- Ізоляція та видоутворення

Содержание
- 2. Каспар Баугін (1560-1624) У 1623 році запропонував бінарну номенклатуру Становлення наукового розуміння поняття виду Джон Рей
- 3. Карл Лінней 1707-1778 Шведський натураліст К.Лінней вперше надав поняттю вид біологічного змісту. Він застосував його для
- 4. Самка та самець крякви (Anas platyrhynchos) Для визначення видів Лінней застосовував типологічний підхід: Віднесення особини до
- 5. Ж.-Б. Ламарк (1744-1829) Чарльз Дарвин (1809-1882) Вид без єволюції чи еволюція без виду? Розвиток уявлень про
- 6. Е. Майр - один з засновників біологічної концепції виду Виникнення синтетичної теорії еволюції дозволило відійти від
- 7. Наприкінці ХІХ ст. розрізняли 12 видів антилоп-оріксів Зараз всі вони віднесені до одного виду: Oryx garella
- 8. М.І.Вавілов: “Ліннеєвський вид – це відокремлена складна, рухома морфологічна та фізіологічна система, пов’язана в своєму генезі
- 9. Критерії виду Первинні критерії виду Похідні критерії виду
- 10. Типи видів Глухар (Tetrao urogallus) - прикліад монотипічного виду Досить легко визначити межі виду, коли ми
- 11. Політипічні види Підвиди жовтої трясогузки та комплекс форм великих білоголових мартинів (чайок)
- 12. Форми у великої синиці Parus major: 1) Parus major major; 2) Parus major minor; 3) Parus
- 13. Проблемні види або напіввиди Хатній та чорногрудий горобець начебто два окремих види, проте між ними у
- 14. Чорна та сіра ворони існують черезполосно по всій теріторії Євразії, проте в зонах контактів регулярно утворюються
- 15. Поліморфні види У богомолів існують дві морфи: зелена і бура. Перша менш помітна на зелених частинах
- 16. «Невдале» видоутворення У білого гуся (Anser caerulescens) дві кольорові морфи: біла і синя.
- 17. Види-двійники Клести (шишкарі) ялиночник та сосновник (Loxia curvirostra та Loxia pytyopsittacus). Миші домова та курганчикова (Mus
- 18. Ізоляція та її механізми
- 19. Докопулятивні механізми ізоляції (механізми, що запобігають міжвидовому схрещуванню)
- 20. Здатність відрізняти своїх та чужих - одна з базових властивостей живих організмів Відрізнити своїх можна за:
- 21. Етологічна ізоляція Звичайна чайка (Мартин звичайний) Мала чайка (Мартин малий)
- 22. Uca tetragonon Uca perplexa Два види крабів мешкають відповідно у нижній та верхній частині літоралі. Цієї
- 23. Різні види жужелець (Carabus sp.) різняться за складом феромонів та будовою статевих органів. Різні види ропух
- 24. Відбір на посилення репродуктивної ізоляції іноді призводить до того, що особи близких видів з області спільного
- 26. Післякопулятивні механізми ізоляції (механізми, що знижують успішність міжвидового схрещування)
- 27. Безплідні гібриди Мул (вислюк та самиця коня ) Лошак (кінь та самиця вислюка) Хонорік (тхір та
- 28. Фертильні гібриди Нар (бактріан та дромадер) Лігр (лев та тигриця) Тумак (зайці-біляк та русак) Вовкособ (вовк
- 29. Кряква та шилохвіст у природі не гібридизують, хоч і гніздяться пліч-о-пліч. Проте у неволі вони утворюють
- 30. «Сексуальний паразитизм» або «гаметні злодії» Rana ridibunda 2n=26 (RR) Rana essonae 2n=26 (LL) Rana kl. esculenta
- 31. В деяких місцях на Далекому Сході основну масу популяцій складають гібриди благородного і плямистого оленей При
- 32. Способи видоутворення Алопатричне (географічне) Симпатричне Викликане екологічними причинами Викликане хромосомними перебудовами Інфекційне видоутворення
- 33. Алопатричне (а) та симпатричне (б) видоутворення
- 34. Видоутворення в умовах островних екосистем Адаптивна радіація гавайських квіткарок - приклад видоутворення, викликаного екологічними причинами (способом
- 35. Симпатричне видоутворення в умовах озерної екосистеми: риби-барбуси озера Тана (Ефіопія). Генералізована форма Їсть комах та планктон
- 36. Хромосомне видоутворення Робертсонівський процес - процес злиття двох акроцентричних хромосом в одну метацентричну або дисоціація метацентрика
- 37. Інфекційне видоутворення Бактерії Wolbachia - у цитоплазмі яйцеклітини комахи-хазяїна Бактерії Wolbachia, Spiroplasma та Cardinium здатні створювати
- 38. Модель ключ-замок. А,Б - сперматогенез інфікованих самців. Зигота розвивається лише якщо самиця теж уражена (Д,Е). Перебудови
- 39. Хромосомне видоутворення в лабораторії.
- 40. Ресинтез існуючих видів
- 41. Алича (2п=16) Терен (2п=32) Слива (2п=48)
- 42. Bufo viridis (2п=22) Bufo danatensis (4п=44) Східна Європа Південний Казахстан, Туркменія
- 43. Глід Чорника Яблуня (з 1647р. ) Rhagoletis pomonella Diachasma alloeum Ланцюгова реакція видоутворення (за А.А. Forbes
- 44. Глодова мухи Rhagoletis розмножуються виключно не глоді, яблучні – на яблуці. Личинки Rhagoletis pomonella у яблуці
- 45. Симпатричне видоутворення в лабораторії Anthriscus nemorosa Chaerophillum bulbosum Chaerophillum maculatum Dysaphys anthrisci 8-10 поколінь Dysaphys sp.
- 46. Симпатричне видоутворення у лабораторії Досліди Diane M.B. Dodd на Drosophila pseudoobscura Крохмаль Мальтоза
- 47. Симпатричне видоутворення у природі: молюски Аральського моря
- 48. Cerastoderma isthmicum - намулоїд (вихідна форма - 1) та Cerastoderma sp. - фільтруюча форма (2) Спеціогенез
- 49. Форми видоутворення в часі
- 51. Скачать презентацию

Каспар Баугін
(1560-1624)
У 1623 році запропонував бінарну номенклатуру
Становлення наукового розуміння поняття виду
Джон
Каспар Баугін
(1560-1624)
У 1623 році запропонував бінарну номенклатуру
Становлення наукового розуміння поняття виду
Джон

Карл Лінней
1707-1778
Шведський натураліст К.Лінней вперше надав поняттю вид біологічного змісту. Він
Карл Лінней
1707-1778
Шведський натураліст К.Лінней вперше надав поняттю вид біологічного змісту. Він

Самка та самець крякви (Anas platyrhynchos)
Для визначення видів Лінней застосовував типологічний
Самка та самець крякви (Anas platyrhynchos)
Для визначення видів Лінней застосовував типологічний

Ж.-Б. Ламарк (1744-1829)
Чарльз Дарвин (1809-1882)
Вид без єволюції чи еволюція без виду?
Розвиток
Ж.-Б. Ламарк (1744-1829)
Чарльз Дарвин (1809-1882)
Вид без єволюції чи еволюція без виду?
Розвиток

Е. Майр - один з засновників біологічної концепції виду
Виникнення синтетичної теорії
Е. Майр - один з засновників біологічної концепції виду
Виникнення синтетичної теорії

Наприкінці ХІХ ст. розрізняли 12 видів антилоп-оріксів
Зараз всі вони віднесені до
Наприкінці ХІХ ст. розрізняли 12 видів антилоп-оріксів
Зараз всі вони віднесені до

М.І.Вавілов: “Ліннеєвський вид – це відокремлена складна, рухома морфологічна та фізіологічна
М.І.Вавілов: “Ліннеєвський вид – це відокремлена складна, рухома морфологічна та фізіологічна
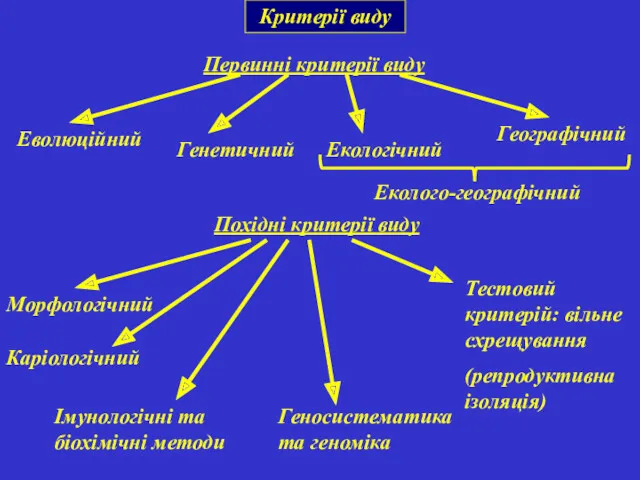
Критерії виду
Первинні критерії виду
Похідні критерії виду
Критерії виду
Первинні критерії виду
Похідні критерії виду

Типи видів
Глухар (Tetrao urogallus) - прикліад монотипічного виду
Досить легко визначити межі
Типи видів
Глухар (Tetrao urogallus) - прикліад монотипічного виду
Досить легко визначити межі
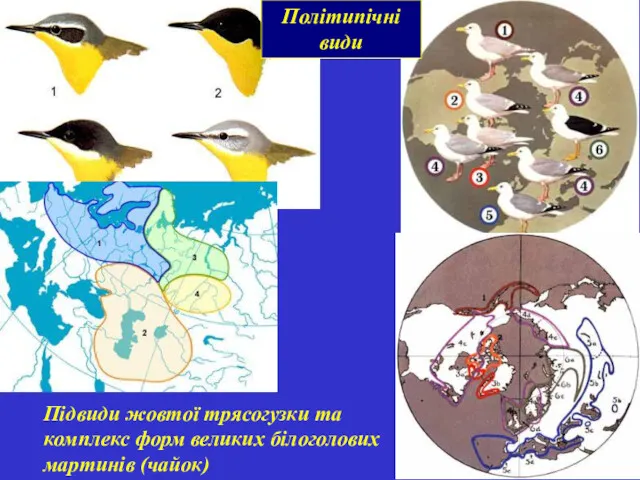
Політипічні види
Підвиди жовтої трясогузки та комплекс форм великих білоголових мартинів (чайок)
Політипічні види
Підвиди жовтої трясогузки та комплекс форм великих білоголових мартинів (чайок)
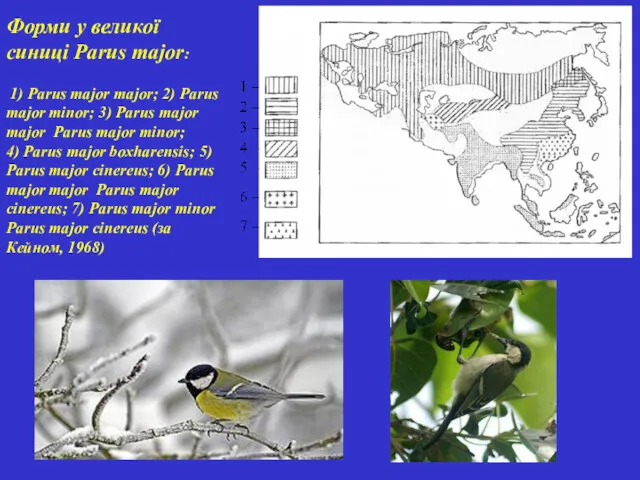
Форми у великої синиці Parus major:
1) Parus major major; 2)
Форми у великої синиці Parus major:
1) Parus major major; 2)

Проблемні види або напіввиди
Хатній та чорногрудий горобець начебто два окремих види,
Проблемні види або напіввиди
Хатній та чорногрудий горобець начебто два окремих види,
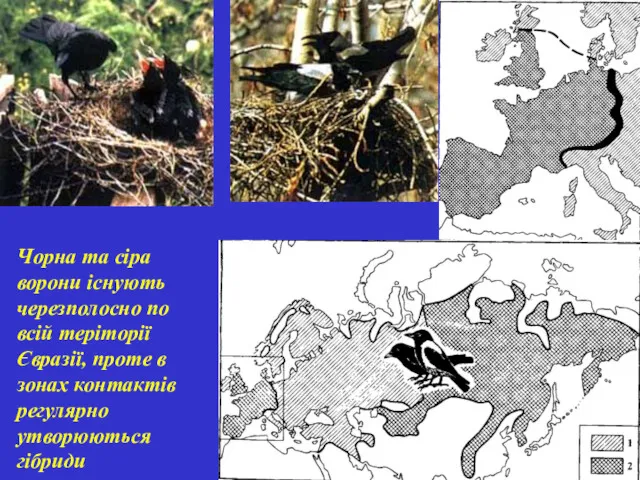
Чорна та сіра ворони існують черезполосно по всій теріторії Євразії, проте
Чорна та сіра ворони існують черезполосно по всій теріторії Євразії, проте

Поліморфні види
У богомолів існують дві морфи: зелена і бура. Перша менш
Поліморфні види
У богомолів існують дві морфи: зелена і бура. Перша менш

«Невдале» видоутворення
У білого гуся (Anser caerulescens) дві кольорові морфи: біла і
«Невдале» видоутворення
У білого гуся (Anser caerulescens) дві кольорові морфи: біла і

Види-двійники
Клести (шишкарі) ялиночник та сосновник (Loxia curvirostra та Loxia pytyopsittacus).
Миші домова
Види-двійники
Клести (шишкарі) ялиночник та сосновник (Loxia curvirostra та Loxia pytyopsittacus).
Миші домова

Ізоляція та її механізми
Ізоляція та її механізми
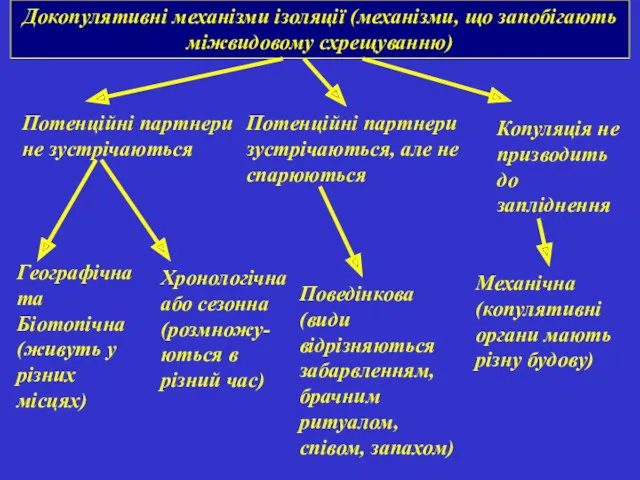
Докопулятивні механізми ізоляції (механізми, що запобігають міжвидовому схрещуванню)
Докопулятивні механізми ізоляції (механізми, що запобігають міжвидовому схрещуванню)

Здатність відрізняти своїх та чужих - одна з базових властивостей живих
Здатність відрізняти своїх та чужих - одна з базових властивостей живих

Етологічна ізоляція
Звичайна чайка (Мартин звичайний)
Мала чайка (Мартин малий)
Етологічна ізоляція
Звичайна чайка (Мартин звичайний)
Мала чайка (Мартин малий)

Uca tetragonon
Uca perplexa
Два види крабів мешкають відповідно у нижній та верхній
Uca tetragonon
Uca perplexa
Два види крабів мешкають відповідно у нижній та верхній

Різні види жужелець (Carabus sp.) різняться за складом феромонів та будовою
Різні види жужелець (Carabus sp.) різняться за складом феромонів та будовою

Відбір на посилення репродуктивної ізоляції іноді призводить до того, що особи
Відбір на посилення репродуктивної ізоляції іноді призводить до того, що особи
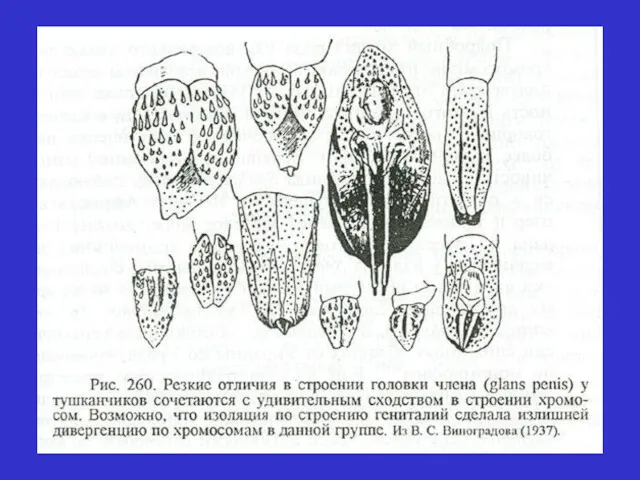
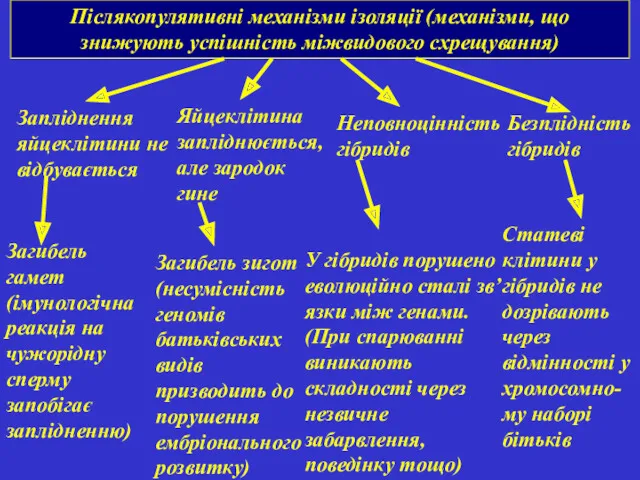
Післякопулятивні механізми ізоляції (механізми, що знижують успішність міжвидового схрещування)
Післякопулятивні механізми ізоляції (механізми, що знижують успішність міжвидового схрещування)

Безплідні гібриди
Мул (вислюк та самиця коня )
Лошак (кінь та самиця вислюка)
Хонорік
Безплідні гібриди
Мул (вислюк та самиця коня )
Лошак (кінь та самиця вислюка)
Хонорік

Фертильні гібриди
Нар (бактріан та дромадер)
Лігр (лев та тигриця)
Тумак (зайці-біляк та русак)
Вовкособ
Фертильні гібриди
Нар (бактріан та дромадер)
Лігр (лев та тигриця)
Тумак (зайці-біляк та русак)
Вовкособ

Кряква та шилохвіст у природі не гібридизують, хоч і гніздяться пліч-о-пліч.
Кряква та шилохвіст у природі не гібридизують, хоч і гніздяться пліч-о-пліч.
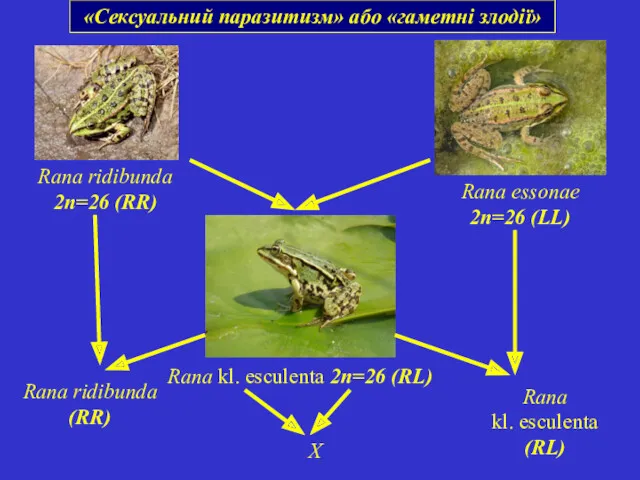
«Сексуальний паразитизм» або «гаметні злодії»
Rana ridibunda 2n=26 (RR)
Rana essonae 2n=26 (LL)
Rana
«Сексуальний паразитизм» або «гаметні злодії»
Rana ridibunda 2n=26 (RR)
Rana essonae 2n=26 (LL)
Rana

В деяких місцях на Далекому Сході основну масу популяцій складають гібриди
В деяких місцях на Далекому Сході основну масу популяцій складають гібриди
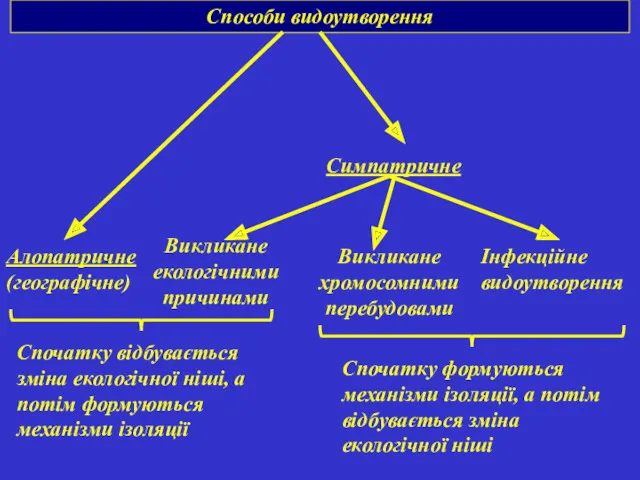
Способи видоутворення
Алопатричне (географічне)
Симпатричне
Викликане екологічними причинами
Викликане хромосомними перебудовами
Інфекційне видоутворення
Способи видоутворення
Алопатричне (географічне)
Симпатричне
Викликане екологічними причинами
Викликане хромосомними перебудовами
Інфекційне видоутворення
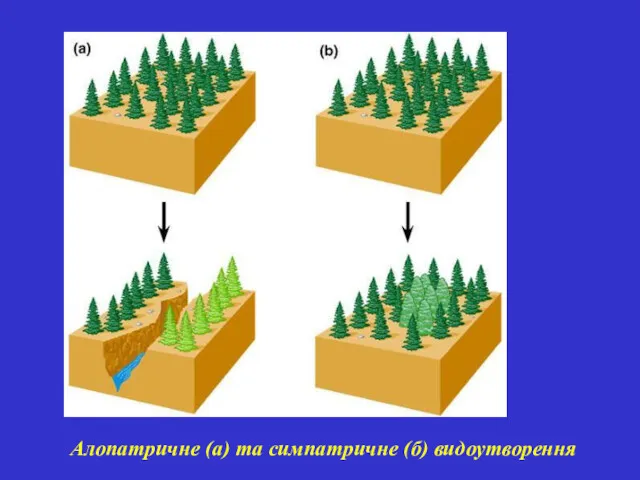
Алопатричне (а) та симпатричне (б) видоутворення
Алопатричне (а) та симпатричне (б) видоутворення

Видоутворення в умовах островних екосистем
Адаптивна радіація гавайських квіткарок - приклад видоутворення,
Видоутворення в умовах островних екосистем
Адаптивна радіація гавайських квіткарок - приклад видоутворення,

Симпатричне видоутворення в умовах озерної екосистеми: риби-барбуси озера Тана (Ефіопія).
Генералізована форма
Їсть
Симпатричне видоутворення в умовах озерної екосистеми: риби-барбуси озера Тана (Ефіопія).
Генералізована форма
Їсть

Хромосомне видоутворення
Робертсонівський процес - процес злиття двох акроцентричних хромосом в одну
Хромосомне видоутворення
Робертсонівський процес - процес злиття двох акроцентричних хромосом в одну
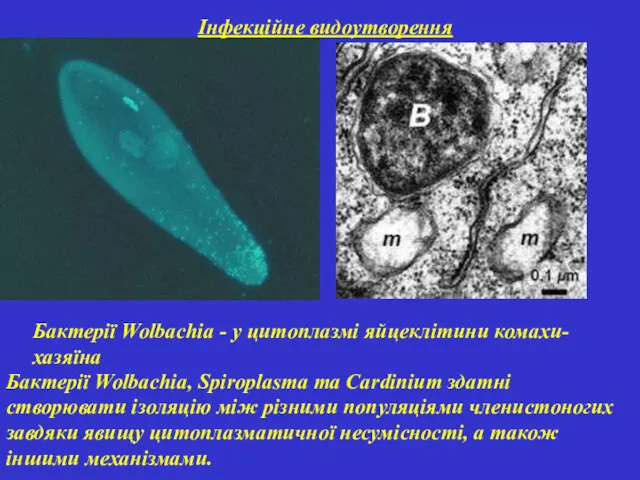
Інфекційне видоутворення
Бактерії Wolbachia - у цитоплазмі яйцеклітини комахи-хазяїна
Бактерії Wolbachia, Spiroplasma
Інфекційне видоутворення
Бактерії Wolbachia - у цитоплазмі яйцеклітини комахи-хазяїна
Бактерії Wolbachia, Spiroplasma
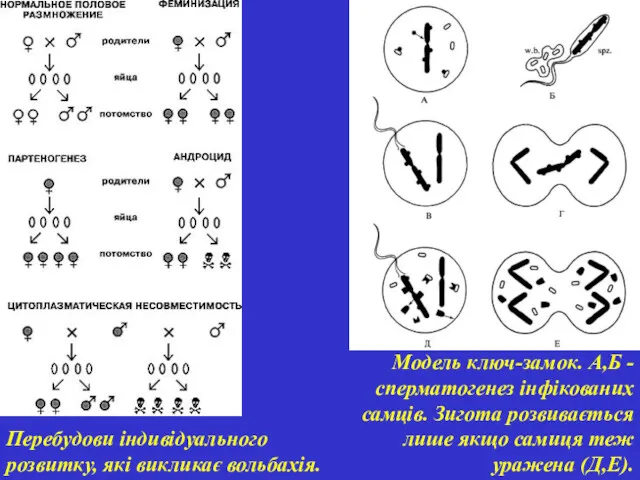
Модель ключ-замок. А,Б - сперматогенез інфікованих самців. Зигота розвивається лише якщо
Модель ключ-замок. А,Б - сперматогенез інфікованих самців. Зигота розвивається лише якщо
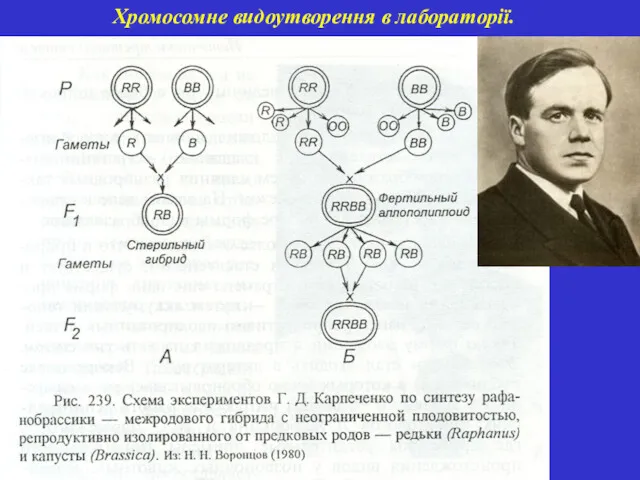
Хромосомне видоутворення в лабораторії.
Хромосомне видоутворення в лабораторії.
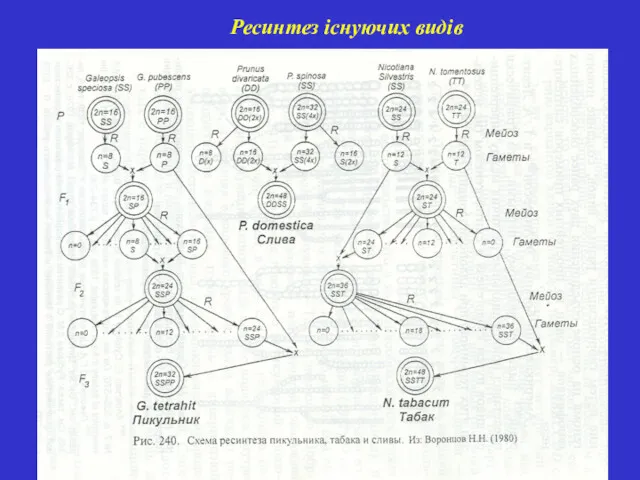
Ресинтез існуючих видів
Ресинтез існуючих видів
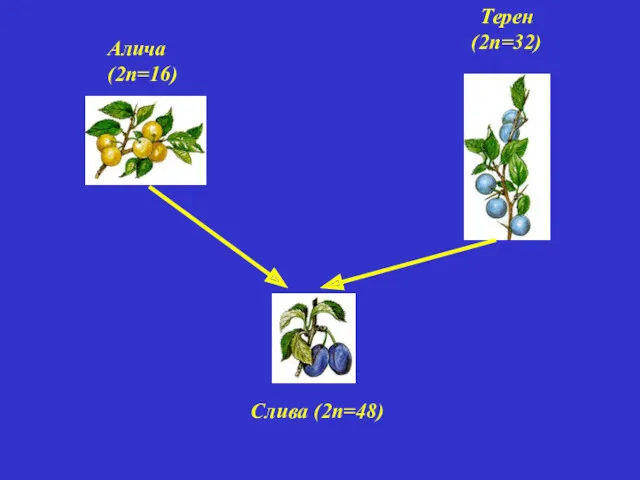
Алича (2п=16)
Терен (2п=32)
Слива (2п=48)
Алича (2п=16)
Терен (2п=32)
Слива (2п=48)
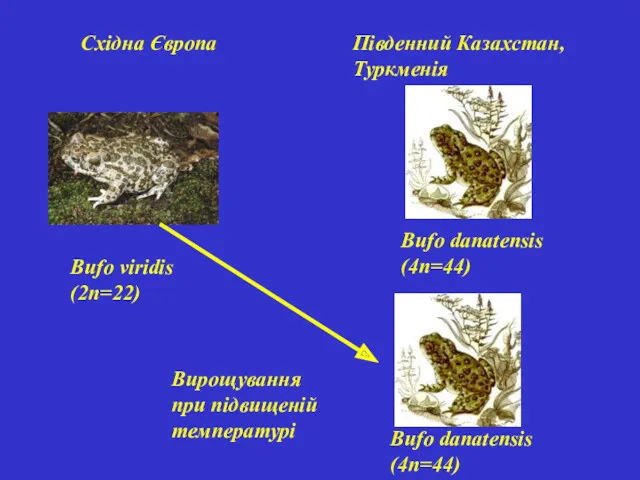
Bufo viridis (2п=22)
Bufo danatensis (4п=44)
Східна Європа
Південний Казахстан, Туркменія
Bufo viridis (2п=22)
Bufo danatensis (4п=44)
Східна Європа
Південний Казахстан, Туркменія

Глід
Чорника
Яблуня (з 1647р. )
Rhagoletis pomonella
Diachasma alloeum
Ланцюгова реакція видоутворення (за А.А. Forbes
Глід
Чорника
Яблуня (з 1647р. )
Rhagoletis pomonella
Diachasma alloeum
Ланцюгова реакція видоутворення (за А.А. Forbes

Глодова мухи Rhagoletis розмножуються виключно не глоді, яблучні – на яблуці.
Личинки
Глодова мухи Rhagoletis розмножуються виключно не глоді, яблучні – на яблуці.
Личинки
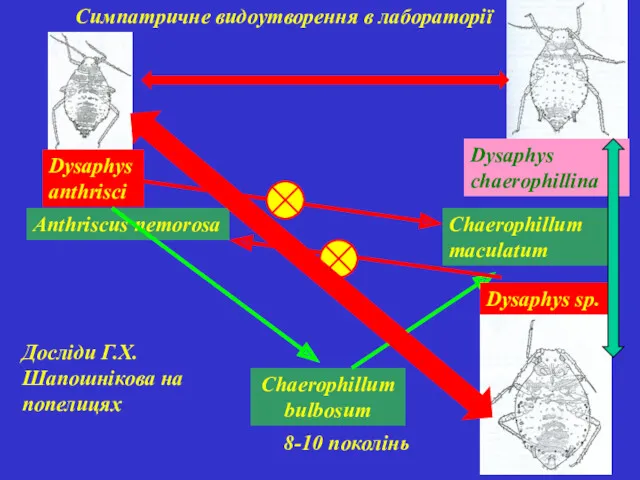
Симпатричне видоутворення в лабораторії
Anthriscus nemorosa
Chaerophillum bulbosum
Chaerophillum maculatum
Dysaphys anthrisci
8-10 поколінь
Dysaphys sp.
Dysaphys chaerophillina
Досліди
Симпатричне видоутворення в лабораторії
Anthriscus nemorosa
Chaerophillum bulbosum
Chaerophillum maculatum
Dysaphys anthrisci
8-10 поколінь
Dysaphys sp.
Dysaphys chaerophillina
Досліди
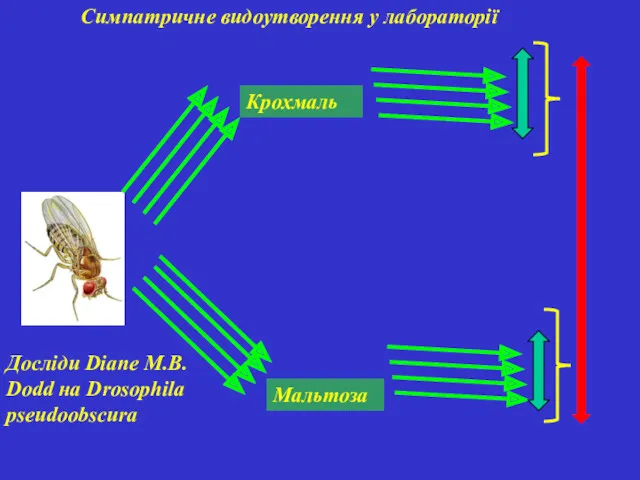
Симпатричне видоутворення у лабораторії
Досліди Diane M.B. Dodd на Drosophila pseudoobscura
Крохмаль
Мальтоза
Симпатричне видоутворення у лабораторії
Досліди Diane M.B. Dodd на Drosophila pseudoobscura
Крохмаль
Мальтоза
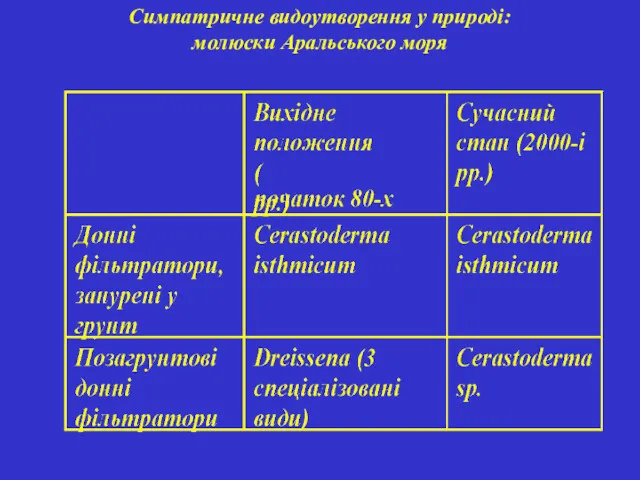
Симпатричне видоутворення у природі: молюски Аральського моря
Симпатричне видоутворення у природі: молюски Аральського моря
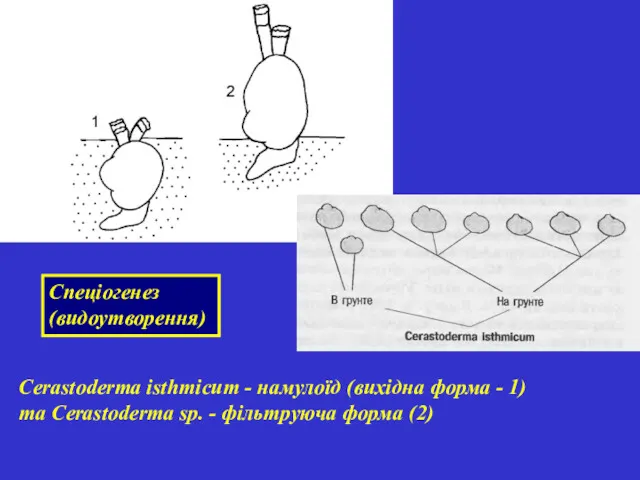
Cerastoderma isthmicum - намулоїд (вихідна форма - 1) та Cerastoderma sp.
Cerastoderma isthmicum - намулоїд (вихідна форма - 1) та Cerastoderma sp.
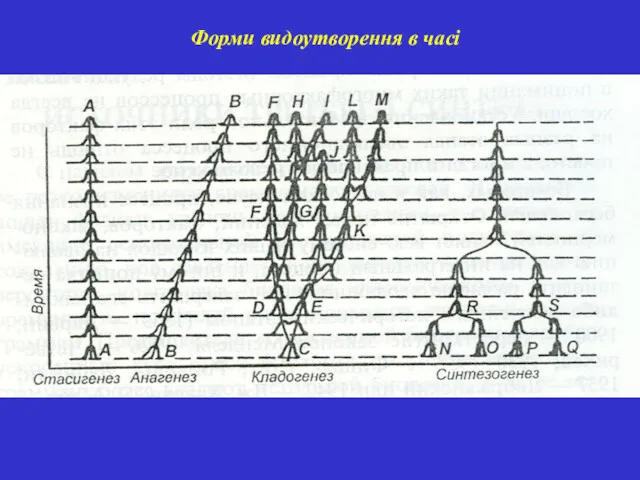
Форми видоутворення в часі
Форми видоутворення в часі
 Отряд Приматы
Отряд Приматы Кость как орган. Скелет туловища
Кость как орган. Скелет туловища Современная эмбриология
Современная эмбриология Ағзаларды клондау
Ағзаларды клондау презентация условия прорастания семян
презентация условия прорастания семян Життя – це спосіб існування білкових тіл
Життя – це спосіб існування білкових тіл Деревья и кустарники
Деревья и кустарники Animals in danger. About other animals
Animals in danger. About other animals Розмноження птахів
Розмноження птахів лекция 2. Направления биотехнологии
лекция 2. Направления биотехнологии Эбриогинез органов мочевыделительной системы
Эбриогинез органов мочевыделительной системы Плодовые растения
Плодовые растения Растения, занесенные в Красную книгу Ульяновской области
Растения, занесенные в Красную книгу Ульяновской области Лес - наше богатство
Лес - наше богатство Метаболизм: пластический и энергетический обмен
Метаболизм: пластический и энергетический обмен Строение раковины моллюска
Строение раковины моллюска Грибы съедобные и несъедобные. Правила сбора грибов
Грибы съедобные и несъедобные. Правила сбора грибов Тамырлар бойымен қан қозғалысынның жалпы физика математикалық заңдылықтары
Тамырлар бойымен қан қозғалысынның жалпы физика математикалық заңдылықтары Заповедник Брянский лес
Заповедник Брянский лес Популяционная генетика
Популяционная генетика Природные сообщества. Биогеоценоз и экосистемы
Природные сообщества. Биогеоценоз и экосистемы Школа Карполова - 2018. Занятие 4. Май-июнь. Активность карпа, ПВА, Термоклин, Маркер
Школа Карполова - 2018. Занятие 4. Май-июнь. Активность карпа, ПВА, Термоклин, Маркер Презентация к урокуДеление клетки. Митоз
Презентация к урокуДеление клетки. Митоз Нейрон. Нейронная теория. Центральная нервная система (ЦНС)
Нейрон. Нейронная теория. Центральная нервная система (ЦНС) Тема: Деление клетки. Митоз
Тема: Деление клетки. Митоз Тип Хордовые
Тип Хордовые Сказочный гербарий
Сказочный гербарий Физиология человека
Физиология человека