- Физиология нервной системы. Часть 2

Содержание
- 2. Общий план центральной регуляции движений Движения могут быть направлены на поддержание позы, перемещение тела в пространстве,
- 3. Ствол мозга обеспечивает регуляцию позы и тонический компонент движений. Спинной мозг осуществляет простейшие рефлексы поддержания длины
- 5. Спинной мозг в регуляции движений Самостоятельная деятельность спинальных двигательных систем обеспечивает простейшие, но очень важные двигательные
- 6. Регуляция двигательной активности в спинном мозге обеспечивается вставочными нейронами, альфа-мотонейронами и гамма-мотонейронами. Эфферентная иннервация скелетных мышц
- 7. Чувствительные волокна афферентных нейронов обвиваются вокруг средней части интрафузального волокна, образуя аннулоспиральное окончание. Это - первичные
- 8. Мышечные веретена - рецепторы растяжения. При растяжении мышцы, соответствующей длине покоя, частота ПД, идущих по афферентным
- 9. Рефлексы поддержания длины скелетных мышц. Суть- рефлекторное укорочение или расслабление скелетных мышц при увеличении или уменьшении
- 10. Рефлексы ограничения напряжения скелетных мышц (тормозные сухожильные рефлексы). Осуществляются при увеличении напряжения мышцы. Возрастающее механическое напряжение
- 11. Полисинаптические двигательные рефлексы. Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов давления и боли скелетных мышц
- 12. Двигательные системы ствола мозга Обеспечивают более высокий уровень регуляции движений, относятся к структурам непосредственного действия. Их
- 13. Децеребрационная ригидность демонстрирует роль стволовых центров в регуляции тонуса и позы. Она возникает при перерезке ЦНС
- 14. КОШКА ПОСЛЕ ПЕРЕРЕЗКИ ЦНС НИЖЕ КРАСНОГО ЯДРА
- 15. Тонические рефлексы ствола мозга статические статокинетические Познотонические рефлексы обеспечиваются преимущественно бульварным отделом. Связаны с определенным перераспределением
- 16. Статокинетические рефлексы возникают при линейном или угловом ускорении. Это наиболее сложные рефлексы ствола мозга. Они осуществляются
- 17. Двигательные рефлексы ствола мозга обеспечивают согласованную работу многих групп мышц в процессе поддержания позы, ее изменения.
- 18. Мозжечок в обеспечении двигательной активности Не является ни сенсорным, ни моторным, ни интегративным образованием ЦНС в
- 19. ПОСЛЕДСТВИЯ УДАЛЕНИЯ МОЗЖЕЧКА Фаза раздражения. Длится несколько суток. Причиной является отек тканей, раздражение мозга, кровоизлияния. Проявляется
- 20. ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА А.Регуляция позы, мышечного тонуса, равновесия, поддерживающих движений. За ее выполнение отвечает червь мозжечка,
- 21. ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА В. Коррекция медленных целенаправленных движений в процессе их выполнения и координация их с
- 22. Таким образом, мозжечок не только модулирует уже начатые двигательные акты, но и обеспечивает создание и хранение
- 23. Функции базальных ганглиев в регуляции движений Программное обеспечение стереотипных, медленных («червеобразных») движений. Эти двигательные программы видовые,
- 24. К стриопаллидарной системе (система базальных ганглиев) относятся структуры мозга: полосатое тело (стриатум), бледный шар (паллидум), черная
- 25. Двигательная кора в обеспечении моторного действия Функционально является низшим звеном организации программы действия и высшим уровнем
- 26. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ двигательной коры в обеспечении моторного действия Эфферентные пути образованы аксонами гигантских пирамидных клеток Беца.
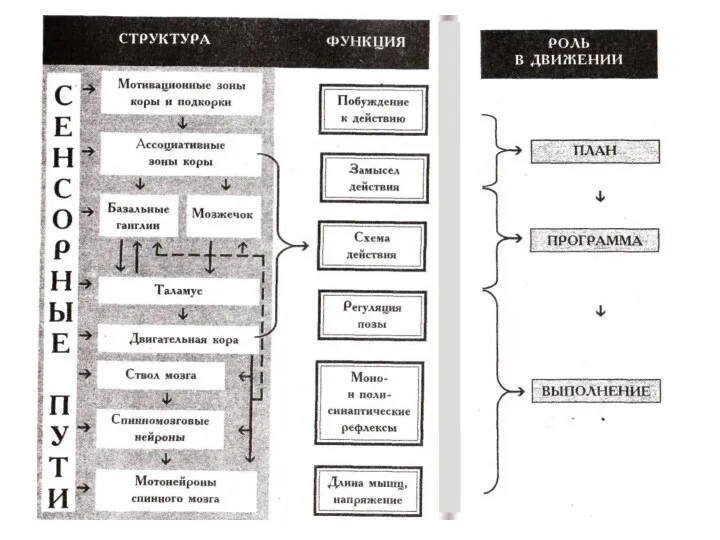
- 27. Побуждение и замысел действия Реализуются на уровне мотивационных зон коры и подкорки, ассоциативных зон коры. Формирование
- 29. Скачать презентацию

Общий план центральной регуляции движений
Движения могут быть направлены на поддержание
Общий план центральной регуляции движений
Движения могут быть направлены на поддержание

Ствол мозга обеспечивает регуляцию позы и тонический компонент движений.
Спинной
Ствол мозга обеспечивает регуляцию позы и тонический компонент движений.
Спинной

Спинной мозг в регуляции движений
Самостоятельная деятельность спинальных двигательных систем обеспечивает
Спинной мозг в регуляции движений
Самостоятельная деятельность спинальных двигательных систем обеспечивает

Регуляция двигательной активности в спинном мозге
обеспечивается вставочными нейронами, альфа-мотонейронами и
Регуляция двигательной активности в спинном мозге
обеспечивается вставочными нейронами, альфа-мотонейронами и
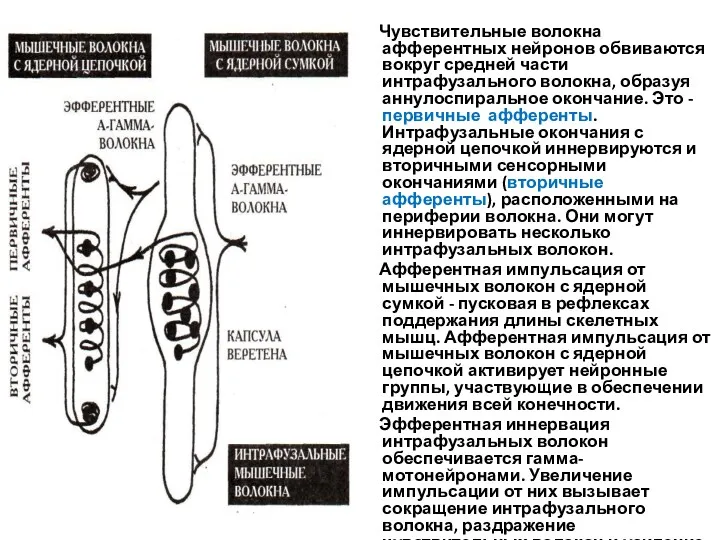
Чувствительные волокна афферентных нейронов обвиваются вокруг средней части интрафузального
Чувствительные волокна афферентных нейронов обвиваются вокруг средней части интрафузального

Мышечные веретена - рецепторы растяжения. При растяжении мышцы, соответствующей длине покоя,
Мышечные веретена - рецепторы растяжения. При растяжении мышцы, соответствующей длине покоя,
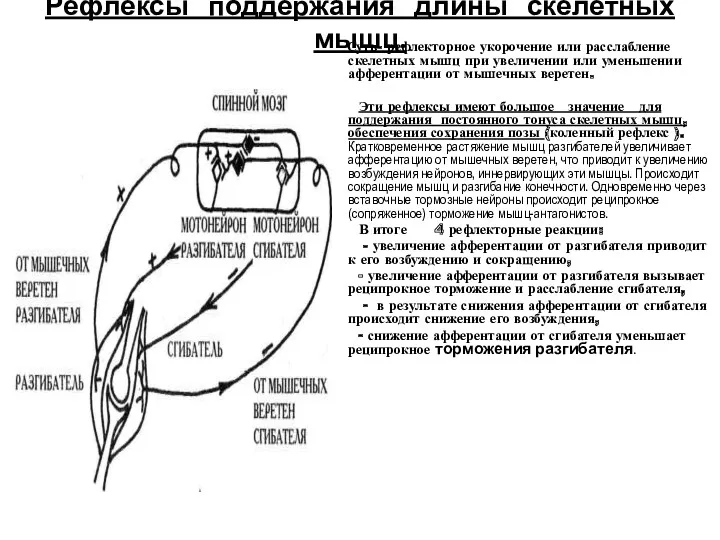
Рефлексы поддержания длины скелетных мышц.
Суть- рефлекторное укорочение или расслабление скелетных
Рефлексы поддержания длины скелетных мышц.
Суть- рефлекторное укорочение или расслабление скелетных
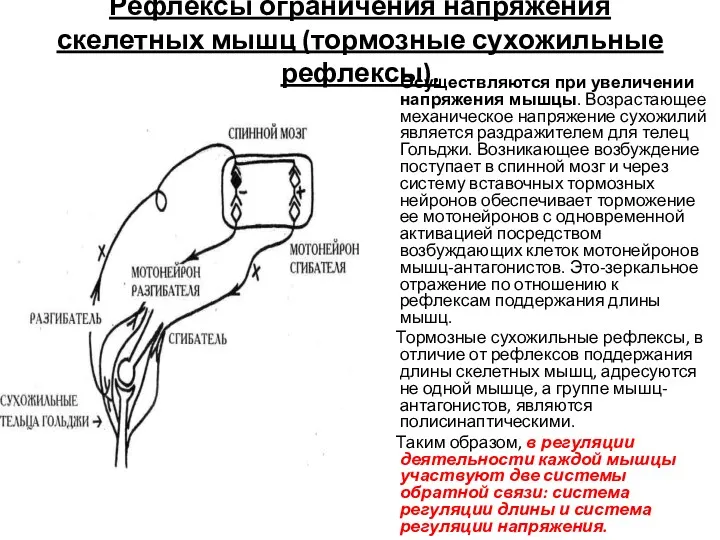
Рефлексы ограничения напряжения скелетных мышц (тормозные сухожильные рефлексы).
Осуществляются при увеличении
Рефлексы ограничения напряжения скелетных мышц (тормозные сухожильные рефлексы).
Осуществляются при увеличении
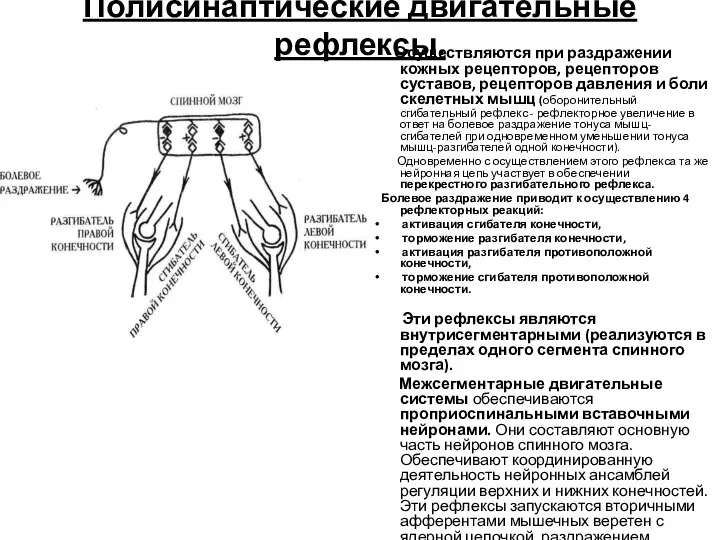
Полисинаптические двигательные рефлексы.
Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов
Полисинаптические двигательные рефлексы.
Осуществляются при раздражении кожных рецепторов, рецепторов суставов, рецепторов
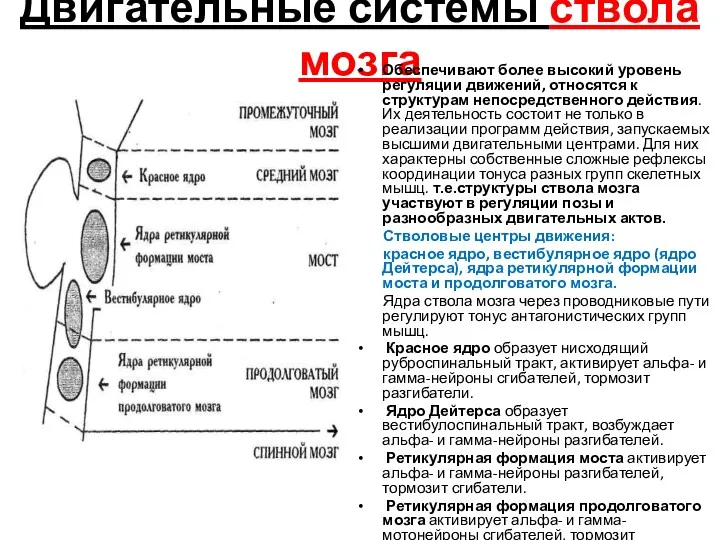
Двигательные системы ствола мозга
Обеспечивают более высокий уровень регуляции движений, относятся к
Двигательные системы ствола мозга
Обеспечивают более высокий уровень регуляции движений, относятся к

Децеребрационная ригидность
демонстрирует роль стволовых центров в регуляции тонуса и позы.
Децеребрационная ригидность
демонстрирует роль стволовых центров в регуляции тонуса и позы.

КОШКА ПОСЛЕ ПЕРЕРЕЗКИ ЦНС НИЖЕ КРАСНОГО ЯДРА
КОШКА ПОСЛЕ ПЕРЕРЕЗКИ ЦНС НИЖЕ КРАСНОГО ЯДРА

Тонические рефлексы ствола мозга
статические статокинетические
Познотонические рефлексы
обеспечиваются преимущественно бульварным
Тонические рефлексы ствола мозга
статические статокинетические
Познотонические рефлексы
обеспечиваются преимущественно бульварным
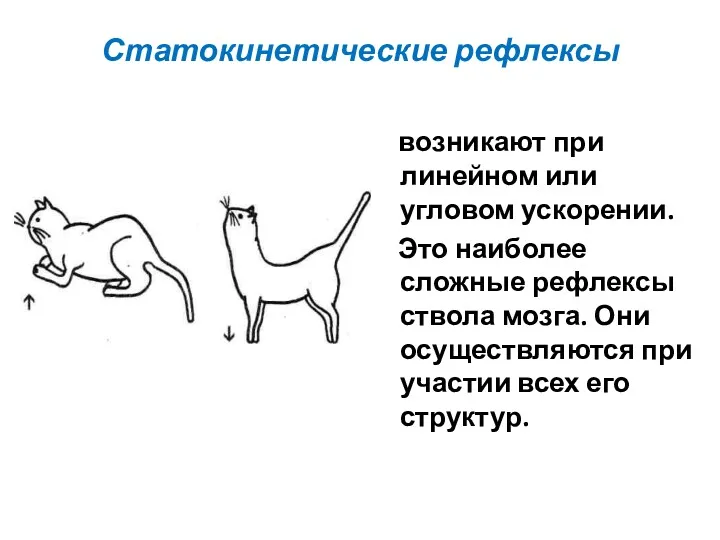
Статокинетические рефлексы
возникают при линейном или угловом ускорении.
Это наиболее сложные
Статокинетические рефлексы
возникают при линейном или угловом ускорении.
Это наиболее сложные

Двигательные рефлексы ствола мозга обеспечивают согласованную работу многих групп
Двигательные рефлексы ствола мозга обеспечивают согласованную работу многих групп

Мозжечок в обеспечении двигательной активности
Не является ни сенсорным, ни моторным, ни
Мозжечок в обеспечении двигательной активности
Не является ни сенсорным, ни моторным, ни

ПОСЛЕДСТВИЯ УДАЛЕНИЯ МОЗЖЕЧКА
Фаза раздражения. Длится несколько суток. Причиной является отек тканей,
ПОСЛЕДСТВИЯ УДАЛЕНИЯ МОЗЖЕЧКА
Фаза раздражения. Длится несколько суток. Причиной является отек тканей,
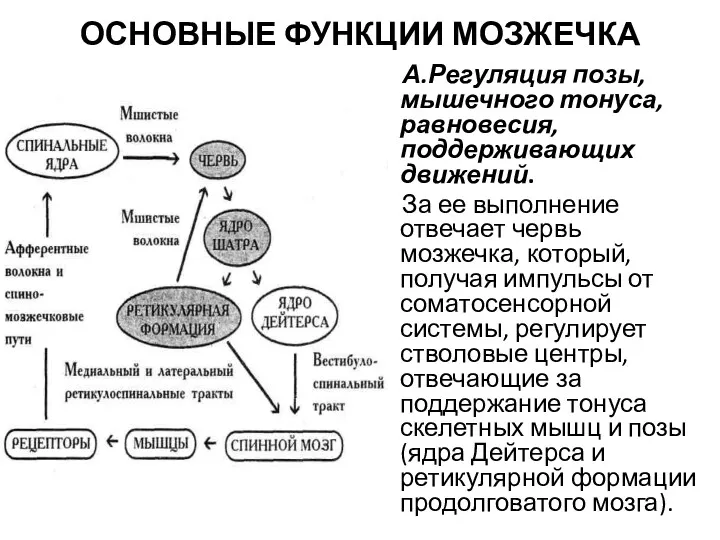
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
А.Регуляция позы, мышечного тонуса, равновесия, поддерживающих движений.
За
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
А.Регуляция позы, мышечного тонуса, равновесия, поддерживающих движений.
За
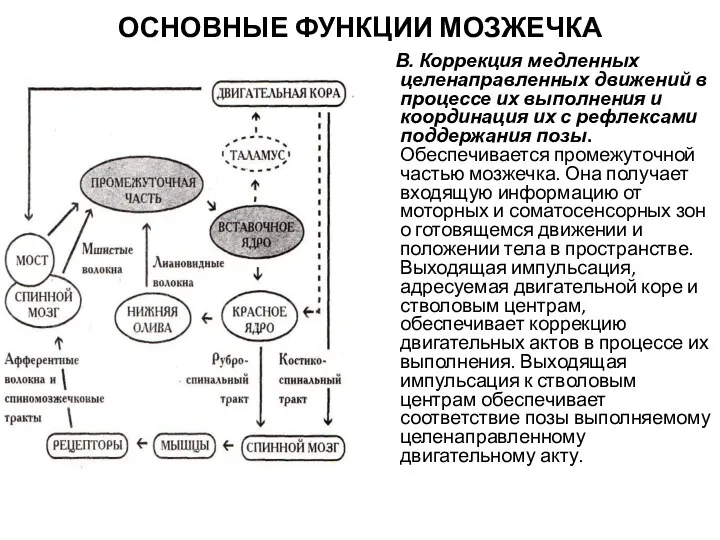
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
В. Коррекция медленных целенаправленных движений в процессе их
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
В. Коррекция медленных целенаправленных движений в процессе их
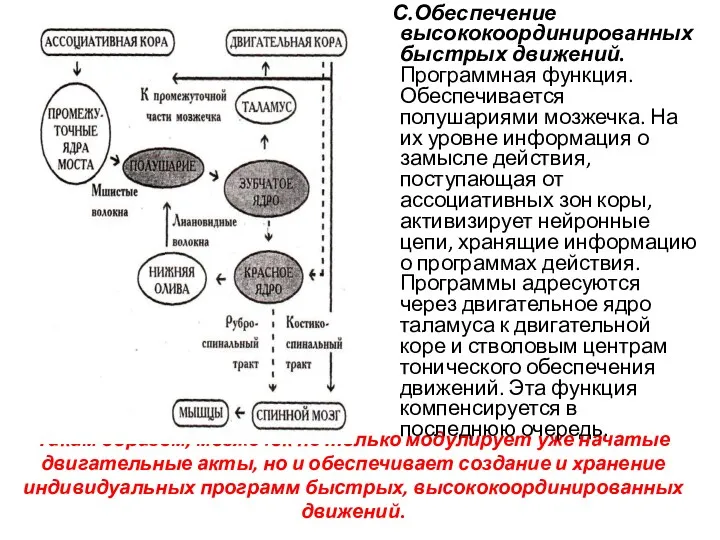
Таким образом, мозжечок не только модулирует уже начатые двигательные акты, но
Таким образом, мозжечок не только модулирует уже начатые двигательные акты, но
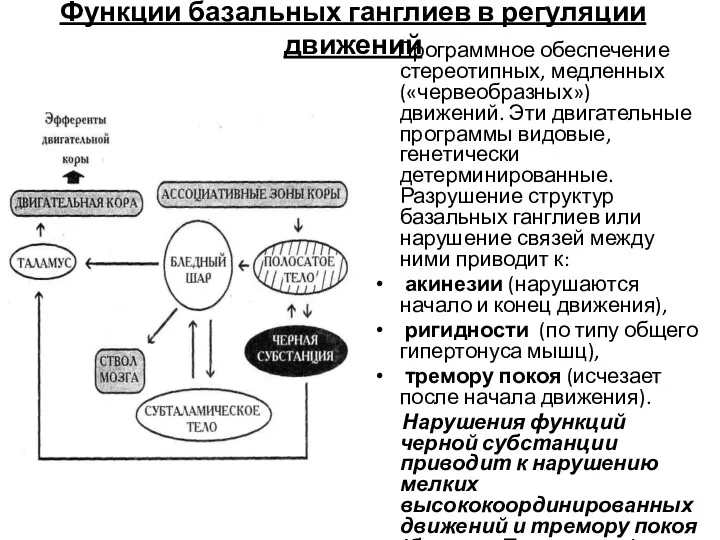
Функции базальных ганглиев в регуляции движений
Программное обеспечение стереотипных, медленных («червеобразных»)
Функции базальных ганглиев в регуляции движений
Программное обеспечение стереотипных, медленных («червеобразных»)

К стриопаллидарной системе (система базальных ганглиев) относятся структуры мозга:
полосатое тело
К стриопаллидарной системе (система базальных ганглиев) относятся структуры мозга: полосатое тело

Двигательная кора в обеспечении моторного действия
Функционально является низшим звеном организации программы
Двигательная кора в обеспечении моторного действия
Функционально является низшим звеном организации программы

ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ двигательной коры в обеспечении моторного действия
Эфферентные пути образованы
ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ двигательной коры в обеспечении моторного действия
Эфферентные пути образованы

Побуждение и замысел действия
Реализуются на уровне мотивационных зон коры и подкорки,
Побуждение и замысел действия
Реализуются на уровне мотивационных зон коры и подкорки,
 Анатомо-физиологические и психологические особенности лиц пожилого и старческого возраста
Анатомо-физиологические и психологические особенности лиц пожилого и старческого возраста Сущность и особенности оценки качества медицинской помощи. Формирование стандартов качества различных видов медицинской помощи
Сущность и особенности оценки качества медицинской помощи. Формирование стандартов качества различных видов медицинской помощи Florence Nightingale
Florence Nightingale Заболевания кожи
Заболевания кожи Ампутации и экзартикуляции конечностей
Ампутации и экзартикуляции конечностей Гепатит С
Гепатит С СПАО Ингосстрах. Предложение по страхованию физических лиц. Добровольное и международное медицинское страхование
СПАО Ингосстрах. Предложение по страхованию физических лиц. Добровольное и международное медицинское страхование Пневмоторакс. Гидроторакс. Гемоторакс
Пневмоторакс. Гидроторакс. Гемоторакс Рани. Види ран
Рани. Види ран Болезнь Крейтцфельда-Якоба
Болезнь Крейтцфельда-Якоба Психосоматика. Психологические тесты в психосоматической практике
Психосоматика. Психологические тесты в психосоматической практике Рак молочной железы
Рак молочной железы Мұрынның және оның қуысының анатомиясы, қан айналым ерекшелігі
Мұрынның және оның қуысының анатомиясы, қан айналым ерекшелігі Трансплантация печени
Трансплантация печени Современные алгоритмы лечения СД и его осложнений
Современные алгоритмы лечения СД и его осложнений Первая помощь при травмах скелета и мышц
Первая помощь при травмах скелета и мышц Первая помощь при ранениях и кровотечениях
Первая помощь при ранениях и кровотечениях ОРВИ и грипп
ОРВИ и грипп Клиническая психология
Клиническая психология Использование бесприборных тест-систем Иммунокомб. Памятка для заводчиков и владельцев питомников кошек
Использование бесприборных тест-систем Иммунокомб. Памятка для заводчиков и владельцев питомников кошек Ишемическая болезнь сердца. Сердечная недостаточность
Ишемическая болезнь сердца. Сердечная недостаточность Обращение товаров аптечного ассортимента (ТАА) на фармацевтическом рынке. Классификация ТАА. Фармацевтическое товароведение
Обращение товаров аптечного ассортимента (ТАА) на фармацевтическом рынке. Классификация ТАА. Фармацевтическое товароведение Организация кабинета медицинского массажа. (Тема 1.1)
Организация кабинета медицинского массажа. (Тема 1.1) Жүрек - қантамырлар жүйесі патологиясы бойынша сканограмма,рентгенограмма және сонограмма альбомын жасау
Жүрек - қантамырлар жүйесі патологиясы бойынша сканограмма,рентгенограмма және сонограмма альбомын жасау Клинические маски стресса в разном возрасте
Клинические маски стресса в разном возрасте Нарушения памяти при локальных поражениях мозга, проблема амнезий
Нарушения памяти при локальных поражениях мозга, проблема амнезий Клеточное строение костного мозга
Клеточное строение костного мозга Вітамін С - потужний антиоксидант
Вітамін С - потужний антиоксидант