- Видовое разнообразие экосистем

Содержание
- 2. . Выделяют три основных уровня биологического разнообразия: I. Молекулярно-генетический, т.е. разнообразие генов в организмах и синтезируемых
- 3. Наука, изучающая закономерности распространении видов живых организмов на нашей планете, называется биогеографией. Она подразделяется на зоогеографию
- 4. Примером реликтов среди животных является пресмыкающееся гаттерия Sphenodon punctatus. Этот единственный современный вид отряда клювоголовых, известный
- 5. Однако некоторые виды, возникшие в предыдущие геологические эпохи, сохранились в неизменном состоянии до наших дней в
- 6. Напротив, виды, роды, семейства или отряды, встречающиеся на обширных территориях, называются космополитичными. К ним относится, например,
- 7. Видовой состав флоры каждой территории предложено выражать в в виде формулы: F = f1 + f2
- 8. Существует ряд способов оценки сходства или различия видового состава разных сообществ. В их числе – различные
- 9. Однако оба эти коэффициента учитывают только число видов в сравниваемых биоценозах, вне зависимости от их численности.
- 10. Пример. В первом биоценозе имеются 4 вида (А, B, C и D), а во втором –
- 11. Биогеографическое районирование земной суши. Вся разных регионов вся территория земной суши подразделяется на ряд крупных зоогеографических
- 12. Большинство исследователей выделяет четыре фаунистических царства. Арктогейское, или Голантарктическое царство; Палеогейское, или Палеотропическое царство; Неогейское, или
- 14. Большинство геоботаников подразделяет территорию земной суши на шесть флористических царств. Четыре из них – Голарктическое, Палеотропическое,
- 16. Разные биоценозы могут значительно различаться как по общей видовой насыщенности, так и по числу видов в
- 17. Видовое разнообразие цветковых растений в сообществах некоторых наземных экосистем
- 18. Основы островной биогеографии Основоположниками этого направления являются П.Макартур и Уилсон. В этой теории остров рассматривается не
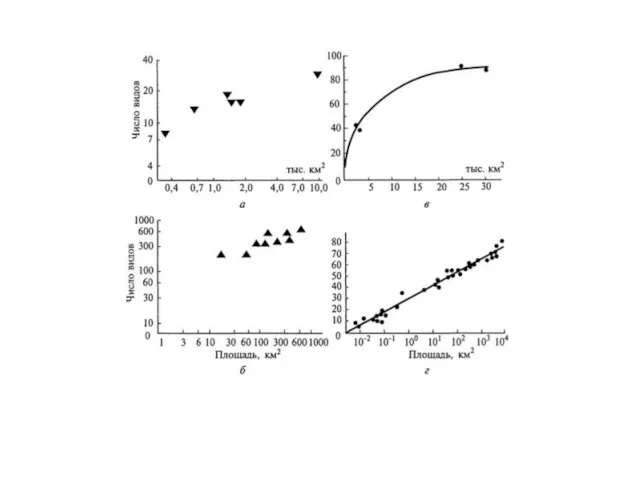
- 19. Связь между N и S (км2) для географических островов имеет параболический характер: N = aSb Значения
- 21. Значения коэффициента b в уравнениях для отдельных групп растений и животных, населяющих разные системы островов
- 22. Скорость заселения острова новыми видами снижается с возрастанием числа видов на острове. По мере того, как
- 23. Допустим, скорость иммиграции, или количество новых видов заселяющих остров в единицу времени (λS) линейно снижается с
- 24. Структура доминирования в биоценозах Практически не встречается сообществ, в которых всех численность составляющих его видов была
- 25. Однако из большого числа видов, входящих в сообщество, лишь немногие оказывают заметное влияние на его функционирование.
- 26. Значимость отдельных видов в сообществе (Рi) рассчитывается следующим образом: Рi = xi/Σxi, где xi - доля
- 27. Другим показателем значимости отдельных видов в сообществе является индекс Симпсона (С). Он обычно рассчитывается по значениям
- 28. Значения индекса Симпсона (C’) для сообществ из более 150 наземных и водных экосистем, различались от 0,05
- 29. В группу с высоким разнообразием сообщества из многих естественных экосистем, не получающих извне энергии и биогенных
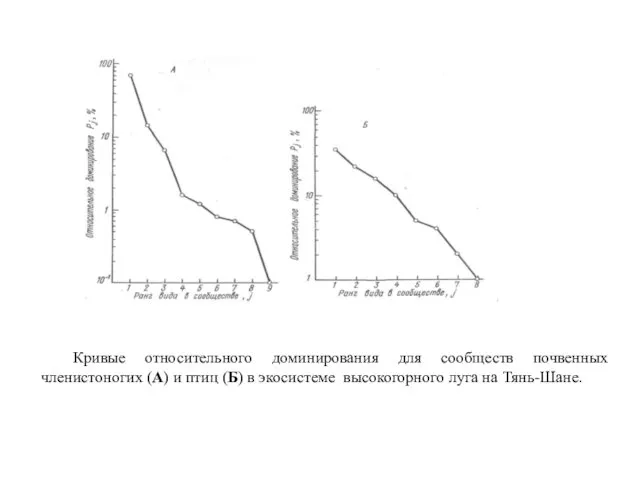
- 30. Кривые относительного доминирования для сообществ почвенных членистоногих (А) и птиц (Б) в экосистеме высокогорного луга на
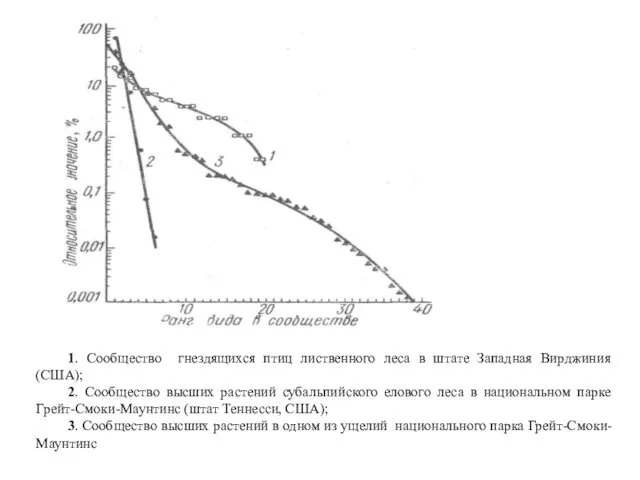
- 31. 1. Сообщество гнездящихся птиц лиственного леса в штате Западная Вирджиния (США); 2. Сообщество высших растений субальпийского
- 32. Кривая 1 отвечает ситуации, когда виды-члены сообщества находятся в сильной зависимости от некоторого ресурса, распределенного вдоль
- 33. Линия 2, представляющая собой прямую, соответствует случаю т. наз. «геометрического распределения». Она отражает ситуацию, когда значимость
- 34. Таким образом, значимость каждого вида последующего ранга в сообществе снижается в геометрической прогрессии. Поэтому прямая 2,
- 35. При всех типах распределения видов в сообществе можно выделить один – два (редко больше) доминирующих вида,
- 37. Скачать презентацию

. Выделяют три основных уровня биологического разнообразия:
I. Молекулярно-генетический, т.е. разнообразие генов
. Выделяют три основных уровня биологического разнообразия:
I. Молекулярно-генетический, т.е. разнообразие генов

Наука, изучающая закономерности распространении видов живых организмов на нашей планете,
Наука, изучающая закономерности распространении видов живых организмов на нашей планете,

Примером реликтов среди животных является пресмыкающееся гаттерия Sphenodon punctatus. Этот
Примером реликтов среди животных является пресмыкающееся гаттерия Sphenodon punctatus. Этот

Однако некоторые виды, возникшие в предыдущие геологические эпохи, сохранились в
Однако некоторые виды, возникшие в предыдущие геологические эпохи, сохранились в

Напротив, виды, роды, семейства или отряды, встречающиеся на обширных территориях,
Напротив, виды, роды, семейства или отряды, встречающиеся на обширных территориях,

Видовой состав флоры каждой территории предложено выражать в в виде формулы:
F
Видовой состав флоры каждой территории предложено выражать в в виде формулы:
F

Существует ряд способов оценки сходства или различия видового состава разных сообществ.
Существует ряд способов оценки сходства или различия видового состава разных сообществ.

Однако оба эти коэффициента учитывают только число видов в сравниваемых биоценозах,
Однако оба эти коэффициента учитывают только число видов в сравниваемых биоценозах,

Пример. В первом биоценозе имеются 4 вида (А, B, C и
Пример. В первом биоценозе имеются 4 вида (А, B, C и

Биогеографическое районирование земной суши.
Вся разных регионов вся территория земной
Биогеографическое районирование земной суши.
Вся разных регионов вся территория земной

Большинство исследователей выделяет четыре фаунистических царства.
Арктогейское, или Голантарктическое царство;
Большинство исследователей выделяет четыре фаунистических царства.
Арктогейское, или Голантарктическое царство;
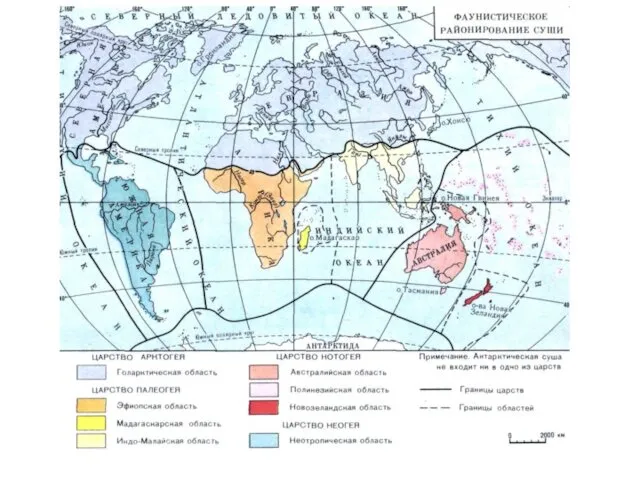

Большинство геоботаников подразделяет территорию земной суши на шесть флористических царств.
Большинство геоботаников подразделяет территорию земной суши на шесть флористических царств.


Разные биоценозы могут значительно различаться как по общей видовой насыщенности, так
Разные биоценозы могут значительно различаться как по общей видовой насыщенности, так
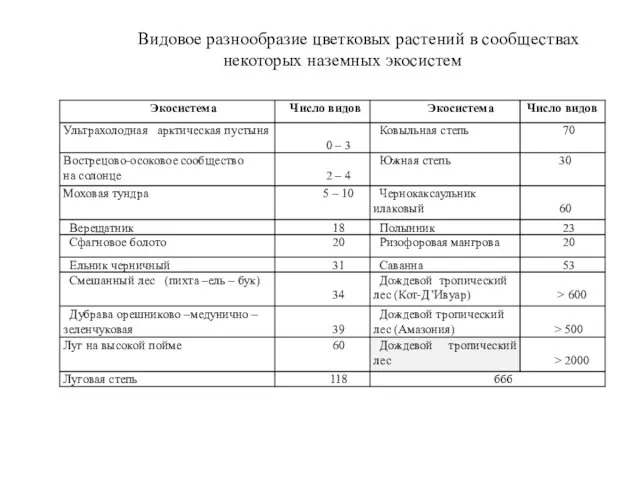
Видовое разнообразие цветковых растений в сообществах некоторых наземных экосистем
Видовое разнообразие цветковых растений в сообществах некоторых наземных экосистем

Основы островной биогеографии
Основоположниками этого направления являются П.Макартур и
Основы островной биогеографии
Основоположниками этого направления являются П.Макартур и

Связь между N и S (км2) для географических островов имеет параболический
Связь между N и S (км2) для географических островов имеет параболический
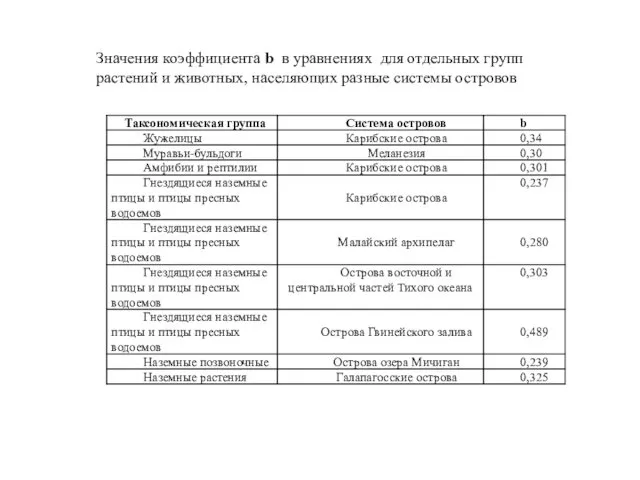
Значения коэффициента b в уравнениях для отдельных групп растений и животных,
Значения коэффициента b в уравнениях для отдельных групп растений и животных,

Скорость заселения острова новыми видами снижается с возрастанием числа видов
Скорость заселения острова новыми видами снижается с возрастанием числа видов

Допустим, скорость иммиграции, или количество новых видов заселяющих остров в
Допустим, скорость иммиграции, или количество новых видов заселяющих остров в

Структура доминирования в биоценозах
Практически не встречается сообществ, в которых всех
Структура доминирования в биоценозах
Практически не встречается сообществ, в которых всех

Однако из большого числа видов, входящих в сообщество, лишь немногие
Однако из большого числа видов, входящих в сообщество, лишь немногие

Значимость отдельных видов в сообществе (Рi) рассчитывается следующим образом:
Рi =
Значимость отдельных видов в сообществе (Рi) рассчитывается следующим образом:
Рi =

Другим показателем значимости отдельных видов в сообществе является индекс Симпсона
Другим показателем значимости отдельных видов в сообществе является индекс Симпсона

Значения индекса Симпсона (C’) для сообществ из более 150 наземных и
Значения индекса Симпсона (C’) для сообществ из более 150 наземных и

В группу с высоким разнообразием сообщества из многих естественных экосистем,
В группу с высоким разнообразием сообщества из многих естественных экосистем,
Кривые относительного доминирования для сообществ почвенных членистоногих (А) и птиц (Б)
Кривые относительного доминирования для сообществ почвенных членистоногих (А) и птиц (Б)
1. Сообщество гнездящихся птиц лиственного леса в штате Западная Вирджиния (США);
1. Сообщество гнездящихся птиц лиственного леса в штате Западная Вирджиния (США);

Кривая 1 отвечает ситуации, когда виды-члены сообщества находятся в сильной зависимости
Кривая 1 отвечает ситуации, когда виды-члены сообщества находятся в сильной зависимости

Линия 2, представляющая собой прямую, соответствует случаю т. наз.
Линия 2, представляющая собой прямую, соответствует случаю т. наз.

Таким образом, значимость каждого вида последующего ранга в сообществе
Таким образом, значимость каждого вида последующего ранга в сообществе

При всех типах распределения видов в сообществе можно выделить один
При всех типах распределения видов в сообществе можно выделить один
 Перспективная система обращения с отходами в Удмуртии
Перспективная система обращения с отходами в Удмуртии Zelen' экологический ресторан
Zelen' экологический ресторан Разработка экологической тропы на примере Московского государственного историко-архитектурного музея-заповедника Коломенское
Разработка экологической тропы на примере Московского государственного историко-архитектурного музея-заповедника Коломенское Почва. Физические и химические свойства. Гигиеническое и экологическое значение
Почва. Физические и химические свойства. Гигиеническое и экологическое значение Экологические факторы
Экологические факторы Загрязнение вод - экологическая проблема современности. 10 класс
Загрязнение вод - экологическая проблема современности. 10 класс Основные виды антропогенного воздействия на почвы
Основные виды антропогенного воздействия на почвы Многозадачность экологического права. Практикум по экологии
Многозадачность экологического права. Практикум по экологии Глобальный экологический мониторинг
Глобальный экологический мониторинг Урок экологии 9 класс Возможности человечества к адаптации
Урок экологии 9 класс Возможности человечества к адаптации Нарушение экологического равновесия в местах проживания и его влияние на здоровье человека
Нарушение экологического равновесия в местах проживания и его влияние на здоровье человека Проблемы народонаселения
Проблемы народонаселения Международный день биологического разнообразия
Международный день биологического разнообразия Полезные ископаемые ХМАО
Полезные ископаемые ХМАО 22 апреля - День Земли
22 апреля - День Земли Профессиональный модуль 3. Организация использования лесов
Профессиональный модуль 3. Организация использования лесов Ecological problems
Ecological problems Методы и основы биоиндикации. Лекция № 5
Методы и основы биоиндикации. Лекция № 5 Экологическое нормирование воздействий на атмосферу
Экологическое нормирование воздействий на атмосферу Экология. Задачи экологии
Экология. Задачи экологии Разберем мусор – спасем Подмосковье
Разберем мусор – спасем Подмосковье Экологическое нормирование антропогенной нагрузки на экосистемы
Экологическое нормирование антропогенной нагрузки на экосистемы Социальная экологическая кампания Крышкин дом
Социальная экологическая кампания Крышкин дом Мұнай-газ өңдеу кешенінің қоршаған ортаға тигізетін әсерін бағалау және мұнай шламын кайта өңдеу
Мұнай-газ өңдеу кешенінің қоршаған ортаға тигізетін әсерін бағалау және мұнай шламын кайта өңдеу Модернизация системы управления твердыми бытовыми отходами Карагандинской области
Модернизация системы управления твердыми бытовыми отходами Карагандинской области Енергозбереження. Проблеми екології
Енергозбереження. Проблеми екології Исследование экологической безопасности материалов, используемых для производства мебели
Исследование экологической безопасности материалов, используемых для производства мебели Влияние окружаюшей среды на здоровье человека
Влияние окружаюшей среды на здоровье человека