклеиновой кислоты являются нуклеозидтрифосфаты.
Соединение нуклеотидов происходит путем
Субстратом для построения цепи нуклеиновой кислоты являются нуклеозидтрифосфаты.
Соединение нуклеотидов происходит путем
Нуклеиновые кислоты
Дезоксирибонуклеиновая кислота (ДНК)
Рибонуклеиновая кислота (РНК)
Нуклеиновые кислоты
Дезоксирибонуклеиновая кислота (ДНК)
Рибонуклеиновая кислота (РНК)

ДНК.
Дезоксирибонуклеотид – состоит из остатков трех веществ:
азотистого основания,
дезоксирибозы (пентозы),
фосфорной кислоты.
ДНК.
Дезоксирибонуклеотид – состоит из остатков трех веществ:
азотистого основания,
дезоксирибозы (пентозы),
фосфорной кислоты.
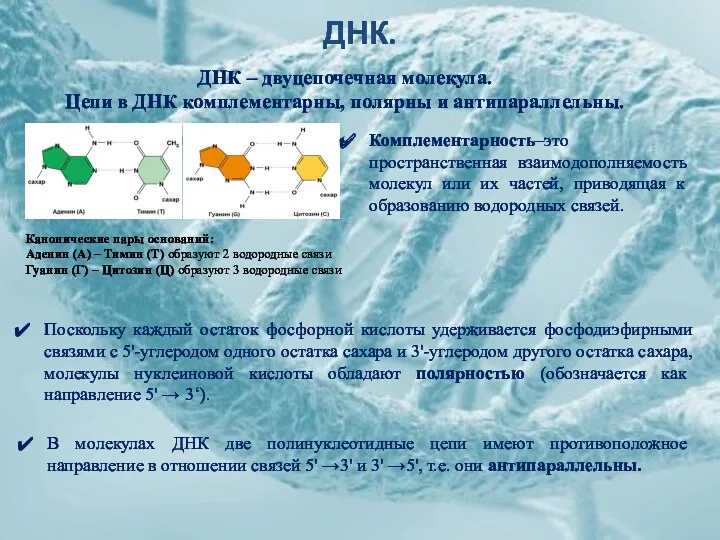
ДНК.
Канонические пары оснований:
Аденин (А) – Тимин (Т) образуют 2 водородные связи
Гуанин
ДНК.
Канонические пары оснований:
Аденин (А) – Тимин (Т) образуют 2 водородные связи
Гуанин

Модель строения ДНК, предложенная Уотсоном
и Криком (1953)
ДНК.
Двуцепочечная молекула
Скелетная основа полинуклеотидных
Модель строения ДНК, предложенная Уотсоном
и Криком (1953)
ДНК.
Двуцепочечная молекула
Скелетная основа полинуклеотидных

При исследовании синтетических молекул ДНК было показано, что ДНК может принимать
При исследовании синтетических молекул ДНК было показано, что ДНК может принимать
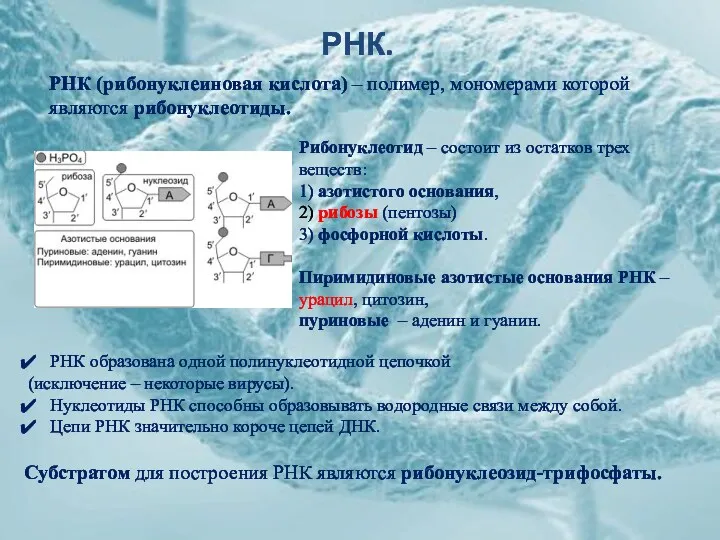
РНК.
Рибонуклеотид – состоит из остатков трех веществ:
1) азотистого основания,
2) рибозы
РНК.
Рибонуклеотид – состоит из остатков трех веществ:
1) азотистого основания,
2) рибозы

Транспортные РНК содержат обычно 76 нуклеотидов (около 10% от общего содержания
Транспортные РНК содержат обычно 76 нуклеотидов (около 10% от общего содержания

3. Некоторые особенности организации наследственного аппарата у прокариот, эукариот и вирусов.
3. Некоторые особенности организации наследственного аппарата у прокариот, эукариот и вирусов.
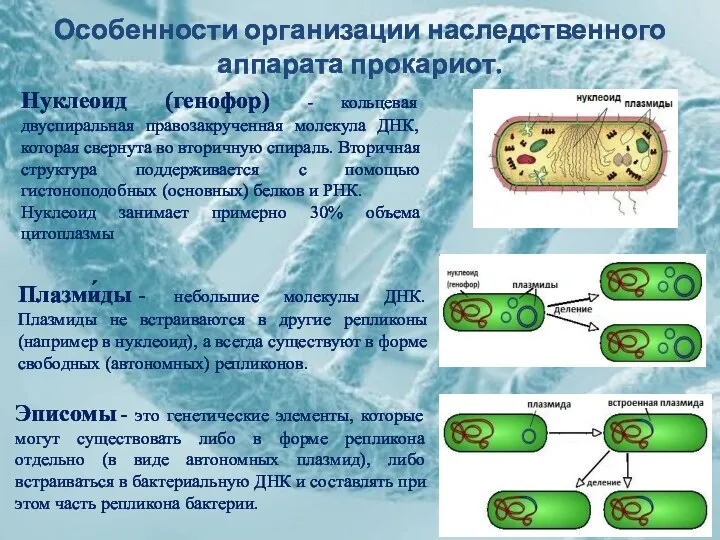
Особенности организации наследственного аппарата прокариот.
Нуклеоид (генофор) - кольцевая двуспиральная правозакрученная молекула
Особенности организации наследственного аппарата прокариот.
Нуклеоид (генофор) - кольцевая двуспиральная правозакрученная молекула

1. Объем=106 пары нуклеотидов
2. Молекула ДНК
3. ДНК не связана с гистонами
4.
1. Объем=106 пары нуклеотидов
2. Молекула ДНК
3. ДНК не связана с гистонами
4.
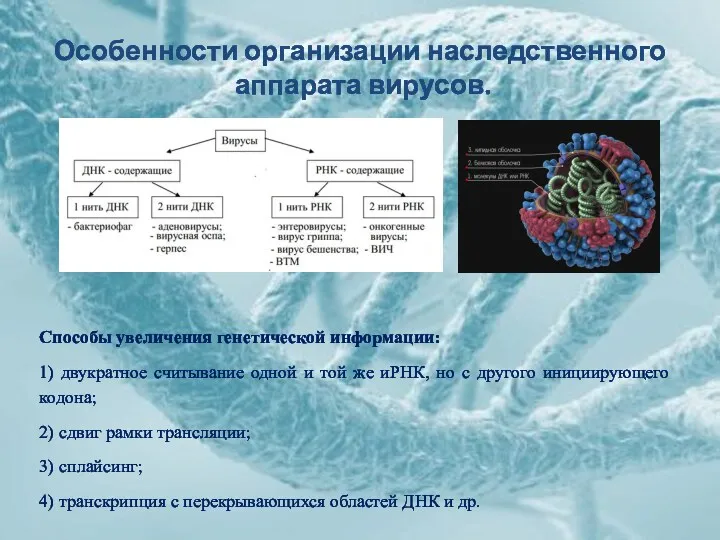
Особенности организации наследственного
аппарата вирусов.
Способы увеличения генетической информации:
1) двукратное считывание
Особенности организации наследственного
аппарата вирусов.
Способы увеличения генетической информации:
1) двукратное считывание

Гены, кодирующие рРНК и тРНК в геноме вирусов обычно отсутствуют.
Геном
Гены, кодирующие рРНК и тРНК в геноме вирусов обычно отсутствуют.
Геном

Вирусные РНК в зависимости от выполняемых функций подразделяются на две группы.
1.
Вирусные РНК в зависимости от выполняемых функций подразделяются на две группы.
1.

Рамка считывания Gag кодирует полипептид, содержащий белок капсида;
pol — ферменты, ответственные за цикл размножения вируса,
Рамка считывания Gag кодирует полипептид, содержащий белок капсида;
pol — ферменты, ответственные за цикл размножения вируса,

4. Реализация генетической информации в клетке.
4. Реализация генетической информации в клетке.
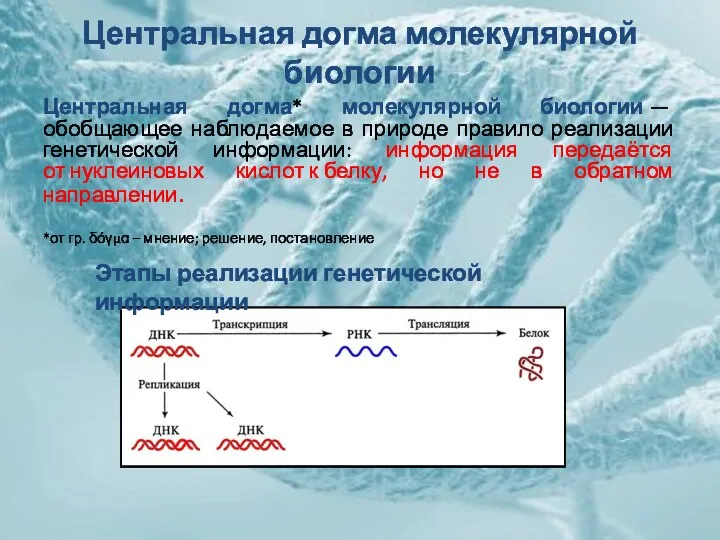
Центральная догма молекулярной биологии
Центральная догма* молекулярной биологии — обобщающее наблюдаемое в природе
Центральная догма молекулярной биологии
Центральная догма* молекулярной биологии — обобщающее наблюдаемое в природе
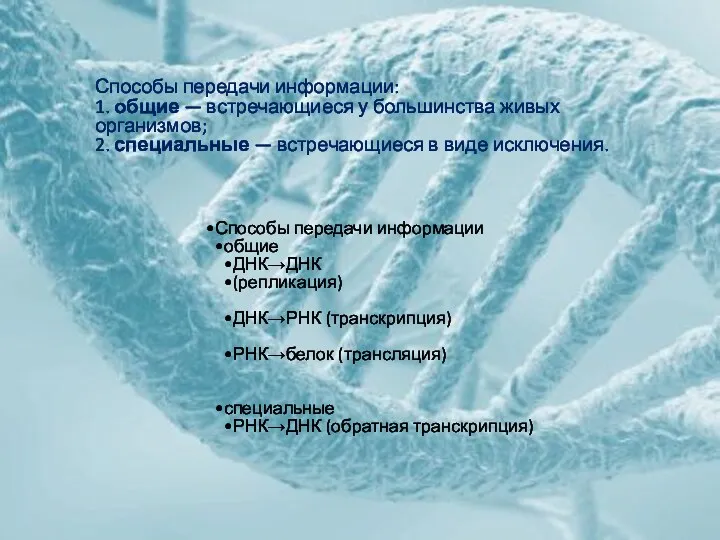
Способы передачи информации
общие
ДНК→ДНК
(репликация)
ДНК→РНК (транскрипция)
РНК→белок (трансляция)
специальные
РНК→ДНК (обратная транскрипция)
Способы передачи информации:
1. общие — встречающиеся
Способы передачи информации
общие
ДНК→ДНК
(репликация)
ДНК→РНК (транскрипция)
РНК→белок (трансляция)
специальные
РНК→ДНК (обратная транскрипция)
Способы передачи информации: 1. общие — встречающиеся
Матричные процессы
репликация
транскрипция
трансляция
репарация
Матричные процессы
Характеристика:
Наличие матрицы
Участие ферментов
Затраты энергии
Три этапа (инициация, элонгация, терминация)
Матричные процессы
репликация
транскрипция
трансляция
репарация
Матричные процессы
Характеристика:
Наличие матрицы
Участие ферментов
Затраты энергии
Три этапа (инициация, элонгация, терминация)
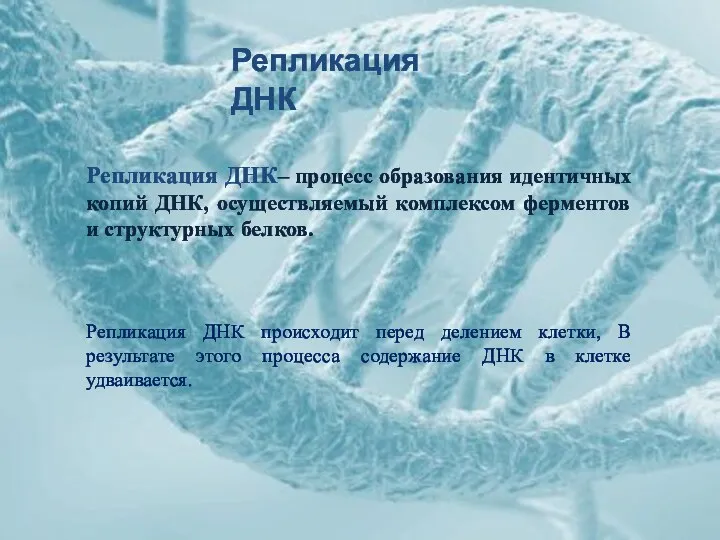
Репликация ДНК
Репликация ДНК– процесс образования идентичных копий ДНК, осуществляемый комплексом ферментов
Репликация ДНК
Репликация ДНК– процесс образования идентичных копий ДНК, осуществляемый комплексом ферментов

ДНК-полимеразы – осуществляют синтез новых цепей ДНК
Ферменты репликации
топоизомераза
геликаза (хеликаза)
праймаза
ДНК-полимеразы – осуществляют синтез новых цепей ДНК
Ферменты репликации
топоизомераза
геликаза (хеликаза)
праймаза
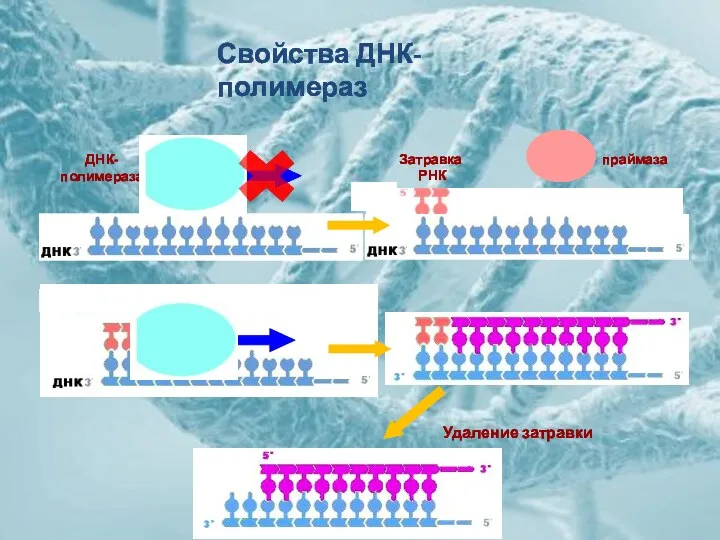
ДНК-
полимераза
праймаза
Затравка
РНК
Удаление затравки
Свойства ДНК-полимераз
ДНК-
полимераза
праймаза
Затравка
РНК
Удаление затравки
Свойства ДНК-полимераз
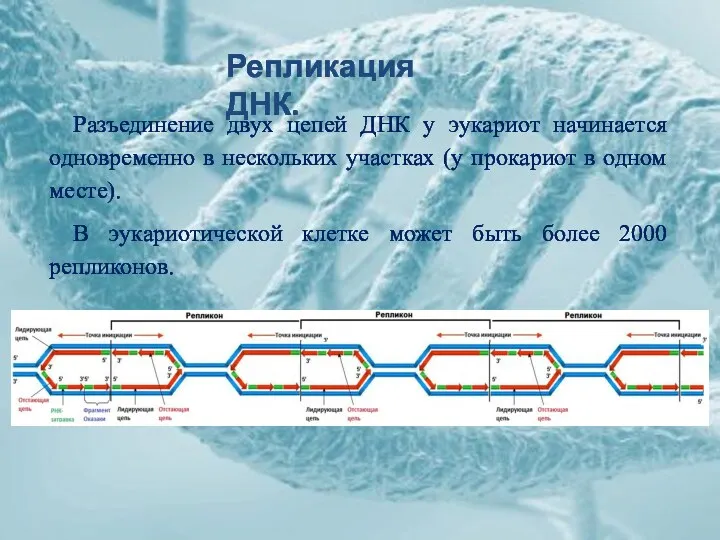
Репликация ДНК.
Разъединение двух цепей ДНК у эукариот начинается одновременно в
Репликация ДНК.
Разъединение двух цепей ДНК у эукариот начинается одновременно в
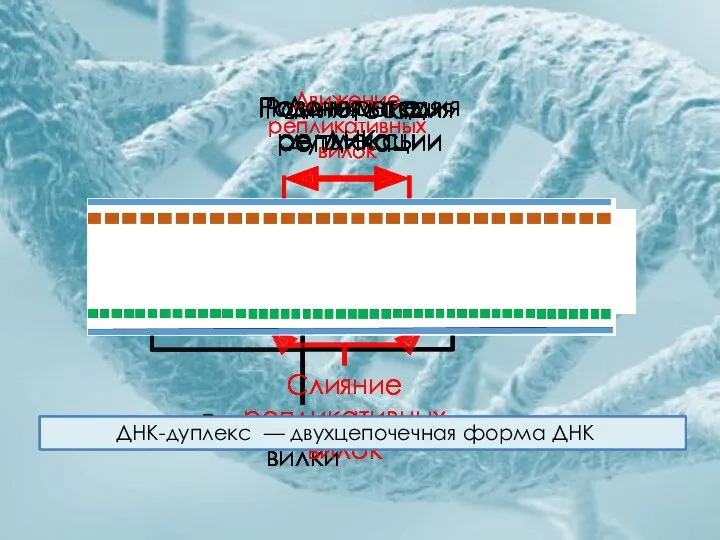
Родительская ДНК
Ранняя стадия репликации
Репликативные вилки
Движение репликативных вилок
Слияние репликативных вилок
Поздняя стадия
Родительская ДНК
Ранняя стадия репликации
Репликативные вилки
Движение репликативных вилок
Слияние репликативных вилок
Поздняя стадия
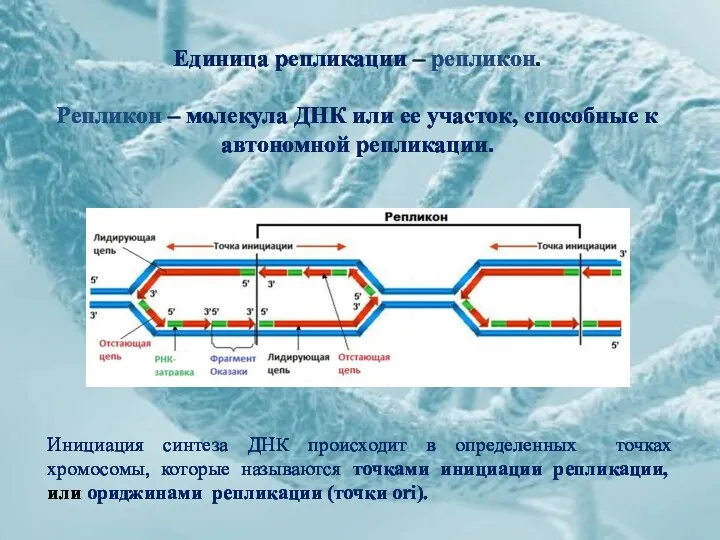
Единица репликации – репликон.
Репликон – молекула ДНК или ее участок, способные
Единица репликации – репликон. Репликон – молекула ДНК или ее участок, способные
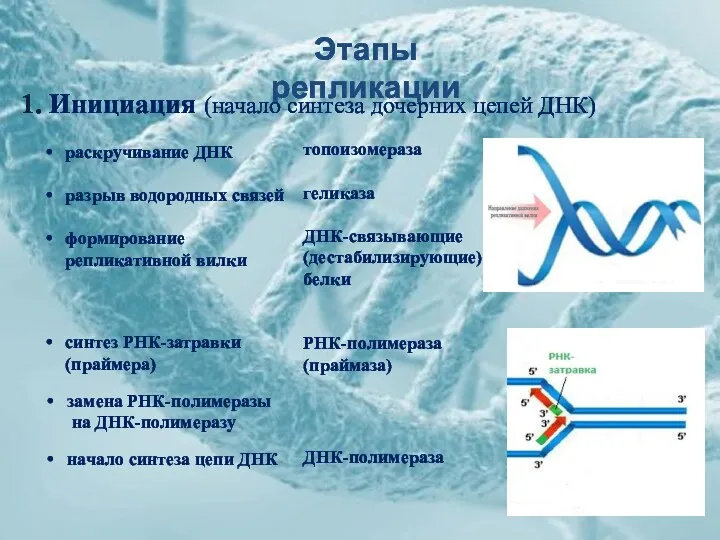
Этапы репликации
1. Инициация (начало синтеза дочерних цепей ДНК)
раскручивание ДНК
разрыв водородных связей
формирование
Этапы репликации
1. Инициация (начало синтеза дочерних цепей ДНК)
раскручивание ДНК
разрыв водородных связей
формирование
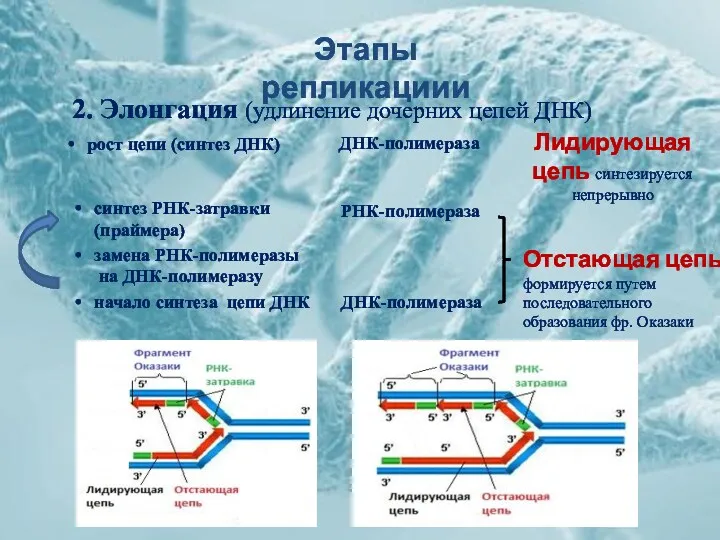
Этапы репликациии
2. Элонгация (удлинение дочерних цепей ДНК)
рост цепи (синтез ДНК)
синтез РНК-затравки
Этапы репликациии
2. Элонгация (удлинение дочерних цепей ДНК)
рост цепи (синтез ДНК)
синтез РНК-затравки
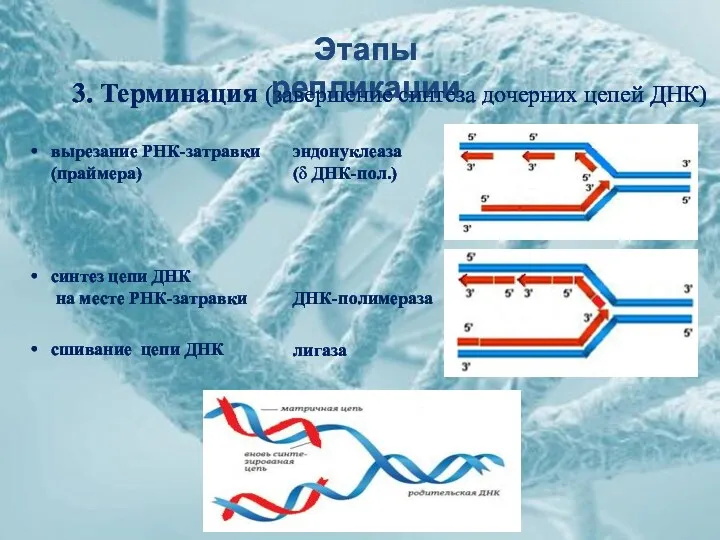
Этапы репликации
3. Терминация (завершение синтеза дочерних цепей ДНК)
вырезание РНК-затравки (праймера)
синтез цепи
Этапы репликации
3. Терминация (завершение синтеза дочерних цепей ДНК)
вырезание РНК-затравки (праймера)
синтез цепи
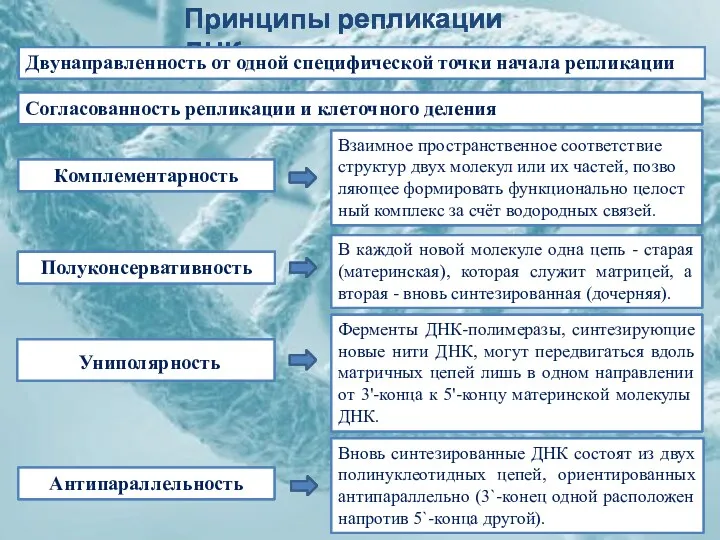
Взаимное пространственное соответствие структур двух молекул или их частей, позволяющее формировать функционально
Взаимное пространственное соответствие структур двух молекул или их частей, позволяющее формировать функционально

Транскрипция
Транскрипция осуществляется при помощи
фермента РНК-полимеразы.
Транскрипция – биосинтез одноцепочечной молекулы РНК
Транскрипция
Транскрипция осуществляется при помощи
фермента РНК-полимеразы.
Транскрипция – биосинтез одноцепочечной молекулы РНК
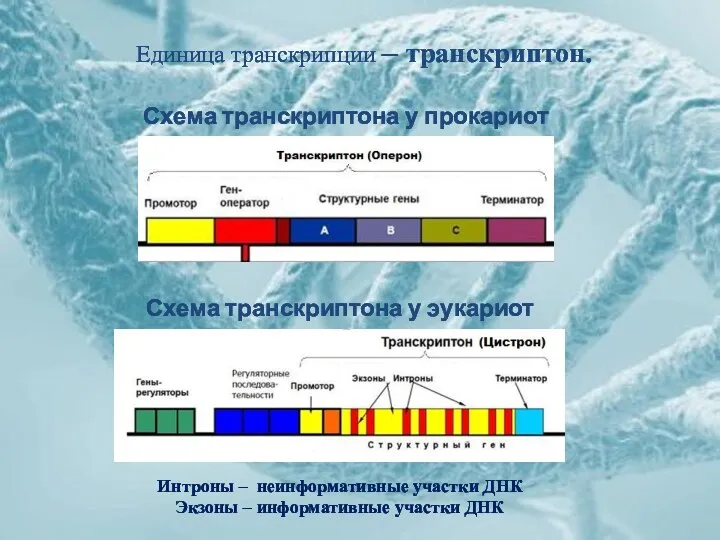
Единица транскрипции – транскриптон.
Схема транскриптона у прокариот
Схема транскриптона у эукариот
Интроны
Единица транскрипции – транскриптон.
Схема транскриптона у прокариот
Схема транскриптона у эукариот
Интроны
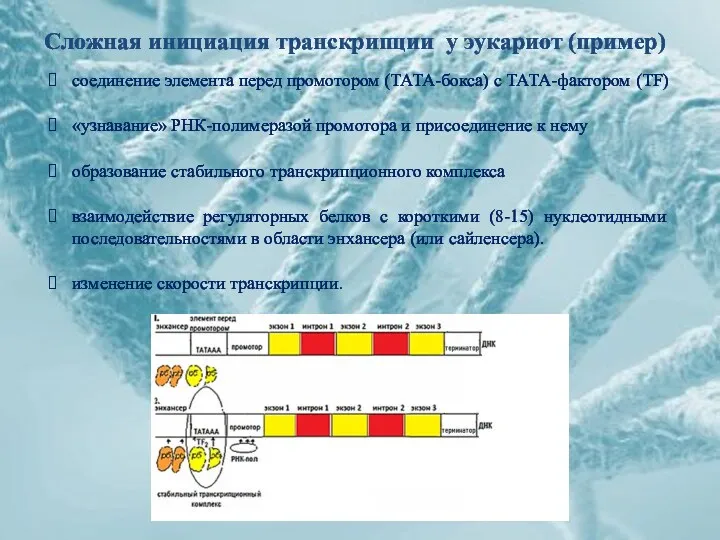
Сложная инициация транскрипции у эукариот (пример)
соединение элемента перед промотором (ТАТА-бокса) с
Сложная инициация транскрипции у эукариот (пример)
соединение элемента перед промотором (ТАТА-бокса) с
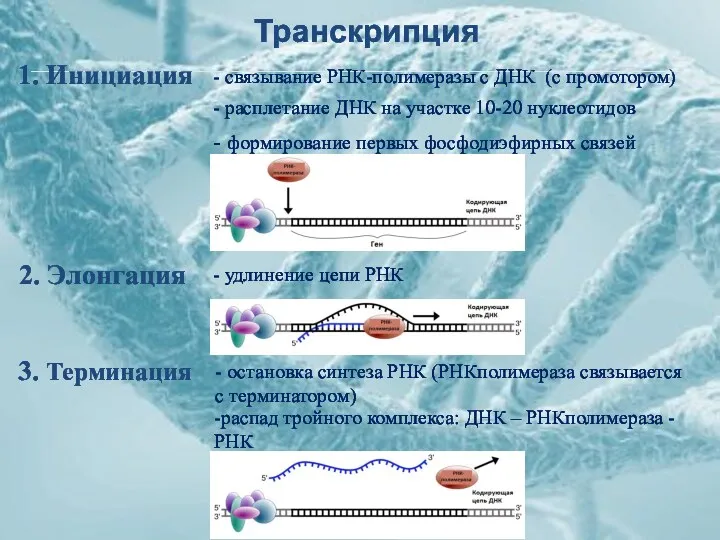
Транскрипция
1. Инициация
2. Элонгация
3. Терминация
- расплетание ДНК на участке 10-20 нуклеотидов
-
Транскрипция
1. Инициация
2. Элонгация
3. Терминация
- расплетание ДНК на участке 10-20 нуклеотидов
-
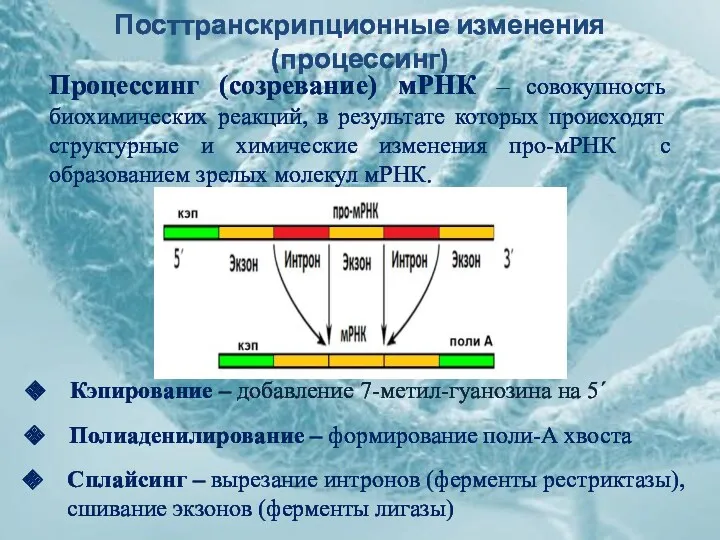
Посттранскрипционные изменения (процессинг)
Процессинг (созревание) мРНК – совокупность биохимических реакций, в результате
Посттранскрипционные изменения (процессинг)
Процессинг (созревание) мРНК – совокупность биохимических реакций, в результате

Генетический код
Генетический код – это способ записи генетической информации о структуре
Генетический код
Генетический код – это способ записи генетической информации о структуре
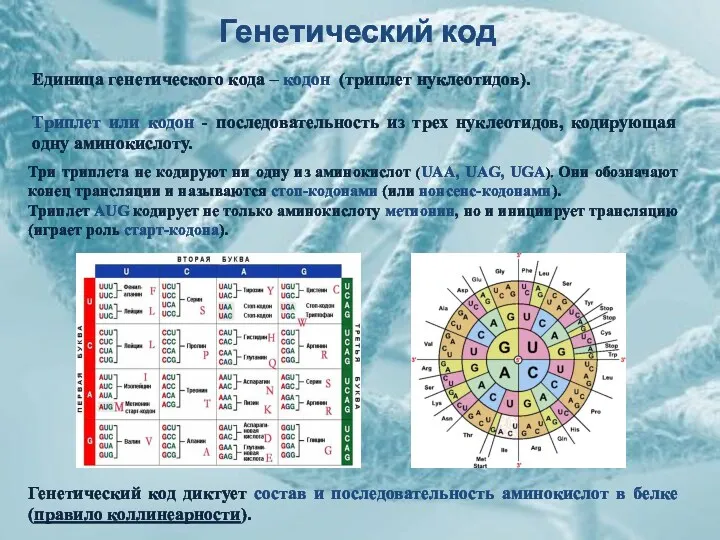
Генетический код
Единица генетического кода – кодон (триплет нуклеотидов).
Триплет или кодон -
Генетический код
Единица генетического кода – кодон (триплет нуклеотидов).
Триплет или кодон -

Свойства генетического кода:
помехоустойчивость – отношение числа консервативных замен к числу радикальных замен.
универсальность
Свойства генетического кода:
помехоустойчивость – отношение числа консервативных замен к числу радикальных замен.
универсальность

Трансляция (синтез белка)
Состоит из двух стадий:
Активирование аминокислоты.
Присоединение аминокислоты к tРНК
Трансляция (синтез белка)
Состоит из двух стадий:
Активирование аминокислоты.
Присоединение аминокислоты к tРНК
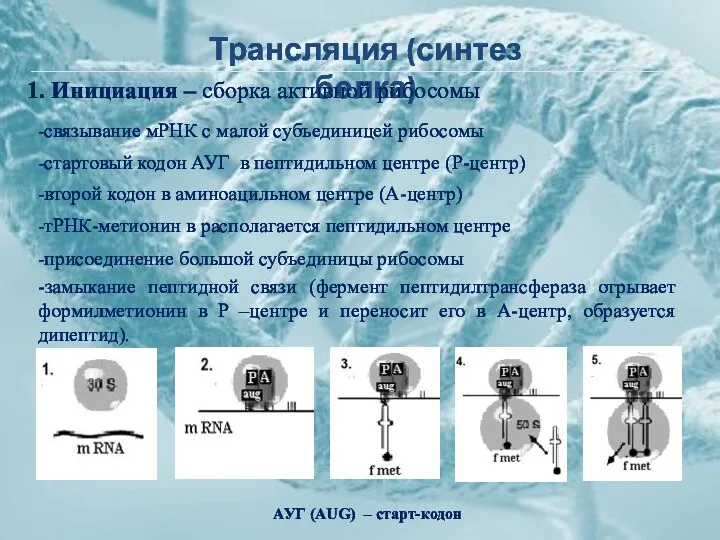
Трансляция (синтез белка)
1. Инициация – сборка активной рибосомы
-связывание мРНК с малой
Трансляция (синтез белка)
1. Инициация – сборка активной рибосомы
-связывание мРНК с малой
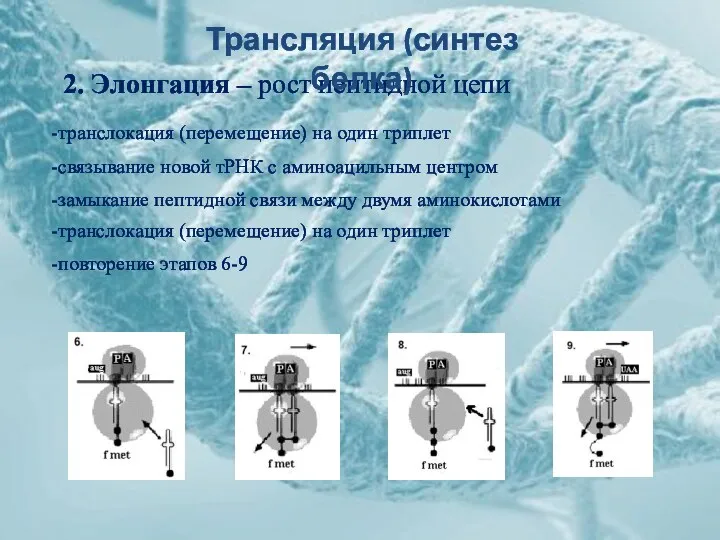
2. Элонгация – рост пептидной цепи
-связывание новой тРНК с аминоацильным
2. Элонгация – рост пептидной цепи
-связывание новой тРНК с аминоацильным
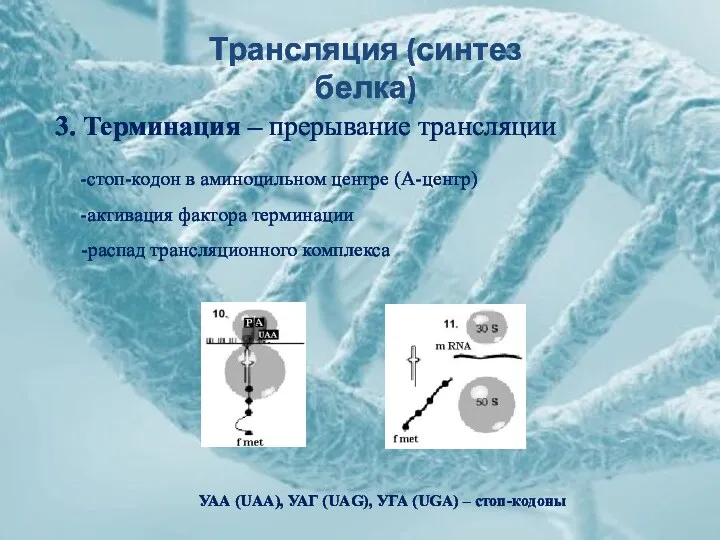
УАА (UAA), УАГ (UAG), УГА (UGA) – стоп-кодоны
3. Терминация – прерывание
УАА (UAA), УАГ (UAG), УГА (UGA) – стоп-кодоны
3. Терминация – прерывание

Передача генетической информации ретровируса.
Обратная транскрипция.
Передача генетической информации ретровируса.
Обратная транскрипция.
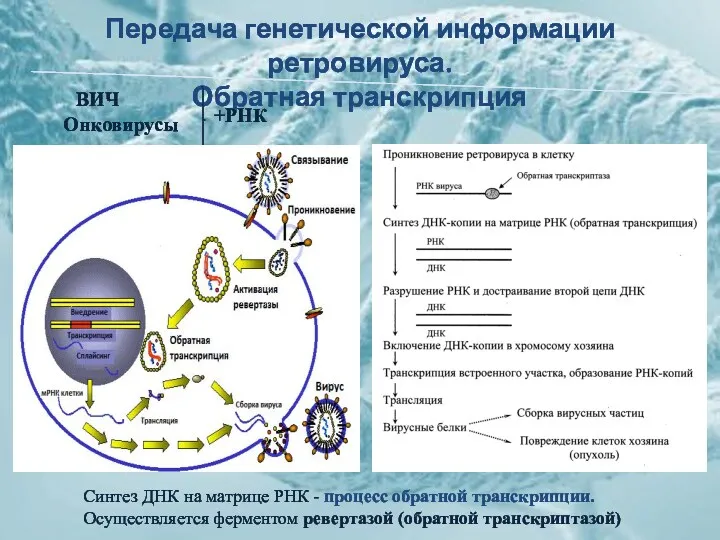
Передача генетической информации ретровируса.
Обратная транскрипция
ВИЧ
Онковирусы
+РНК
Синтез ДНК на матрице РНК - процесс
Передача генетической информации ретровируса.
Обратная транскрипция
ВИЧ
Онковирусы
+РНК
Синтез ДНК на матрице РНК - процесс

Открытие явления обратной транскрипции сыграло большую роль в развитии генной инженерии,
Открытие явления обратной транскрипции сыграло большую роль в развитии генной инженерии,

Репарация -процесс устранения повреждений нуклеотидной последовательности ДНК.
Восстановления структуры ДНК осуществляются
Репарация -процесс устранения повреждений нуклеотидной последовательности ДНК.
Восстановления структуры ДНК осуществляются
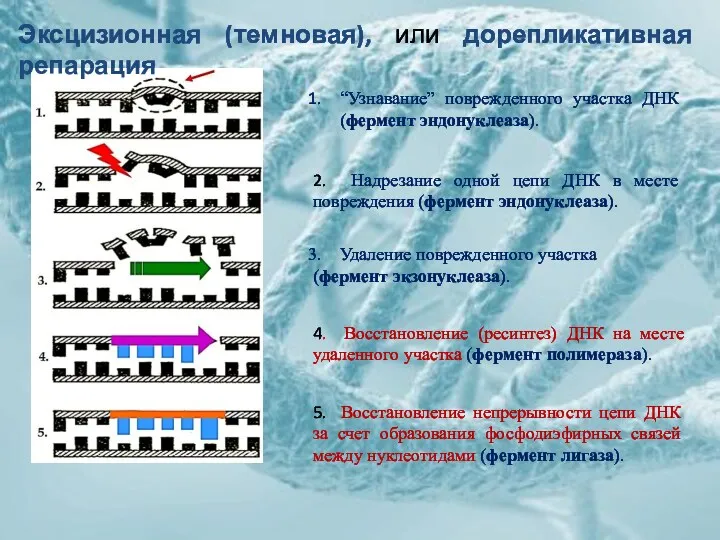
“Узнавание” поврежденного участка ДНК (фермент эндонуклеаза).
2. Надрезание одной цепи ДНК в
“Узнавание” поврежденного участка ДНК (фермент эндонуклеаза).
2. Надрезание одной цепи ДНК в
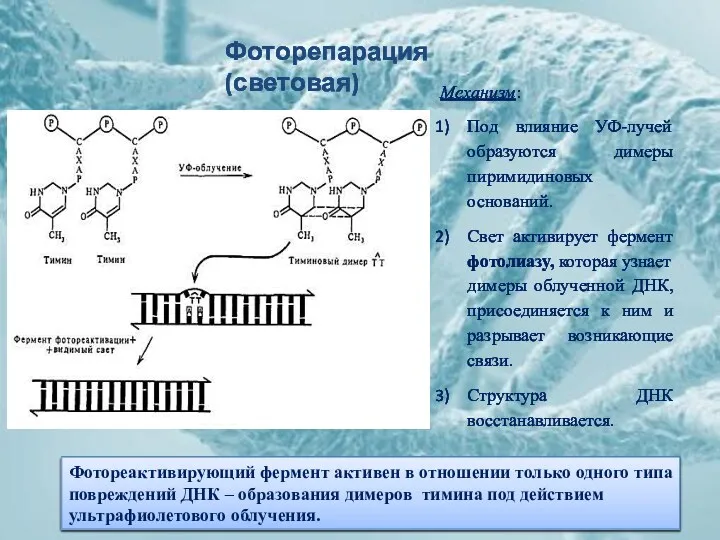
Фоторепарация (световая)
Механизм:
Под влияние УФ-лучей образуются димеры пиримидиновых оснований.
Свет активирует фермент
Фоторепарация (световая)
Механизм:
Под влияние УФ-лучей образуются димеры пиримидиновых оснований.
Свет активирует фермент
 Значення та охорона членистоногих
Значення та охорона членистоногих Особенности строения прокариотической клетки
Особенности строения прокариотической клетки Симбиотические организмы лишайники
Симбиотические организмы лишайники Вода, її роль у життєдіяльності організмів
Вода, її роль у життєдіяльності організмів Прокариотты және эукариотты өсімдіктер клеткасы
Прокариотты және эукариотты өсімдіктер клеткасы Мы в ответе за тех, кого приручили
Мы в ответе за тех, кого приручили Внешнее строение листа
Внешнее строение листа Строение, виды и значение вирусов. Прионы. Вироиды
Строение, виды и значение вирусов. Прионы. Вироиды Покормите птиц 2020. Эколого-культурная акция
Покормите птиц 2020. Эколого-культурная акция Гигиена органов пищеварения. Предупреждение желудочно–кишечных инфекций
Гигиена органов пищеварения. Предупреждение желудочно–кишечных инфекций Ткани животных
Ткани животных Простые и сложные соцветия
Простые и сложные соцветия Дәріс №2. Жасушалық технологияның негізі– жасуша өсіндісі
Дәріс №2. Жасушалық технологияның негізі– жасуша өсіндісі Введение генов в клетки растений - основные способы
Введение генов в клетки растений - основные способы Воздействие человека и его деятельности на животных
Воздействие человека и его деятельности на животных Строение, размножение и развитие рыб
Строение, размножение и развитие рыб Функциональная анатомия мышц конечностей
Функциональная анатомия мышц конечностей Физиология пищеварения
Физиология пищеварения Семейство зайцев. Школьная научно практическая конференция
Семейство зайцев. Школьная научно практическая конференция Развитие и устойчивость экосистем
Развитие и устойчивость экосистем Регуляция кровообращения
Регуляция кровообращения Способи годівлі птахів
Способи годівлі птахів Методы биологических исследований
Методы биологических исследований Промежуточные филаменты
Промежуточные филаменты Ақтөбеде сирек кездесетін өсімдіктер
Ақтөбеде сирек кездесетін өсімдіктер Систематика класса Птицы
Систематика класса Птицы 20231221_baranova_gmo
20231221_baranova_gmo Экология и природопользование. Экосистемы
Экология и природопользование. Экосистемы