- Биофизика мембранных процессов. Мембранный транспорт

Содержание
- 2. Виды мембранного транспорта (МТ) Мембранный транспорт (МТ) 1.Пассивный транспорт нейтральная 1.1.Простая диффузия ионная 1.1.1. Осмос 1.1.2.
- 3. Виды транспорта (массопереноса) через биомембрану. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 4. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Виды пассивного транспорта
- 5. Энергия пассивного транспорта создается различными градиентами: концентрационным осмотическим электрическим градиентом гидростатического давления жидкости электрохимическим (совокупность концентрационного
- 6. Виды пассивного транспорта Диффузия Осмос Фильтрация Облегченная диффузия ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 7. Диффузия – это самопроизвольный процесс проникновения вещества из области большей в область меньшей его концентрации в
- 8. Простая диффузия описывается уравнением Фика : скорость диффузии dm/dt прямо пропорциональна градиенту концентрации dc/dx, площади S,
- 9. Скорость диффузии - это количество вещества, диффундирующего в единицу времени через данную площадь. Градиент концентрации -
- 10. Осмос - движение молекулы воды через полупроницаемые мембраны из места с меньшей концентрацией растворенного вещества в
- 11. Уравнение, описывающее осмотический перенос воды. где - количество воды, проходящей через мембрану площадью S за единицу
- 12. Фильтрация – это движение раствора через поры в мембране под действием градиента гидростатического давления. Явление фильтрации
- 13. Фильтрация – движение жидкости через поры какой-либо перегородки под действием гидростатического давления. где r – радиус
- 14. Уравнение Коллендера-Бернульда описывает транспорт веществ через липидный бислой где С1 и С2 – концентрации вещества по
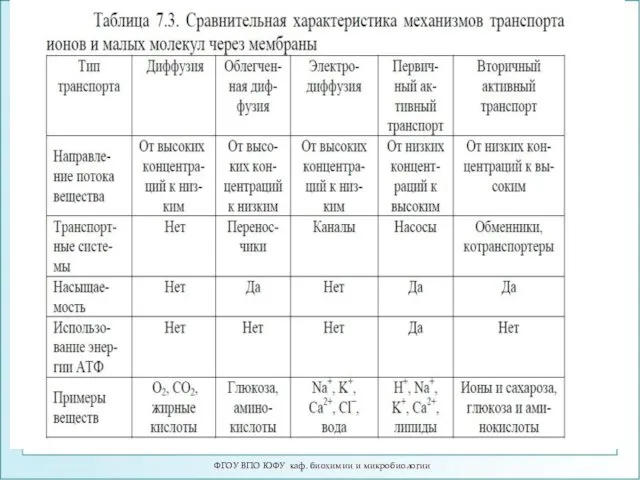
- 15. Различные виды транспорта ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Переносчик Канал
- 16. Облегчённая диффузия. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии I = Imax [C]/Kt + [C], где
- 17. Виды мембранного транспорта по направлению и количеству транспортируемых веществ ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 18. Активный транспорт. Виды активного транспорта. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Вторичный активный транспорт –
- 19. Активный транспорт: типы насосов (транспортных АТФаз) Семейство АТФаз Р-типа Семейство АТФаз F- и V-типа Суперсемейство АТФаз
- 20. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии 1. АТФ-азы Р-типа – переносчики ионов, которые обратимо фосфорилируются
- 21. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии АВС-транспортер, «выкачивающий» липофильные вещества из клетки
- 22. АТФ-синтаза/АТФ-аза: «вальсирующий» комплекс Структура: Две субъединицы: F0 и F1 F1: α3β3γδε α - 59, β -
- 23. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Механизм действия Р-гликопротеина (P-gp)
- 24. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 25. Роль онкотического и гидростатического давления в транспорте воды через мембрану ФГОУ ВПО ЮФУ каф. биохимии и
- 26. Строение Na+/K+-ATФазы ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Протомер фермента состоит из одной альфа- и
- 27. Реакционный цикл Na+/K+-ATФазы ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Шесть основных последовательньх реакций включают: 1.)
- 28. Связывание ионов Na+ и K+ в ионных центрах Na+/K+-ATФазы ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 29. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Все морские птицы (например, альбатрос) имеют ряд специальных приспособлений
- 30. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Биопотенциалы
- 31. Генерация и распространение биоэлектрических потенциалов - важнейшее физическое явление в живых клетках и тканях, которое лежит
- 32. Для возникновения биопотенциалов решающее значение имеют потенциалы, обусловленные асимметричным, неравномерным распределением ионов.
- 34. Диффузионный потенциал возникает на границе раздела двух жидких сред в результате различной подвижности ионов. 0 1
- 35. Диффузионный потенциал определяется из уравнения Гендерсона Где U – подвижность катионов V – подвижность анионов R
- 36. Мембранный потенциал возникает на границе раздела полупроницаемой мембраны, имеющей фиксированный отрицательный заряд – катионобменная мембрана. ▬
- 37. Мембранный потенциал определяют из уравнения Нернста Где R – универсальная газовая постоянная Т – абсолютная температура
- 38. Фазовые потенциалы возникают на границе раздела двух несмешивающихся фаз. нитробензол + + + + + +
- 39. Электродный потенциал возникает в результате диффузии ионов из электрода в раствор электролита. AgCl + + +
- 40. В 1902 году Бернштейном была выдвинута мембранная теория биопотенциалов. В 50-60-х годах теория была развита и
- 41. Сущность мембранной теории биопотенциалов Потенциал покоя и потенциал действия является по своей природе мембранными потенциалами, обусловленными
- 42. Мембранным потенциалом называется разность потенциалов между внутренней и наружной поверхностями мембраны.
- 43. Потенциал покоя – стационарная разность электрических потенциалов, регистрируемых между внутренней и наружной поверхностями мембраны в невозбужденном
- 44. Потенциал покоя. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 45. Потенциал покоя
- 46. При образовании потенциала покоя мембрана внутри заряжена отрицательно (за счет выхода из клетки большого количества ионов
- 47. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Потенциал покоя. В мембране открыты калиевые ионные каналы постоянного
- 48. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Нервная клетка и потенциал действия Схема распределения зарядов по
- 49. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Причины отрицательности мембранного потенциала покоя: • Покоящаяся клеточная мембрана
- 50. Уравнение Нернста для потенциала покоя
- 51. Уравнение Гольдмана для потенциала покоя
- 52. Отношение коэффициентов проницаемости для состояния покоя PК : РNa : PCl = 1 : 0,04 :
- 53. С учетом работы электрогенных ионных насосов для мембранного потенциала было получено уравнение Томаса (1972 г.) Где
- 54. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Потенциал покоя и потенциал действия
- 55. Потенциал действия (ПД) – это последовательность изменений, мембранного потенциала, которая запускается в ответ на воздействие надпороговых
- 56. Потенциалом действия называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам
- 57. Потенциал действия
- 58. При генерации потенциала действия происходит перезарядка мембраны: внутри она становится зараженной положительно (за счет входа ионов
- 59. Возбудимость – это способность клеток к быстрому ответу на раздражение, проявляющемуся через совокупность физических, физико-химических процессов
- 60. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Фазы потенциала действия
- 61. Стимуляция нервной клетки достигает порога возбудимости, необходимого для возникновения потенциала действия. Начальное изменение мембранного потенциала приводит
- 62. Поскольку деполяризация в клетке продолжается, открывается больше потенциал-зависимых К+-каналов, и К+ начинает по электрохимическому градиенту выходить
- 63. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Изменение проницаемости мембраны для ионов натрия и калия при
- 64. Физический механизм деполяризации Увеличение проницаемости для Na+ Вход Na+ в клетку Деполяризация мембраны
- 65. Отношение коэффициентов проницаемости ионов для фазы деполяризации PК : РNa : PCl = 1 : 20
- 66. Физический механизм реполяризации Выход К+ в среду Уменьшение потока Na+ в клетку Реполяризация мембраны
- 67. Формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к
- 68. МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ Е1 – потенциал покоя Е2 – мембранный потенциал при возбуждении Е3 –
- 69. Уравнение Нернста для потенциала действия
- 70. Уравнение Ходжкина-Хаксли, описывающее мембранный потенциал, который складывается из потенциала покоя и потенциала действия
- 71. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Ионные каналы
- 72. Ионный канал. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 73. Ионный транспорт ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Где z - валентность иона e -
- 74. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Вероятность перехода иона из воды в липидную фазу: Непосредственный
- 75. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Изменение энергии при переходе гидрофильного вещества через липидный бислой
- 76. Свойства ионных каналов Селективность Независимость работы отдельных каналов Дискретный характер проводимости Зависимость параметров каналов от мембранного
- 77. Свойства ионных каналов 1.Селективность – способность пропускать ионы одного типа. ИК обладают абсолютной селективностью по отношению
- 78. 2.Независимость работы отдельных каналов Прохождение тока через отдельный ионный канал не зависит от того, идет ли
- 79. 3.Дискретный характер проводимости Проводимость ионного канала дискретна, и он может находится в двух состояниях: открытом и
- 80. 4.Зависимость параметров каналов от мембранного потенциала Ионные каналы нервных волокон чувствительны к мембранному потенциалу, например К+-каналы
- 81. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии В 2003 году Нобелевская премия по химии была присуждена
- 82. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Строение потенциал-чувствительного калиевого канала
- 83. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Строение натриевого канала
- 84. Потенциал-чувствительный натриевый канал Канал состоит из четырёх похожих фрагментов-повторов (а), обозначенных I–IV. Каждый фрагмент содержит шесть
- 85. Сравнение параметров ионных каналов ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 86. Строение калиевого канала. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 87. Энергетические профили ионных каналов. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Входящий и выходящий потоки Na+
- 88. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Блокатор натриевых каналов тетродотоксин Рыба фугу — японский деликатес,
- 89. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Подобно пробке, тетродотоксин входит во внешний вестибюль натриевого канала.
- 90. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Хищный брюхоногий моллюск Conus textile и структура конотоксина. Справа
- 91. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Третий класс соединений, воздействующих на потенциал-зависимые натриевые каналы, представляет
- 92. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Местные анестетики Местные анестетики блокируют быстрый вход Na+, возникновение
- 93. Действие местных анестетиков. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 94. Химическая структура блокаторов ионных каналов. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
- 95. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Метод пэтч-кламп, разработанный Неером и Закманом, позволяет изучать отдельные
- 96. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Нобелевская премия по химии 2003 г. была вручена «за
- 97. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Аквапорины образуют гидрофильные трансмембранные каналы для переноса воды
- 98. Индуцированный ионный транспорт осуществляется ионофорами, которые делятся на 2 группы: 1. Подвижные переносчики 2. Каналообразующие агенты
- 99. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Подвижные переносчики. Перенос иона через мембрану с участием подвижного
- 100. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Подвижные переносчики. Валиномицин. Антибиотик валиномицин впервые был выделен из
- 101. Ионофоры. Валиномицин. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Молекула валиномицина (а) напоминает своей конфигурацией браслет,
- 102. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Кристаллическая структура ионофора - комплекса К+-валиномицин
- 103. Механизм работы валиномицина – малая «карусель» ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Молекула валиномицина переносит
- 104. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Скульптурная композиция комплекса К+-валиномицин перед входом в институт биоорганической
- 105. Индуцированный транспорт ионов при участии различных переносчиков I - нейтральные ионофоры (валиномицин) II - слабокислые ионофоры
- 106. Грамицидин – каналообразующий антибиотик ФГОУ ВПО ЮФУ каф. биохимии и микробиологии 1) Грамицидин сформирован 15 гидрофобными
- 107. Структура амфипатической α-спирали и расположение спиралей в мембране при формировании гидрофильного канала
- 108. Аламетицин – каналообразующий антибиотик ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Аламетицин, пептидный антибиотик с молекулярной
- 109. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Проведение потенциала действия в безмиелиновых (А) и миелиновых (Б)
- 110. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии Проведение возбуждения в нервных волокнах. А – безмиелиновое волокно
- 112. Скачать презентацию

Виды мембранного транспорта (МТ)
Мембранный транспорт (МТ)
1.Пассивный транспорт нейтральная
1.1.Простая диффузия ионная
Виды мембранного транспорта (МТ)
Мембранный транспорт (МТ)
1.Пассивный транспорт нейтральная
1.1.Простая диффузия ионная
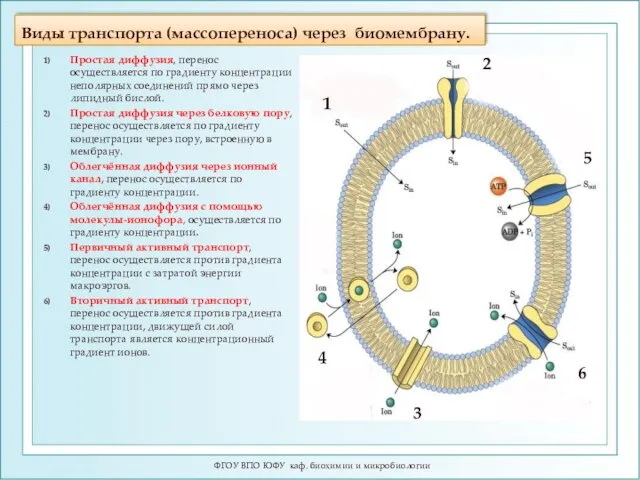
Виды транспорта (массопереноса) через биомембрану.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Виды транспорта (массопереноса) через биомембрану.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
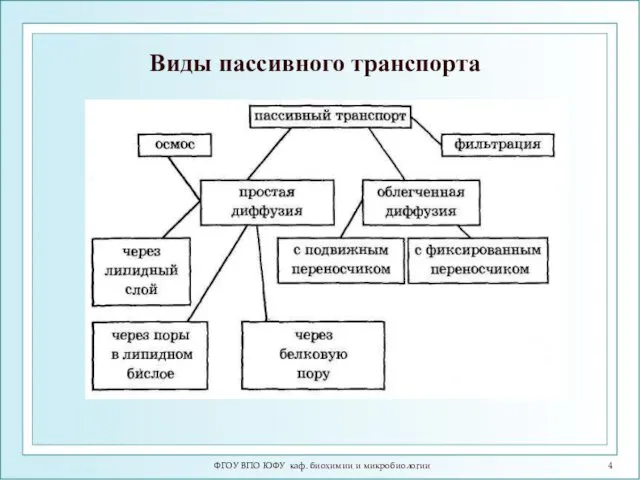
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Виды пассивного транспорта
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Виды пассивного транспорта

Энергия пассивного транспорта создается различными градиентами:
концентрационным
осмотическим
электрическим
градиентом гидростатического давления жидкости
электрохимическим (совокупность концентрационного
Энергия пассивного транспорта создается различными градиентами:
концентрационным
осмотическим
электрическим
градиентом гидростатического давления жидкости
электрохимическим (совокупность концентрационного

Виды пассивного транспорта
Диффузия
Осмос
Фильтрация
Облегченная диффузия
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Виды пассивного транспорта
Диффузия
Осмос
Фильтрация
Облегченная диффузия
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии

Диффузия –
это самопроизвольный процесс проникновения вещества из области большей в
Диффузия –
это самопроизвольный процесс проникновения вещества из области большей в

Простая диффузия описывается уравнением Фика : скорость диффузии dm/dt прямо пропорциональна
Простая диффузия описывается уравнением Фика : скорость диффузии dm/dt прямо пропорциональна

Скорость диффузии - это количество вещества, диффундирующего в единицу времени
Скорость диффузии - это количество вещества, диффундирующего в единицу времени

Осмос - движение молекулы воды через полупроницаемые мембраны из места
Осмос - движение молекулы воды через полупроницаемые мембраны из места

Уравнение, описывающее осмотический перенос воды.
где
- количество воды, проходящей через мембрану
Уравнение, описывающее осмотический перенос воды.
где
- количество воды, проходящей через мембрану

Фильтрация –
это движение раствора через поры в мембране под действием
Фильтрация – это движение раствора через поры в мембране под действием

Фильтрация –
движение жидкости через поры
какой-либо перегородки под действием гидростатического
Фильтрация – движение жидкости через поры какой-либо перегородки под действием гидростатического

Уравнение Коллендера-Бернульда описывает транспорт веществ через липидный бислой
где
С1 и С2 –
Уравнение Коллендера-Бернульда описывает транспорт веществ через липидный бислой
где
С1 и С2 –
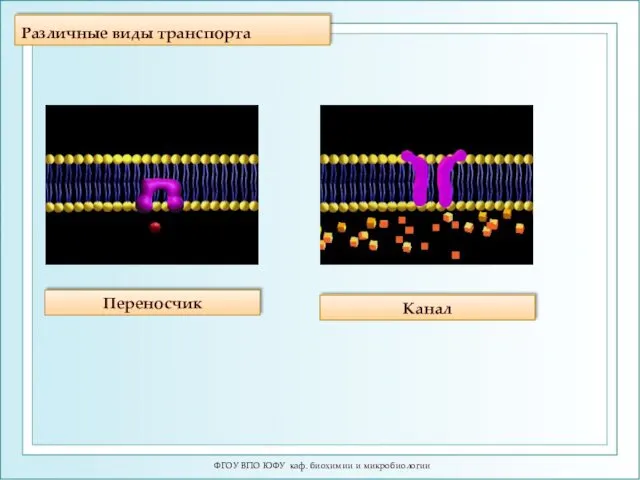
Различные виды транспорта
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Переносчик
Канал
Различные виды транспорта
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Переносчик
Канал
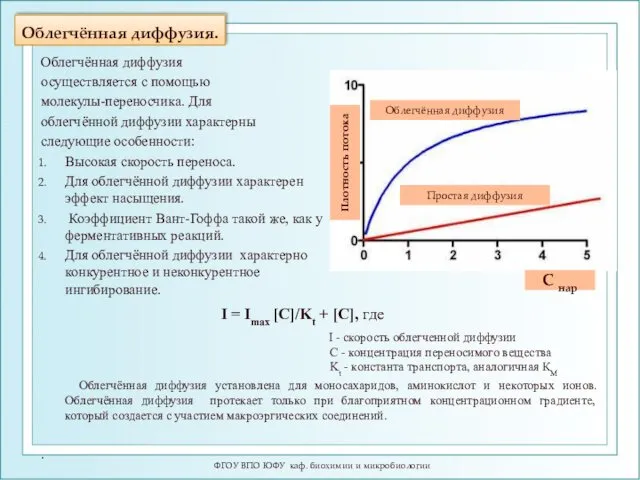
Облегчённая диффузия.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
I = Imax [C]/Kt
Облегчённая диффузия.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
I = Imax [C]/Kt
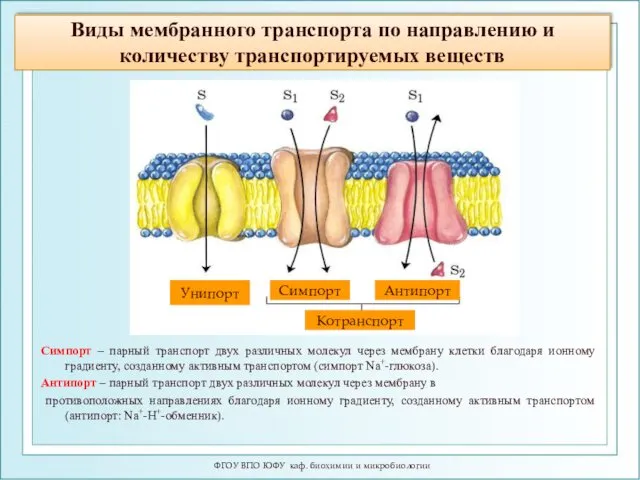
Виды мембранного транспорта по направлению и количеству транспортируемых веществ
ФГОУ ВПО ЮФУ
Виды мембранного транспорта по направлению и количеству транспортируемых веществ
ФГОУ ВПО ЮФУ
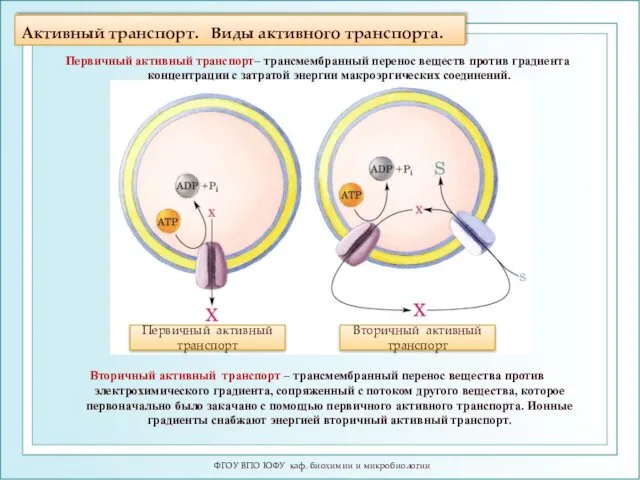
Активный транспорт. Виды активного транспорта.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Вторичный
Активный транспорт. Виды активного транспорта.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Вторичный
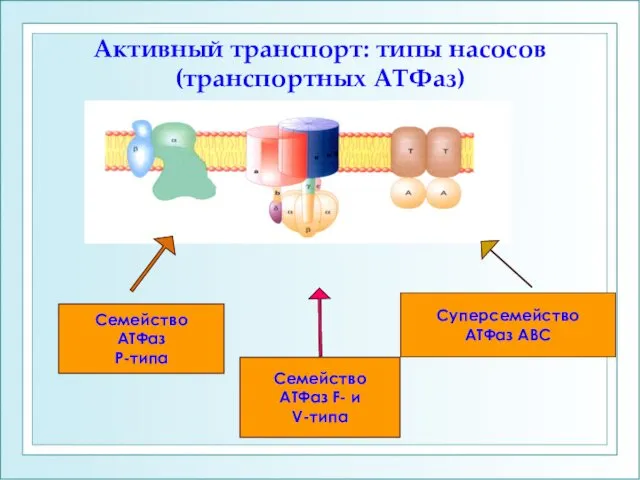
Активный транспорт: типы насосов (транспортных АТФаз)
Семейство
АТФаз
Р-типа
Семейство
АТФаз F- и
V-типа
Суперсемейство
АТФаз АВС
Активный транспорт: типы насосов (транспортных АТФаз)
Семейство
АТФаз
Р-типа
Семейство
АТФаз F- и
V-типа
Суперсемейство
АТФаз АВС

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
1. АТФ-азы Р-типа – переносчики
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
1. АТФ-азы Р-типа – переносчики
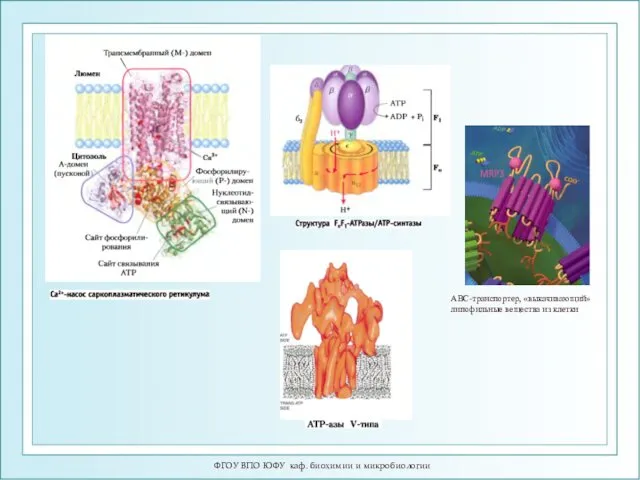
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
АВС-транспортер, «выкачивающий» липофильные вещества из
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
АВС-транспортер, «выкачивающий» липофильные вещества из
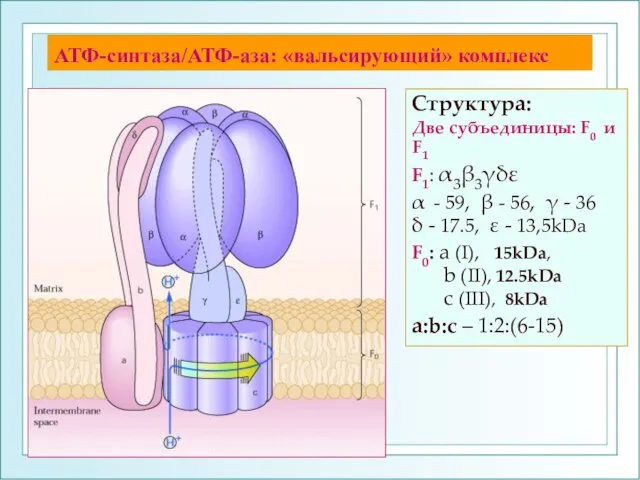
АТФ-синтаза/АТФ-аза: «вальсирующий» комплекс
Структура:
Две субъединицы: F0 и F1
F1: α3β3γδε
α - 59, β
АТФ-синтаза/АТФ-аза: «вальсирующий» комплекс
Структура:
Две субъединицы: F0 и F1
F1: α3β3γδε
α - 59, β
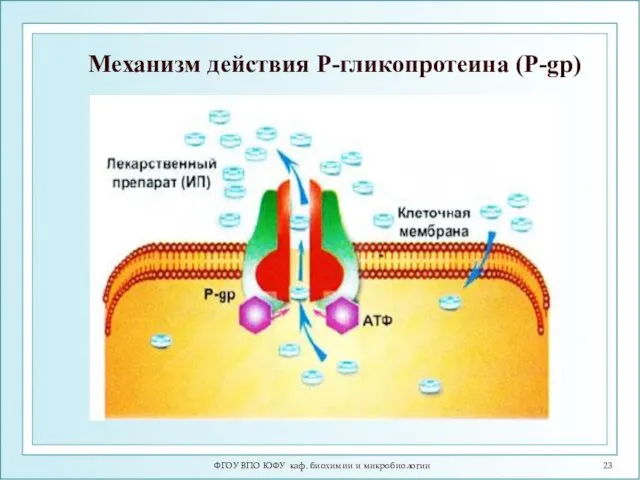
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Механизм действия Р-гликопротеина (P-gp)
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Механизм действия Р-гликопротеина (P-gp)
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
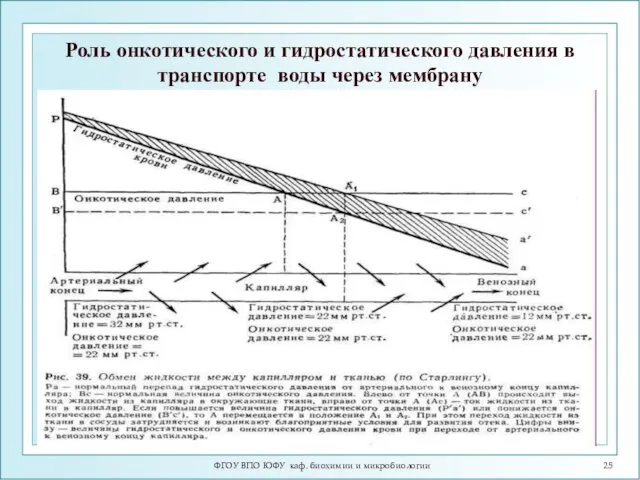
Роль онкотического и гидростатического давления в транспорте воды через мембрану
ФГОУ ВПО
Роль онкотического и гидростатического давления в транспорте воды через мембрану
ФГОУ ВПО
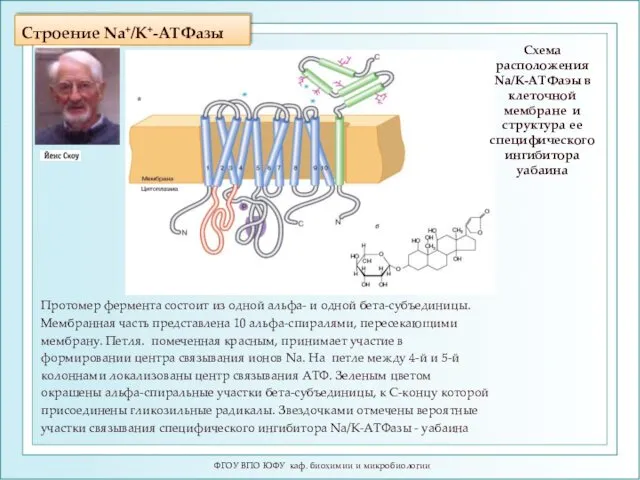
Строение Na+/K+-ATФазы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Протомер фермента состоит из
Строение Na+/K+-ATФазы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Протомер фермента состоит из
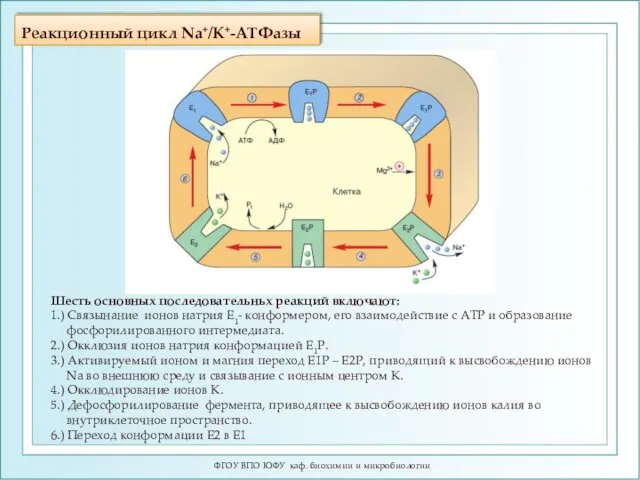
Реакционный цикл Na+/K+-ATФазы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Шесть основных последовательньх
Реакционный цикл Na+/K+-ATФазы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Шесть основных последовательньх
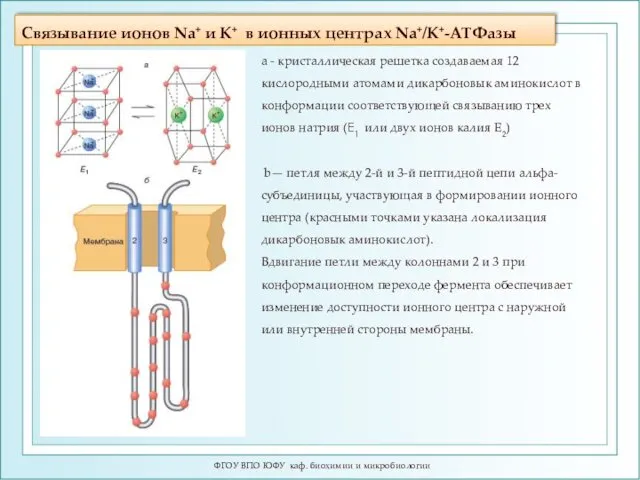
Связывание ионов Na+ и K+ в ионных центрах Na+/K+-ATФазы
ФГОУ ВПО ЮФУ
Связывание ионов Na+ и K+ в ионных центрах Na+/K+-ATФазы
ФГОУ ВПО ЮФУ
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
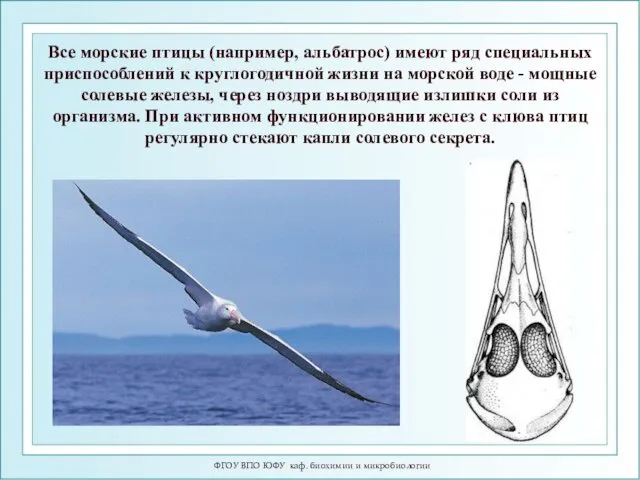
Все морские птицы (например, альбатрос)
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Все морские птицы (например, альбатрос)

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Биопотенциалы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Биопотенциалы

Генерация и распространение биоэлектрических потенциалов - важнейшее физическое явление в живых
Генерация и распространение биоэлектрических потенциалов - важнейшее физическое явление в живых

Для возникновения биопотенциалов решающее значение имеют потенциалы, обусловленные асимметричным, неравномерным распределением
Для возникновения биопотенциалов решающее значение имеют потенциалы, обусловленные асимметричным, неравномерным распределением
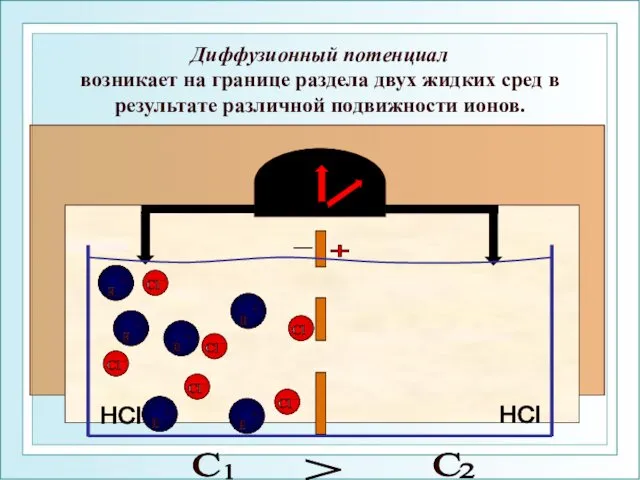
Диффузионный потенциал
возникает на границе раздела двух жидких сред в результате
Диффузионный потенциал возникает на границе раздела двух жидких сред в результате

Диффузионный потенциал определяется из уравнения Гендерсона
Где
U – подвижность катионов
V –
Диффузионный потенциал определяется из уравнения Гендерсона
Где
U – подвижность катионов
V –

Мембранный потенциал
возникает на границе раздела полупроницаемой мембраны, имеющей фиксированный отрицательный
Мембранный потенциал возникает на границе раздела полупроницаемой мембраны, имеющей фиксированный отрицательный

Мембранный потенциал определяют из уравнения Нернста
Где
R – универсальная газовая постоянная
Т –
Мембранный потенциал определяют из уравнения Нернста
Где
R – универсальная газовая постоянная
Т –

Фазовые потенциалы
возникают на границе раздела двух несмешивающихся фаз.
нитробензол
+
+
Фазовые потенциалы
возникают на границе раздела двух несмешивающихся фаз.
нитробензол
+
+
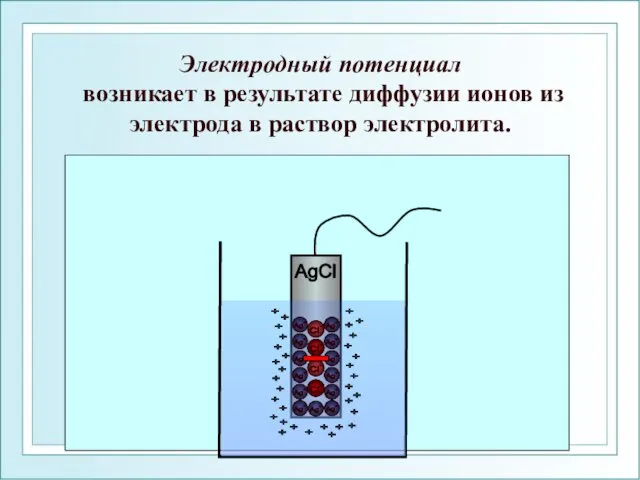
Электродный потенциал
возникает в результате диффузии ионов из электрода в раствор
Электродный потенциал возникает в результате диффузии ионов из электрода в раствор

В 1902 году Бернштейном была выдвинута мембранная теория биопотенциалов.
В 50-60-х
В 1902 году Бернштейном была выдвинута мембранная теория биопотенциалов. В 50-60-х

Сущность мембранной теории биопотенциалов
Потенциал покоя и потенциал действия является по
Сущность мембранной теории биопотенциалов
Потенциал покоя и потенциал действия является по

Мембранным потенциалом
называется разность потенциалов между внутренней и наружной поверхностями мембраны.
Мембранным потенциалом
называется разность потенциалов между внутренней и наружной поверхностями мембраны.

Потенциал покоя – стационарная разность электрических потенциалов, регистрируемых между внутренней и
Потенциал покоя – стационарная разность электрических потенциалов, регистрируемых между внутренней и
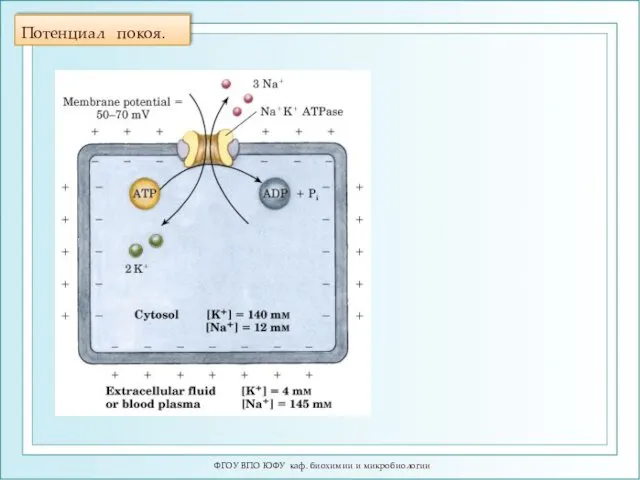
Потенциал покоя.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Потенциал покоя.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
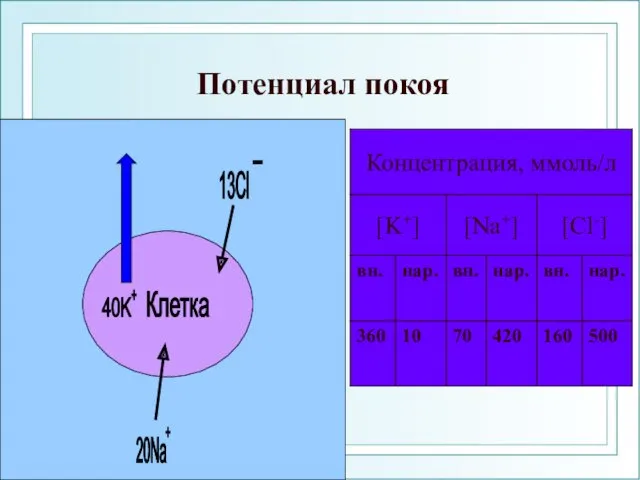
Потенциал покоя
Потенциал покоя
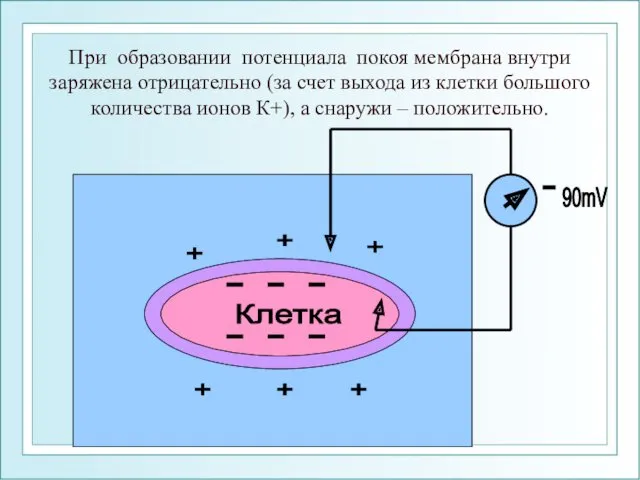
При образовании потенциала покоя мембрана внутри заряжена отрицательно (за счет выхода
При образовании потенциала покоя мембрана внутри заряжена отрицательно (за счет выхода
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
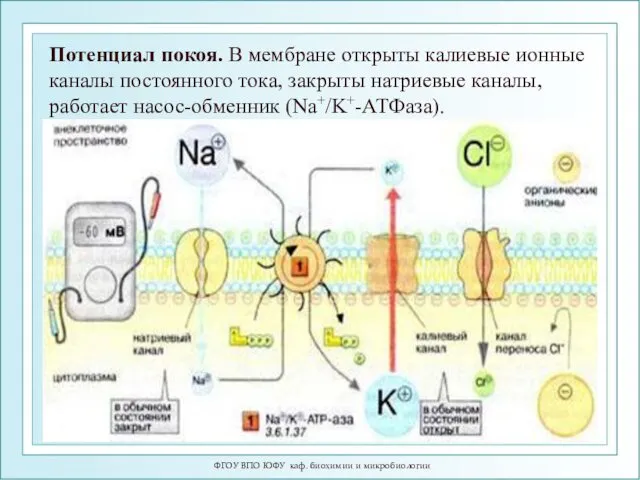
Потенциал покоя. В мембране открыты калиевые
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Потенциал покоя. В мембране открыты калиевые
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
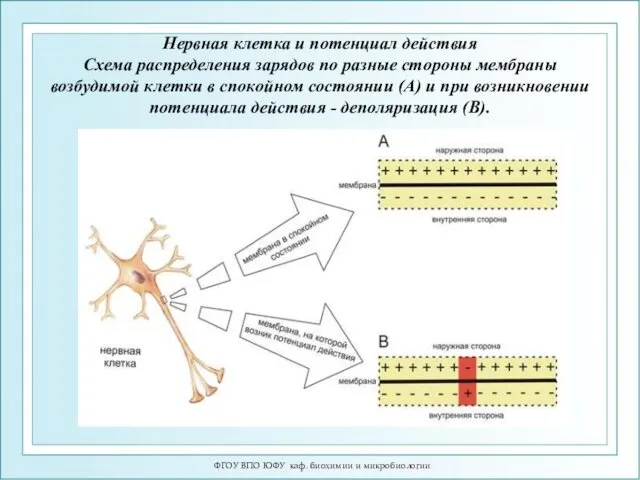
Нервная клетка и потенциал действия
Схема
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Нервная клетка и потенциал действия Схема

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Причины отрицательности мембранного потенциала покоя:
•
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Причины отрицательности мембранного потенциала покоя:
•
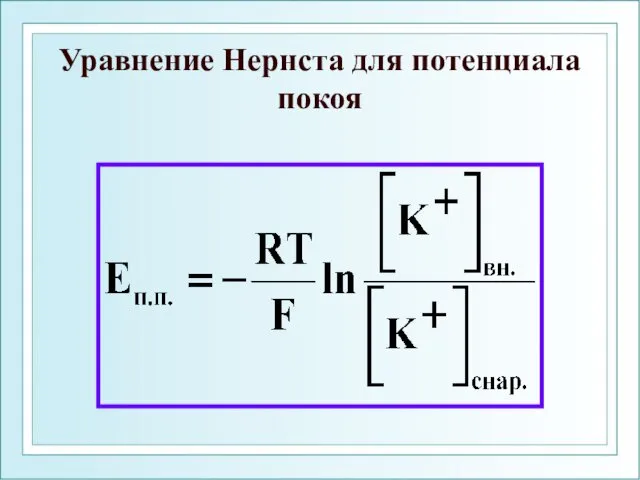
Уравнение Нернста для потенциала покоя
Уравнение Нернста для потенциала покоя

Уравнение Гольдмана
для потенциала покоя
Уравнение Гольдмана
для потенциала покоя

Отношение коэффициентов проницаемости для состояния покоя
PК : РNa : PCl =
Отношение коэффициентов проницаемости для состояния покоя
PК : РNa : PCl =

С учетом работы электрогенных ионных насосов для мембранного потенциала было получено
С учетом работы электрогенных ионных насосов для мембранного потенциала было получено
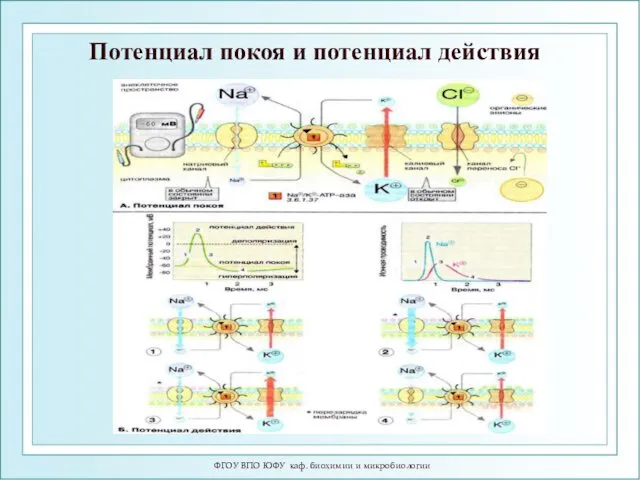
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Потенциал покоя и потенциал действия
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Потенциал покоя и потенциал действия

Потенциал действия (ПД) – это последовательность изменений, мембранного потенциала, которая запускается
Потенциал действия (ПД) – это последовательность изменений, мембранного потенциала, которая запускается

Потенциалом действия называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и
Потенциалом действия называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и
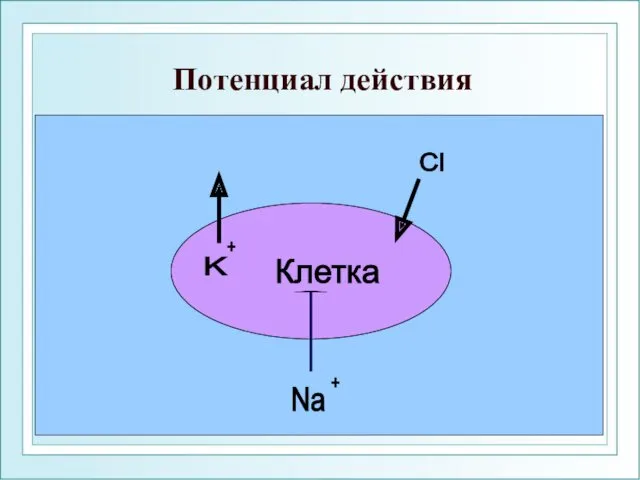
Потенциал действия
Потенциал действия
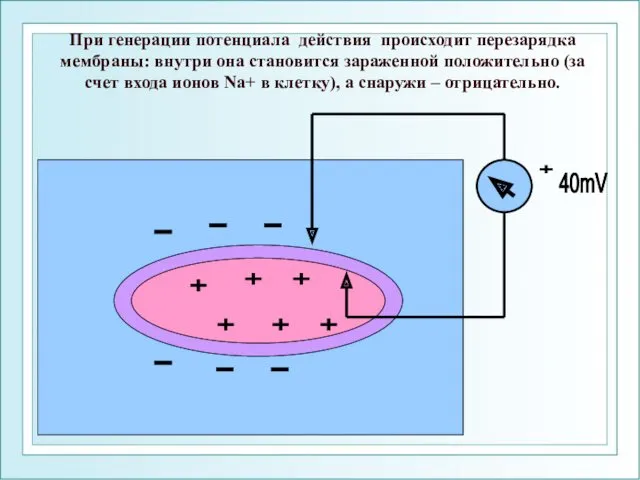
При генерации потенциала действия происходит перезарядка мембраны: внутри она становится зараженной
При генерации потенциала действия происходит перезарядка мембраны: внутри она становится зараженной

Возбудимость – это способность клеток к быстрому ответу на раздражение, проявляющемуся
Возбудимость – это способность клеток к быстрому ответу на раздражение, проявляющемуся

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Фазы потенциала действия
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Фазы потенциала действия

Стимуляция нервной клетки достигает порога возбудимости, необходимого для возникновения потенциала действия.
Начальное
Стимуляция нервной клетки достигает порога возбудимости, необходимого для возникновения потенциала действия.
Начальное

Поскольку деполяризация в клетке продолжается, открывается больше потенциал-зависимых К+-каналов, и К+
Поскольку деполяризация в клетке продолжается, открывается больше потенциал-зависимых К+-каналов, и К+
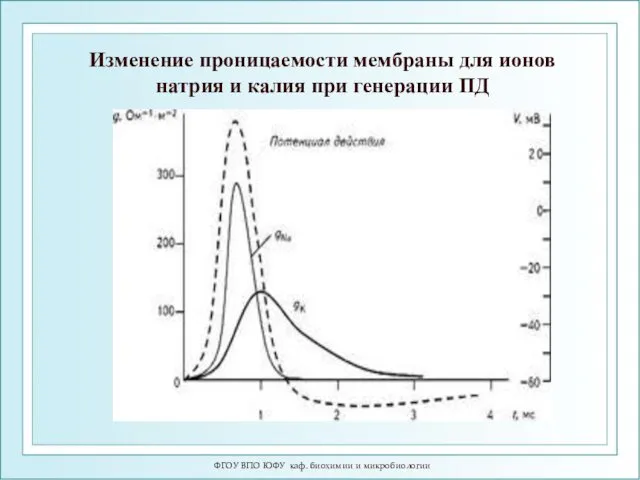
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Изменение проницаемости мембраны для ионов
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Изменение проницаемости мембраны для ионов
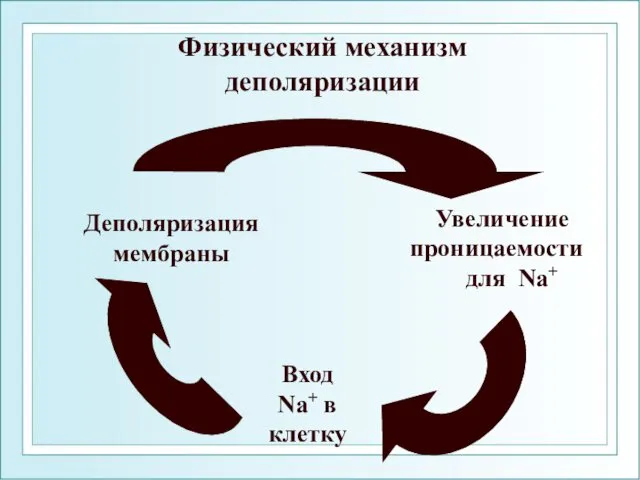
Физический механизм деполяризации
Увеличение
проницаемости
для Na+
Вход
Na+ в
клетку
Деполяризация
Физический механизм деполяризации
Увеличение
проницаемости
для Na+
Вход
Na+ в
клетку
Деполяризация

Отношение коэффициентов проницаемости ионов для фазы деполяризации
PК : РNa : PCl
Отношение коэффициентов проницаемости ионов для фазы деполяризации
PК : РNa : PCl
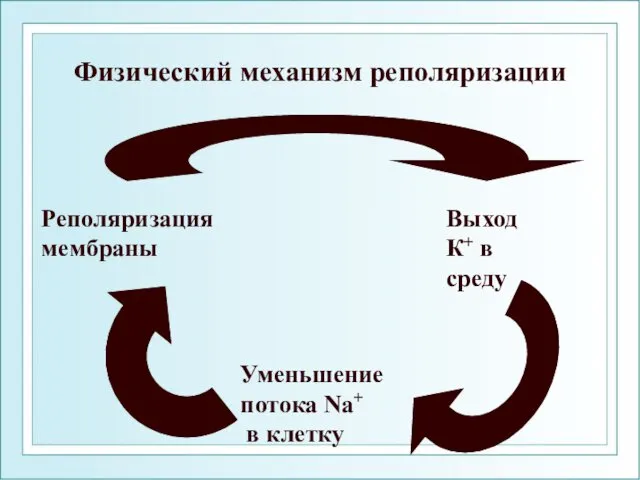
Физический механизм реполяризации
Выход
К+ в
среду
Уменьшение
потока Na+
в клетку
Реполяризация
мембраны
Физический механизм реполяризации
Выход
К+ в
среду
Уменьшение
потока Na+
в клетку
Реполяризация
мембраны

Формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов
Формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов
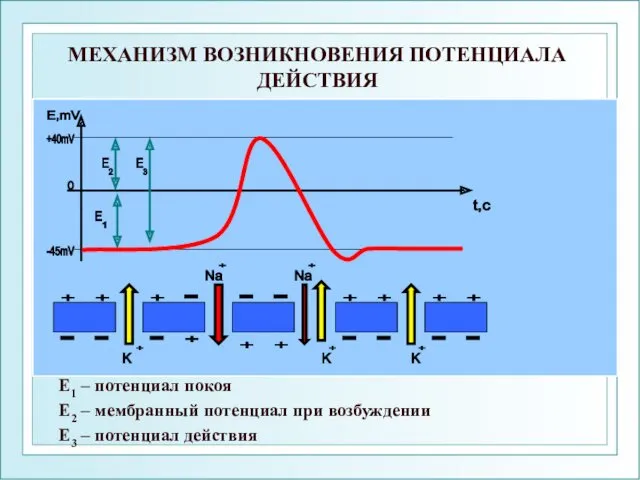
МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Е1 – потенциал покоя
Е2 – мембранный потенциал при
МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Е1 – потенциал покоя
Е2 – мембранный потенциал при
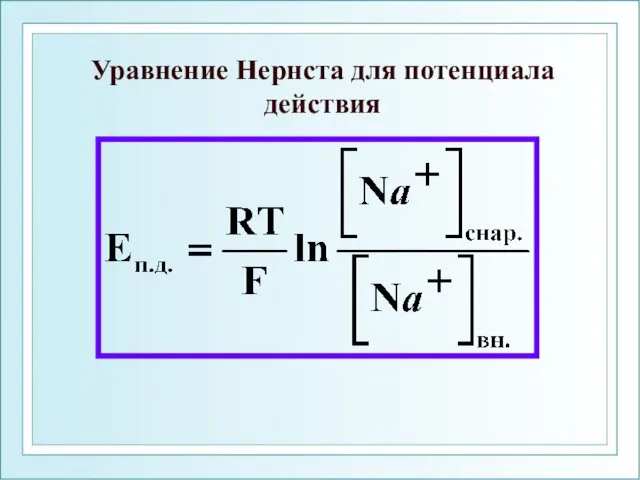
Уравнение Нернста для потенциала действия
Уравнение Нернста для потенциала действия
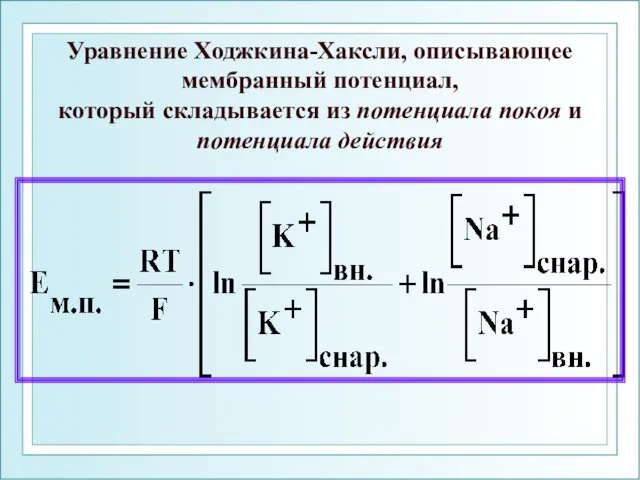
Уравнение Ходжкина-Хаксли, описывающее
мембранный потенциал,
который складывается из потенциала покоя и
Уравнение Ходжкина-Хаксли, описывающее мембранный потенциал, который складывается из потенциала покоя и

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Ионные каналы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Ионные каналы

Ионный канал.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Ионный канал.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии

Ионный транспорт
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Где
z - валентность иона
e
Ионный транспорт
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Где
z - валентность иона
e

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Вероятность перехода иона из воды
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Вероятность перехода иона из воды
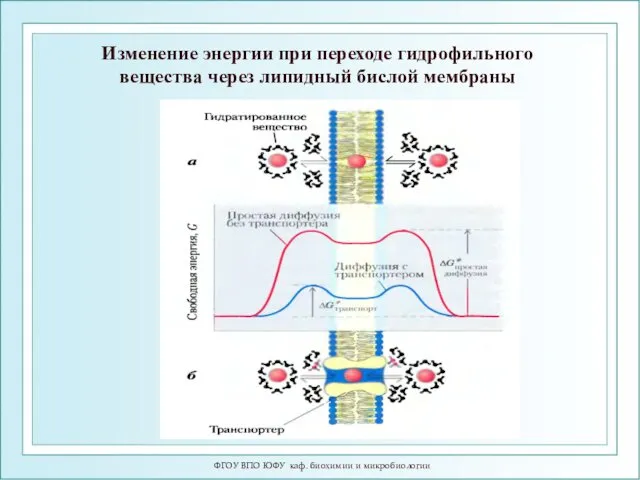
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Изменение энергии при переходе гидрофильного
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Изменение энергии при переходе гидрофильного

Свойства ионных каналов
Селективность
Независимость работы отдельных каналов
Дискретный характер проводимости
Зависимость параметров каналов от
Свойства ионных каналов
Селективность
Независимость работы отдельных каналов
Дискретный характер проводимости
Зависимость параметров каналов от

Свойства ионных каналов
1.Селективность – способность пропускать ионы одного типа.
ИК обладают абсолютной
Свойства ионных каналов
1.Селективность – способность пропускать ионы одного типа.
ИК обладают абсолютной

2.Независимость работы отдельных каналов
Прохождение тока через отдельный ионный канал не
2.Независимость работы отдельных каналов
Прохождение тока через отдельный ионный канал не

3.Дискретный характер проводимости
Проводимость ионного канала дискретна, и он может находится
3.Дискретный характер проводимости
Проводимость ионного канала дискретна, и он может находится

4.Зависимость параметров каналов от мембранного потенциала
Ионные каналы нервных волокон чувствительны
4.Зависимость параметров каналов от мембранного потенциала
Ионные каналы нервных волокон чувствительны

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
В 2003 году Нобелевская премия по химии
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
В 2003 году Нобелевская премия по химии
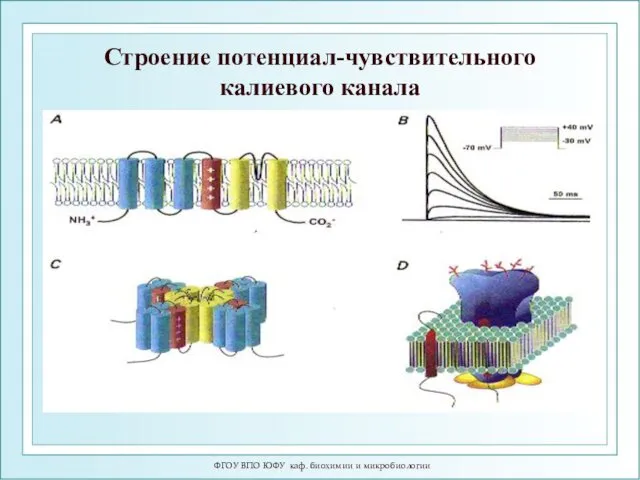
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Строение потенциал-чувствительного калиевого канала
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Строение потенциал-чувствительного калиевого канала
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
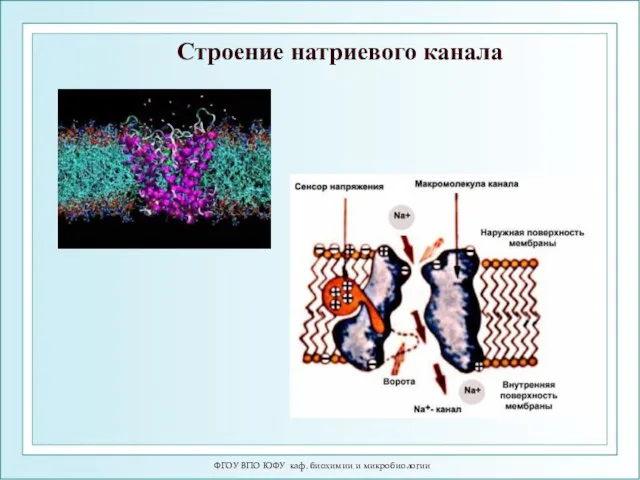
Строение натриевого канала
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Строение натриевого канала
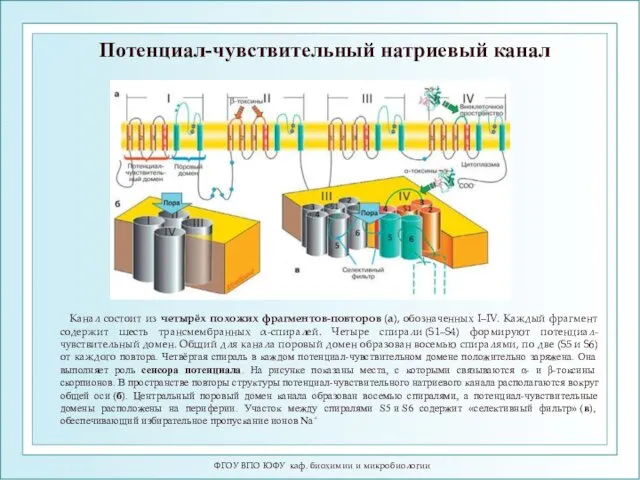
Потенциал-чувствительный натриевый канал
Канал состоит из четырёх похожих фрагментов-повторов (а), обозначенных I–IV. Каждый
Потенциал-чувствительный натриевый канал
Канал состоит из четырёх похожих фрагментов-повторов (а), обозначенных I–IV. Каждый

Сравнение параметров ионных каналов
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Сравнение параметров ионных каналов
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
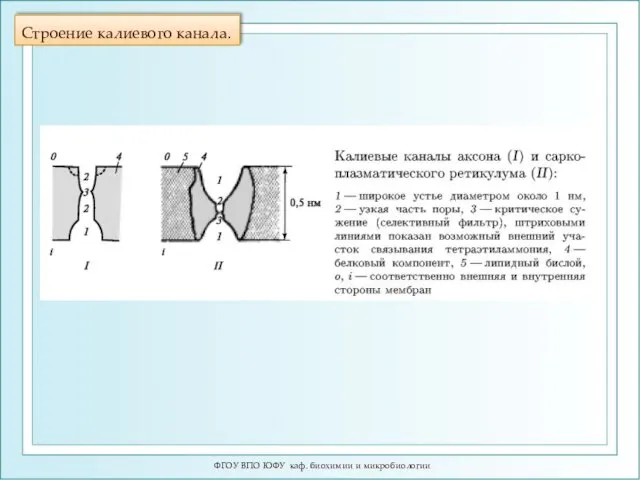
Строение калиевого канала.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Строение калиевого канала.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
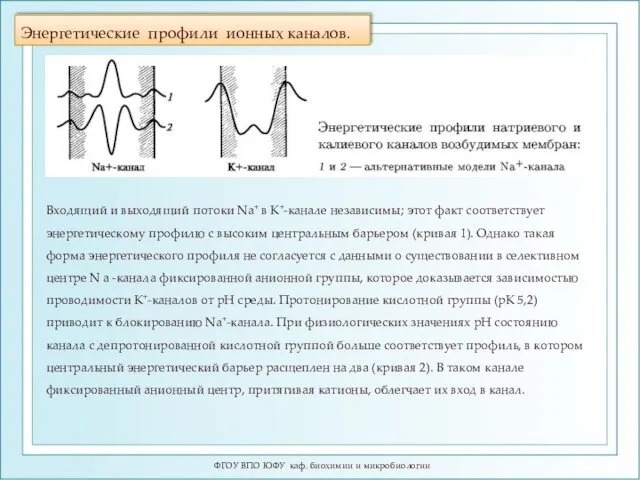
Энергетические профили ионных каналов.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Входящий и
Энергетические профили ионных каналов.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Входящий и

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Блокатор натриевых каналов тетродотоксин
Рыба фугу —
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Блокатор натриевых каналов тетродотоксин
Рыба фугу —
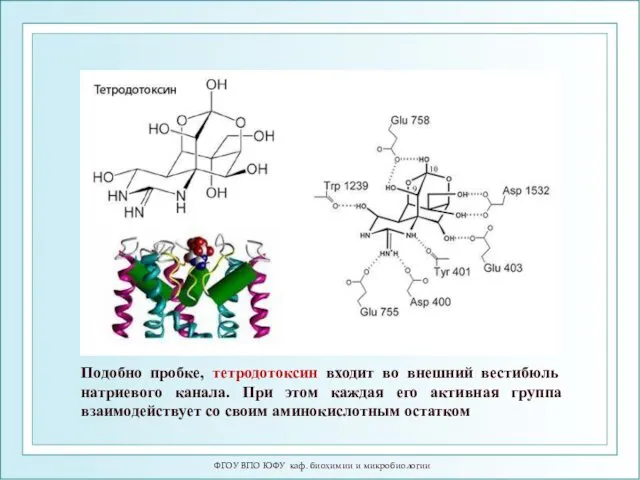
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Подобно пробке, тетродотоксин входит во
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Подобно пробке, тетродотоксин входит во

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Хищный брюхоногий моллюск Conus textile и структура
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Хищный брюхоногий моллюск Conus textile и структура

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Третий класс соединений, воздействующих на
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Третий класс соединений, воздействующих на
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
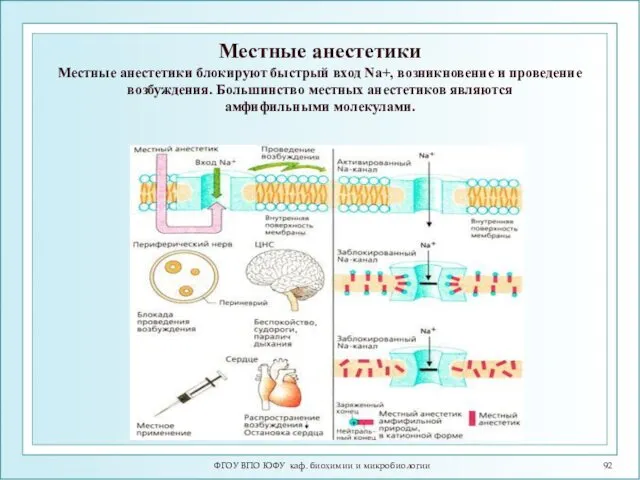
Местные анестетики
Местные анестетики блокируют быстрый
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Местные анестетики
Местные анестетики блокируют быстрый
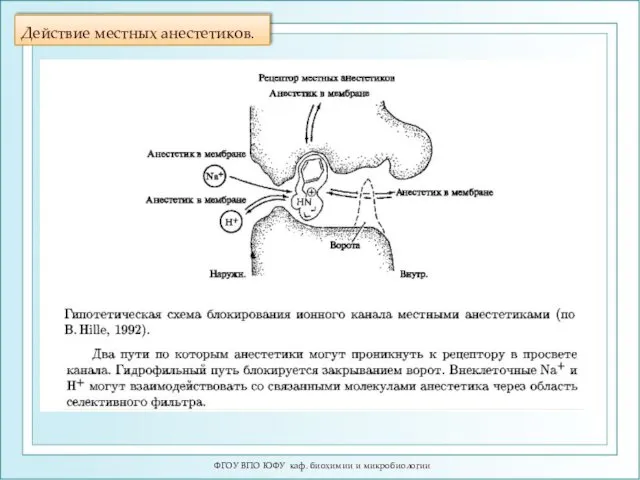
Действие местных анестетиков.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Действие местных анестетиков.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
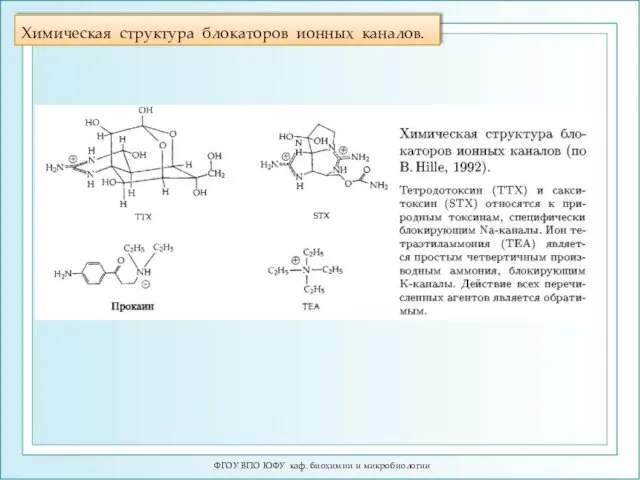
Химическая структура блокаторов ионных каналов.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Химическая структура блокаторов ионных каналов.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
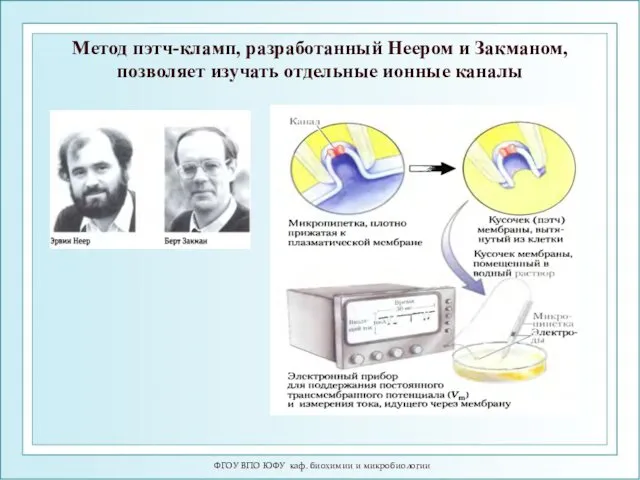
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Метод пэтч-кламп, разработанный Неером и
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Метод пэтч-кламп, разработанный Неером и

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Нобелевская премия по химии 2003
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Нобелевская премия по химии 2003
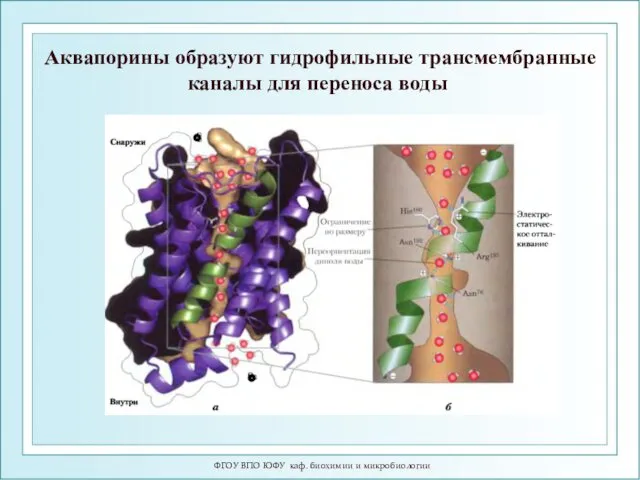
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Аквапорины образуют гидрофильные трансмембранные каналы
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Аквапорины образуют гидрофильные трансмембранные каналы

Индуцированный ионный транспорт осуществляется ионофорами, которые делятся на 2 группы:
1. Подвижные
Индуцированный ионный транспорт осуществляется ионофорами, которые делятся на 2 группы:
1. Подвижные

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Подвижные переносчики.
Перенос иона через
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Подвижные переносчики.
Перенос иона через

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Подвижные переносчики. Валиномицин.
Антибиотик валиномицин впервые
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Подвижные переносчики. Валиномицин.
Антибиотик валиномицин впервые
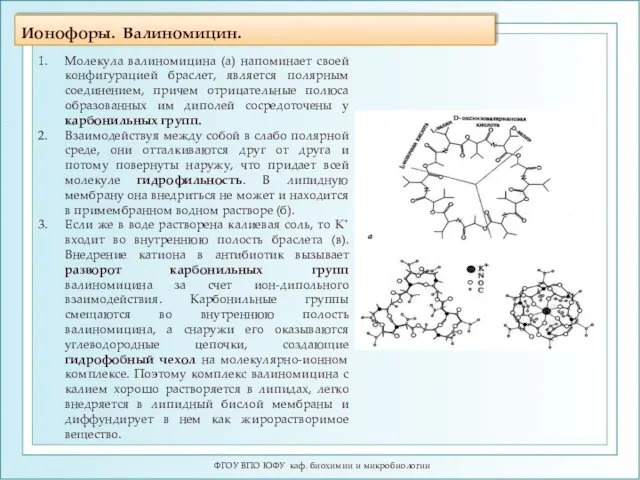
Ионофоры. Валиномицин.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Молекула валиномицина (а)
Ионофоры. Валиномицин.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Молекула валиномицина (а)
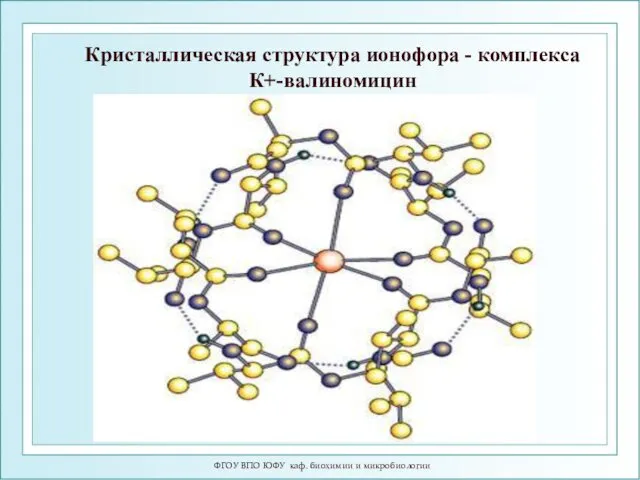
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Кристаллическая структура ионофора - комплекса
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Кристаллическая структура ионофора - комплекса
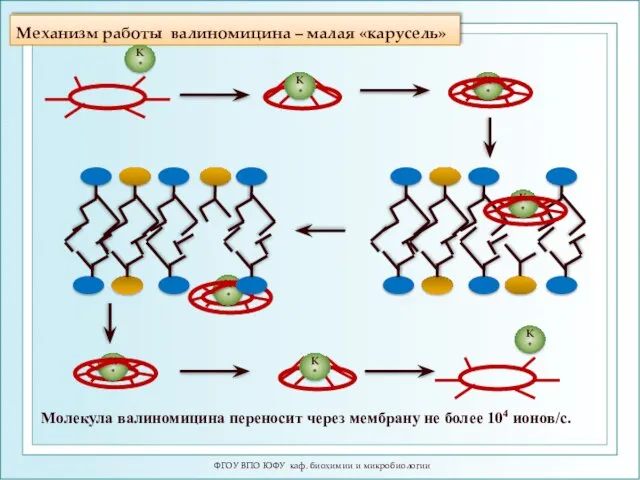
Механизм работы валиномицина – малая «карусель»
ФГОУ ВПО ЮФУ каф. биохимии
Механизм работы валиномицина – малая «карусель»
ФГОУ ВПО ЮФУ каф. биохимии

ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Скульптурная композиция комплекса
К+-валиномицин перед
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Скульптурная композиция комплекса
К+-валиномицин перед
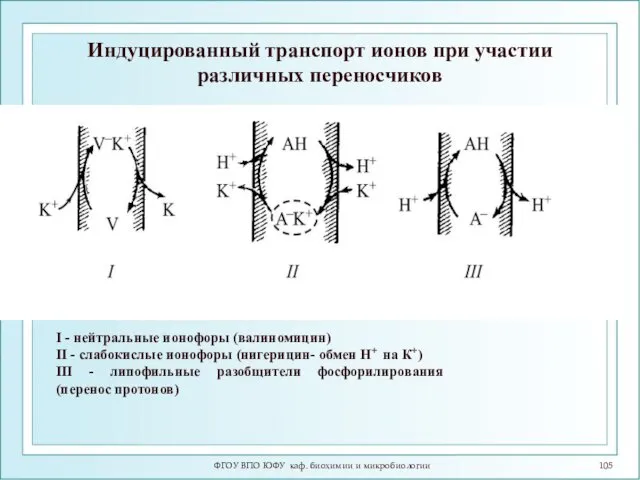
Индуцированный транспорт ионов при участии различных переносчиков
I - нейтральные ионофоры (валиномицин)
II
Индуцированный транспорт ионов при участии различных переносчиков
I - нейтральные ионофоры (валиномицин)
II
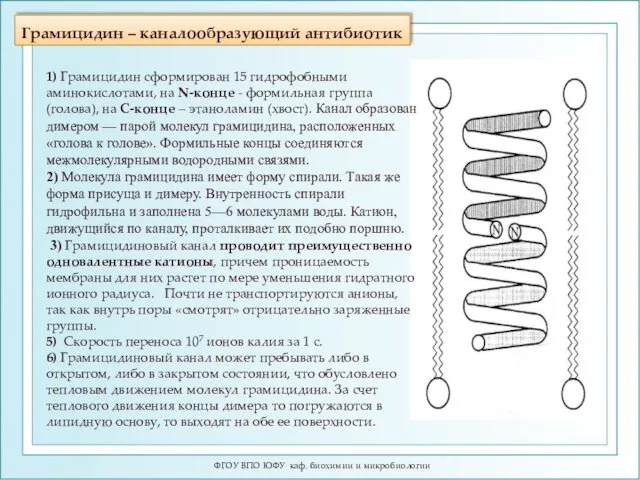
Грамицидин – каналообразующий антибиотик
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
1) Грамицидин
Грамицидин – каналообразующий антибиотик
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
1) Грамицидин
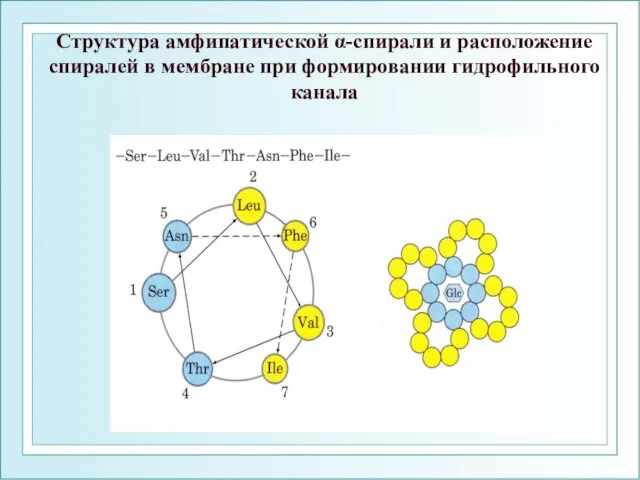
Структура амфипатической α-спирали и расположение спиралей в мембране при формировании гидрофильного
Структура амфипатической α-спирали и расположение спиралей в мембране при формировании гидрофильного
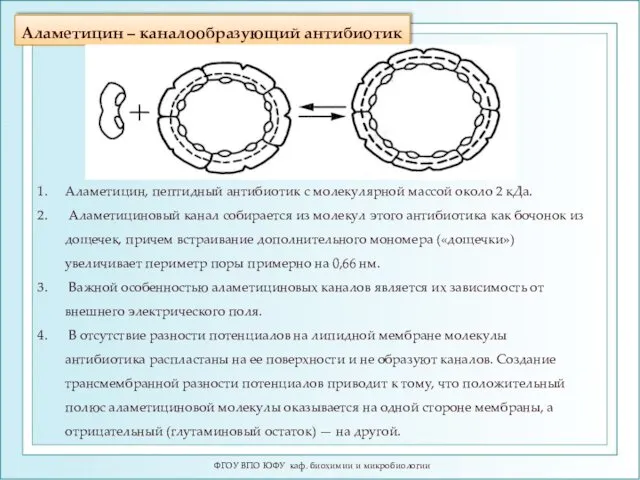
Аламетицин – каналообразующий антибиотик
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Аламетицин, пептидный
Аламетицин – каналообразующий антибиотик
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Аламетицин, пептидный
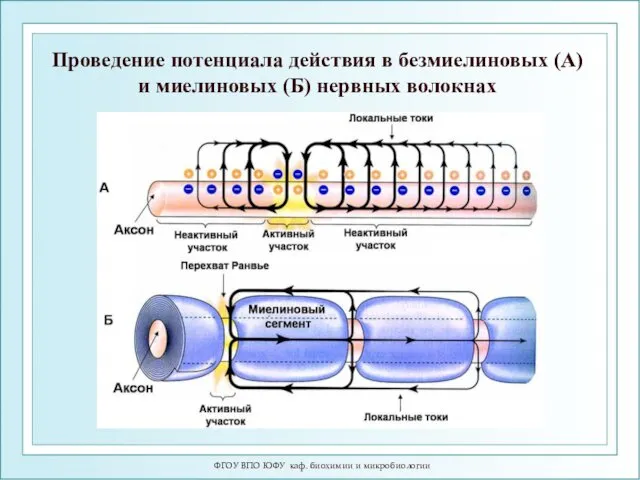
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Проведение потенциала действия в безмиелиновых
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Проведение потенциала действия в безмиелиновых
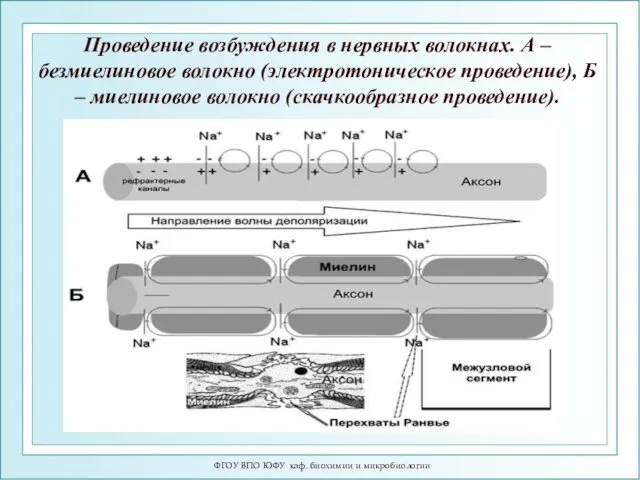
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Проведение возбуждения в нервных волокнах.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии
Проведение возбуждения в нервных волокнах.
 Характеристики тел и веществ презентация к уроку естествознания 5 класс ФГОС
Характеристики тел и веществ презентация к уроку естествознания 5 класс ФГОС nanotehnologii_za_i_protiv
nanotehnologii_za_i_protiv Ошибочная активация Bluelink. Технический запрос
Ошибочная активация Bluelink. Технический запрос Фотоэффект құбылысы. Эйнштейн формуласы. Фотоэффект құбылысын техникада пайдалану
Фотоэффект құбылысы. Эйнштейн формуласы. Фотоэффект құбылысын техникада пайдалану Сила трения
Сила трения Дифракция света
Дифракция света Электротехнические материалы. ТЭО
Электротехнические материалы. ТЭО Презентация к уроку Реактивные двигатели
Презентация к уроку Реактивные двигатели Испытания на сдвиг, методы определения модулей сдвига при кручении плоских образцов и пластин по периоду крутильных колебаний
Испытания на сдвиг, методы определения модулей сдвига при кручении плоских образцов и пластин по периоду крутильных колебаний Напряженное и деформированное состояния элемента в окрестности точки
Напряженное и деформированное состояния элемента в окрестности точки Goorui Company Introduction
Goorui Company Introduction Кинетическая энергия
Кинетическая энергия открытый урок-презентация на тему История Российской атомной энергетики
открытый урок-презентация на тему История Российской атомной энергетики Електромагнітні коливання та хвилі
Електромагнітні коливання та хвилі Генератор переменного тока. Виды генераторов
Генератор переменного тока. Виды генераторов Открытие протона и нейтрона. Строение атомного ядра
Открытие протона и нейтрона. Строение атомного ядра Проектирование участка для ремонта двигателя внутреннего сгорания трактора Т-40М ремонтного предприятия на 200 тракторов в год
Проектирование участка для ремонта двигателя внутреннего сгорания трактора Т-40М ремонтного предприятия на 200 тракторов в год Тележка, модели 18-194-1
Тележка, модели 18-194-1 Открытие электромагнитной индукции
Открытие электромагнитной индукции Организация работ по диагностированию, техническому обслуживанию ремонту ходовой части ВАЗ 2109
Организация работ по диагностированию, техническому обслуживанию ремонту ходовой части ВАЗ 2109 Определение перемещений сечений стержневых систем
Определение перемещений сечений стержневых систем Игра Звёдный час по физике 7-8 класс
Игра Звёдный час по физике 7-8 класс Сила. Законы Ньютона
Сила. Законы Ньютона Определение ударной вязкости и порога хладноломкости стали
Определение ударной вязкости и порога хладноломкости стали Термометр. Виды термометров
Термометр. Виды термометров Законы динамики. Уравнения движения. Лекция 1
Законы динамики. Уравнения движения. Лекция 1 Динамика вращательного движения
Динамика вращательного движения Электрический ток. Закон Ома для участка цепи
Электрический ток. Закон Ома для участка цепи