- Дифференцировка Т-лимфоцитов

Содержание
- 2. Т-лимфоциты
- 3. T-лимфоциты, или Т-клетки — лимфоциты, развивающиеся у млекопитающих в тимусе из предшественников — претимоцитов, поступающих в
- 4. Основные субпопуляции Т-лимфоцитов Относительное содержание Т-лимфоцитов в крови составляет в среднем около 73% (55–85%) от общего
- 5. Миграция в тимус клеток-предшественников В ходе этих перемещений происходит созревание Т-клеток. На этом отрезке миграции проходят
- 6. Стадии дифференцировки
- 7. DN1-стадия Клетки пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению их числа. Основной ростовой фактор
- 8. DN2-стадия Тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого эпителиальными клетками наружной коры;
- 9. αβT или γδT?
- 10. DN3-стадия Запускается основное событие дифференцировки Т-лимфоцитов — перестройка V-генов TCR (β, γ, δ). Реаранжировка V-генов TCR
- 11. ФЕНОТИП: CD44-CD25+ , 2 суток до β-селекции, β-селекция – 1 сутки На заключительном этапе фазы DN3
- 12. DN3-стадия Внутриклеточный механизм Notch-опосредованной регуляции
- 13. Определенную роль в индукции перестройки V-генов играют цитокины, в частности IL-7, который особенно важен для запуска
- 14. DP-стадия Продолжительность: 1,5-2 сут Перестройка V-гена α-цепи – завершающий этап формирования αβTCR. Все это время не
- 15. Селекция тимоцитов Положительная селекция - обеспечивает отбор только тех тимоцитов, которые экспрессируют ТСR, обладающие сродством к
- 16. Отрицательная селекция - ограничивает степень агрессивности Т-клеток в отношении собственных молекул. На этом этапе тимоциты представлены
- 17. Т-клетки как бы ≪натаскиваются≫ на распознавание собственных антигенов, а не чужеродных молекул, но степень агрессивности Т-клеток
- 18. Дифференцировка CD4+ и CD8+ лимфоцитов Дифференцировка Т-клеток регулируется как внутренними (дифференцировочные факторы), так и внешними (сигналы,
- 19. Дифференцировка CD4+ и CD8+ лимфоцитов Факторы, контролирующие дифференцировку αβТ-клеток
- 20. Созревшие тимоциты довольно долго (7—14 сут) не покидают тимус, пребывая преимущественно в наружных слоях мозгового слоя;
- 22. Характеристика «классических» αβТ-клеток На поверхности αβТ-клеток экспрессируется примерно в 5 раз меньше молекул TCR, чем BCR
- 23. Т-лимфоциты относят к долгоживущим клеткам. Срок жизни различных субпопуляций наивных αβТ-лимфоцитов составляет месяцы и годы; Основное
- 25. Скачать презентацию
Т-лимфоциты
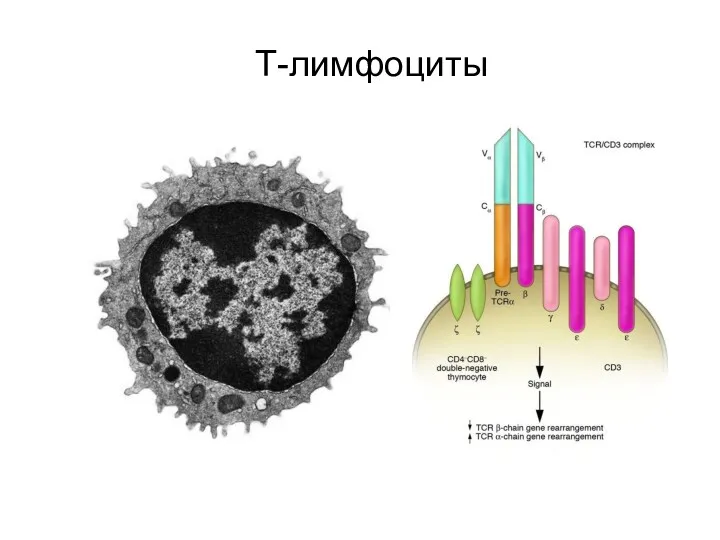
Т-лимфоциты

T-лимфоциты, или Т-клетки — лимфоциты, развивающиеся у млекопитающих в тимусе из предшественников — претимоцитов, поступающих в него из красного костного мозга.
T-лимфоциты, или Т-клетки — лимфоциты, развивающиеся у млекопитающих в тимусе из предшественников — претимоцитов, поступающих в него из красного костного мозга.
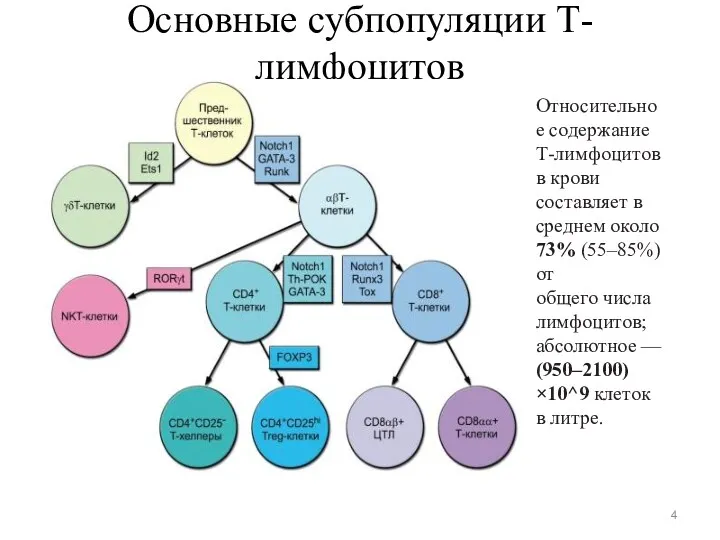
Основные субпопуляции Т-лимфоцитов
Относительное содержание Т-лимфоцитов в крови составляет в среднем около
Основные субпопуляции Т-лимфоцитов
Относительное содержание Т-лимфоцитов в крови составляет в среднем около
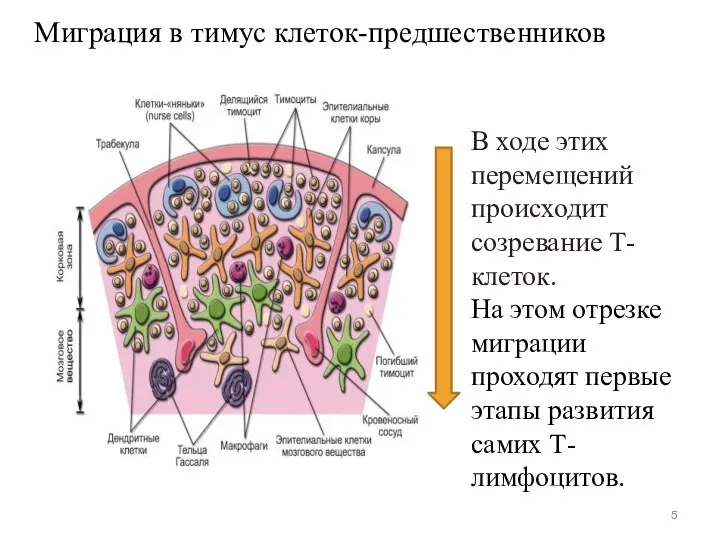
Миграция в тимус клеток-предшественников
В ходе этих перемещений происходит созревание Т-клеток.
На этом
Миграция в тимус клеток-предшественников
В ходе этих перемещений происходит созревание Т-клеток.
На этом
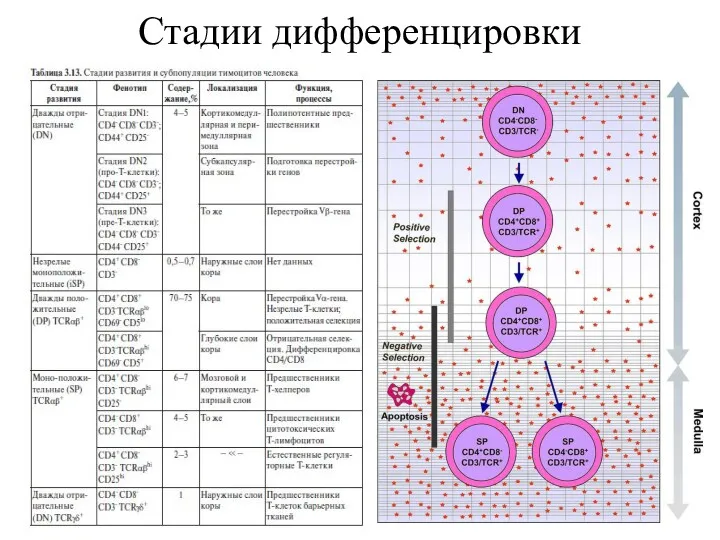
Стадии дифференцировки
Стадии дифференцировки

DN1-стадия
Клетки пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению их
DN1-стадия
Клетки пролиферируют (до 10 делений), что приводит к 1000-кратному увеличению их

DN2-стадия
Тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого
DN2-стадия
Тимоциты перемещаются в сторону наружных слоев коры по градиенту CXCL12, секретируемого
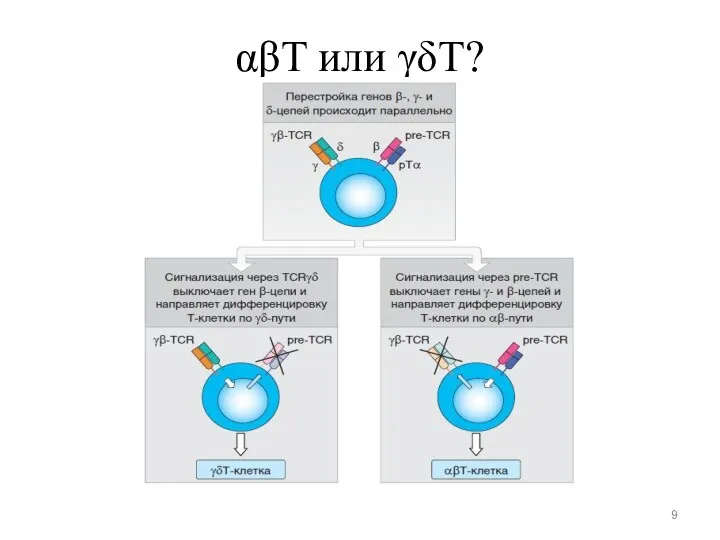
αβT или γδT?
αβT или γδT?

DN3-стадия
Запускается основное событие дифференцировки Т-лимфоцитов — перестройка V-генов TCR (β, γ,
DN3-стадия
Запускается основное событие дифференцировки Т-лимфоцитов — перестройка V-генов TCR (β, γ,

ФЕНОТИП: CD44-CD25+ ,
2 суток до β-селекции, β-селекция – 1 сутки
На заключительном
ФЕНОТИП: CD44-CD25+ ,
2 суток до β-селекции, β-селекция – 1 сутки
На заключительном
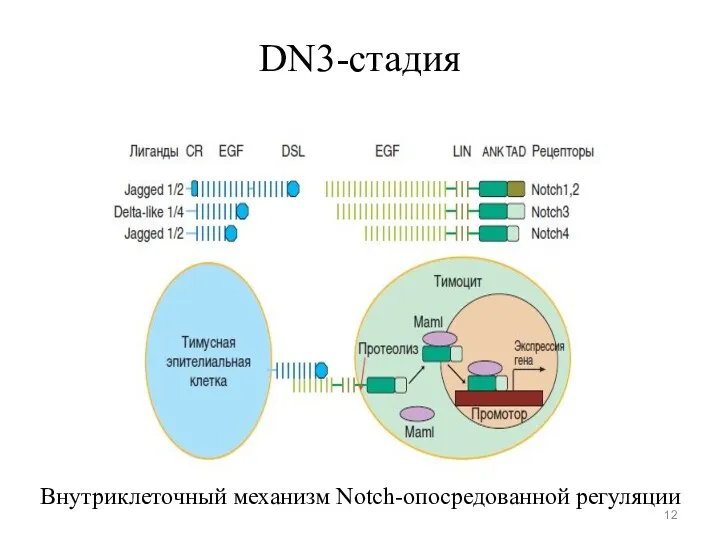
DN3-стадия
Внутриклеточный механизм Notch-опосредованной регуляции
DN3-стадия
Внутриклеточный механизм Notch-опосредованной регуляции

Определенную роль в индукции перестройки V-генов играют цитокины, в частности IL-7,
Определенную роль в индукции перестройки V-генов играют цитокины, в частности IL-7,

DP-стадия
Продолжительность: 1,5-2 сут
Перестройка V-гена α-цепи – завершающий этап формирования αβTCR.
Все это
DP-стадия
Продолжительность: 1,5-2 сут
Перестройка V-гена α-цепи – завершающий этап формирования αβTCR.
Все это

Селекция тимоцитов
Положительная селекция - обеспечивает отбор только тех тимоцитов, которые экспрессируют
Селекция тимоцитов
Положительная селекция - обеспечивает отбор только тех тимоцитов, которые экспрессируют

Отрицательная селекция - ограничивает степень агрессивности Т-клеток в отношении собственных молекул.
На
Отрицательная селекция - ограничивает степень агрессивности Т-клеток в отношении собственных молекул.
На

Т-клетки как бы ≪натаскиваются≫ на распознавание собственных антигенов, а не чужеродных
Т-клетки как бы ≪натаскиваются≫ на распознавание собственных антигенов, а не чужеродных

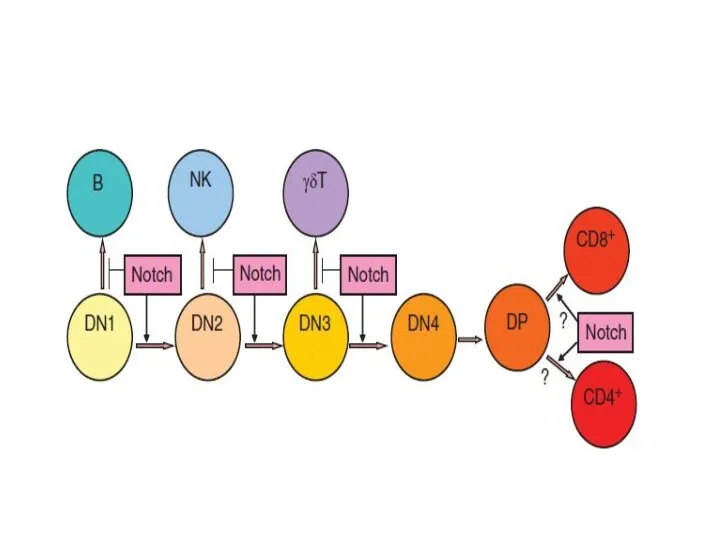
Дифференцировка CD4+ и CD8+ лимфоцитов
Дифференцировка Т-клеток регулируется как внутренними (дифференцировочные факторы),
Дифференцировка CD4+ и CD8+ лимфоцитов
Дифференцировка Т-клеток регулируется как внутренними (дифференцировочные факторы),
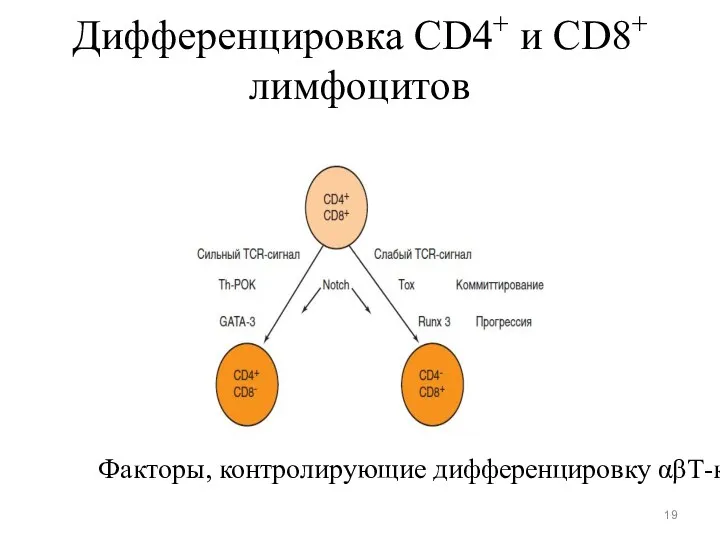
Дифференцировка CD4+ и CD8+ лимфоцитов
Факторы, контролирующие дифференцировку αβТ-клеток
Дифференцировка CD4+ и CD8+ лимфоцитов
Факторы, контролирующие дифференцировку αβТ-клеток

Созревшие тимоциты довольно долго (7—14 сут) не покидают тимус, пребывая преимущественно
Созревшие тимоциты довольно долго (7—14 сут) не покидают тимус, пребывая преимущественно

Характеристика «классических» αβТ-клеток
На поверхности αβТ-клеток экспрессируется примерно в 5 раз меньше
Характеристика «классических» αβТ-клеток
На поверхности αβТ-клеток экспрессируется примерно в 5 раз меньше

Т-лимфоциты относят к долгоживущим клеткам. Срок жизни различных субпопуляций наивных αβТ-лимфоцитов
Т-лимфоциты относят к долгоживущим клеткам. Срок жизни различных субпопуляций наивных αβТ-лимфоцитов
 Оценка функционального состояния
Оценка функционального состояния Asphyxia
Asphyxia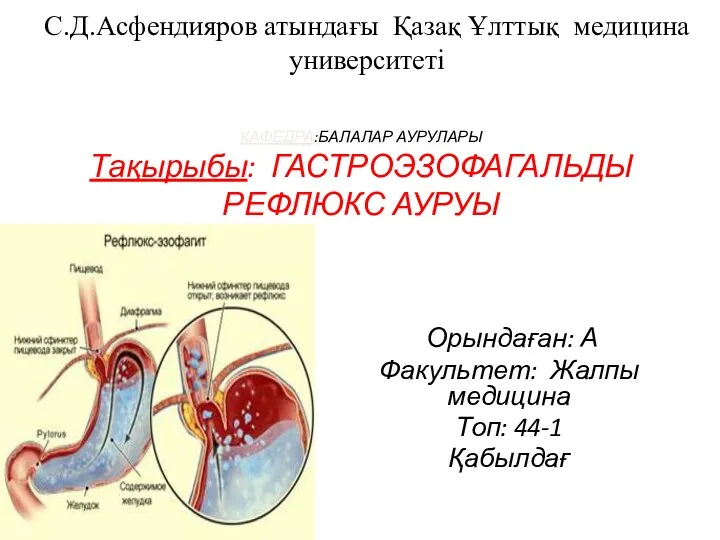 Гастроэзофагальды рефлюкс ауруы
Гастроэзофагальды рефлюкс ауруы Общественное здравоохранение
Общественное здравоохранение Жұқпалы аурумен ауырған малдың ұшасы мен ағзаларын малдәрiгерлiк-санитариялық сараптау
Жұқпалы аурумен ауырған малдың ұшасы мен ағзаларын малдәрiгерлiк-санитариялық сараптау Туберкулез, алкоголизм и наркомания
Туберкулез, алкоголизм и наркомания Коклюш у детей раннего возраста
Коклюш у детей раннего возраста Неспецифический язвенный колит и болезнь Крона
Неспецифический язвенный колит и болезнь Крона Внутрибольничные инфекции. Масштаб, проблемы и структура. Инфекционный процесс
Внутрибольничные инфекции. Масштаб, проблемы и структура. Инфекционный процесс Клинические рекомендации по диагностике, лечению и профилактике тяжелой внебольничной пневмонии у взрослых
Клинические рекомендации по диагностике, лечению и профилактике тяжелой внебольничной пневмонии у взрослых Внебольничная пневмония у взрослых: диагностика, лечение, профилактика
Внебольничная пневмония у взрослых: диагностика, лечение, профилактика Нарушения менструального цикла
Нарушения менструального цикла Комалар кезіндегі диагностикалау және жедел көмек көрсету алгоритмі
Комалар кезіндегі диагностикалау және жедел көмек көрсету алгоритмі Острая дыхательная недостаточность
Острая дыхательная недостаточность Травма шейного отдела позвоночника
Травма шейного отдела позвоночника Дифференциальный диагноз при аритмиях. Клиническая и ЭКГ-диагностика. Принципы лечения и неотложная помощь
Дифференциальный диагноз при аритмиях. Клиническая и ЭКГ-диагностика. Принципы лечения и неотложная помощь Слайд-лекция №12. Нейролептики. Транквилизаторы. Седативные. Соли лития
Слайд-лекция №12. Нейролептики. Транквилизаторы. Седативные. Соли лития Влияние наушников на слух школьников
Влияние наушников на слух школьников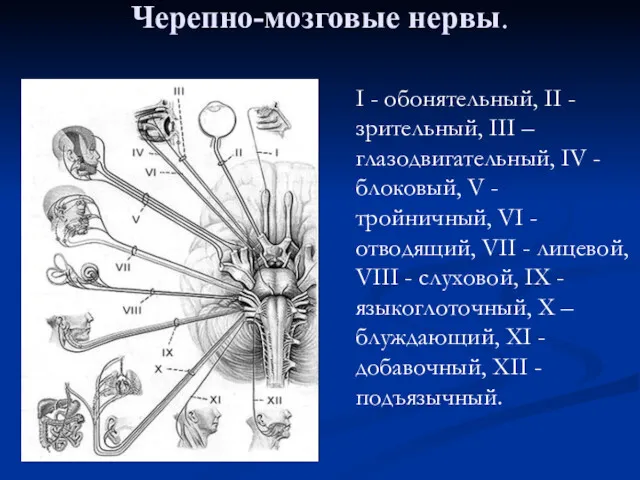 Черепно-мозговые нервы и их поражение
Черепно-мозговые нервы и их поражение Державна санітарноепідеміологічна експертиза товарів для дітей та підлітків
Державна санітарноепідеміологічна експертиза товарів для дітей та підлітків Расстройства восприятия
Расстройства восприятия Авиценна и народная медицина
Авиценна и народная медицина Гастрит. Жедел гастрит
Гастрит. Жедел гастрит Патогенез туберкулеза
Патогенез туберкулеза Кости верхней конечности и их соединения
Кости верхней конечности и их соединения Государственная регистрация лекарственных средств
Государственная регистрация лекарственных средств Плевриты. Инфекционные и неинфекционные плевриты
Плевриты. Инфекционные и неинфекционные плевриты Covid-19. Симптомы и признаки коронавируса у человека
Covid-19. Симптомы и признаки коронавируса у человека