- Методы спектроскопии в исследовании биополимеров

Содержание
- 2. Распространение электромагнитной волны в пространстве. Векторы Е и Н все время взаимно перпендикулярны. 1. Спекроскопия
- 3. 1. Спекроскопия Спектральный анализ - совокупность методов качественного и количественного определения состава объекта, основанная на изучении
- 4. Шкала электромагнитных излучений, используемая для исследований в области биологии 1. Спекроскопия γ – спектроскопия Рентгеновская спектроскопия
- 5. 1. Спекроскопия
- 6. 1. Спекроскопия
- 7. Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy) 2. Методы исследования вторичной структуры Исследует колебания атомов в молекуле 2 основных
- 8. Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy) 2. Методы исследования вторичной структуры ИК-спектр поглощения связан с переходами на различные
- 9. Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy) 2. Методы исследования вторичной структуры ИК-диапазон электромагнитного излучения – 750-100000 нм
- 10. Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy) 2. Методы исследования вторичной структуры
- 11. Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy) 2. Методы исследования вторичной структуры
- 12. 2. Методы исследования вторичной структуры Исследование вторичной структуры белка методом ИК-спектроскопии Амид I
- 13. Круговой дихроизм (КД-спектроскопия,CD-spectroscopy) 2. Методы исследования вторичной структуры Исследует взаимодействие плоскополяризованного света с оптически активными молекулами
- 14. Круговой дихроизм (КД-спектроскопия,CD-spectroscopy) 2. Методы исследования вторичной структуры Исследует взаимодействие плоскополяризованного света с оптически активными молекулами
- 15. Круговой дихроизм (КД-спектроскопия,CD-spectroscopy) 2. Методы исследования вторичной структуры Оптически активные вещества – вещества, имеющие хиральный центр
- 16. Круговой дихроизм (КД-спектроскопия,CD-spectroscopy) 2. Методы исследования вторичной структуры КД-спектр – зависимость кругового дихроизма от длины волны
- 17. Круговой дихроизм (КД-спектроскопия,CD-spectroscopy) 2. Методы исследования вторичной структуры КД-спектр – зависимость кругового дихроизма от длины волны
- 18. 3. Методы исследования третичной структуры белка Исследуются ядра атомов, имеющих непарное число протонов и нейтронов. Такие
- 19. 3. Методы исследования третичной структуры белка Химический сдвиг νобр - νэт νэт δ = ⋅106 [м.д.]
- 20. 3. Методы исследования третичной структуры белка
- 21. 3. Методы исследования третичной структуры белка ЯМР (водные растворы белков) Рентгеноструктурный анализ (кристаллы)
- 22. 4. Методы исследования мембран клеток
- 23. 4. Методы исследования мембран клеток Электронный парамагнитный резонанс (ЭПР)
- 24. 4. Методы исследования мембран клеток Электронный парамагнитный резонанс (ЭПР)
- 25. 4. Методы исследования мембран клеток Электронный парамагнитный резонанс (ЭПР)
- 26. 4. Методы исследования мембран клеток Электронный парамагнитный резонанс (ЭПР)
- 27. 4. Методы исследования мембран клеток Электронный парамагнитный резонанс (ЭПР) Зависимость формы сигнала подвижности группы
- 28. 5. Флуоресцентный анализ
- 30. 5. Флуоресцентный анализ
- 31. 5. Флуоресцентный анализ
- 32. 5. Флуоресцентный анализ
- 33. Флуоресцентный анализ мембран (встраивание и свечение) определяется разницей потенциалов на поверхностях мембраны АНС
- 34. Флуоресцентный анализ мембран Пирен
- 35. Флуоресцентный анализ мембран Флуоресцеиндиацетат Свечение определяется диффузией зонда в клетку, активностью эстераз и рН среды в
- 37. БИОЛЮМИНЕСЦЕНЦИЯ Биолюминесценцией называют свечение живых организмов. Оно происходит за счет специфичных химических реакций с выделением света.
- 38. Биолюминесцентные организмы
- 39. БИОЛЮМИНЕСЦЕНЦИЯ Многоклеточные животные обычно излучают свет не всем телом, а только специальными клетками фотоцитами, часто сгруппированными
- 40. Биолюминесценция наблюдается вследствие дезактивации возбужденных молекул – эмиттеров. Эмиттеры возникают в организмах в ходе биохимических реакций.
- 41. Общий механизм биолюминесценции Люциферины разных светящихся организмов: (1) агариковых грибов (2) жуков-светляков (3) динофлагеллят (4) кишечнополостных,
- 42. БИОЛЮМИНЕСЦЕНЦИЯ Химически разные биолюминесцентные реакции объединяет несколько характерных черт. 1. Всем этим реакциям абсолютно необходим кислород.
- 43. Биолюминесцентная реакция светляков Различные виды светляков (насекомые семейства Lampyridae) встречаются на юге Северной Америки, в Японии,
- 44. Биолюминесцентная реакция светляков Люцифераза светляков состоит из двух субъединиц с молекулярной массой 50000 каждая и не
- 45. Биолюминесцентная реакция светляков Максимумы биолюминесценции разных видов светляков лежат в интервале от 540 до 590 нм.
- 46. Биолюминесценция кишечнополостных Биолюминесценция среди кишечнополостных (Coelenterata) распространена довольно широко. Известно несколько десятков видов гидромедуз, излучающих свет
- 47. Биолюминесценция кишечнополостных Наиболее изученными фотопротеинами являются обелин из гидроидных полипов рода Obelia и акворин из гидромедуз
- 48. Биолюминесценция кишечнополостных Для люминесценции морских перьев Renilla характерна стадия предварительной активации люциферина. На этой стадии неактивный
- 49. Биолюминесценция кишечнополостных В 1962 г. О. Шимомура, Ф. Джонсон и Ю. Сайга отметили разницу между цветом
- 52. GFP – green fluorescent protein
- 53. Биолюминесценция бактерий Все известные в настоящее время светящиеся бактерии делятся на две группы: морские и пресноводные.
- 54. Биолюминесценция бактерий Химической основой свечения бактерий является ферментативное окисление восстановленного флавинмононуклеотида ФМН⋅Н2 и длинноцепочечного альдегида RCHO
- 55. Биолюминесценция бактерий
- 56. Биолюминесценция динофлагеллят Динофлагелляты или жгутиконостцы являются основными продуцентами света в море, по крайней мере в его
- 57. Свечение червей Среди кольчатых червей много биолюминесцентных видов, как морских, так и наземных. Морские полихеты Chaetopterus
- 58. Биолюминесценция грибов К настоящему времени описано более 40 разновидностей люминесцентных грибов. Из них большинство принадлежит ко
- 59. Биолюминесцентный анализ Биолюминесценция in vivo и in vitro нашла широкое применение в качестве аналитических методов и
- 60. Биолюминесцентный анализ В настоящее время развиваются два направления биолюминесцентного анализа: методы избирательного анализа и биолюминесцентные интегральные
- 61. Биолюминесцентный анализ Светящиеся бактерии применяются в различных областях в трех основных видах: природном (интактном), мутантном и
- 62. Биолюминесцентный анализ
- 63. Биолюминесцентный анализ Прогресс в области генной инженерии позволил создать новые светящиеся организмы. Было проведено клонирование и
- 64. Биолюминесцентный анализ Можно выделить семь основных направлений применения биолюминесцентных реакций, полученных из различных светящихся организмов: 1.
- 65. Биолюминесцентный анализ Анализ субстратов люцифераз Как уже упоминалось, люциферазы проявляют высокую специфичность к своим субстратам. Интенсивность
- 66. Биолюминесцентный анализ Мониторинг АТФ Биолюминесцентный анализ - наиболее специфичный, чувствительный и быстрый метод определения АТФ. В
- 67. Биолюминесцентный анализ Мониторинг АТФ Области применения АТФ-метрии: 1. Медицина: - обнаружение микробных инфекций в крови, моче
- 68. Биолюминесцентный анализ Определение содержания ионов кальция Концентрация ионов кальция является внутриклеточным сигналом, ответственным за контролирование многочисленных
- 69. Детекция взаимодействия биомолекул. Иммуноанализ Люциферазы применяются в качестве ферментов-маркеров в иммуноанализе наряду с пероксидазой хрена и
- 70. Биолюминесцентный анализ Биолюминесцентные репортерные гены Технология репортерных генов – одно из главных новых достижений молекулярной биологии
- 71. Биолюминесцентный анализ Биолюминесцентная визуализация Визуализация биологических процессов in vitro и in vivo стала возможной благодаря созданию
- 74. Скачать презентацию
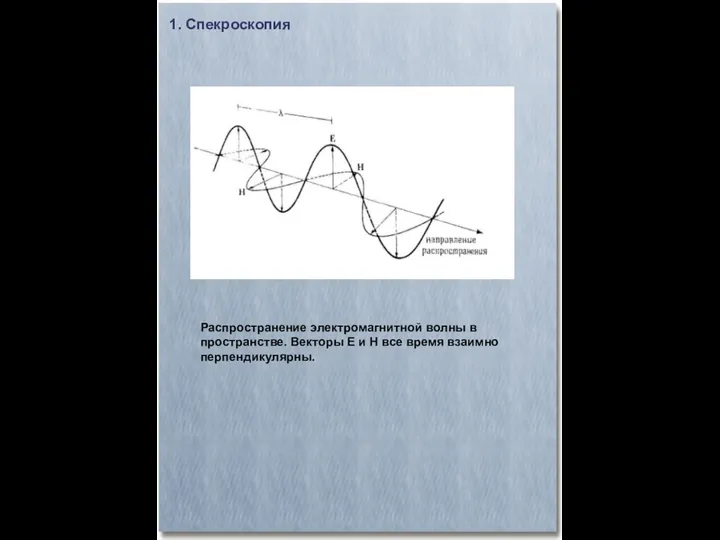
Распространение электромагнитной волны в пространстве. Векторы Е и Н все время
Распространение электромагнитной волны в пространстве. Векторы Е и Н все время

1. Спекроскопия
Спектральный анализ - совокупность методов качественного и количественного определения состава
1. Спекроскопия
Спектральный анализ - совокупность методов качественного и количественного определения состава

Шкала электромагнитных излучений, используемая
для исследований в области биологии
1. Спекроскопия
γ –
Шкала электромагнитных излучений, используемая
для исследований в области биологии
1. Спекроскопия
γ –

1. Спекроскопия
1. Спекроскопия

1. Спекроскопия
1. Спекроскопия

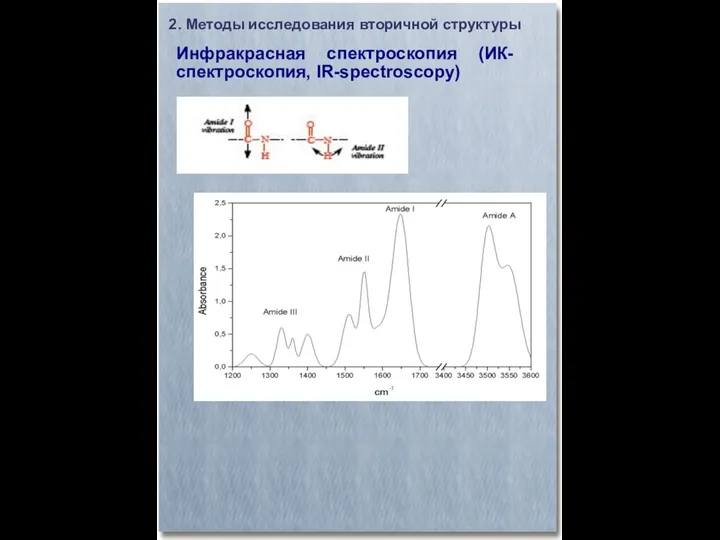
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
Исследует колебания атомов в
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
Исследует колебания атомов в

Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
ИК-спектр поглощения связан с
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
ИК-спектр поглощения связан с

Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
ИК-диапазон электромагнитного излучения –
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
ИК-диапазон электромагнитного излучения –
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
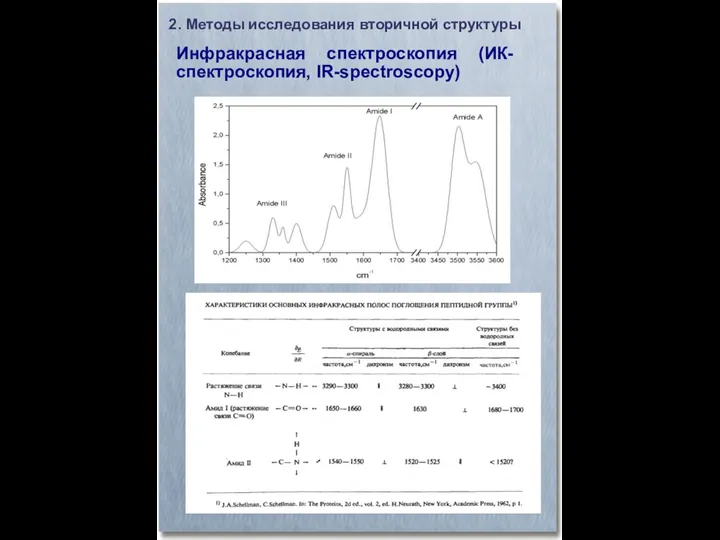
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
Инфракрасная спектроскопия (ИК-спектроскопия, IR-spectroscopy)
2. Методы исследования вторичной структуры
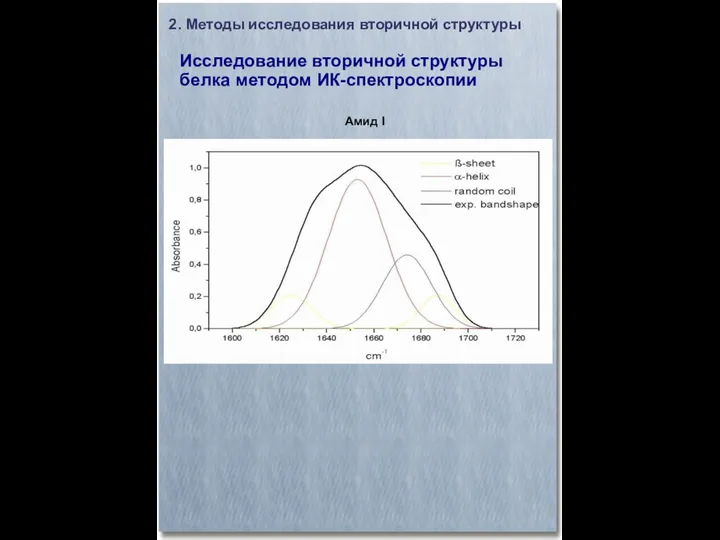
2. Методы исследования вторичной структуры
Исследование вторичной структуры белка методом ИК-спектроскопии
Амид I
2. Методы исследования вторичной структуры
Исследование вторичной структуры белка методом ИК-спектроскопии
Амид I

Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
Исследует взаимодействие плоскополяризованного света с оптически
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
Исследует взаимодействие плоскополяризованного света с оптически

Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
Исследует взаимодействие плоскополяризованного света с оптически
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
Исследует взаимодействие плоскополяризованного света с оптически

Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
Оптически активные вещества – вещества, имеющие
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
Оптически активные вещества – вещества, имеющие
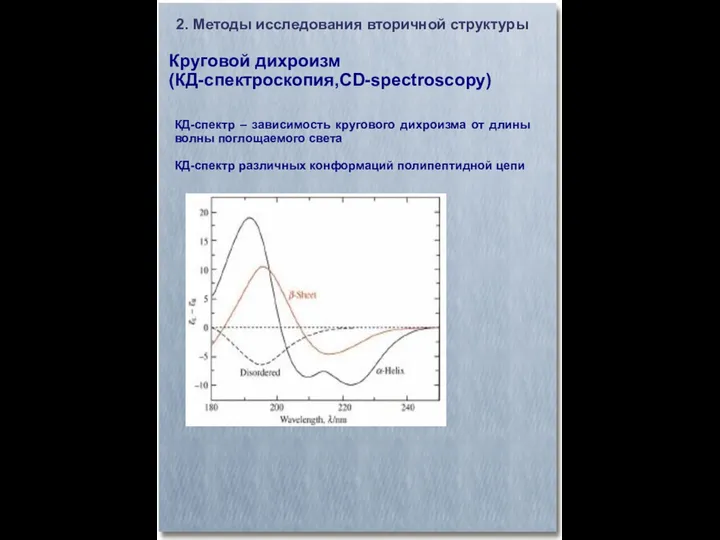
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
КД-спектр – зависимость кругового дихроизма от
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
КД-спектр – зависимость кругового дихроизма от
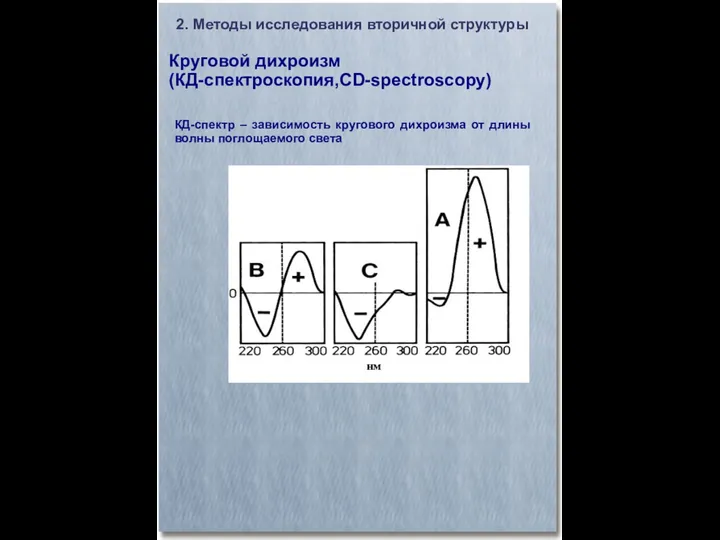
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
КД-спектр – зависимость кругового дихроизма от
Круговой дихроизм
(КД-спектроскопия,CD-spectroscopy)
2. Методы исследования вторичной структуры
КД-спектр – зависимость кругового дихроизма от

3. Методы исследования третичной структуры
белка
Исследуются ядра атомов, имеющих непарное число протонов
3. Методы исследования третичной структуры
белка
Исследуются ядра атомов, имеющих непарное число протонов
![3. Методы исследования третичной структуры белка Химический сдвиг νобр - νэт νэт δ = ⋅106 [м.д.]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/383537/slide-18.jpg)
3. Методы исследования третичной структуры
белка
Химический сдвиг
νобр - νэт
νэт
δ =
⋅106 [м.д.]
3. Методы исследования третичной структуры
белка
Химический сдвиг
νобр - νэт
νэт
δ =
⋅106 [м.д.]

3. Методы исследования третичной структуры
белка
3. Методы исследования третичной структуры
белка
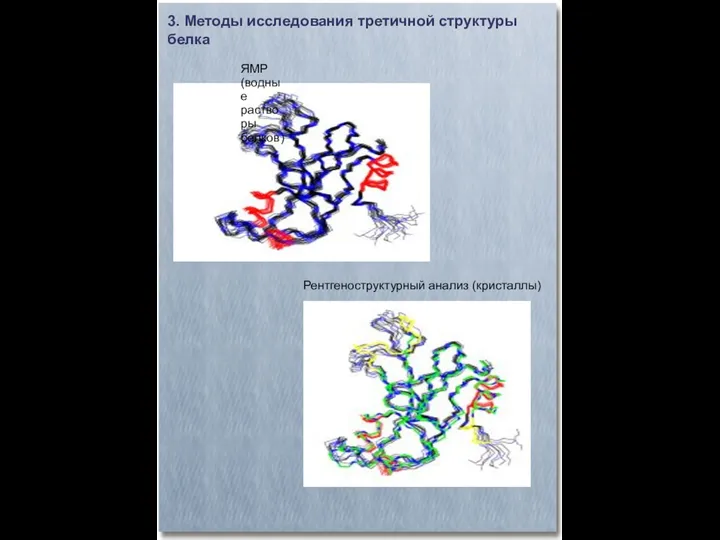
3. Методы исследования третичной структуры
белка
ЯМР (водные растворы белков)
Рентгеноструктурный анализ (кристаллы)
3. Методы исследования третичной структуры
белка
ЯМР (водные растворы белков)
Рентгеноструктурный анализ (кристаллы)
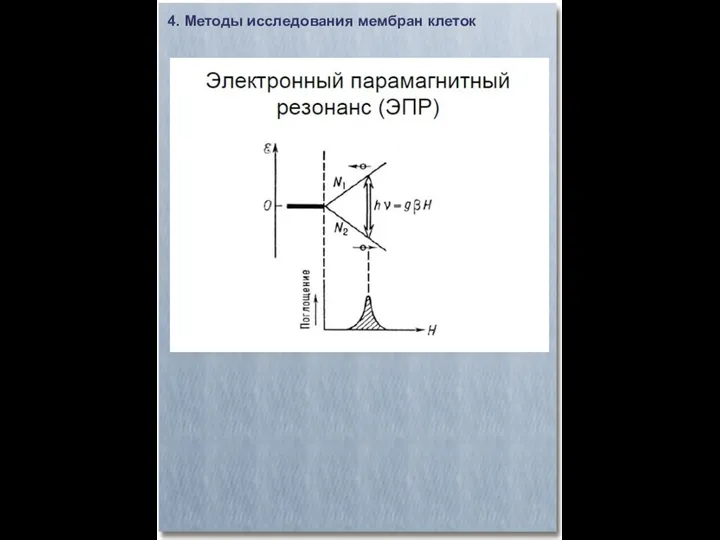
4. Методы исследования мембран клеток
4. Методы исследования мембран клеток

4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
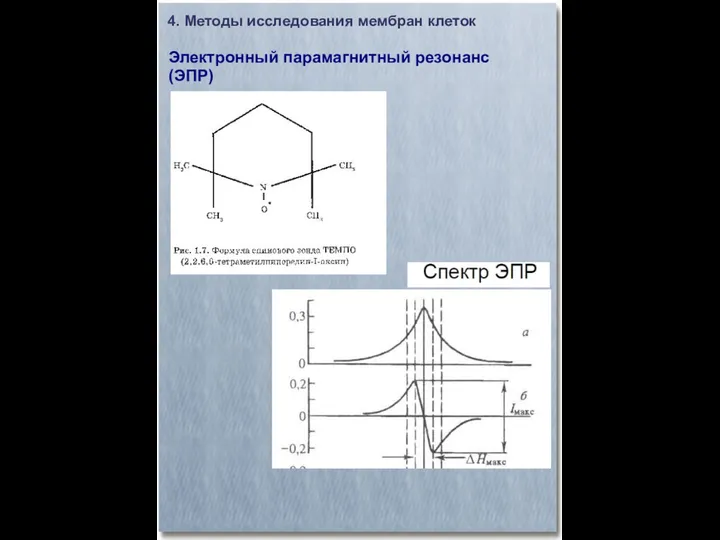
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
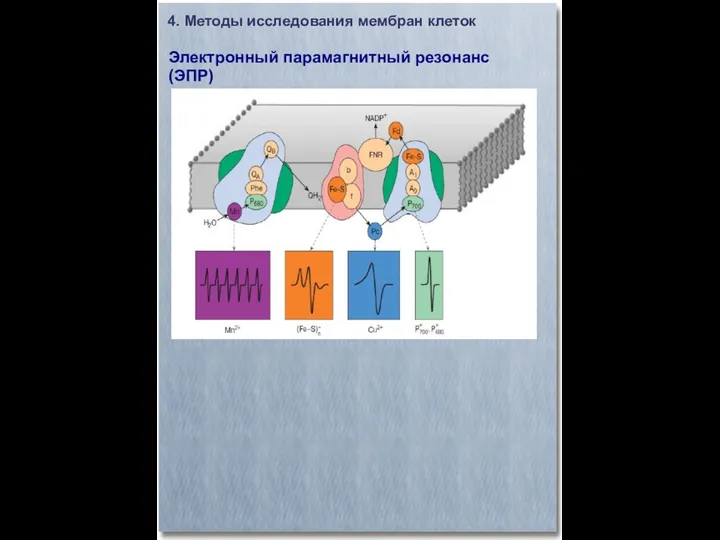
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
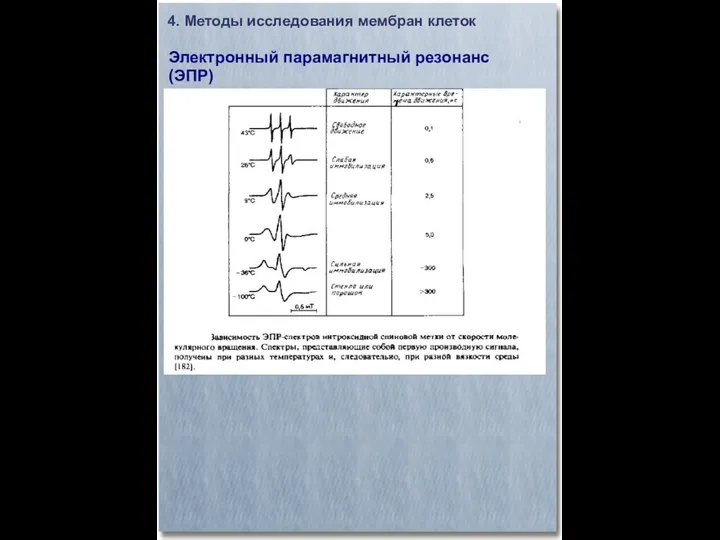
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
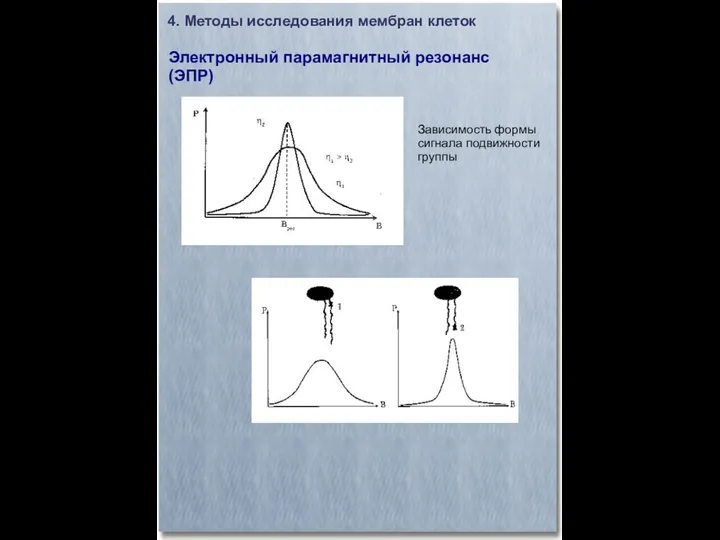
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
Зависимость формы
сигнала подвижности
группы
4. Методы исследования мембран клеток
Электронный парамагнитный резонанс (ЭПР)
Зависимость формы
сигнала подвижности
группы
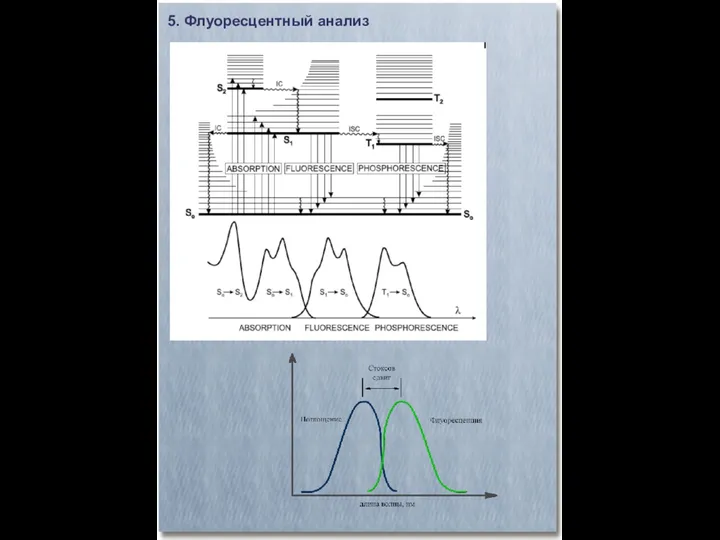
5. Флуоресцентный анализ
5. Флуоресцентный анализ

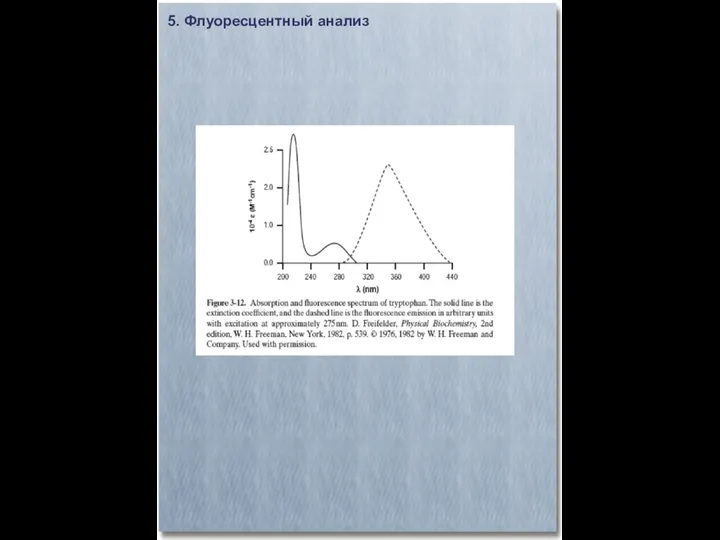
5. Флуоресцентный анализ
5. Флуоресцентный анализ
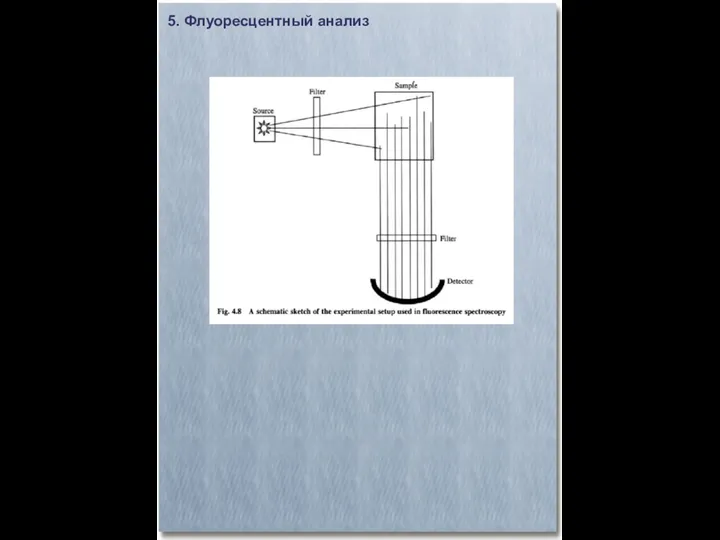
5. Флуоресцентный анализ
5. Флуоресцентный анализ
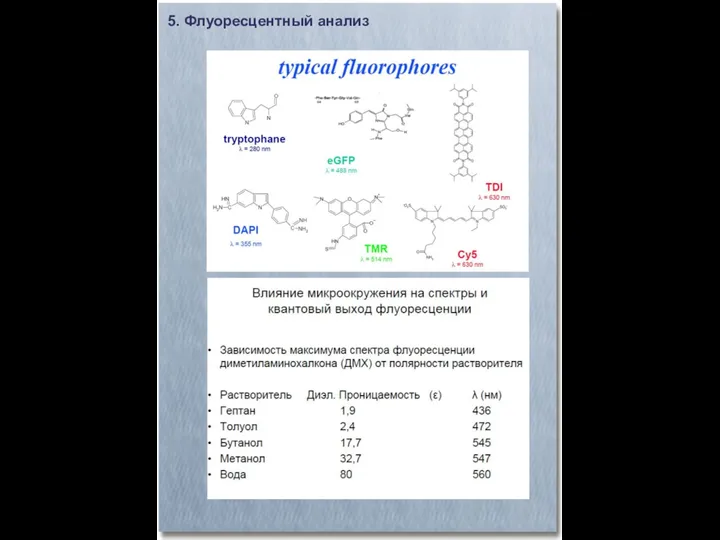
5. Флуоресцентный анализ
5. Флуоресцентный анализ
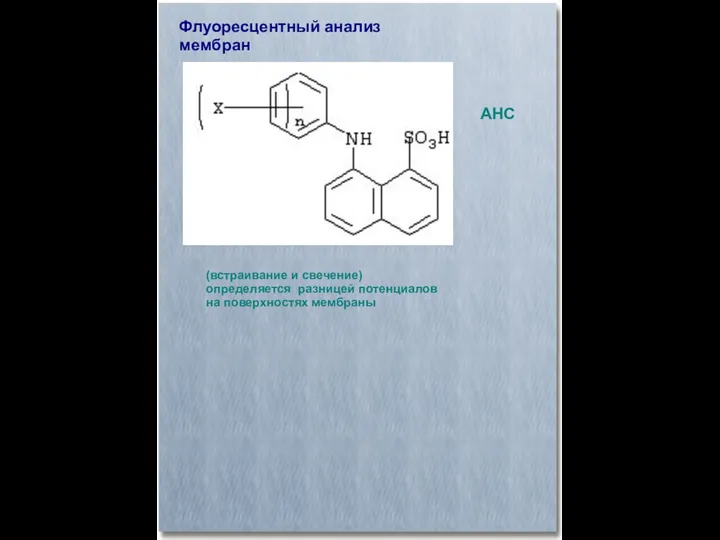
Флуоресцентный анализ мембран
(встраивание и свечение) определяется разницей потенциалов на поверхностях мембраны
АНС
Флуоресцентный анализ мембран
(встраивание и свечение) определяется разницей потенциалов на поверхностях мембраны
АНС
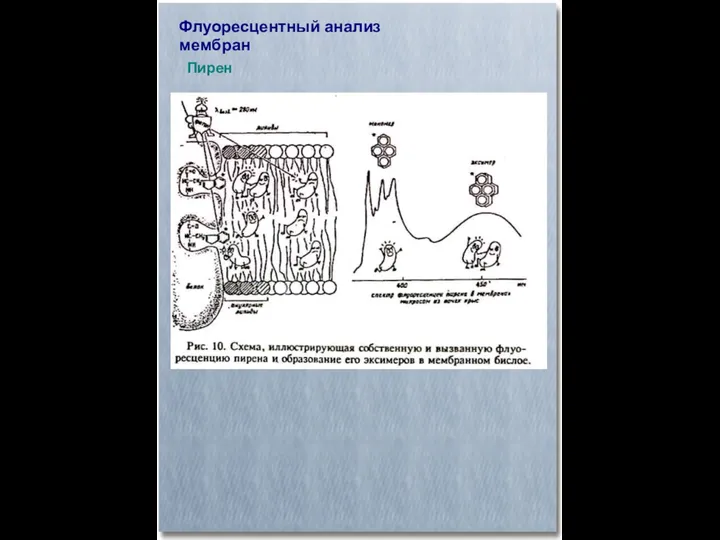
Флуоресцентный анализ мембран
Пирен
Флуоресцентный анализ мембран
Пирен
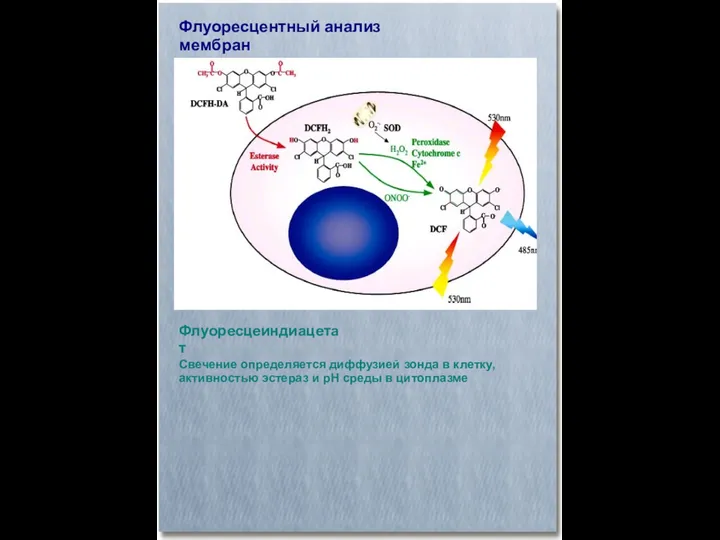
Флуоресцентный анализ мембран
Флуоресцеиндиацетат
Свечение определяется диффузией зонда в клетку, активностью эстераз и
Флуоресцентный анализ мембран
Флуоресцеиндиацетат
Свечение определяется диффузией зонда в клетку, активностью эстераз и


БИОЛЮМИНЕСЦЕНЦИЯ
Биолюминесценцией называют свечение живых организмов. Оно происходит за счет специфичных химических
БИОЛЮМИНЕСЦЕНЦИЯ
Биолюминесценцией называют свечение живых организмов. Оно происходит за счет специфичных химических
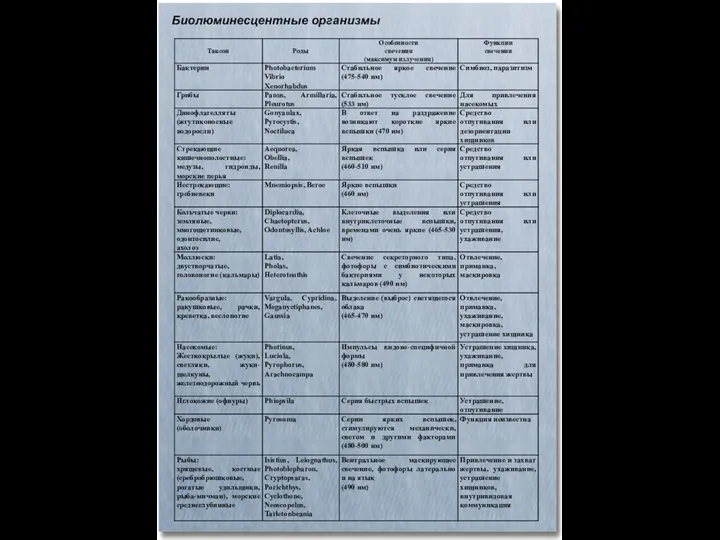
Биолюминесцентные организмы
Биолюминесцентные организмы

БИОЛЮМИНЕСЦЕНЦИЯ
Многоклеточные животные обычно излучают свет не всем телом, а только специальными
БИОЛЮМИНЕСЦЕНЦИЯ
Многоклеточные животные обычно излучают свет не всем телом, а только специальными

Биолюминесценция наблюдается вследствие дезактивации возбужденных молекул – эмиттеров. Эмиттеры возникают в
Биолюминесценция наблюдается вследствие дезактивации возбужденных молекул – эмиттеров. Эмиттеры возникают в
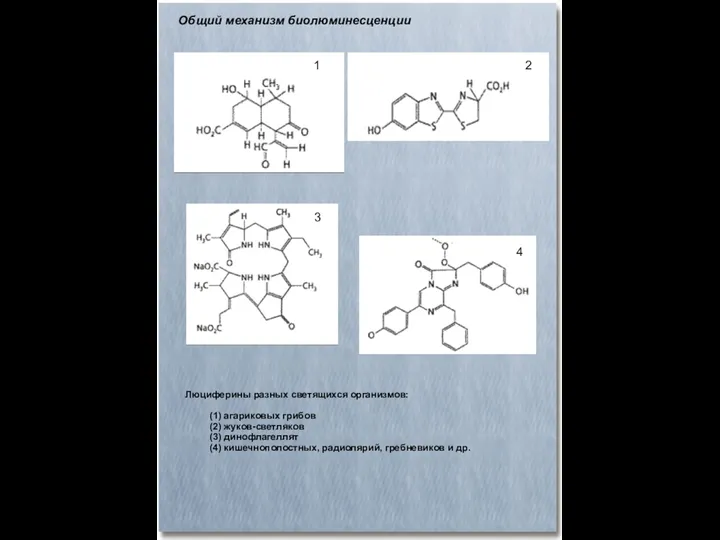
Общий механизм биолюминесценции
Люциферины разных светящихся организмов:
(1) агариковых грибов
(2) жуков-светляков
(3)
Общий механизм биолюминесценции
Люциферины разных светящихся организмов:
(1) агариковых грибов
(2) жуков-светляков
(3)

БИОЛЮМИНЕСЦЕНЦИЯ
Химически разные биолюминесцентные реакции объединяет несколько характерных черт.
1. Всем этим реакциям
БИОЛЮМИНЕСЦЕНЦИЯ
Химически разные биолюминесцентные реакции объединяет несколько характерных черт.
1. Всем этим реакциям

Биолюминесцентная реакция светляков
Различные виды светляков (насекомые семейства Lampyridae) встречаются на юге
Биолюминесцентная реакция светляков
Различные виды светляков (насекомые семейства Lampyridae) встречаются на юге

Биолюминесцентная реакция светляков
Люцифераза светляков состоит из двух субъединиц с молекулярной массой
Биолюминесцентная реакция светляков
Люцифераза светляков состоит из двух субъединиц с молекулярной массой
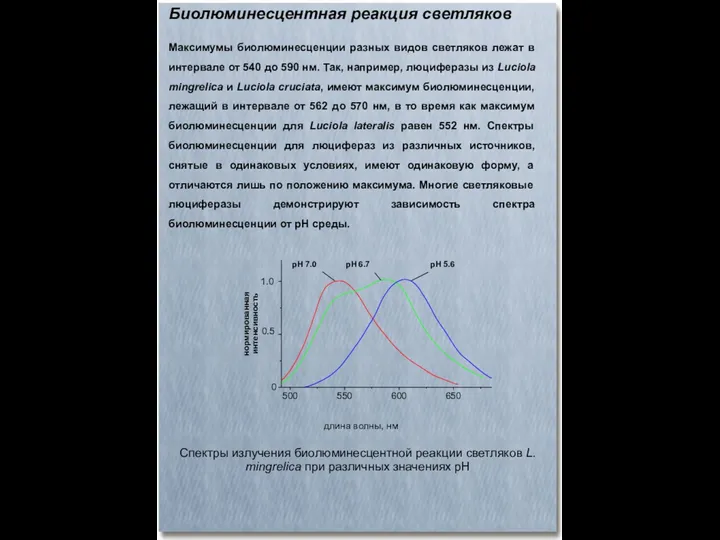
Биолюминесцентная реакция светляков
Максимумы биолюминесценции разных видов светляков лежат в интервале от
Биолюминесцентная реакция светляков
Максимумы биолюминесценции разных видов светляков лежат в интервале от

Биолюминесценция кишечнополостных
Биолюминесценция среди кишечнополостных (Coelenterata) распространена довольно широко. Известно несколько десятков
Биолюминесценция кишечнополостных
Биолюминесценция среди кишечнополостных (Coelenterata) распространена довольно широко. Известно несколько десятков
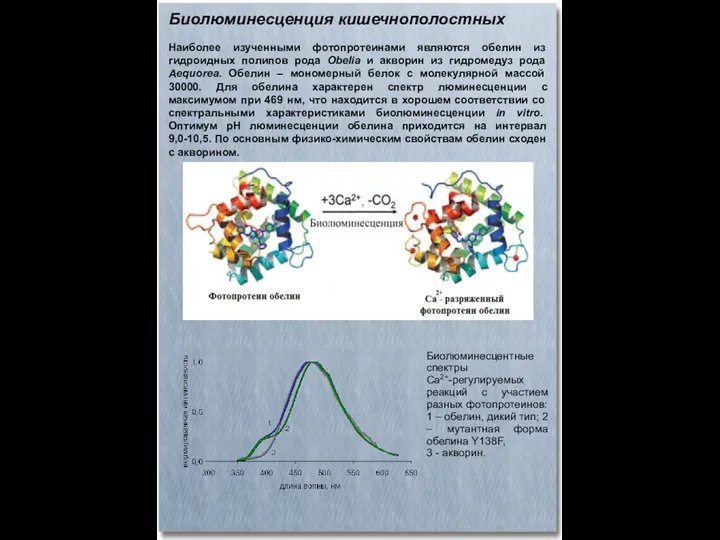
Биолюминесценция кишечнополостных
Наиболее изученными фотопротеинами являются обелин из гидроидных полипов рода Obelia
Биолюминесценция кишечнополостных
Наиболее изученными фотопротеинами являются обелин из гидроидных полипов рода Obelia

Биолюминесценция кишечнополостных
Для люминесценции морских перьев Renilla характерна стадия предварительной активации люциферина.
Биолюминесценция кишечнополостных
Для люминесценции морских перьев Renilla характерна стадия предварительной активации люциферина.

Биолюминесценция кишечнополостных
В 1962 г. О. Шимомура, Ф. Джонсон и Ю. Сайга отметили
Биолюминесценция кишечнополостных
В 1962 г. О. Шимомура, Ф. Джонсон и Ю. Сайга отметили

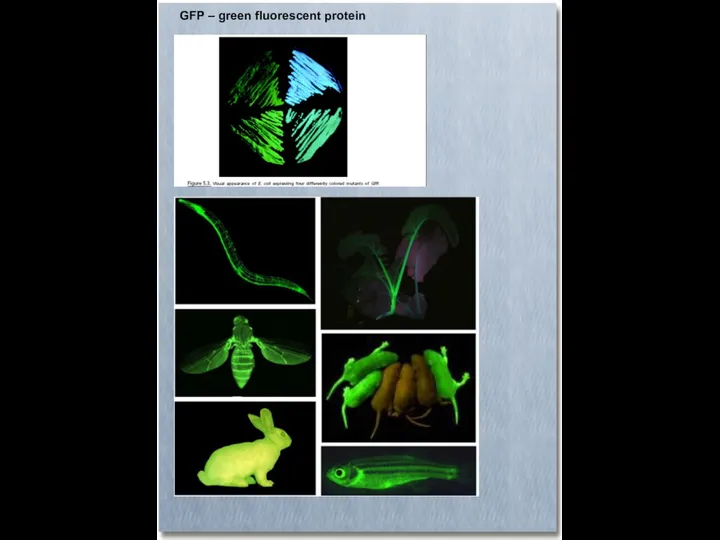
GFP – green fluorescent protein
GFP – green fluorescent protein

Биолюминесценция бактерий
Все известные в настоящее время светящиеся бактерии делятся на две
Биолюминесценция бактерий
Все известные в настоящее время светящиеся бактерии делятся на две

Биолюминесценция бактерий
Химической основой свечения бактерий является ферментативное окисление восстановленного флавинмононуклеотида ФМН⋅Н2
Биолюминесценция бактерий
Химической основой свечения бактерий является ферментативное окисление восстановленного флавинмононуклеотида ФМН⋅Н2
Биолюминесценция бактерий
Биолюминесценция бактерий

Биолюминесценция динофлагеллят
Динофлагелляты или жгутиконостцы являются основными продуцентами света в море, по
Биолюминесценция динофлагеллят
Динофлагелляты или жгутиконостцы являются основными продуцентами света в море, по

Свечение червей
Среди кольчатых червей много биолюминесцентных видов, как морских, так и
Свечение червей
Среди кольчатых червей много биолюминесцентных видов, как морских, так и

Биолюминесценция грибов
К настоящему времени описано более 40 разновидностей люминесцентных грибов. Из
Биолюминесценция грибов
К настоящему времени описано более 40 разновидностей люминесцентных грибов. Из

Биолюминесцентный анализ
Биолюминесценция in vivo и in vitro нашла широкое применение в
Биолюминесцентный анализ
Биолюминесценция in vivo и in vitro нашла широкое применение в
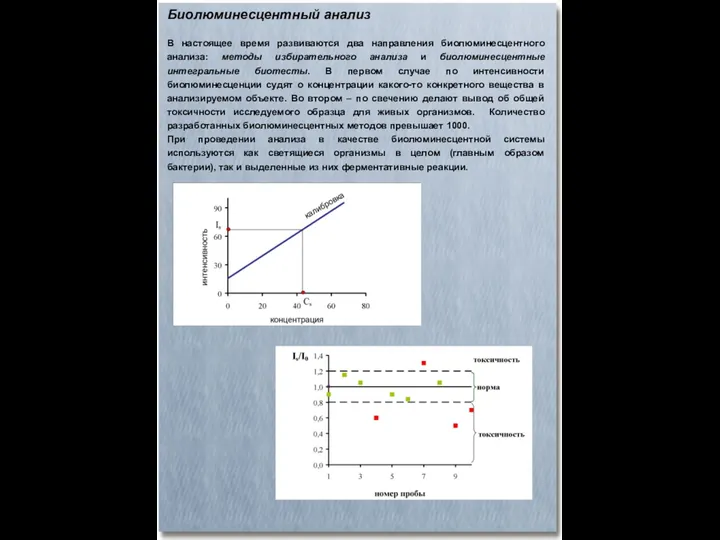
Биолюминесцентный анализ
В настоящее время развиваются два направления биолюминесцентного анализа: методы избирательного
Биолюминесцентный анализ
В настоящее время развиваются два направления биолюминесцентного анализа: методы избирательного

Биолюминесцентный анализ
Светящиеся бактерии применяются в различных областях в трех основных видах:
Биолюминесцентный анализ
Светящиеся бактерии применяются в различных областях в трех основных видах:

Биолюминесцентный анализ
Биолюминесцентный анализ

Биолюминесцентный анализ
Прогресс в области генной инженерии позволил создать новые светящиеся организмы.
Биолюминесцентный анализ
Прогресс в области генной инженерии позволил создать новые светящиеся организмы.

Биолюминесцентный анализ
Можно выделить семь основных направлений применения биолюминесцентных реакций, полученных из
Биолюминесцентный анализ
Можно выделить семь основных направлений применения биолюминесцентных реакций, полученных из

Биолюминесцентный анализ
Анализ субстратов люцифераз
Как уже упоминалось, люциферазы проявляют высокую специфичность к
Биолюминесцентный анализ
Анализ субстратов люцифераз
Как уже упоминалось, люциферазы проявляют высокую специфичность к

Биолюминесцентный анализ
Мониторинг АТФ
Биолюминесцентный анализ - наиболее специфичный, чувствительный и быстрый метод
Биолюминесцентный анализ
Мониторинг АТФ
Биолюминесцентный анализ - наиболее специфичный, чувствительный и быстрый метод

Биолюминесцентный анализ
Мониторинг АТФ
Области применения АТФ-метрии:
1. Медицина:
- обнаружение микробных инфекций в крови,
Биолюминесцентный анализ
Мониторинг АТФ
Области применения АТФ-метрии:
1. Медицина:
- обнаружение микробных инфекций в крови,

Биолюминесцентный анализ
Определение содержания ионов кальция
Концентрация ионов кальция является внутриклеточным сигналом, ответственным
Биолюминесцентный анализ
Определение содержания ионов кальция
Концентрация ионов кальция является внутриклеточным сигналом, ответственным

Детекция взаимодействия биомолекул. Иммуноанализ
Люциферазы применяются в качестве ферментов-маркеров в иммуноанализе наряду
Детекция взаимодействия биомолекул. Иммуноанализ
Люциферазы применяются в качестве ферментов-маркеров в иммуноанализе наряду

Биолюминесцентный анализ
Биолюминесцентные репортерные гены
Технология репортерных генов – одно из главных новых
Биолюминесцентный анализ
Биолюминесцентные репортерные гены
Технология репортерных генов – одно из главных новых
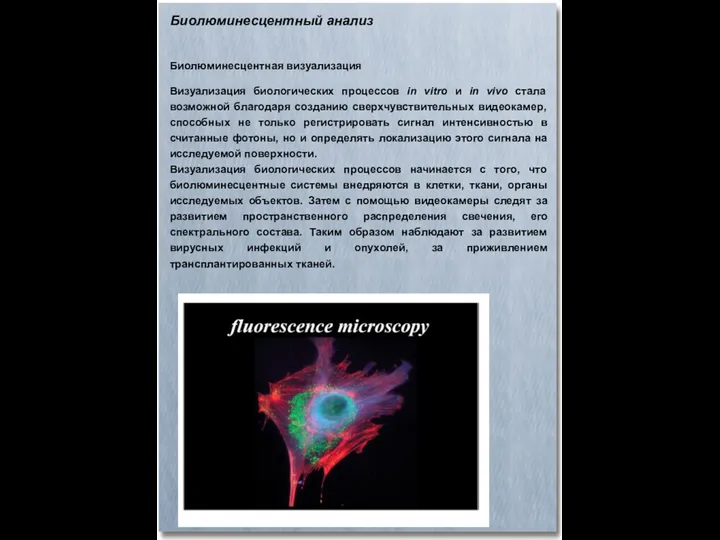
Биолюминесцентный анализ
Биолюминесцентная визуализация
Визуализация биологических процессов in vitro и in vivo стала
Биолюминесцентный анализ
Биолюминесцентная визуализация
Визуализация биологических процессов in vitro и in vivo стала
 Сила упругости. Закон Гука (7 класс)
Сила упругости. Закон Гука (7 класс) Условия плавания тел в жидкости
Условия плавания тел в жидкости Основы механики деформации горных пород. Тема 3
Основы механики деформации горных пород. Тема 3 Технология работ по ТО и ремонту механической коробки переключения передач и устройству
Технология работ по ТО и ремонту механической коробки переключения передач и устройству Теоретические основы электротехники
Теоретические основы электротехники Сила трения скольжения
Сила трения скольжения Асинхронный двигатель
Асинхронный двигатель Тепломассообмен
Тепломассообмен Презентация-игра, 7-8 класс
Презентация-игра, 7-8 класс Постоянный электрический ток
Постоянный электрический ток Внутренняя энергия. Температура
Внутренняя энергия. Температура Семей ядролық полигоны
Семей ядролық полигоны Барометр – анероид. Атмосферное давление на различных высотах (7 класс)
Барометр – анероид. Атмосферное давление на различных высотах (7 класс) Перемещение при прямолинейном равноускоренном движении
Перемещение при прямолинейном равноускоренном движении Вертолет Ми-8МТВ. Масляная система двигателя
Вертолет Ми-8МТВ. Масляная система двигателя Решение задач на вычисление силы Ампера
Решение задач на вычисление силы Ампера Устройство железнодорожных вагонов
Устройство железнодорожных вагонов Схемы интерферометров
Схемы интерферометров Электроразведка
Электроразведка Режимы работы и функциональные возможности САРП
Режимы работы и функциональные возможности САРП Магнитное поле катушки с током
Магнитное поле катушки с током Урок по физике на тему: закон Архимеда
Урок по физике на тему: закон Архимеда Урок по физике Реактивное движение
Урок по физике Реактивное движение Основы электротехники и электроники
Основы электротехники и электроники Аэродинамический нагрев
Аэродинамический нагрев Конструкция автомобиля. Сцепление
Конструкция автомобиля. Сцепление Криволинейное движение
Криволинейное движение Модульные технологии как технологии здоровьесбережения.
Модульные технологии как технологии здоровьесбережения.