- Белок биосинтезі Көмірсу Нуклеин қышқылы синтезі

Содержание
- 2. ЖОСПАР Кіріспе Негізгі бөлім Белок биосинтезі Көмірсу құрылымы Нуклеин қышқылы синтезі Қорытынды Пайдаланылған әдебиеттер
- 3. КІРІСПЕ Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесінде белок молекулалары әртүрлі биохимиялық реакцияларға
- 4. Клеткадағы зат және энергия алмасулар Тірі организмдердің ерекше қасиеті — олар зат алмасуға (метаболизмге) және көптеген
- 5. Биосинтез Көмірсу биосинтезі Белок биосинтезі Күн энергиясы Химиялық байланыстар энергиясы Күн АТФ
- 6. НЕГІЗГІ БӨЛІМ Белок биосинтезі Генетикалық код
- 7. Белок туралы алғашқы мәліметтер XVIII ғасырдан белгілі. 1745 ж. италиялық ғалым Беккори бидай ұнынан лейковина деген
- 8. Ақуыз құрамы Ақыздардың қасиеттерін олардың құрамы мен құрылымы анықтайды. Ақуыз молекуласындағы а-аминқышқылдары қалдықтарының саны әр түрлі
- 9. Молекулалық биологияның орталық догмасы
- 10. Клеткадағы рибосомалар локализациясы
- 11. Рибосома морфологиясы
- 12. Про- және эукариоттар рибосомалары Прокариоттық: 70S = 30S + 50S Эукариоттық: 80S = 40S + 60S
- 13. Рибосома суббірліктері
- 14. Белоктардың синтезі рибосомада жүреді Белоктардың синтезі үшін қажет энергия АТФ және ГТФ арқылы қамтамасыз етіледі, айта
- 15. Рибосоманың негізгі қызметі – белок молекуласын синтездеу Кіші бөлігінде трансляция (көшіріп аудару) процессі жүреді Үлкен бөлігінде
- 16. РНҚ-ның құрылысы мен функциясы Әрбір жасушада ДНҚ-ға қарағанда РНҚ-ның мөлшері 5-10 есе көп кездеседі. РНҚ-ның бастапқы
- 17. РНҚ-ның ерекшеліктері – бір полинуклеотидтік тізбектен тұрады. 2-жіпшелік РНҚ өте сирек кездеседі. Бірақ, РНҚ -РНҚ деген
- 18. РНҚ-ның түрлері. Көптеген жасушада мынадай РНҚ-ның түрлері кездеседі: рибосомды (рРНҚ) транспортты (тРНҚ) ақпараттық немесе матрицалық (мРНҚ).
- 19. и-РНҚ құрылысы ГЕНЕТИКАЛЫҚ КОД
- 20. Биологиялық код - ақуыздардың бірінші реттік құрылысы туралы ақпаратты нуклеин қышқылдары арқылы анықтау әдісі Генетикалық кодтың
- 21. Төрт түрлі нуклеотидтен 64 кодон түзіледі, ондағы 61-і амин қьшқылдарын кодтайды және бірі метионинді кодтайтын АУГ-триплеті
- 22. Маңызы қызметі: триплеттілігі, биологиялық кодта код саны - 3. Үш нуклеотидтік қалдықтан бір амин қышкылы қолданылады.
- 23. 1. Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос тізбегінің біреуінің комплементарлы көшірмесі болып табылатын и-РНҚ синтезі
- 24. Белок синтезі сатылары:
- 25. ТРАНСКРИПЦИЯ (лат. transcrіptіo – қайта көшіріп жазу) – тірі клеткалардағы рибонуклеин қышқылының биосинтез процесі. Ол дезоксирибонуклеин
- 27. ТРАНСЛЯЦИЯ — полипептид тізбегінің гендегі иРНҚ негізінде ақпаратқа сай түзілуі. Трансляция болашақ белокқа тән иРНҚ-на жазылған
- 28. Трансляцияның жалпы кезеңдерінің сызбанұсқасы
- 29. Бұл пептид синтезінің басталуы. Осы жерде рибосомалардың не суббірліктерінің бірлесуі және алғашқы амино-ацил т-РНҚ-ң қосылуы жүреді.
- 30. Инициация кезінде РНҚ полимераза промотормен қосылып ДНҚ шынжырын тарқатады. М-РНҚ бір шынжырлы. ДНҚ –да РНҚ-ның орналасуы
- 31. Элонгация кезеңі немесе РНҚ – транскриптатының ұзаруы. Бұл кезде ДНҚ тарқатылып және нуклеосома бұзылады. Синтез жылдамдылығы
- 32. Терминация
- 33. Трансляцияның 4-ші кезеңі – Терминация, яғни синтездің бітуі, аяқталу кезеңі, керекті эаттар: 1)АТФ; 2) Белок синтезінің
- 34. Трансляцияның 5-ші кезеңі - кеңістіктегі полипептидтік тізбектің орналасуы және процессинг. Бұл кезеңде полипептид өзінің кеңістіктегі екінші-
- 35. Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция (яғни белок синтезі) кезеңінде жүреді. Бұл
- 36. НЕГІЗГІ БӨЛІМ Көмірсулар түрлері Көмірсулардың қасиеттері
- 37. Көмірсулар – барлық тірі ағзада энергия беретін органикалық зат және көміртек көзі жалпы формуласы Cn(H2O)m
- 38. Таза күйінде қызылша қанты 1747 жылы неміс химигі А. Маргграф ашқан болатын 3. 1837жылы швед химигі
- 39. Көмірсулар жіктемесі
- 40. Көмірсулар биологиялық қызметтері Энергетикалық Тіректік қызмет Қорғаныштық (майлау)қызметі Гидросмостық және ионды реттеушілік Кофакторлық
- 41. Моносахаридтердің физикалық қасиеттері Түссіз Кристалды заттар Суда ериді Дәмі тәтті Cn(H2O)m
- 42. Моносахаридтер изомериясы. Оптикалық изомерия Көмірсу молекуласы оптикалы белсенді және құрамынды бірнеше ассиметриялы көміртегі атомдары бар. ОН-
- 43. Моносахаридтертің химиялық қасиеттері 1. Альдегидтер мен кетондар сияқты сутегі атомдары әсерінен оңай тотықсызданады. C6H12O6 + НАД-Н
- 44. Моносахаридтертің химиялық қасиеттері
- 45. 3. Моносахаридтер метаболизмге негізінен фосфор қышқылының эфирі (фосфоэфирлер) түрінде қатысады. Мұнда фосфат тобының доноры қызметін АТФ
- 46. 5. Эпимеризация – изомерияның бір түрі. Молекуланың хиральдық орталықтарының біреуінің қарама-қарсысына өзгеруі (айналуы, бұрылуы)
- 47. Моносахарид - пентоздар Рибоза С5Н10О5 маңызы: РНҚ құрамына кіреді, витаминдердің В тобының, ферменттердің АТФ көзі Дезоксирибоза
- 48. Глюкоза С6Н12О6 маңызы: Энергия көзі; гликозидтердің құрамына кіреді, бос күйінде өсімдіктер, жануарлар және адам тканьдерінде болады;
- 49. Фруктоза С6Н12О6 Маңызы: Жеміс қанты; бос күйінде бал құрамында болады; сахарозамен салыстырғанда тәттілеу, байланысқан күйде сахарозада,
- 50. Дисахаридтер Молекулалық формуласы С12Н22О11, судың бір молекуласын бөліп шығару арқылы гликозидтік байланыспен байланысқан моносахаридтердің екі молекуласынан
- 51. Дисахаридтер Сахароза «қызылша қанты» Глюкоза + фруктоза Бос карбонильдік тобы жоқ, сондықтан оның тотықсыздандырғыштық қасиеті болмайды;
- 52. Мальтоза Глюкоза + Глюкоза Крахмал мен гликогеннің шала гидролизденуі нәтижесінде алынатын өнім болап табылады. Ячмень солодында,
- 53. Полисахаридтер Күрделі көмірсулар, олар гликозидтік байланыстармен жалғасқан 1000 дейін, тіпті одан да көп моносахаридтерден құралады. Полисахаридтерге
- 54. Өсімдіктердің негізгі қоры болатын көмірсу және адамға азық-түлік, малға жемшөп болатын аса маңызды полисахарид. Екі бөліктен
- 55. Крахмал Амилоза Амилопектин амилозы (10-20%) және амилопектина (80-90%), D-глюкозаға дейін гидролизденеді. n = 1000-3000 n =
- 56. Фотосинтез процесіне күннің көзі қажет Фотосинтез процесі жасыл өсімдіктерде өтеді Өсімдіктер СО2 газын сіңіреді О2 бөліп
- 57. Гликоген Жануарлардың полисахарид қоры. Бауыр мен бұлшық еттерде, қанда да жиналады. Ол – организмге қуат беретін
- 58. Целлюлоза Өсімдіктер жасушасының негізгі материалы. Таза целлюлозаның талшықтанған құрылымы бар және органикалық еріткіштерде ерімейді. Ол Швейцар
- 60. Хитин целлюлозаға жақын; ол саңырауқұлақтардың кейбір түрлерінде кездеседі; және де кейбір жануарлардың сыртқы қаңқасының маңызды бөлігі
- 61. Көмірсудың қолданылуы
- 62. Көмірсулардың қорытылуы
- 63. НЕГІЗГІ БӨЛІМ Нуклеин қышқылы синтезі Нуклеин қышқылы қасиеттері
- 64. Швейцария биологы Э.Мишер (1868) ірің жасушалары ядросынан құрамында фосфоры бар зат бөліп алды.Ядродан бөліп алынған бұл
- 65. РНҚ жасуша ядросында және цитоплазмада (РНҚ-ның 90%-ға жуығы цитоплазмада) орналасқан және ақуыз синтезінің белсенділігі РНҚ мөлшеріне
- 66. Нуклеин қышқылы - пішіні жіп тәрізді, жоғарғы молекулалы, ерітінділерінің тұтқырлығы жоғары қосылыс. Адамның 23 жұп хромасомасында
- 67. ДНҚ геннің материалдық негізі, ол геннің келесі қасиеттеріне ие болады: 1. Өз-өзін дәл қайталау, соның арқасында
- 68. Дезоксирибонуклеотидтер - ағзадағы ДНҚ-ң түзілуіне қатысады. Рибонуклеотидтер РНҚ- түзілуіне жүмсалады. Рибонуклеотидтердің кейбірі трансферазалық реакцияларда коферменттік қызмет
- 69. Нуклеин қышқылдары - нуклеотид полимерлерінен құралған. Нуклеотидтердің химиялық құрамы - пиримидиндік немесе пуриндік негіздер, пентоз, фосфор
- 70. Нуклепротеид Ақуызды бөлігі ақуызды емес бөлігі Полипептид Амин қышқылы Азоттық негіздер Мононуклеидтер Пуриндік Пириминдиндік Көмірсулар H3PO4
- 71. 1) ДНҚ- дезоксирибонуклеин қышқылдары, толық гидроизденгенде аденин, гуанин, цитозин, тимин, дезоксирибоза және фосфор қышқылына; 2)РНҚ-рибонуклеин қышқылы,
- 72. Алтымүшелі бір гетероатомды гетероциклдердің өкілі – пиридин; алтымүшелі екі гетероатомды гетероциклдердің өкілі – пиримидин. Пиридин пиримидин
- 73. Пиримидин туындылары: урацил, цитозин, тимин: Урацил – 2,6-диоксипиримидин, екі таутомерлі формада Цитозин – 2-окси-6-аминопиримидин Тимин –
- 74. Пиримидин және имидазол ядролары конденсирленгенде пурин гетероциклі түзіледі. Пурин Пурин туындылары аденин және гуанин: Аденин гуанин
- 75. Дезоксирибоза Рибоза
- 76. Нуклеопротеидтердің гидролизденуі сатылай жүреді және бұл тізбек кез-келген сатыда тоқтауы мүмкін. Азоттық негізді трифосфор қышқылы ағзада
- 78. РНҚ (рибонуклеозидтер) құрамына кіретін нуклеозидтер: аденил қышқылы (аденозин), гуанин қышқылы (гуанозин), цитидил қышқылы (цитидин) т.б.
- 79. Нуклеотидтердің атауы: нуклеин негіздердің тривиалды атауына сәйкес жалғаулар “идин” пиримидиндік және “озин”пуринді нуклеозидтерге жалғанады: Цитозин+рибоза =
- 81. Нуклеин қышқылдарының құрамындағы мононуклеотидтер бір-бірімен оттектік көпірше арқылы, яғни пентоздардың гидроксил тобы (-ОН) (үшінші көміртегінің жанындағы)
- 82. Нуклеин қышқылдарының бірінші реттік құрылымы: мононуклеотидтер бір-бірімен «3́-5́́ байланыс» арқылы, мұнда фосфор қышқылының қалдығы 5′- фосфатты
- 83. Америка ғалымдары Дж.Уотсон және ағылшын физигі Фрэнсис Крик (1953 ж.) ұсынған ДНҚ молекуласының екіншілік құрылымы қос
- 84. ДНҚ-ның екінші реттік құрылымында молекула оралым спираль тәрізді тізбек құрады. ДНҚның екіншілік құрылымында пуриндік негіз міндетті
- 86. Бір тізбектегі нуклеотидтің орналасуы өз бетінмен еркінірек болғанымен, екінші тізбектегі нуклеотидтердің орны бірінші тізбектің нуклеотидтік құрамына
- 87. Әрбір тізбек төрт нуклеотидтің: аденин = тимин, цитозин ≡ гуанин байланыс арқылы байланысады. ДНҚ –ның екіншілік
- 89. ДНҚ синтезіне қатысатын негізгі фермент - ДНҚ-на тәуелді ДНҚ полимераза. ДНҚ молекуласының синтезіне қажетті негізгі компоненттер:
- 90. ДНҚ синтезіне қатысатын ферменттер: 1. ДНҚ-на тәуелді ДНҚ-полимеразасы (I, П, III). III ДНҚ-полимеразасы элонгацияға тікелей қатысатын
- 91. ДНҚ-ның биосинтезі үш кезеңнен түрады: 1. Инициация. ДНҚ-на тәуелді РНҚ-полимеразасы жұмыс істейді, сннтездің басы. ДНҚ-ны екі
- 92. ҚОРЫТЫНДЫ Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесінде белок молекулалары әртүрлі биохимиялық реакцияларға
- 94. Скачать презентацию

ЖОСПАР
Кіріспе
Негізгі бөлім
Белок биосинтезі
Көмірсу құрылымы
Нуклеин қышқылы синтезі
Қорытынды
Пайдаланылған әдебиеттер
ЖОСПАР
Кіріспе
Негізгі бөлім
Белок биосинтезі
Көмірсу құрылымы
Нуклеин қышқылы синтезі
Қорытынды
Пайдаланылған әдебиеттер

КІРІСПЕ
Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесінде белок
КІРІСПЕ
Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесінде белок

Клеткадағы зат және энергия алмасулар
Тірі организмдердің ерекше қасиеті — олар зат
Клеткадағы зат және энергия алмасулар
Тірі организмдердің ерекше қасиеті — олар зат
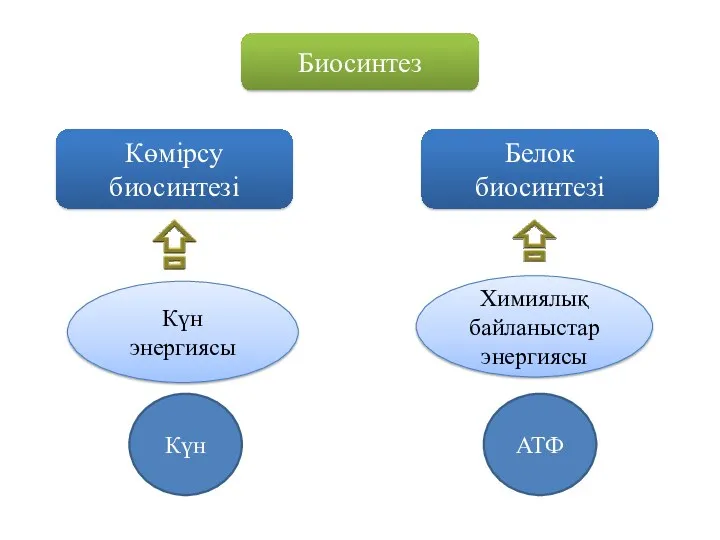
Биосинтез
Көмірсу биосинтезі
Белок
биосинтезі
Күн энергиясы
Химиялық байланыстар энергиясы
Күн
АТФ
Биосинтез
Көмірсу биосинтезі
Белок
биосинтезі
Күн энергиясы
Химиялық байланыстар энергиясы
Күн
АТФ
НЕГІЗГІ БӨЛІМ
Белок биосинтезі
Генетикалық код
НЕГІЗГІ БӨЛІМ
Белок биосинтезі
Генетикалық код

Белок туралы алғашқы мәліметтер XVIII ғасырдан белгілі. 1745 ж. италиялық ғалым
Белок туралы алғашқы мәліметтер XVIII ғасырдан белгілі. 1745 ж. италиялық ғалым

Ақуыз құрамы
Ақыздардың қасиеттерін олардың құрамы мен құрылымы анықтайды. Ақуыз молекуласындағы
Ақуыз құрамы
Ақыздардың қасиеттерін олардың құрамы мен құрылымы анықтайды. Ақуыз молекуласындағы
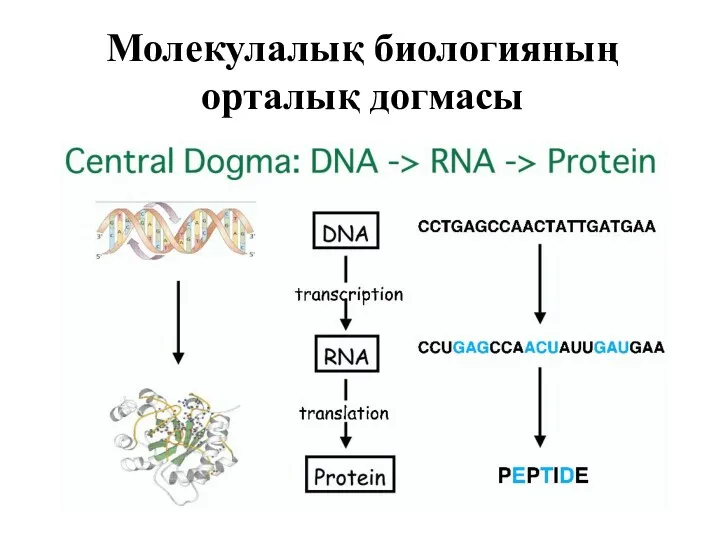
Молекулалық биологияның орталық догмасы
Молекулалық биологияның орталық догмасы
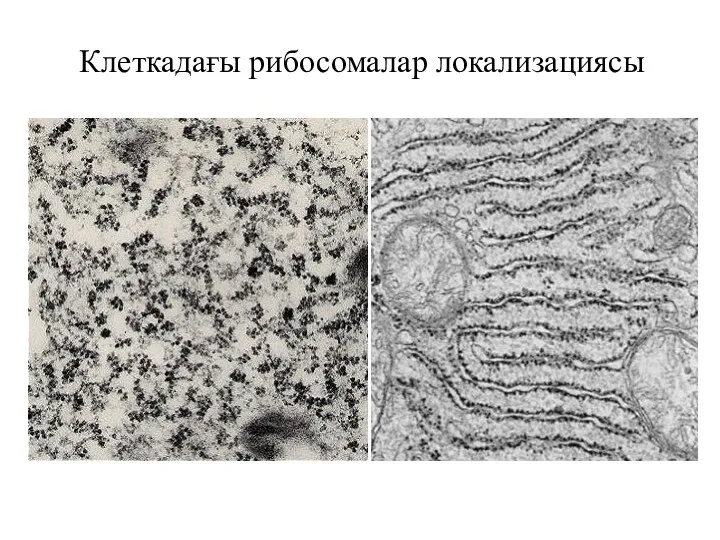
Клеткадағы рибосомалар локализациясы
Клеткадағы рибосомалар локализациясы
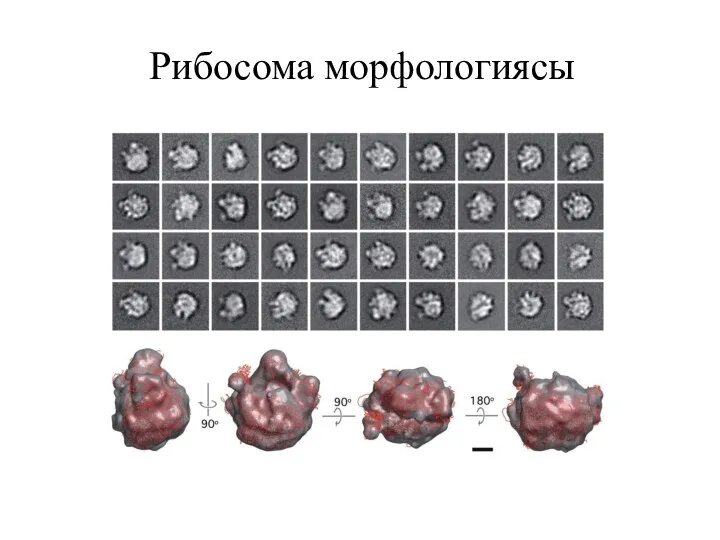
Рибосома морфологиясы
Рибосома морфологиясы

Про- және эукариоттар рибосомалары
Прокариоттық:
70S = 30S + 50S
Эукариоттық:
80S = 40S +
Про- және эукариоттар рибосомалары
Прокариоттық:
70S = 30S + 50S
Эукариоттық:
80S = 40S +
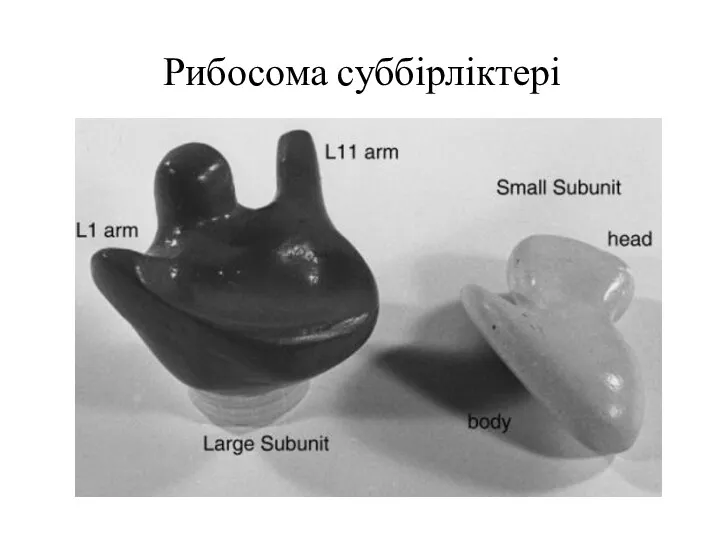
Рибосома суббірліктері
Рибосома суббірліктері

Белоктардың синтезі рибосомада жүреді
Белоктардың синтезі үшін қажет энергия АТФ және
Белоктардың синтезі рибосомада жүреді
Белоктардың синтезі үшін қажет энергия АТФ және
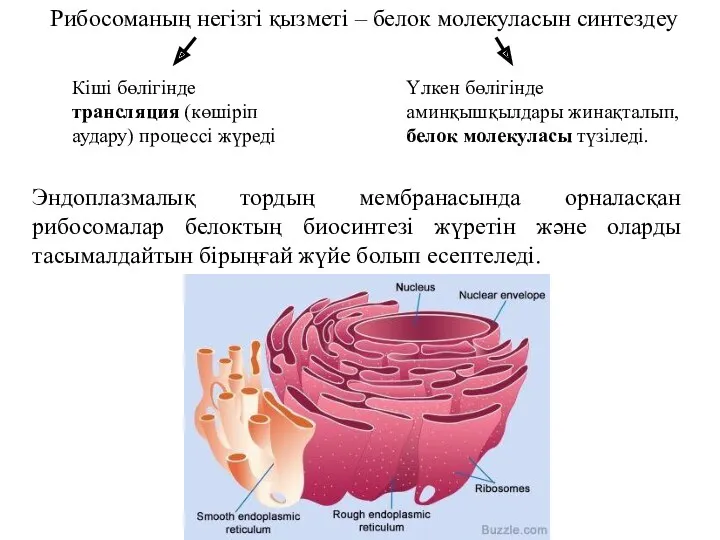
Рибосоманың негізгі қызметі – белок молекуласын синтездеу
Кіші бөлігінде трансляция (көшіріп
Рибосоманың негізгі қызметі – белок молекуласын синтездеу
Кіші бөлігінде трансляция (көшіріп

РНҚ-ның құрылысы мен функциясы
Әрбір жасушада ДНҚ-ға қарағанда РНҚ-ның мөлшері 5-10 есе
РНҚ-ның құрылысы мен функциясы
Әрбір жасушада ДНҚ-ға қарағанда РНҚ-ның мөлшері 5-10 есе

РНҚ-ның ерекшеліктері – бір полинуклеотидтік тізбектен тұрады.
2-жіпшелік РНҚ өте сирек
РНҚ-ның ерекшеліктері – бір полинуклеотидтік тізбектен тұрады.
2-жіпшелік РНҚ өте сирек

РНҚ-ның түрлері.
Көптеген жасушада мынадай РНҚ-ның түрлері кездеседі:
рибосомды (рРНҚ)
транспортты (тРНҚ)
РНҚ-ның түрлері.
Көптеген жасушада мынадай РНҚ-ның түрлері кездеседі:
рибосомды (рРНҚ)
транспортты (тРНҚ)

и-РНҚ құрылысы
ГЕНЕТИКАЛЫҚ КОД
и-РНҚ құрылысы
ГЕНЕТИКАЛЫҚ КОД

Биологиялық код - ақуыздардың бірінші реттік
құрылысы туралы ақпаратты нуклеин
Биологиялық код - ақуыздардың бірінші реттік
құрылысы туралы ақпаратты нуклеин

Төрт түрлі нуклеотидтен 64 кодон түзіледі,
ондағы 61-і амин қьшқылдарын кодтайды
Төрт түрлі нуклеотидтен 64 кодон түзіледі,
ондағы 61-і амин қьшқылдарын кодтайды

Маңызы қызметі: триплеттілігі, биологиялық кодта код саны - 3. Үш нуклеотидтік
Маңызы қызметі: триплеттілігі, биологиялық кодта код саны - 3. Үш нуклеотидтік

1. Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос тізбегінің біреуінің комплементарлы
1. Ядролық кезең немесе транскрипция. Мұнда ДНҚ қос тізбегінің біреуінің комплементарлы
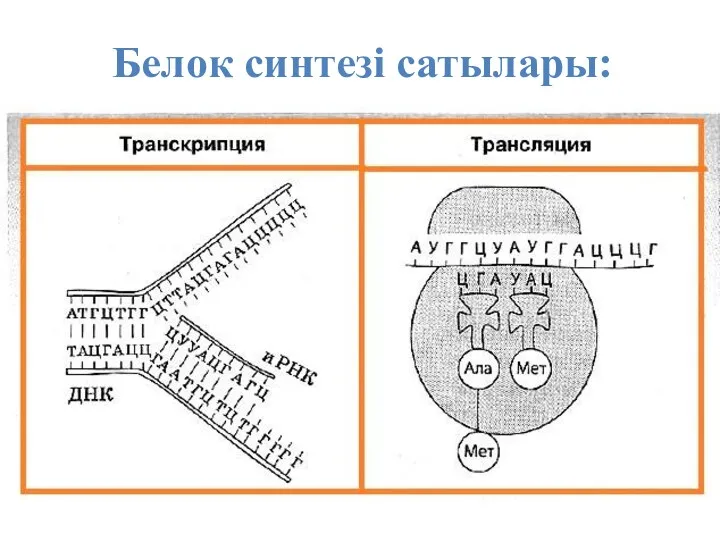
Белок синтезі сатылары:
Белок синтезі сатылары:
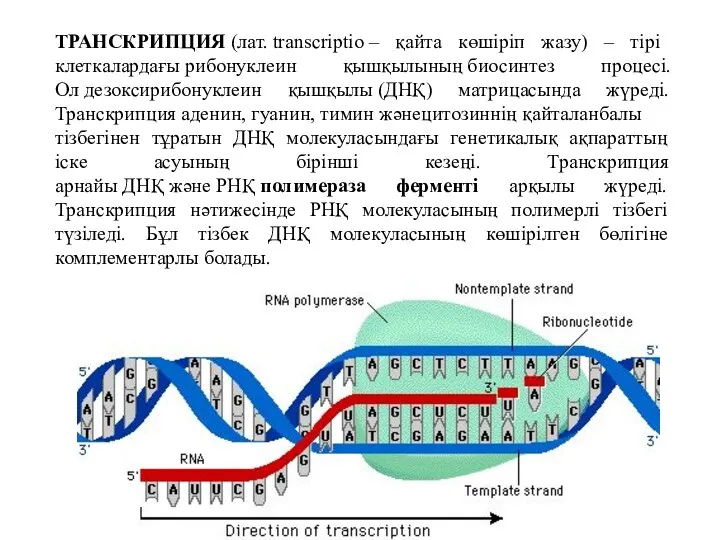
ТРАНСКРИПЦИЯ (лат. transcrіptіo – қайта көшіріп жазу) – тірі клеткалардағы рибонуклеин қышқылының биосинтез процесі. Ол дезоксирибонуклеин қышқылы (ДНҚ)
ТРАНСКРИПЦИЯ (лат. transcrіptіo – қайта көшіріп жазу) – тірі клеткалардағы рибонуклеин қышқылының биосинтез процесі. Ол дезоксирибонуклеин қышқылы (ДНҚ)
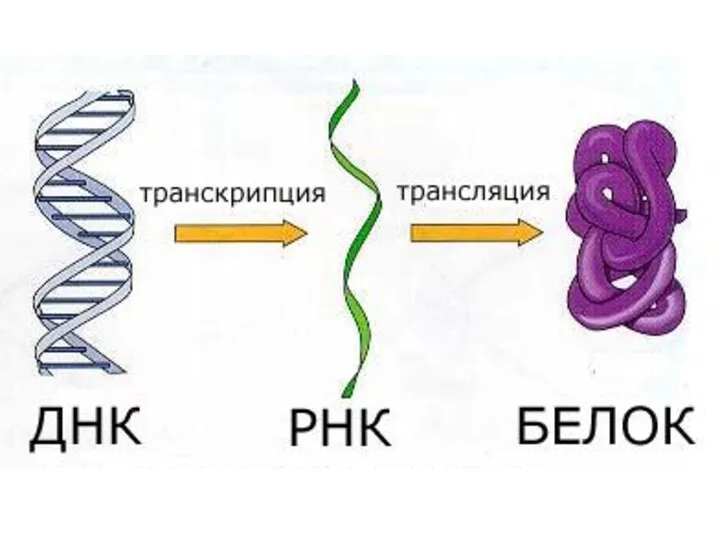
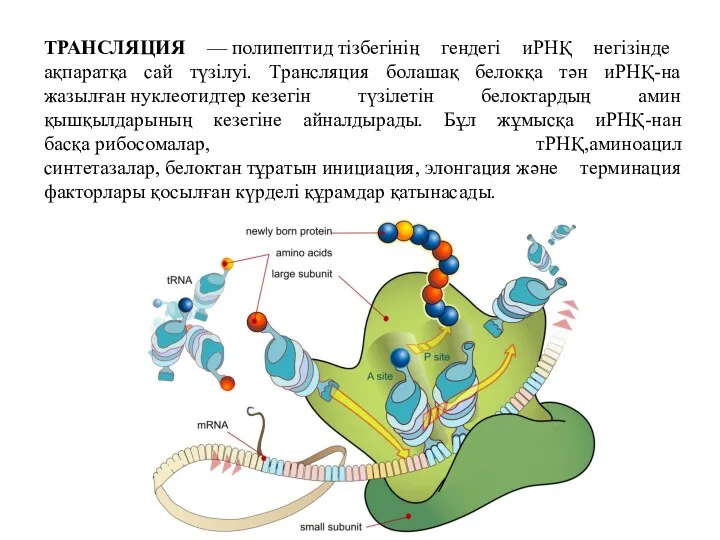
ТРАНСЛЯЦИЯ — полипептид тізбегінің гендегі иРНҚ негізінде ақпаратқа сай түзілуі. Трансляция болашақ белокқа
ТРАНСЛЯЦИЯ — полипептид тізбегінің гендегі иРНҚ негізінде ақпаратқа сай түзілуі. Трансляция болашақ белокқа
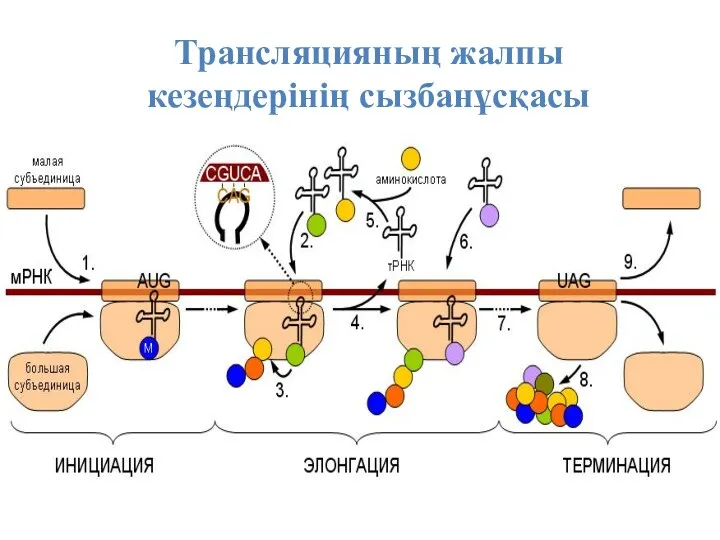
Трансляцияның жалпы кезеңдерінің сызбанұсқасы
Трансляцияның жалпы кезеңдерінің сызбанұсқасы
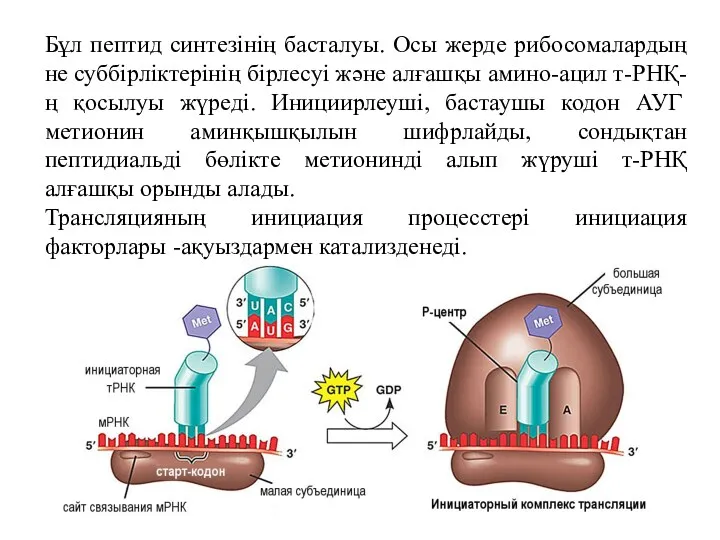
Бұл пептид синтезінің басталуы. Осы жерде рибосомалардың не суббірліктерінің бірлесуі және
Бұл пептид синтезінің басталуы. Осы жерде рибосомалардың не суббірліктерінің бірлесуі және

Инициация кезінде РНҚ полимераза промотормен қосылып ДНҚ шынжырын тарқатады. М-РНҚ бір
Инициация кезінде РНҚ полимераза промотормен қосылып ДНҚ шынжырын тарқатады. М-РНҚ бір
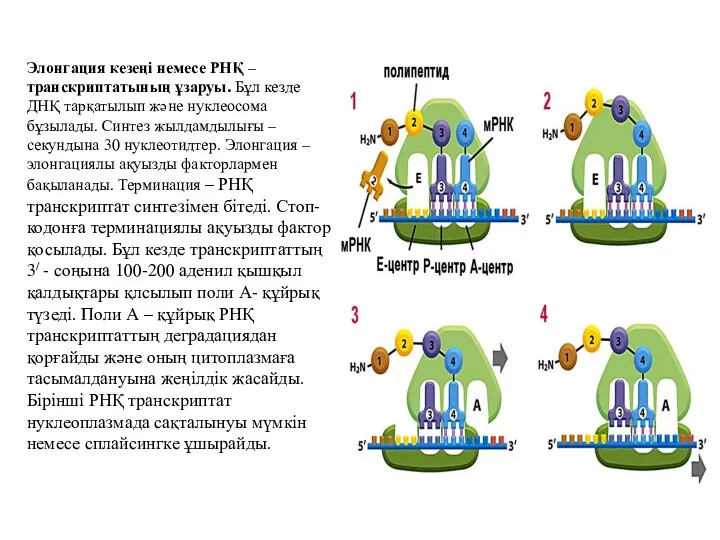
Элонгация кезеңі немесе РНҚ – транскриптатының ұзаруы. Бұл кезде ДНҚ тарқатылып
Элонгация кезеңі немесе РНҚ – транскриптатының ұзаруы. Бұл кезде ДНҚ тарқатылып
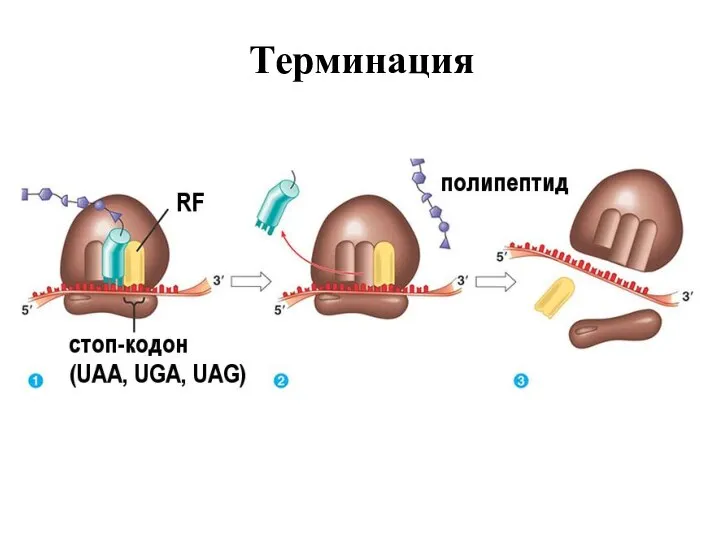
Терминация
Терминация

Трансляцияның 4-ші кезеңі – Терминация, яғни синтездің бітуі, аяқталу кезеңі, керекті
Трансляцияның 4-ші кезеңі – Терминация, яғни синтездің бітуі, аяқталу кезеңі, керекті

Трансляцияның 5-ші кезеңі - кеңістіктегі полипептидтік тізбектің орналасуы және процессинг. Бұл
Трансляцияның 5-ші кезеңі - кеңістіктегі полипептидтік тізбектің орналасуы және процессинг. Бұл

Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция (яғни
Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция (яғни
НЕГІЗГІ БӨЛІМ
Көмірсулар түрлері
Көмірсулардың қасиеттері
НЕГІЗГІ БӨЛІМ
Көмірсулар түрлері
Көмірсулардың қасиеттері
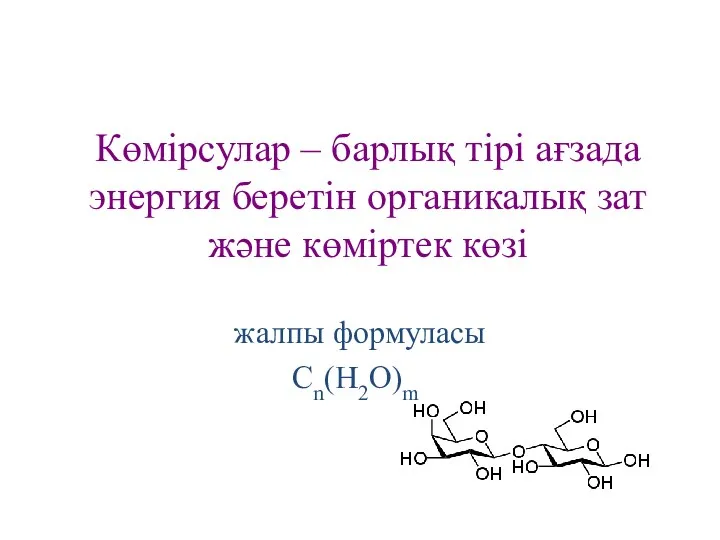
Көмірсулар – барлық тірі ағзада энергия беретін органикалық зат және көміртек
Көмірсулар – барлық тірі ағзада энергия беретін органикалық зат және көміртек

Таза күйінде қызылша қанты 1747 жылы неміс химигі А. Маргграф
Таза күйінде қызылша қанты 1747 жылы неміс химигі А. Маргграф

Көмірсулар жіктемесі
Көмірсулар жіктемесі

Көмірсулар биологиялық қызметтері
Энергетикалық
Тіректік қызмет
Қорғаныштық (майлау)қызметі
Гидросмостық және ионды реттеушілік
Кофакторлық
Көмірсулар биологиялық қызметтері
Энергетикалық
Тіректік қызмет
Қорғаныштық (майлау)қызметі
Гидросмостық және ионды реттеушілік
Кофакторлық
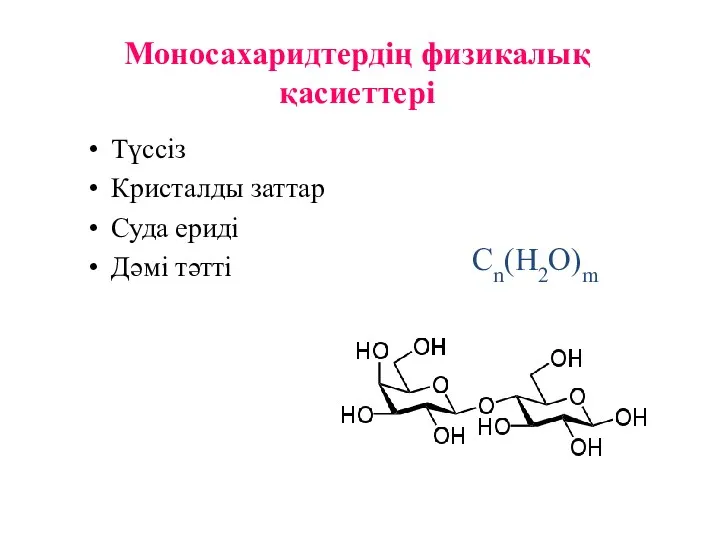
Моносахаридтердің физикалық қасиеттері
Түссіз
Кристалды заттар
Суда ериді
Дәмі тәтті
Cn(H2O)m
Моносахаридтердің физикалық қасиеттері
Түссіз
Кристалды заттар
Суда ериді
Дәмі тәтті
Cn(H2O)m

Моносахаридтер изомериясы. Оптикалық изомерия
Көмірсу молекуласы оптикалы белсенді және құрамынды бірнеше ассиметриялы
Моносахаридтер изомериясы. Оптикалық изомерия
Көмірсу молекуласы оптикалы белсенді және құрамынды бірнеше ассиметриялы

Моносахаридтертің химиялық қасиеттері
1. Альдегидтер мен кетондар сияқты сутегі атомдары әсерінен оңай
Моносахаридтертің химиялық қасиеттері
1. Альдегидтер мен кетондар сияқты сутегі атомдары әсерінен оңай
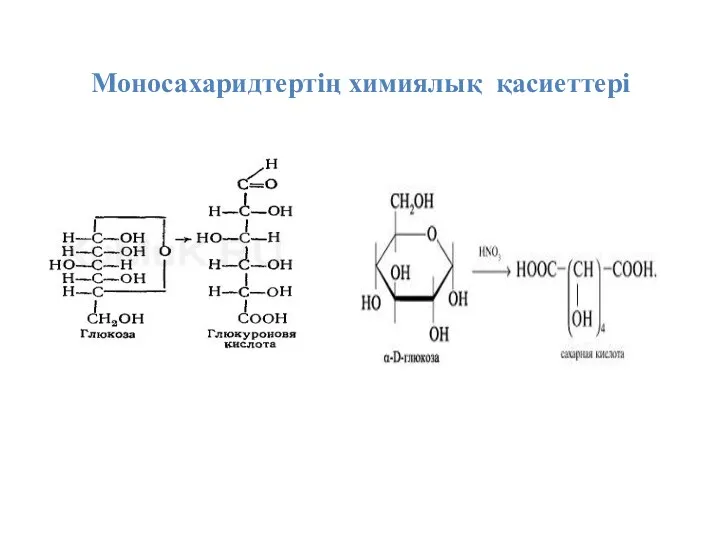
Моносахаридтертің химиялық қасиеттері
Моносахаридтертің химиялық қасиеттері
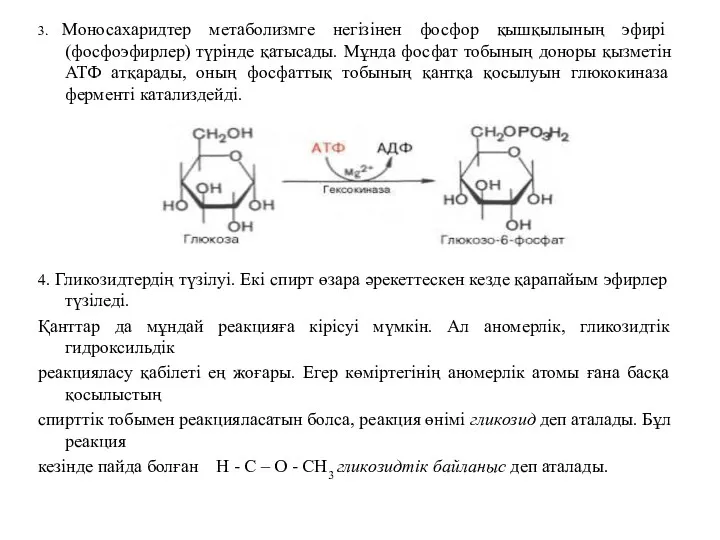
3. Моносахаридтер метаболизмге негізінен фосфор қышқылының эфирі (фосфоэфирлер) түрінде қатысады. Мұнда
3. Моносахаридтер метаболизмге негізінен фосфор қышқылының эфирі (фосфоэфирлер) түрінде қатысады. Мұнда
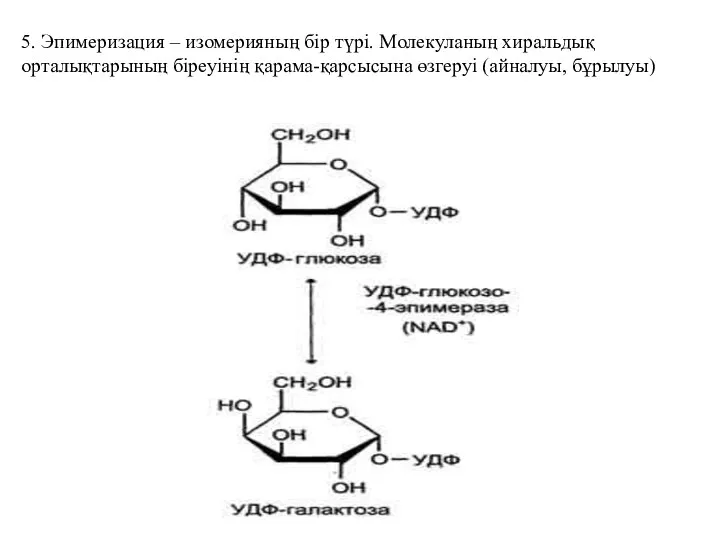
5. Эпимеризация – изомерияның бір түрі. Молекуланың хиральдық орталықтарының біреуінің қарама-қарсысына
5. Эпимеризация – изомерияның бір түрі. Молекуланың хиральдық орталықтарының біреуінің қарама-қарсысына
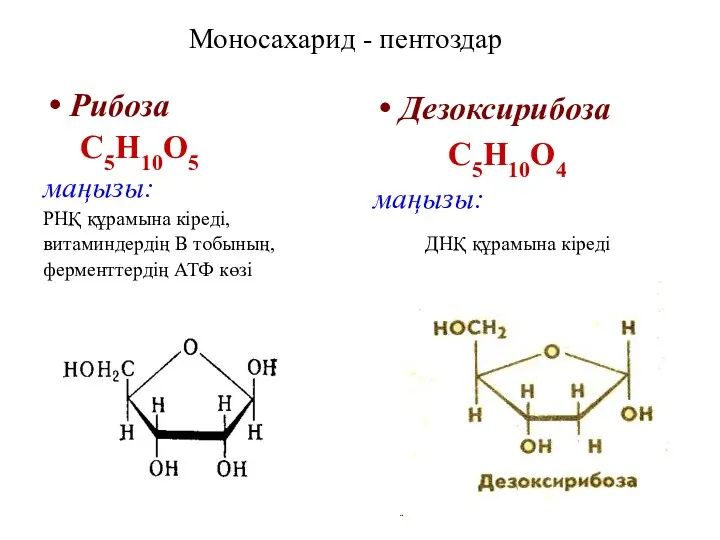
Моносахарид - пентоздар
Рибоза
С5Н10О5
маңызы:
РНҚ құрамына кіреді,
витаминдердің В тобының,
ферменттердің АТФ
Моносахарид - пентоздар
Рибоза
С5Н10О5
маңызы:
РНҚ құрамына кіреді,
витаминдердің В тобының,
ферменттердің АТФ
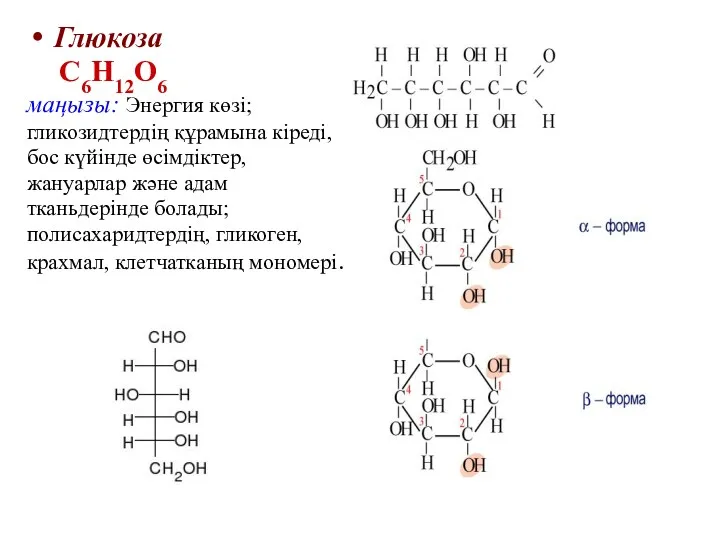
Глюкоза
С6Н12О6
маңызы: Энергия көзі;
гликозидтердің құрамына кіреді,
бос күйінде өсімдіктер,
жануарлар
Глюкоза
С6Н12О6
маңызы: Энергия көзі;
гликозидтердің құрамына кіреді,
бос күйінде өсімдіктер,
жануарлар
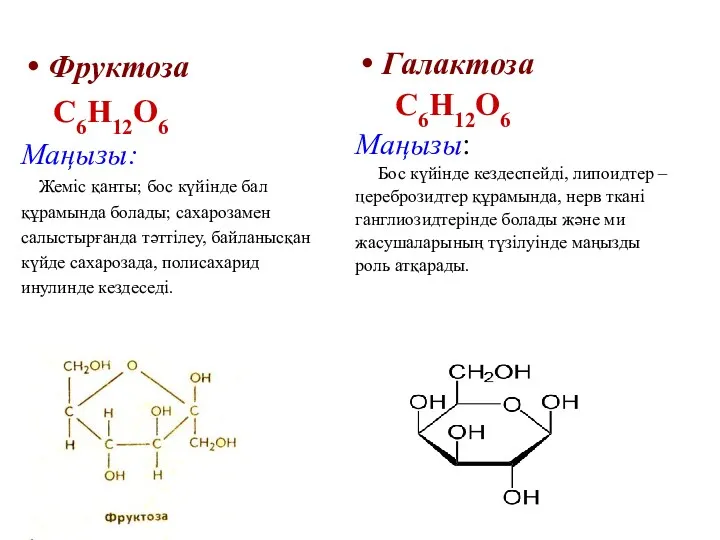
Фруктоза
С6Н12О6
Маңызы:
Жеміс қанты; бос күйінде бал
құрамында болады; сахарозамен
Фруктоза
С6Н12О6
Маңызы:
Жеміс қанты; бос күйінде бал
құрамында болады; сахарозамен

Дисахаридтер
Молекулалық формуласы С12Н22О11, судың бір молекуласын бөліп шығару арқылы гликозидтік байланыспен
Дисахаридтер
Молекулалық формуласы С12Н22О11, судың бір молекуласын бөліп шығару арқылы гликозидтік байланыспен
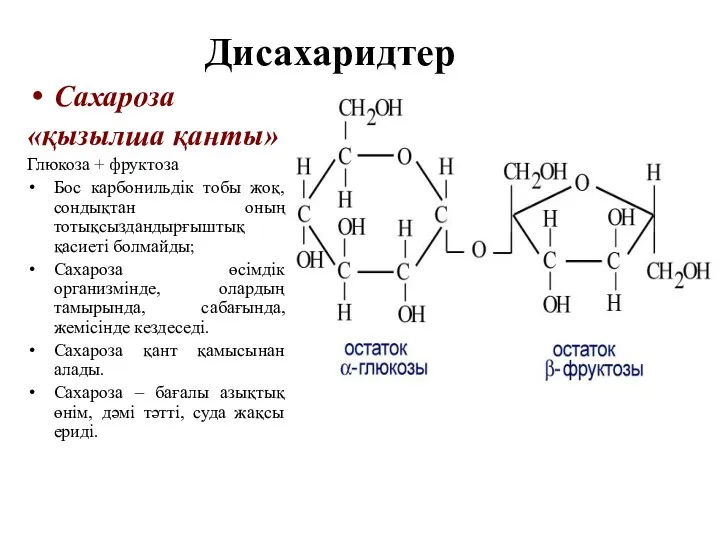
Дисахаридтер
Сахароза
«қызылша қанты»
Глюкоза + фруктоза
Бос карбонильдік тобы жоқ, сондықтан оның тотықсыздандырғыштық
Дисахаридтер
Сахароза
«қызылша қанты»
Глюкоза + фруктоза
Бос карбонильдік тобы жоқ, сондықтан оның тотықсыздандырғыштық
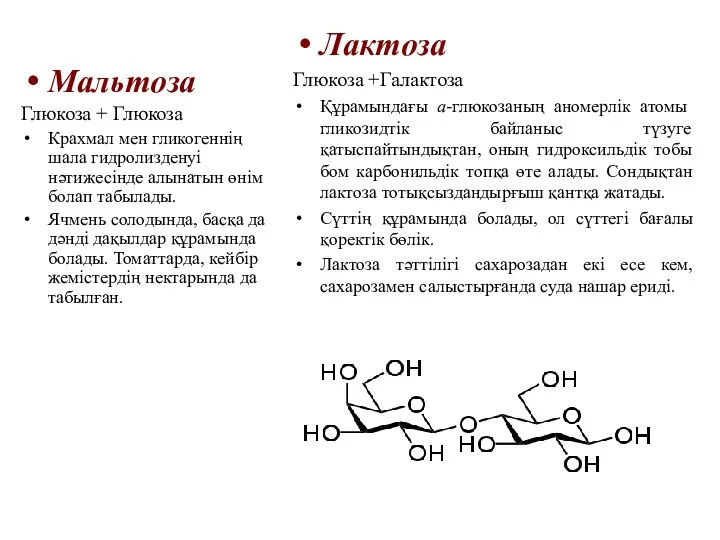
Мальтоза
Глюкоза + Глюкоза
Крахмал мен гликогеннің шала гидролизденуі нәтижесінде алынатын өнім болап
Мальтоза
Глюкоза + Глюкоза
Крахмал мен гликогеннің шала гидролизденуі нәтижесінде алынатын өнім болап

Полисахаридтер
Күрделі көмірсулар, олар гликозидтік байланыстармен жалғасқан 1000 дейін, тіпті одан да
Полисахаридтер
Күрделі көмірсулар, олар гликозидтік байланыстармен жалғасқан 1000 дейін, тіпті одан да

Өсімдіктердің негізгі қоры болатын көмірсу және адамға азық-түлік, малға жемшөп болатын
Өсімдіктердің негізгі қоры болатын көмірсу және адамға азық-түлік, малға жемшөп болатын
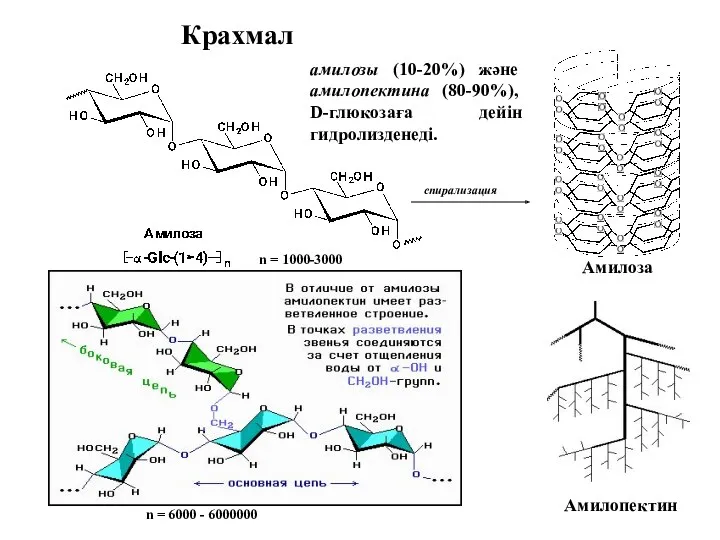
Крахмал
Амилоза
Амилопектин
амилозы (10-20%) және амилопектина (80-90%), D-глюкозаға дейін гидролизденеді.
n = 1000-3000
n =
Крахмал
Амилоза
Амилопектин
амилозы (10-20%) және амилопектина (80-90%), D-глюкозаға дейін гидролизденеді.
n = 1000-3000
n =
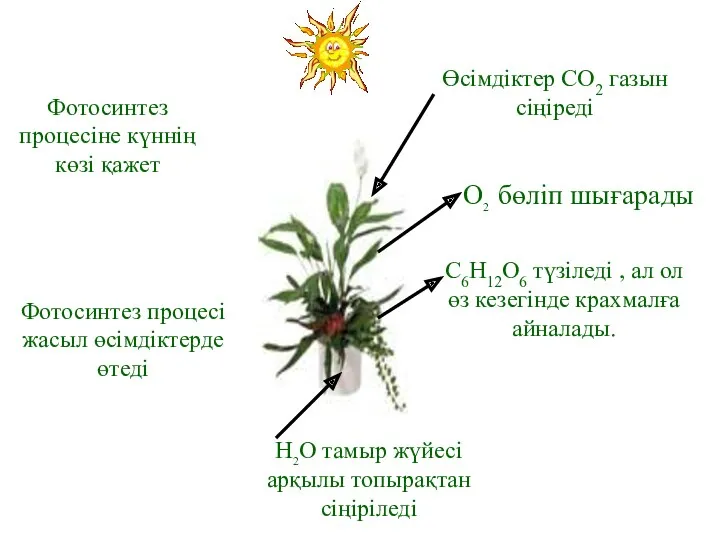
Фотосинтез процесіне күннің көзі қажет
Фотосинтез процесі жасыл өсімдіктерде өтеді
Өсімдіктер СО2 газын
Фотосинтез процесіне күннің көзі қажет
Фотосинтез процесі жасыл өсімдіктерде өтеді
Өсімдіктер СО2 газын
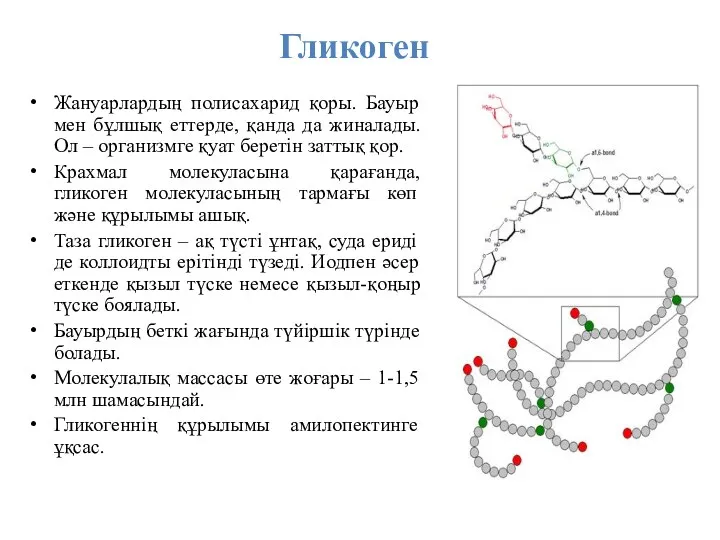
Гликоген
Жануарлардың полисахарид қоры. Бауыр мен бұлшық еттерде, қанда да жиналады.
Гликоген
Жануарлардың полисахарид қоры. Бауыр мен бұлшық еттерде, қанда да жиналады.

Целлюлоза
Өсімдіктер жасушасының негізгі материалы.
Таза целлюлозаның талшықтанған құрылымы бар және
Целлюлоза
Өсімдіктер жасушасының негізгі материалы.
Таза целлюлозаның талшықтанған құрылымы бар және

Хитин целлюлозаға жақын; ол саңырауқұлақтардың кейбір түрлерінде кездеседі; және де
Хитин целлюлозаға жақын; ол саңырауқұлақтардың кейбір түрлерінде кездеседі; және де

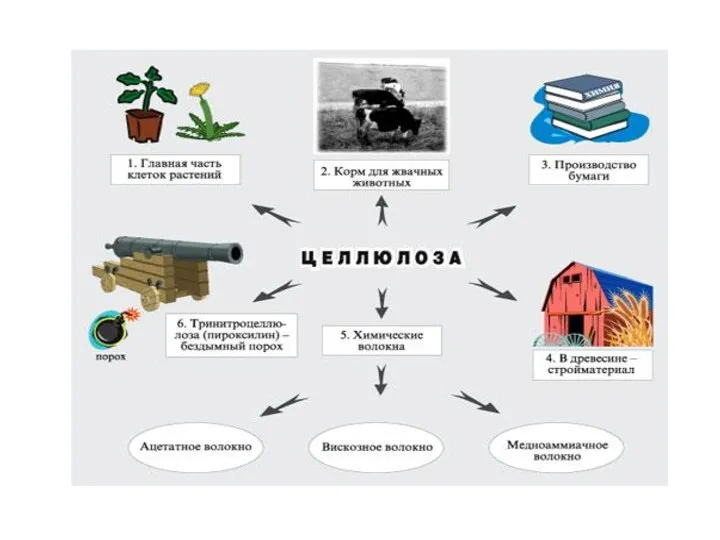
Көмірсудың қолданылуы
Көмірсудың қолданылуы
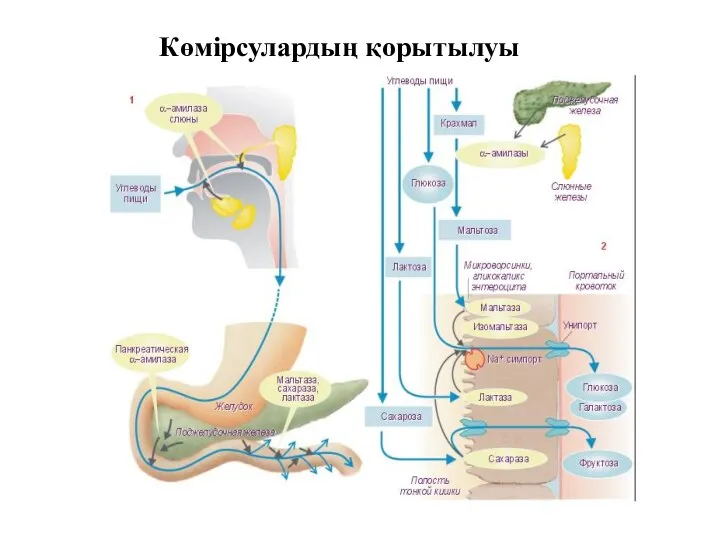
Көмірсулардың қорытылуы
Көмірсулардың қорытылуы

НЕГІЗГІ БӨЛІМ
Нуклеин қышқылы синтезі
Нуклеин қышқылы қасиеттері
НЕГІЗГІ БӨЛІМ
Нуклеин қышқылы синтезі
Нуклеин қышқылы қасиеттері

Швейцария биологы Э.Мишер (1868) ірің жасушалары ядросынан құрамында фосфоры бар
Швейцария биологы Э.Мишер (1868) ірің жасушалары ядросынан құрамында фосфоры бар

РНҚ жасуша ядросында және цитоплазмада (РНҚ-ның 90%-ға жуығы цитоплазмада) орналасқан және
РНҚ жасуша ядросында және цитоплазмада (РНҚ-ның 90%-ға жуығы цитоплазмада) орналасқан және

Нуклеин қышқылы - пішіні жіп тәрізді,
жоғарғы молекулалы, ерітінділерінің
тұтқырлығы жоғары
Нуклеин қышқылы - пішіні жіп тәрізді,
жоғарғы молекулалы, ерітінділерінің
тұтқырлығы жоғары

ДНҚ геннің материалдық негізі, ол геннің келесі қасиеттеріне ие болады:
1. Өз-өзін
ДНҚ геннің материалдық негізі, ол геннің келесі қасиеттеріне ие болады:
1. Өз-өзін

Дезоксирибонуклеотидтер - ағзадағы ДНҚ-ң түзілуіне қатысады.
Рибонуклеотидтер РНҚ- түзілуіне жүмсалады. Рибонуклеотидтердің кейбірі
Дезоксирибонуклеотидтер - ағзадағы ДНҚ-ң түзілуіне қатысады.
Рибонуклеотидтер РНҚ- түзілуіне жүмсалады. Рибонуклеотидтердің кейбірі

Нуклеин қышқылдары - нуклеотид полимерлерінен құралған.
Нуклеотидтердің химиялық құрамы - пиримидиндік
Нуклеин қышқылдары - нуклеотид полимерлерінен құралған.
Нуклеотидтердің химиялық құрамы - пиримидиндік
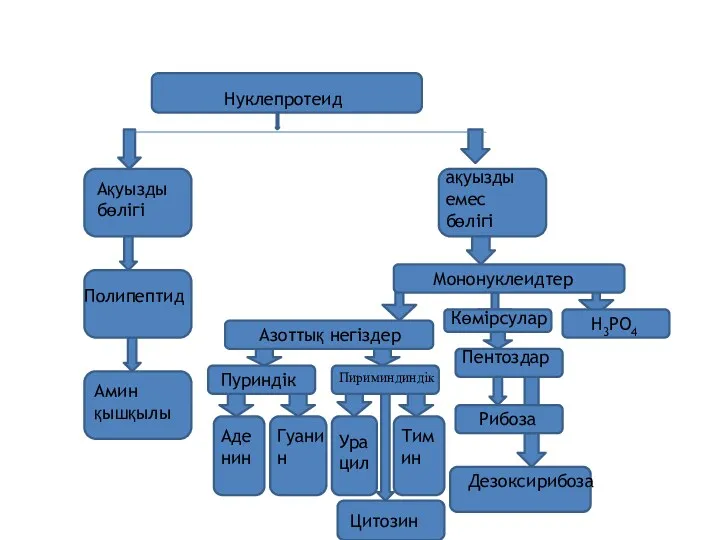
Нуклепротеид
Ақуызды бөлігі
ақуызды емес бөлігі
Полипептид
Амин
қышқылы
Азоттық негіздер
Нуклепротеид
Ақуызды бөлігі
ақуызды емес бөлігі
Полипептид
Амин
қышқылы
Азоттық негіздер
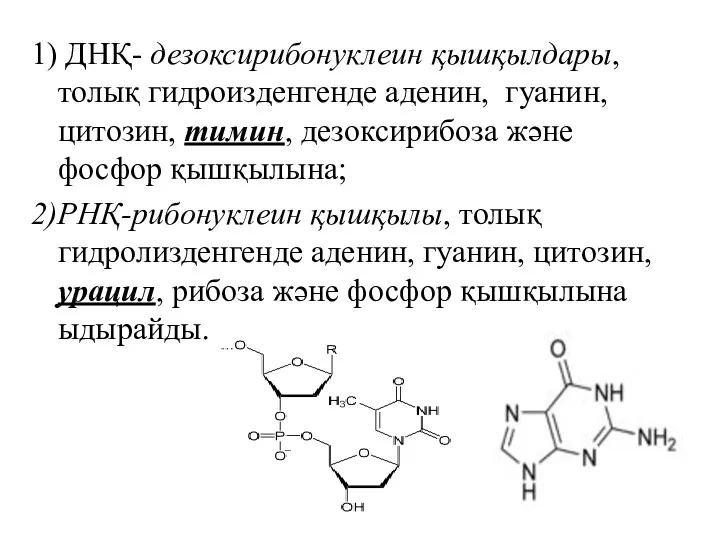
1) ДНҚ- дезоксирибонуклеин қышқылдары, толық гидроизденгенде аденин, гуанин, цитозин, тимин, дезоксирибоза
1) ДНҚ- дезоксирибонуклеин қышқылдары, толық гидроизденгенде аденин, гуанин, цитозин, тимин, дезоксирибоза
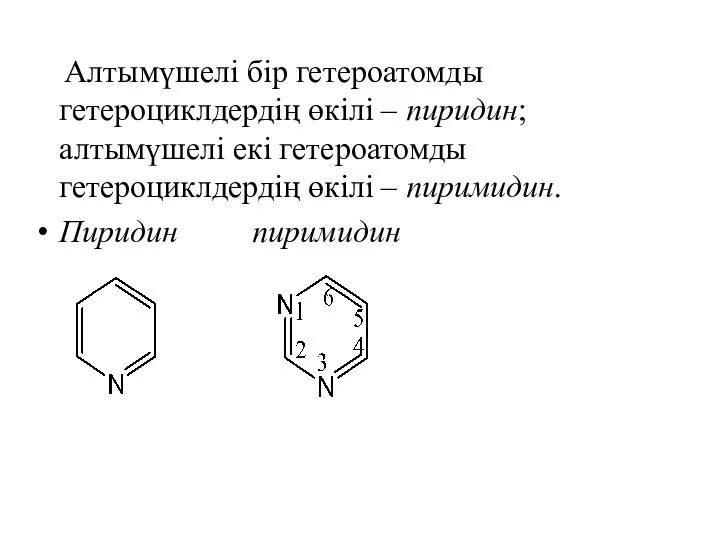
Алтымүшелі бір гетероатомды гетероциклдердің өкілі – пиридин; алтымүшелі екі гетероатомды
Алтымүшелі бір гетероатомды гетероциклдердің өкілі – пиридин; алтымүшелі екі гетероатомды
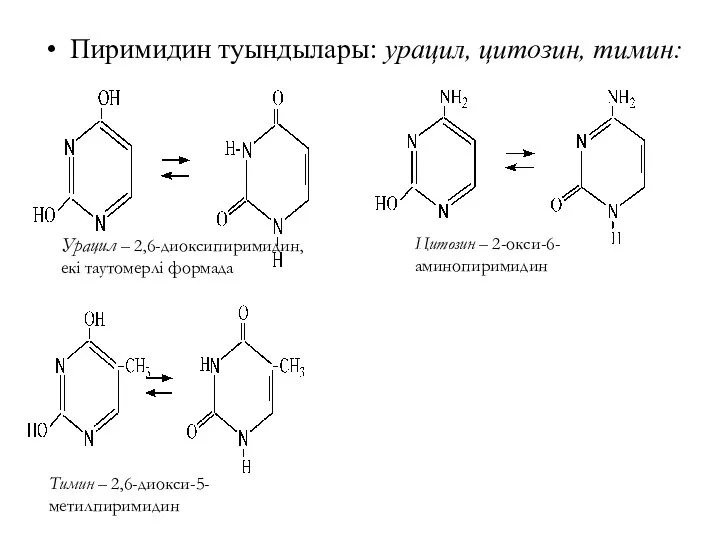
Пиримидин туындылары: урацил, цитозин, тимин:
Урацил – 2,6-диоксипиримидин, екі таутомерлі формада
Пиримидин туындылары: урацил, цитозин, тимин:
Урацил – 2,6-диоксипиримидин, екі таутомерлі формада
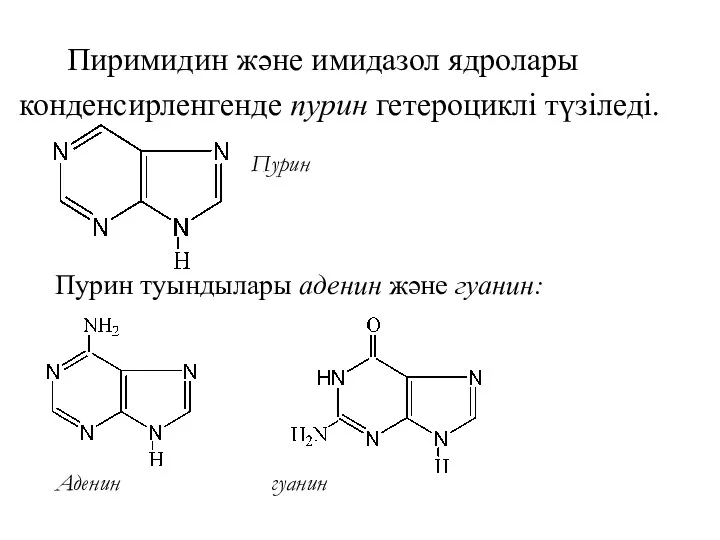
Пиримидин және имидазол ядролары
конденсирленгенде пурин гетероциклі түзіледі.
Пурин
Пиримидин және имидазол ядролары
конденсирленгенде пурин гетероциклі түзіледі.
Пурин
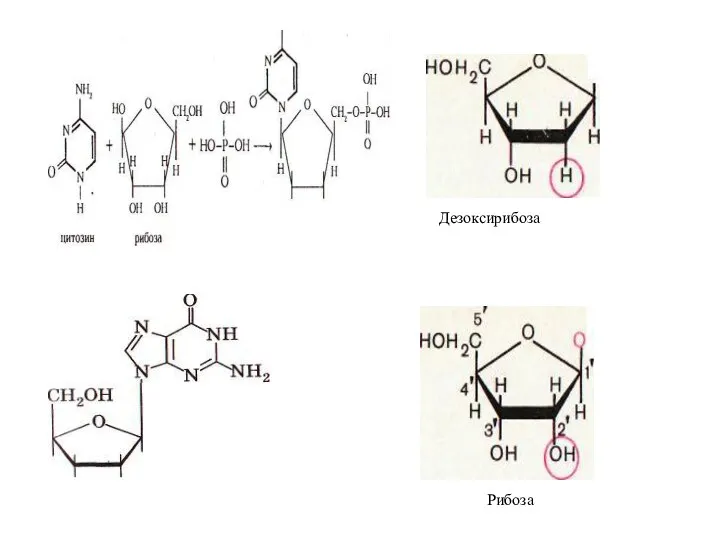
Дезоксирибоза
Рибоза
Дезоксирибоза
Рибоза

Нуклеопротеидтердің гидролизденуі сатылай жүреді және бұл тізбек кез-келген сатыда тоқтауы мүмкін.
Нуклеопротеидтердің гидролизденуі сатылай жүреді және бұл тізбек кез-келген сатыда тоқтауы мүмкін.
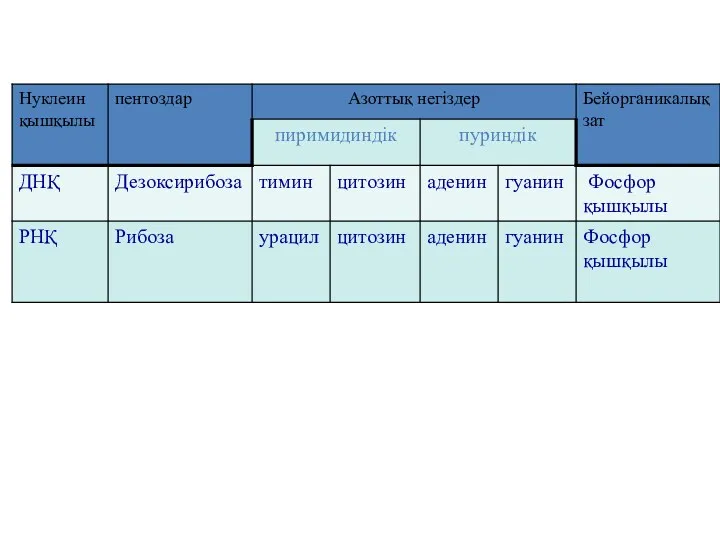

РНҚ (рибонуклеозидтер) құрамына кіретін нуклеозидтер:
аденил қышқылы (аденозин),
гуанин
РНҚ (рибонуклеозидтер) құрамына кіретін нуклеозидтер:
аденил қышқылы (аденозин),
гуанин

Нуклеотидтердің атауы:
нуклеин негіздердің тривиалды атауына сәйкес жалғаулар “идин” пиримидиндік және
Нуклеотидтердің атауы:
нуклеин негіздердің тривиалды атауына сәйкес жалғаулар “идин” пиримидиндік және

Нуклеин қышқылдарының құрамындағы
мононуклеотидтер бір-бірімен оттектік көпірше
арқылы, яғни пентоздардың гидроксил тобы (-ОН)
(үшінші
Нуклеин қышқылдарының құрамындағы
мононуклеотидтер бір-бірімен оттектік көпірше
арқылы, яғни пентоздардың гидроксил тобы (-ОН)
(үшінші

Нуклеин қышқылдарының бірінші реттік құрылымы: мононуклеотидтер бір-бірімен
«3́-5́́ байланыс» арқылы, мұнда
Нуклеин қышқылдарының бірінші реттік құрылымы: мононуклеотидтер бір-бірімен
«3́-5́́ байланыс» арқылы, мұнда
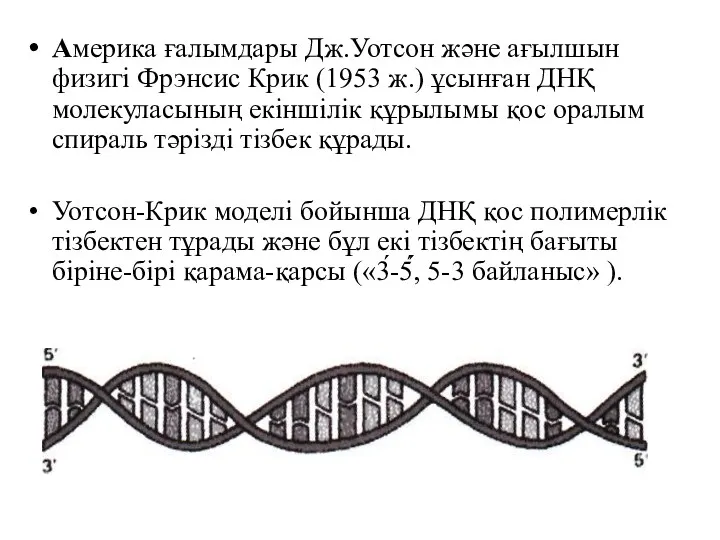
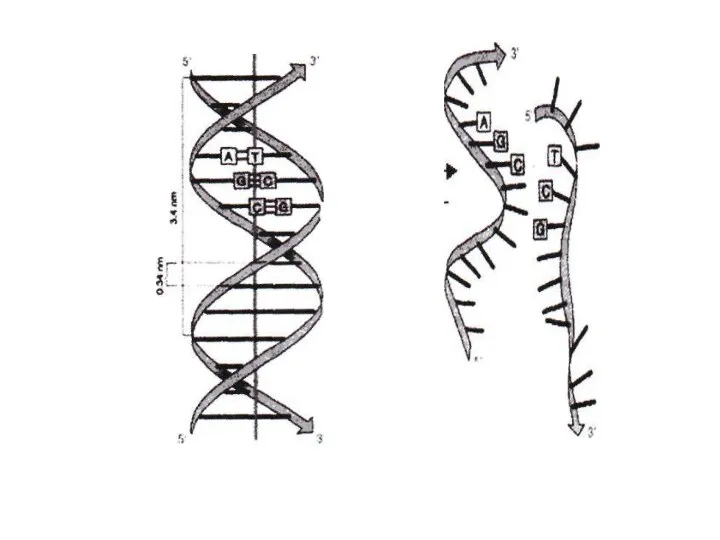
Америка ғалымдары Дж.Уотсон және ағылшын физигі Фрэнсис Крик (1953 ж.) ұсынған
Америка ғалымдары Дж.Уотсон және ағылшын физигі Фрэнсис Крик (1953 ж.) ұсынған

ДНҚ-ның екінші реттік құрылымында молекула оралым спираль тәрізді тізбек құрады.
ДНҚның
ДНҚ-ның екінші реттік құрылымында молекула оралым спираль тәрізді тізбек құрады.
ДНҚның
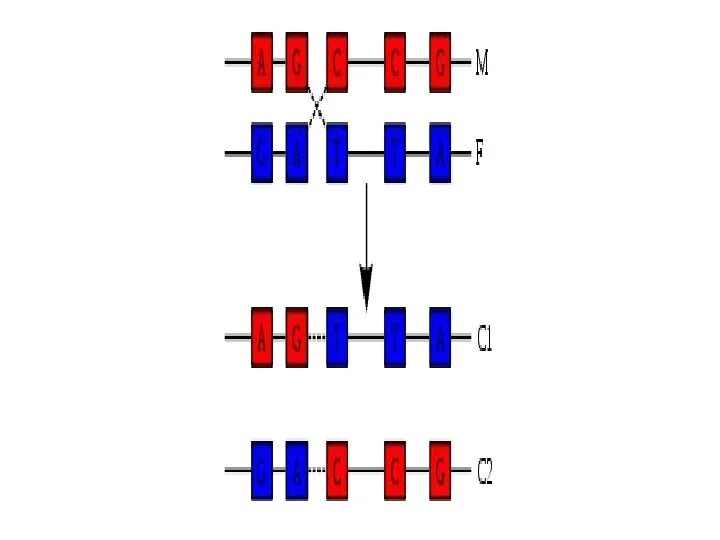

Бір тізбектегі нуклеотидтің орналасуы өз бетінмен еркінірек болғанымен, екінші тізбектегі нуклеотидтердің
Бір тізбектегі нуклеотидтің орналасуы өз бетінмен еркінірек болғанымен, екінші тізбектегі нуклеотидтердің
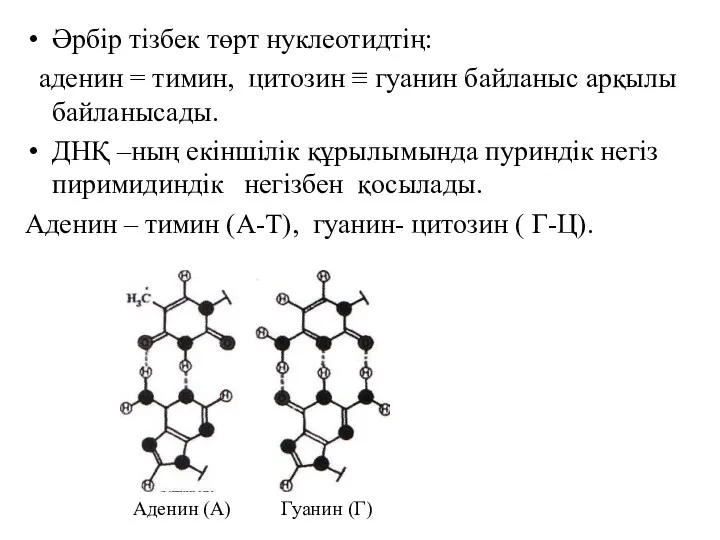
Әрбір тізбек төрт нуклеотидтің:
аденин = тимин, цитозин ≡ гуанин байланыс
Әрбір тізбек төрт нуклеотидтің:
аденин = тимин, цитозин ≡ гуанин байланыс

ДНҚ синтезіне қатысатын негізгі фермент - ДНҚ-на тәуелді ДНҚ полимераза.
ДНҚ молекуласының
ДНҚ синтезіне қатысатын негізгі фермент - ДНҚ-на тәуелді ДНҚ полимераза.
ДНҚ молекуласының

ДНҚ синтезіне қатысатын ферменттер:
1. ДНҚ-на тәуелді ДНҚ-полимеразасы (I, П, III).
ДНҚ синтезіне қатысатын ферменттер:
1. ДНҚ-на тәуелді ДНҚ-полимеразасы (I, П, III).

ДНҚ-ның биосинтезі үш кезеңнен түрады:
1. Инициация. ДНҚ-на тәуелді РНҚ-полимеразасы жұмыс
ДНҚ-ның биосинтезі үш кезеңнен түрады:
1. Инициация. ДНҚ-на тәуелді РНҚ-полимеразасы жұмыс

ҚОРЫТЫНДЫ
Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесінде белок
ҚОРЫТЫНДЫ
Белок биосинтезі тіршіліктің ең маңызды қасиеттерінің бірі, себебі тіршілік процесінде белок
 Болезни внчс у детей и подростков
Болезни внчс у детей и подростков Внутрибольничные инфекции. Меры профилактики
Внутрибольничные инфекции. Меры профилактики Этико-правовые аспекты проведения исследований на животных
Этико-правовые аспекты проведения исследований на животных Невідкладна допомога на догоспітальному етапі. Медсестринський процес
Невідкладна допомога на догоспітальному етапі. Медсестринський процес Невынашивание беременности
Невынашивание беременности Медико-биологическое сопровождение физической культуры и спорта
Медико-биологическое сопровождение физической культуры и спорта Дәрігердің кәсіби деформациясы. Дәрігердің көшбасшылық қасиеті және олардың кәсіби қызметіндегі маңызы
Дәрігердің кәсіби деформациясы. Дәрігердің көшбасшылық қасиеті және олардың кәсіби қызметіндегі маңызы Патофизиология печени
Патофизиология печени Профилактика внутрибольничных инфекций
Профилактика внутрибольничных инфекций Введение в теорию сестринского дела. Понятие о теориях и моделях сестринского дела
Введение в теорию сестринского дела. Понятие о теориях и моделях сестринского дела Лейкоз ауруы кезіндегі зертханалық әдістер
Лейкоз ауруы кезіндегі зертханалық әдістер Преаналитический этап в лабораторной диагностике. Требования по подготовке пациента к лабораторным исследованиям
Преаналитический этап в лабораторной диагностике. Требования по подготовке пациента к лабораторным исследованиям Вопросы смерти и сердечно-легочной реанимации
Вопросы смерти и сердечно-легочной реанимации Желчекаменная болезнь
Желчекаменная болезнь презентация восполит.заб
презентация восполит.заб Балалар мен жасөспірімдер мекемелерінде оқу-тәрбиелеу үрдісінің гигиеналық қағидалары. Күн тәртібінің физиологиялық негізі
Балалар мен жасөспірімдер мекемелерінде оқу-тәрбиелеу үрдісінің гигиеналық қағидалары. Күн тәртібінің физиологиялық негізі Здоровьесберегающие образовательные технологии в педиатрии. Роль центров здоровья в укреплении здоровья детей и подростков
Здоровьесберегающие образовательные технологии в педиатрии. Роль центров здоровья в укреплении здоровья детей и подростков Общая фармакология
Общая фармакология Маршрутизация пациентов и особенности эвакуационных мероприятий больных или лиц с подозрением на Covid-19
Маршрутизация пациентов и особенности эвакуационных мероприятий больных или лиц с подозрением на Covid-19 Амилоидоз
Амилоидоз Антигендер және антиденелер
Антигендер және антиденелер Особенности диагностики и лечения пациента с легочной гипертензией
Особенности диагностики и лечения пациента с легочной гипертензией Классификация заболеваний, проявляющихся гиперандрогенией. Алгоритм дифференциальной диагностики. Принципы терапии
Классификация заболеваний, проявляющихся гиперандрогенией. Алгоритм дифференциальной диагностики. Принципы терапии Уход за лихорадящими пациентами
Уход за лихорадящими пациентами Современные подходы в лечении диабетической полинейропатии. Тиогамма. Мильгамма
Современные подходы в лечении диабетической полинейропатии. Тиогамма. Мильгамма Өкпе артериясының тромбоэмболиясы
Өкпе артериясының тромбоэмболиясы Нейропротезирование. История нейропротезирования
Нейропротезирование. История нейропротезирования Послеродовая депрессия (начало)
Послеродовая депрессия (начало)