- Применение методов флуоресцентных зондов для исследования белковых комплексов

Содержание
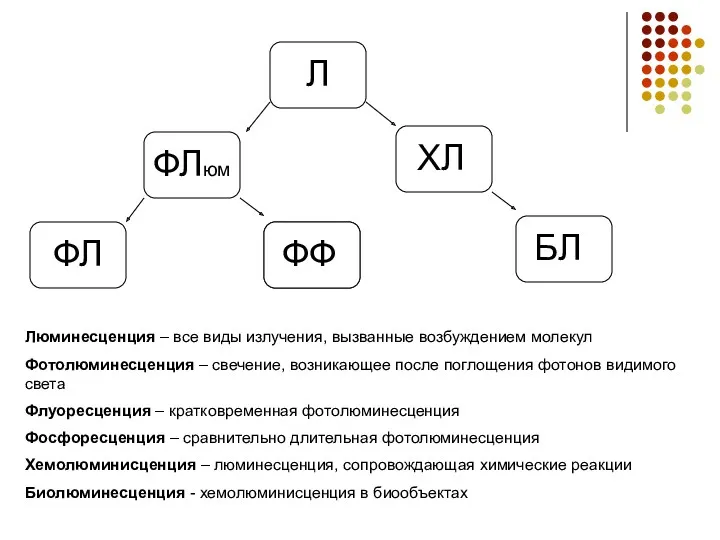
- 2. Л ФЛюм ХЛ ФЛ ФФ БЛ Люминесценция – все виды излучения, вызванные возбуждением молекул Фотолюминесценция –
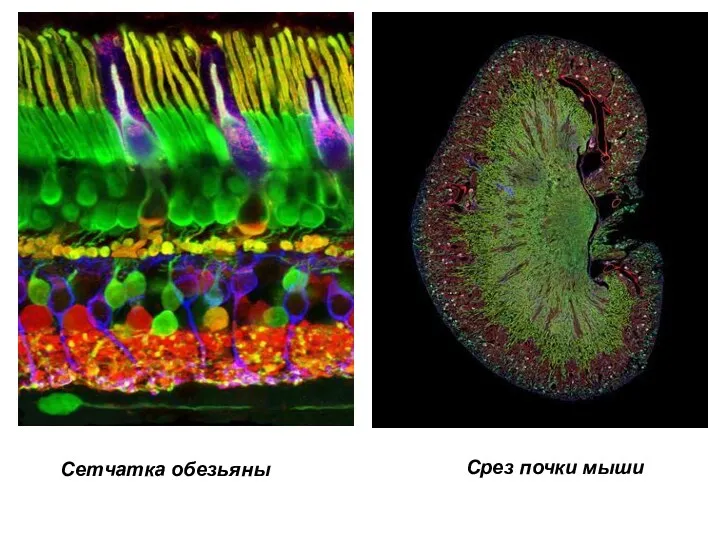
- 3. Срез почки мыши Сетчатка обезьяны
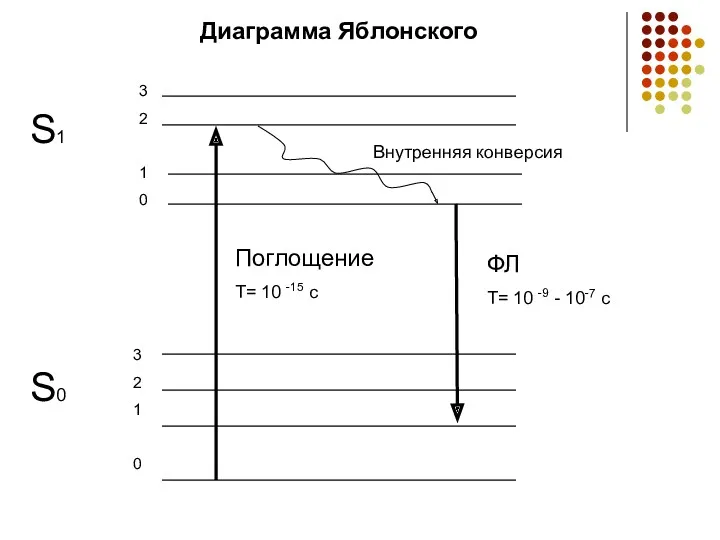
- 4. S0 3 2 1 0 3 2 1 0 S1 Внутренняя конверсия Поглощение T= 10 -15
- 5. Флуоресцентные красители различной химической природы
- 6. Флуорохромы – все виды флуоресцирующих молекул Флуорофоры – флуоресцирующие групировки крупной молекулы Молекулярный репортер – флуорохром,
- 7. Micrasterias furcata из образца озерной воды Эмбриональные фибробласты мыши (Нити актина - красный, митохондрии –зеленые и
- 8. Структуры типичных флуоресцирующих соединений
- 9. Параметры флуоресценции 1) Спектры поглощения и ФЛ – длины волн света, которые преимущественно поглощает и излучает
- 10. Параметры флуоресценции 3) Время жизни ФЛ – усредненное время в течении которого молекулы флуорофоров находятся в
- 11. Дополнительные информационные возможности ФЛ Р 5) Безызлучательная передача энергия 6) Тушение ФЛ Молекулы – тушители: кислород,
- 12. Морской червь Phascolion cryptum. Вид с нижней и передней верхней поверхности. (Синий - ДНК; зеленый -
- 13. Спектры Квантовый выход Время жизни Анизотропия БПЭ Тушение Сообщение о свойствах окружения, в котором находится ФЛ
- 15. Стволовые и пролиферирующие клетки волосяного фолликула хвоста мыши Слой нервных волокон сетчатки мыши
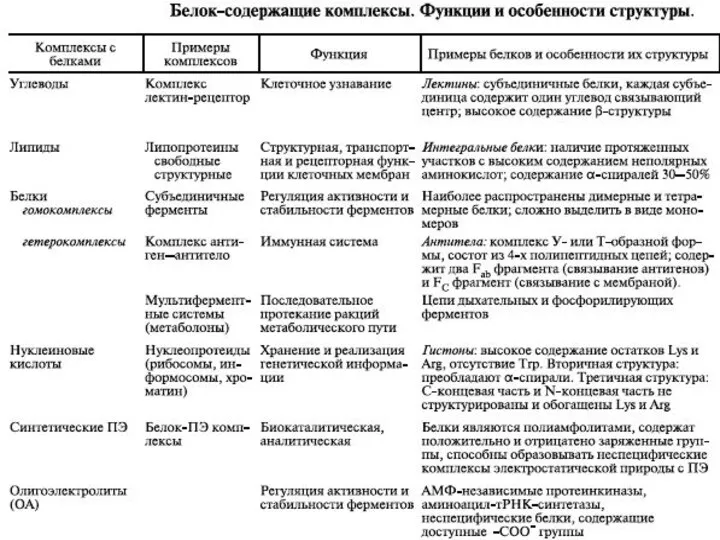
- 16. ФИЗИКО ХИМИЧЕСКИЕ МЕТОДЫ АНАЛИЗА СТРУКТУРЫ БЕЛОК СОДЕРЖАЩИХ КОМПЛЕКСОВ В РАСТВОРЕ
- 17. ФЛУОРЕСЦЕНТНЫЕ МЕТОДЫ
- 18. МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ Спектры поглощения и испускания ФЛ перилена хинина
- 19. МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ Спектр собственной флуоресценции альфа -химотрипсина (ХТ) (кривая 1). Длина волны возбуждения 296 нм.
- 20. Методы тушения флуоресценции
- 21. Флуоресцентная анизотропия Спектры поляризации 9-винилантрацена в пропиленгликоле при -50С
- 22. Водное полотоядное растение –горбатая пузычатка Utricularia gibba с одноклетчными организмами внутри
- 23. Спасибо за внимание! Мозг мыши, субвентрикулярная зона, сеть кровеносных сосудов
- 24. МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ Метод флуоресцентной спектроскопии является одним из наиболее распространенных для изучения физикохимических свойств биологических
- 25. Методы тушения флуоресценции Обычно рассматривают два вида тушения флуоресценции: статическое — связанное с образованием нефлуоресцирующего комплекса,
- 27. Скачать презентацию
Л
ФЛюм
ХЛ
ФЛ
ФФ
БЛ
Люминесценция – все виды излучения, вызванные возбуждением молекул
Фотолюминесценция – свечение, возникающее
Л
ФЛюм
ХЛ
ФЛ
ФФ
БЛ
Люминесценция – все виды излучения, вызванные возбуждением молекул
Фотолюминесценция – свечение, возникающее
Срез почки мыши
Сетчатка обезьяны
Срез почки мыши
Сетчатка обезьяны
S0
3
2
1
0
3
2
1
0
S1
Внутренняя конверсия
Поглощение
T= 10 -15 c
ФЛ
T= 10 -9 - 10-7 c
Диаграмма Яблонского
S0
3
2
1
0
3
2
1
0
S1
Внутренняя конверсия
Поглощение
T= 10 -15 c
ФЛ
T= 10 -9 - 10-7 c
Диаграмма Яблонского
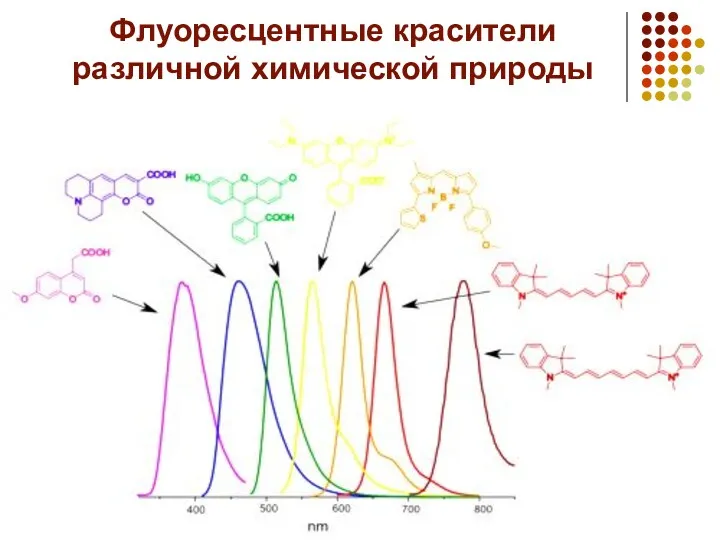
Флуоресцентные красители различной химической природы
Флуоресцентные красители различной химической природы
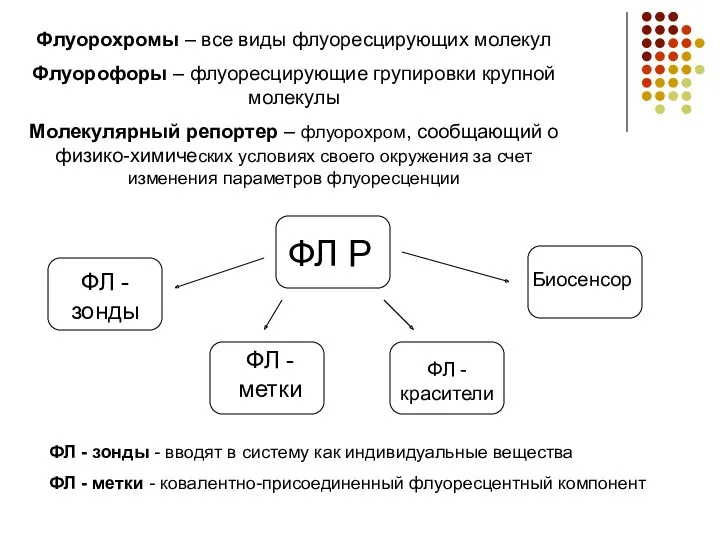
Флуорохромы – все виды флуоресцирующих молекул
Флуорофоры – флуоресцирующие групировки крупной молекулы
Молекулярный
Флуорохромы – все виды флуоресцирующих молекул
Флуорофоры – флуоресцирующие групировки крупной молекулы
Молекулярный
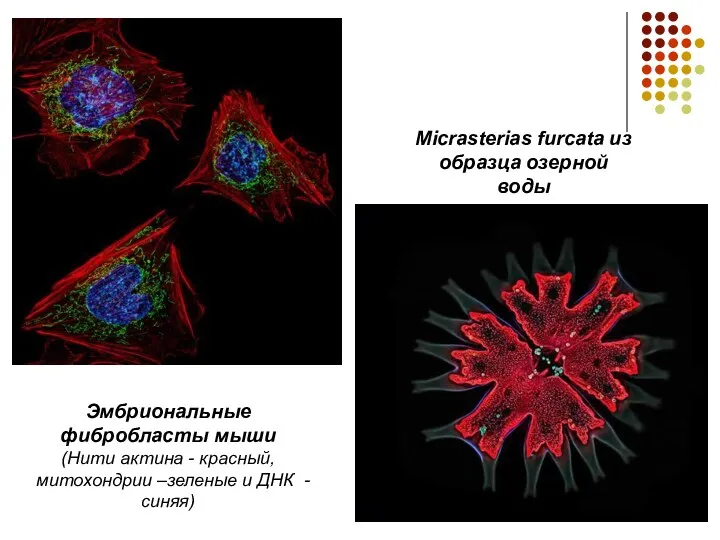
Micrasterias furcata из образца озерной воды
Эмбриональные фибробласты мыши
(Нити актина -
Micrasterias furcata из образца озерной воды
Эмбриональные фибробласты мыши
(Нити актина -
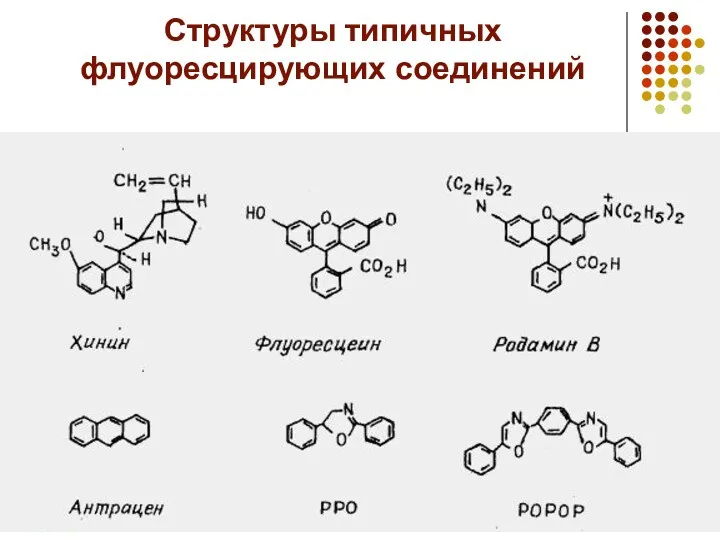
Структуры типичных флуоресцирующих соединений
Структуры типичных флуоресцирующих соединений

Параметры флуоресценции
1) Спектры поглощения и ФЛ – длины волн света, которые
Параметры флуоресценции
1) Спектры поглощения и ФЛ – длины волн света, которые

Параметры флуоресценции
3) Время жизни ФЛ – усредненное время в течении которого
Параметры флуоресценции
3) Время жизни ФЛ – усредненное время в течении которого
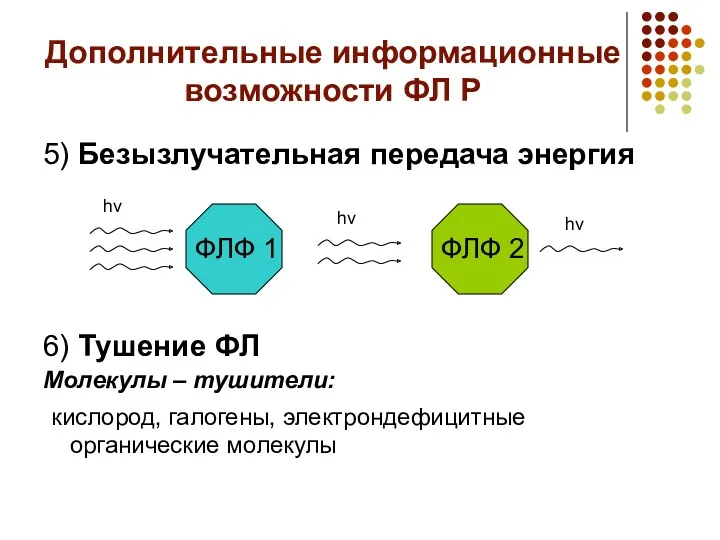
Дополнительные информационные возможности ФЛ Р
5) Безызлучательная передача энергия
6) Тушение ФЛ
Молекулы –
Дополнительные информационные возможности ФЛ Р
5) Безызлучательная передача энергия
6) Тушение ФЛ
Молекулы –
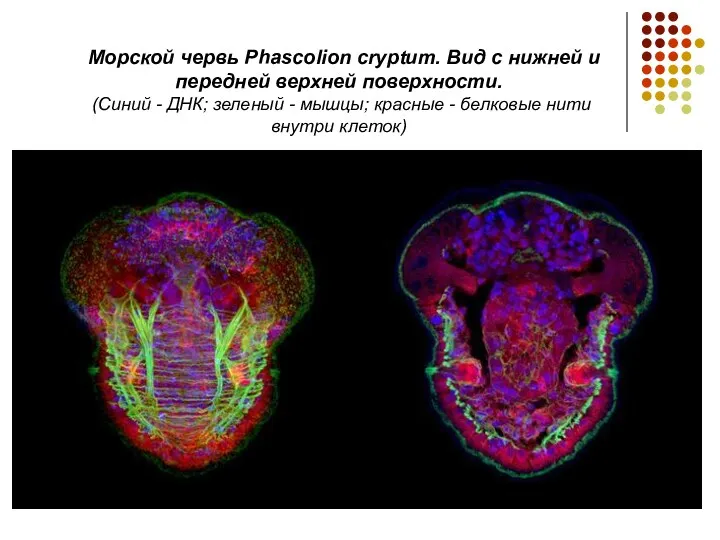
Морской червь Phascolion cryptum. Вид с нижней и передней верхней
Морской червь Phascolion cryptum. Вид с нижней и передней верхней
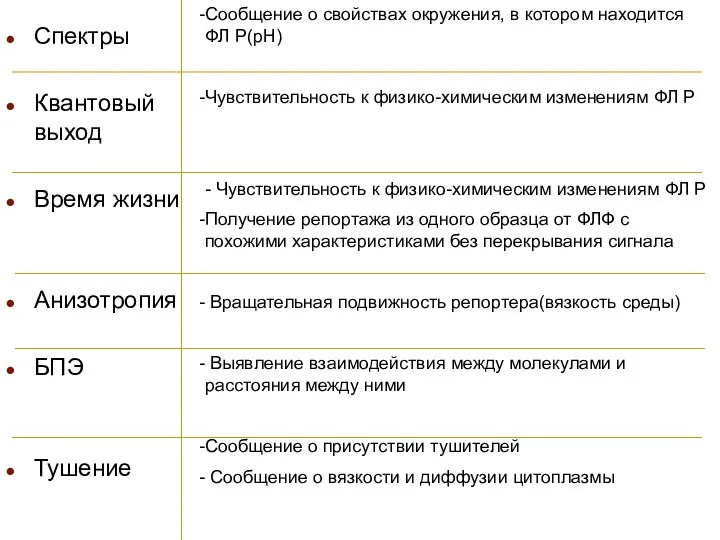
Спектры
Квантовый выход
Время жизни
Анизотропия
БПЭ
Тушение
Сообщение о свойствах окружения, в котором находится ФЛ
Спектры
Квантовый выход
Время жизни
Анизотропия
БПЭ
Тушение
Сообщение о свойствах окружения, в котором находится ФЛ
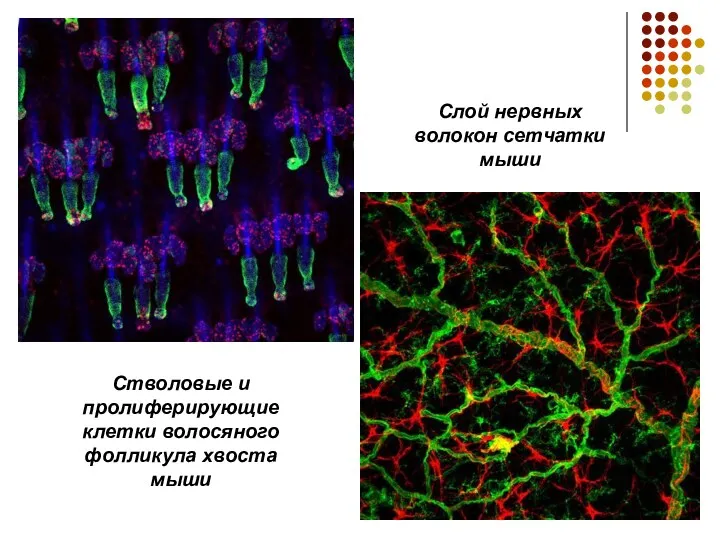
Стволовые и пролиферирующие клетки волосяного фолликула хвоста мыши
Слой нервных волокон сетчатки
Стволовые и пролиферирующие клетки волосяного фолликула хвоста мыши
Слой нервных волокон сетчатки

ФИЗИКО ХИМИЧЕСКИЕ МЕТОДЫ АНАЛИЗА
СТРУКТУРЫ БЕЛОК СОДЕРЖАЩИХ КОМПЛЕКСОВ
В РАСТВОРЕ
ФИЗИКО ХИМИЧЕСКИЕ МЕТОДЫ АНАЛИЗА
СТРУКТУРЫ БЕЛОК СОДЕРЖАЩИХ КОМПЛЕКСОВ
В РАСТВОРЕ
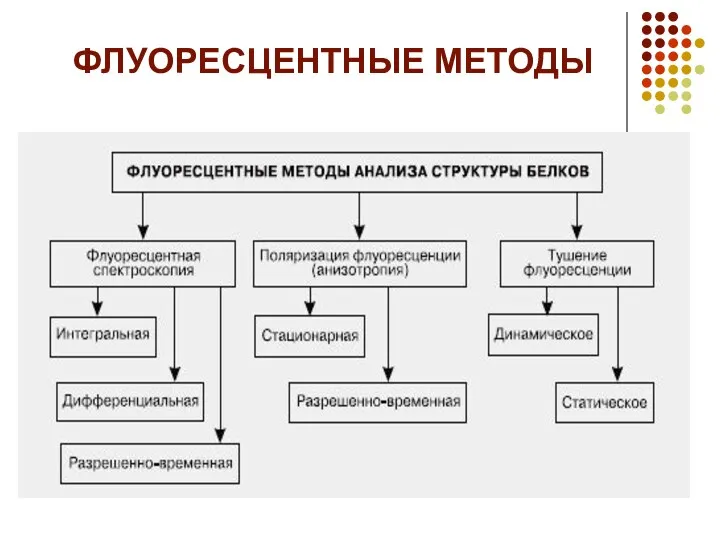
ФЛУОРЕСЦЕНТНЫЕ МЕТОДЫ
ФЛУОРЕСЦЕНТНЫЕ МЕТОДЫ
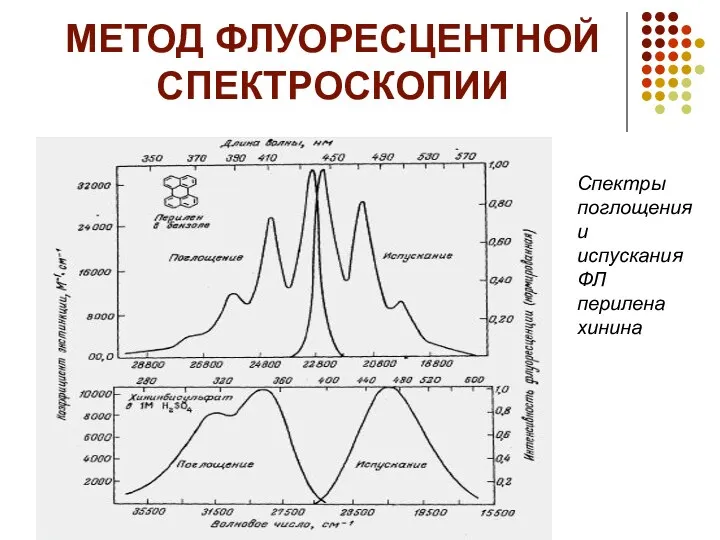
МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ
Спектры поглощения и испускания ФЛ перилена хинина
МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ
Спектры поглощения и испускания ФЛ перилена хинина
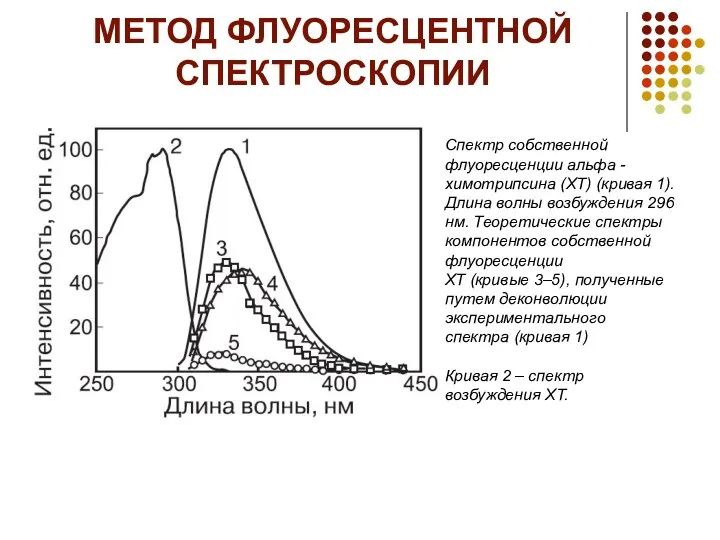
МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ
Спектр собственной флуоресценции альфа -химотрипсина (ХТ) (кривая 1).
Длина
МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ
Спектр собственной флуоресценции альфа -химотрипсина (ХТ) (кривая 1).
Длина
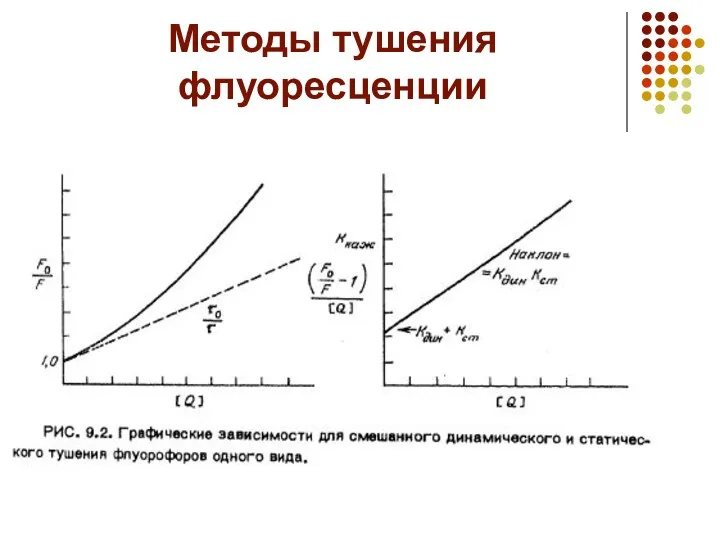
Методы тушения флуоресценции
Методы тушения флуоресценции
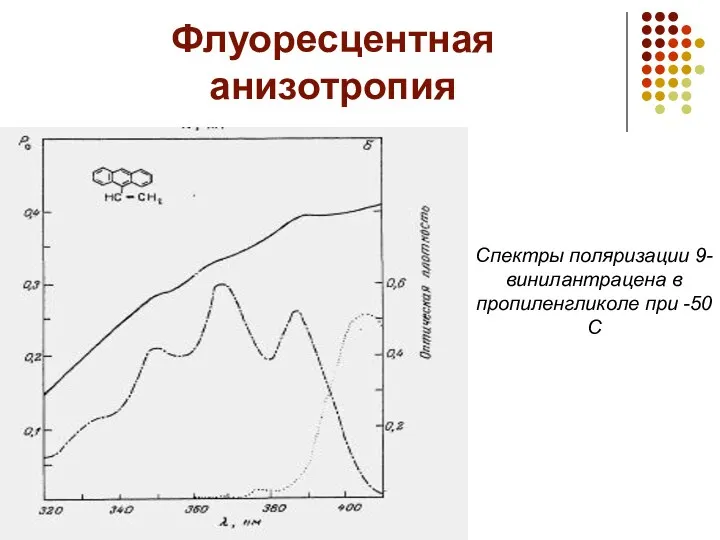
Флуоресцентная анизотропия
Спектры поляризации 9-винилантрацена в пропиленгликоле при -50С
Флуоресцентная анизотропия
Спектры поляризации 9-винилантрацена в пропиленгликоле при -50С
Водное полотоядное растение –горбатая пузычатка Utricularia gibba с одноклетчными организмами внутри
Водное полотоядное растение –горбатая пузычатка Utricularia gibba с одноклетчными организмами внутри
Спасибо за внимание!
Мозг мыши, субвентрикулярная зона, сеть кровеносных сосудов
Спасибо за внимание!
Мозг мыши, субвентрикулярная зона, сеть кровеносных сосудов

МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ
Метод флуоресцентной спектроскопии является одним из наиболее распространенных
МЕТОД ФЛУОРЕСЦЕНТНОЙ СПЕКТРОСКОПИИ
Метод флуоресцентной спектроскопии является одним из наиболее распространенных

Методы тушения флуоресценции
Обычно рассматривают два вида тушения флуоресценции: статическое —
Методы тушения флуоресценции
Обычно рассматривают два вида тушения флуоресценции: статическое —
 Теплопередача
Теплопередача Масс – спектрометрия
Масс – спектрометрия Применение фотоэффекта. Давление света. Химическое действие света. Фотография
Применение фотоэффекта. Давление света. Химическое действие света. Фотография Маноматры. Автор Максимова Наталья Сергеевна
Маноматры. Автор Максимова Наталья Сергеевна Интерференция света
Интерференция света Магнитное поле и его графическое изображение. Неоднородное и однородное магнитное поле
Магнитное поле и его графическое изображение. Неоднородное и однородное магнитное поле Інерція та інертність. Маса та імпульс тіла. Другий закон Ньютона
Інерція та інертність. Маса та імпульс тіла. Другий закон Ньютона ПЛАН-КОНСПЕКТ УРОКА Электроскоп. Проводники и непроводники электричества физика 8класс
ПЛАН-КОНСПЕКТ УРОКА Электроскоп. Проводники и непроводники электричества физика 8класс Явление смачивания
Явление смачивания Материаловедение. Технология конструкционных материалов (лекция № 4). Пластическая деформация. Механические свойства металлов
Материаловедение. Технология конструкционных материалов (лекция № 4). Пластическая деформация. Механические свойства металлов Оптика – это раздел физики, в котором изучаются световые явления
Оптика – это раздел физики, в котором изучаются световые явления Механическое движение.
Механическое движение. Основы теории цепей
Основы теории цепей Теплообменные процессы
Теплообменные процессы Урок и презентация по теме Плавание тел 7 класс
Урок и презентация по теме Плавание тел 7 класс 8Последовательное и параллельное соединение проводников
8Последовательное и параллельное соединение проводников Динамика бокового возмущенного движения
Динамика бокового возмущенного движения Теоретическая механика. Модуль 1
Теоретическая механика. Модуль 1 Антенны и распространение радиоволн
Антенны и распространение радиоволн Инфракрасная Фурье - спектроскопия
Инфракрасная Фурье - спектроскопия Презентация к уроку Сила упругости 7 класс
Презентация к уроку Сила упругости 7 класс Оптика − жарықты зерттейтін физиканың бөлімі
Оптика − жарықты зерттейтін физиканың бөлімі Тепловые явления. Решение задач
Тепловые явления. Решение задач Электрический ток в металлах
Электрический ток в металлах Электродинамика
Электродинамика Машиноведение. История создания швейной машины. Классификация швейных машин
Машиноведение. История создания швейной машины. Классификация швейных машин Агрегатные состояния вещества. Плавление и отвердевание кристаллических тел
Агрегатные состояния вещества. Плавление и отвердевание кристаллических тел Прямолинейное равноускоренное движение
Прямолинейное равноускоренное движение