- Иммунный ответ

Содержание
- 2. СОДЕРЖАНИЕ 1 Стадии иммунного ответа 2 Взаимодействие клеток в ходе иммунных ответов 3 Процессинг антигена и
- 3. Суть любого иммунного ответа состоит в антигензависимой дифференцировке или прайминге лимфоцитарного клона до его способности к
- 4. МАГИСТРАЛЬНЫЕ ПУТИ ИММУННОГО ОТВЕТА Имеется два магистральных пути иммунного ответа: Клеточный или T-клеточно-опосредованный ответ Гуморальный или
- 5. СТАДИИ ИММУННОГО ОТВЕТА 1. Эндоцитоз антигена, его процессинг и загрузка на молекулы HLA I или II
- 6. КЛОНАЛЬНАЯ ЭКСПАНСИЯ МОДЕЛЬ T-КЛЕТОЧНЫХ ОТВЕТОВ Macrophage Naive CD4+ T cell CD4+ T helper 1 AКТИВАЦИЯ КЛОНАЛЬНАЯ
- 7. МОДЕЛЬ B-КЛЕТОЧНЫХ ОТВЕТОВ B cell РАСПОЗНАВАНИЕ AКТИВАЦИЯ КЛОНАЛЬНАЯ ЭКСПАНСИЯ Dendritic cell (type 2) Naive CD4+T cell
- 8. AНТИГЕНПРЕДСТАВЛЯЮЩИЕ КЛЕТКИ Антигенпредставляющая клетка (макрофаг, дендритная клетка и В-лимфоцит) сталкивается с нативным антигеном и эндоцитирует его.
- 9. ПРОЦЕССИНГ АНТИГЕНА Следующее событие, процессинг, предполагает разборку молекулы антигена внутри антигенпредставляющей клетки. Процессированный антиген приобретает иммуногенные
- 10. ПУТЬ ПРОЦЕССИНГА, СВЯЗАННЫЙ С HLA II Этот путь предполагает процессинг экзогенных антигенов, которые презентируются в ассоциации
- 11. Эндогенные и внутриклеточно расположенные антигены микробного происхождения загружаются на молекулы HLA I для представления наивным CD8+
- 12. ПРОЦЕССИНГ И ПРЕЗЕНТАЦИЯ АНТИГЕНА
- 14. РАСПОЗНАВАНИЕ АНТИГЕНА Многие антигены попадают внутрь человеческого тела, но для включения специфических иммунных ответов Т- и
- 15. ТИПЫ СТИМУЛОВ Распознавание в ходе иммунного ответа предполагает восприятие лимфоцитами трёх типов стимулов, одного специфического и
- 16. РАСПОЗНАВАНИЕ КОМПЛЕКСА АНТИГЕН/HLA I или II Специфический αβTCR и кoрецепторы CD4 или CD8 распознают иммуноген/HLA II
- 18. Антигенпредставляющая клетка Т-хелпер 1 или 2 Ag/HLA II TCR/CD3 CD4
- 19. НОБЕЛЕВСКАЯ ПРЕМИЯ (1996) P.C.DOHERTY R.M.ZINKERNAGEL Открытие «двойного распознавания»
- 20. РАСПОЗНАВАНИЕ Тh1-ЗАВИСИМЫЙ ПУТЬ Специфический сигнал Специфический сигнал Неспецифический сигнал от костимулирующих молекул Неспецифический сигнал от костимулирующих
- 21. РАСПОЗНАВАНИЕ Тh2-ЗАВИСИМЫЙ ПУТЬ Специфический сигнал Неспецифический сигнал от костимулирующих молекул Неспецифический цитокиновый сигнал Специфический сигнал был
- 22. Костимулирующие молекулы семейства B7 (на антигенпредставляющих клетках), главным образом, B7-1 (CD80) и B7-2 (CD86), через их
- 23. CD4+T-ХЕЛПЕР ТИПА 1 ПРЕДОСТАВЛЯЕТ «ПОМОЩЬ» НАИВНОЙ CD8+T-КЛЕТКЕ 1. Прямое взаимодействие CD40L-CD40 ? 2. Стимуляция повышенной экспрессии
- 24. Т- и В-лимфоциты получают неспецифические цито-киновые сигналы не только от антигенпредстав-ляющих, но и других клеток (например,
- 25. TРАНСДУКЦИЯ СИГНАЛА Специфические сигналы, чтобы иметь последующую эффективность, должны быть преобразованы и усилены внутри принимающей клетки
- 27. Сигнальный путь к транскрипции гена IL2
- 28. Сигнальный путь к транскрипции генов структурных белков
- 29. НОБЕЛЕВСКАЯ ПРЕМИЯ (1999) G. BLOBEL Работы в области сигнальной трансдукции
- 30. КЛОНАЛЬНАЯ ЭКСПАНСИЯ Т-КЛЕТОК Клональная экспансия Т-лимфоцитов происходит в паракортикальных зонах лимфатических узлов и периартериолярных пространствах селезёнки.
- 31. КЛОНАЛЬНАЯ ЭКСПАНСИЯ В-КЛЕТОК Клональная экспансия В-лимфоцитов происходит в селезёнке и первичных фолликулах (зародышевых центрах) лимфатических узлов
- 32. СЕЛЕКЦИЯ ВЫСОКОАФФИННЫХ BCR ПРИ КЛОНАЛЬНОЙ ЭКСПАНСИИ CD19?CD23 Bcl-2+ Апоптоз отменяется Если BCR и кoрецептор CD19 (на
- 33. ДИФФЕРЕНЦИРОВКА Т-КЛЕТОК Созревание Т-лимфоцитов происходит в пери-ферических органах иммунной системы, где пред-шественники Т-эффекторов под влиянием цитокинов
- 34. ДИФФЕРЕНЦИРОВКА В-КЛЕТОК Созревание В-клеток начинается в лимфатических органах, а заканчивается в костном мозге. Некоторые В-клетки мигрируют
- 35. ПЕРЕКЛЮЧЕНИЕ ИЗОТИПОВ ИММУНОГЛОБУЛИНОВ CD40L ? CD40 Острый инфекционный эпизод 1-й день 5-й день 9-й день
- 36. ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ ПЕРЕКЛЮЧЕНИЯ ИЗОТИПОВ IgM IgG1 IgG2/3 IgA IgG4 IgE T helper 1 Plasma cell IL4,IL5,IL6,IL13
- 37. ЭФФЕКТОРНЫЕ Т- И В-КЛЕТКИ Дифференцирующиеся T-клетки только изменяют свой фенотип, тогда как B-клетки через несколько морфологических
- 39. КЛЕТКИ ПАМЯТИ T-клетки памяти имеют фенотип: CD45RO, VLA-4hi, CD44hi, LFA-1hi, CD2hi, LFA-3hi, быстрый рециклинг, не зависящий
- 40. В-КЛЕТКИ ПЯМЯТИ Первичная инфекция ? Клинические симптомы есть Tа же самая инфекция Нет клинических проявлений ?
- 42. Скачать презентацию

СОДЕРЖАНИЕ
1 Стадии иммунного ответа
2 Взаимодействие клеток в ходе иммунных ответов
3 Процессинг
СОДЕРЖАНИЕ
1 Стадии иммунного ответа
2 Взаимодействие клеток в ходе иммунных ответов
3 Процессинг

Суть любого иммунного ответа состоит в антигензависимой дифференцировке или прайминге лимфоцитарного
Суть любого иммунного ответа состоит в антигензависимой дифференцировке или прайминге лимфоцитарного

МАГИСТРАЛЬНЫЕ ПУТИ ИММУННОГО ОТВЕТА
Имеется два магистральных пути иммунного ответа:
Клеточный или
МАГИСТРАЛЬНЫЕ ПУТИ ИММУННОГО ОТВЕТА
Имеется два магистральных пути иммунного ответа:
Клеточный или

СТАДИИ ИММУННОГО ОТВЕТА
1. Эндоцитоз антигена, его процессинг и загрузка на
СТАДИИ ИММУННОГО ОТВЕТА
1. Эндоцитоз антигена, его процессинг и загрузка на
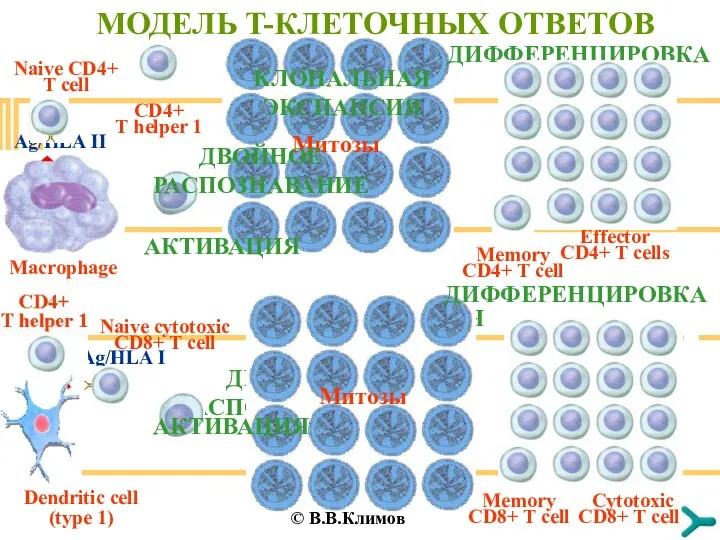
КЛОНАЛЬНАЯ ЭКСПАНСИЯ
МОДЕЛЬ T-КЛЕТОЧНЫХ ОТВЕТОВ
Macrophage
Naive CD4+ T cell
CD4+ T helper 1
AКТИВАЦИЯ
КЛОНАЛЬНАЯ ЭКСПАНСИЯ
ДИФФЕРЕНЦИРОВКА
Effector
КЛОНАЛЬНАЯ ЭКСПАНСИЯ
МОДЕЛЬ T-КЛЕТОЧНЫХ ОТВЕТОВ
Macrophage
Naive CD4+ T cell
CD4+ T helper 1
AКТИВАЦИЯ
КЛОНАЛЬНАЯ ЭКСПАНСИЯ
ДИФФЕРЕНЦИРОВКА
Effector
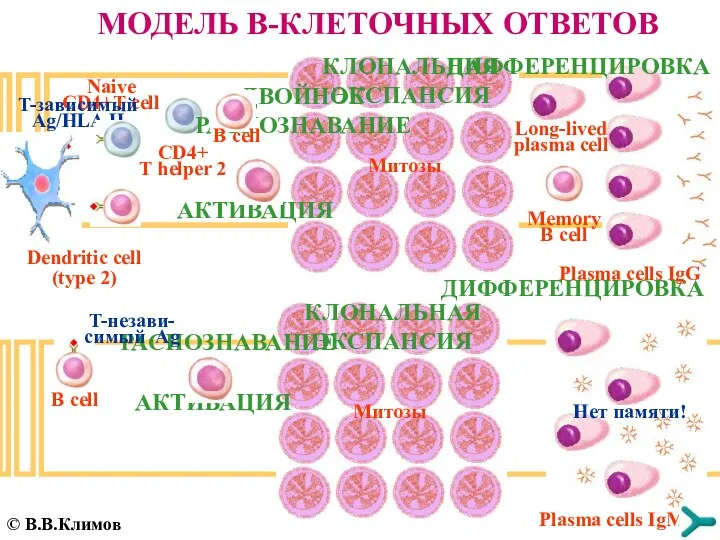
МОДЕЛЬ B-КЛЕТОЧНЫХ ОТВЕТОВ
B cell
РАСПОЗНАВАНИЕ
AКТИВАЦИЯ
КЛОНАЛЬНАЯ ЭКСПАНСИЯ
Dendritic cell (type 2)
Naive CD4+T cell
CD4+ T
МОДЕЛЬ B-КЛЕТОЧНЫХ ОТВЕТОВ
B cell
РАСПОЗНАВАНИЕ
AКТИВАЦИЯ
КЛОНАЛЬНАЯ ЭКСПАНСИЯ
Dendritic cell (type 2)
Naive CD4+T cell
CD4+ T
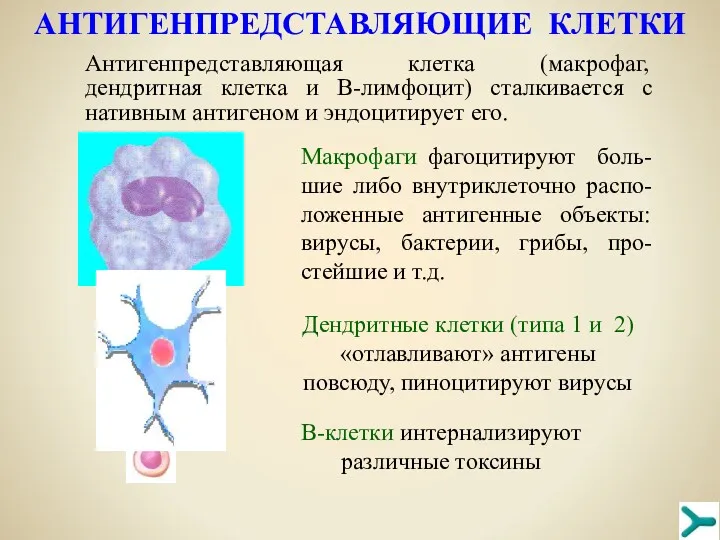
AНТИГЕНПРЕДСТАВЛЯЮЩИЕ КЛЕТКИ
Антигенпредставляющая клетка (макрофаг, дендритная клетка и В-лимфоцит) сталкивается с нативным
AНТИГЕНПРЕДСТАВЛЯЮЩИЕ КЛЕТКИ
Антигенпредставляющая клетка (макрофаг, дендритная клетка и В-лимфоцит) сталкивается с нативным

ПРОЦЕССИНГ АНТИГЕНА
Следующее событие, процессинг, предполагает разборку молекулы антигена внутри антигенпредставляющей клетки.
ПРОЦЕССИНГ АНТИГЕНА
Следующее событие, процессинг, предполагает разборку молекулы антигена внутри антигенпредставляющей клетки.

ПУТЬ ПРОЦЕССИНГА, СВЯЗАННЫЙ С HLA II
Этот путь предполагает процессинг экзогенных антигенов,
ПУТЬ ПРОЦЕССИНГА, СВЯЗАННЫЙ С HLA II
Этот путь предполагает процессинг экзогенных антигенов,

Эндогенные и внутриклеточно расположенные антигены микробного происхождения загружаются на молекулы HLA
Эндогенные и внутриклеточно расположенные антигены микробного происхождения загружаются на молекулы HLA
ПРОЦЕССИНГ И ПРЕЗЕНТАЦИЯ АНТИГЕНА
ПРОЦЕССИНГ И ПРЕЗЕНТАЦИЯ АНТИГЕНА
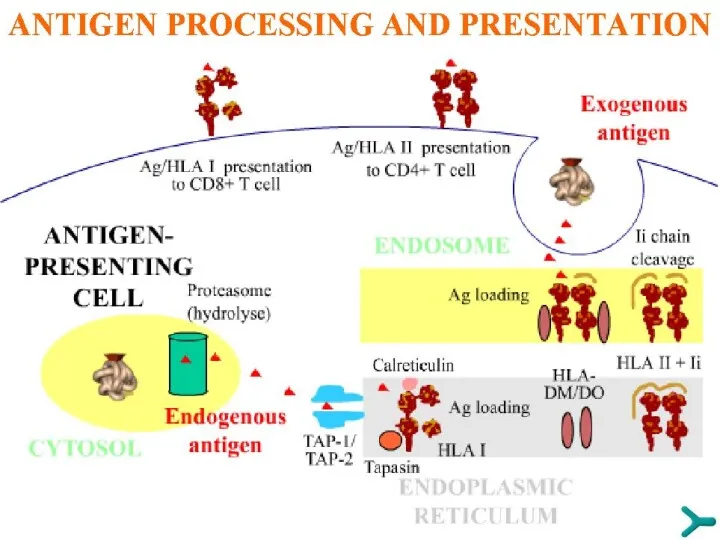

РАСПОЗНАВАНИЕ АНТИГЕНА
Многие антигены попадают внутрь человеческого тела, но для включения специфических
РАСПОЗНАВАНИЕ АНТИГЕНА
Многие антигены попадают внутрь человеческого тела, но для включения специфических

ТИПЫ СТИМУЛОВ
Распознавание в ходе иммунного ответа предполагает восприятие лимфоцитами трёх типов
ТИПЫ СТИМУЛОВ
Распознавание в ходе иммунного ответа предполагает восприятие лимфоцитами трёх типов

РАСПОЗНАВАНИЕ КОМПЛЕКСА АНТИГЕН/HLA I или II
Специфический αβTCR и кoрецепторы CD4
РАСПОЗНАВАНИЕ КОМПЛЕКСА АНТИГЕН/HLA I или II
Специфический αβTCR и кoрецепторы CD4
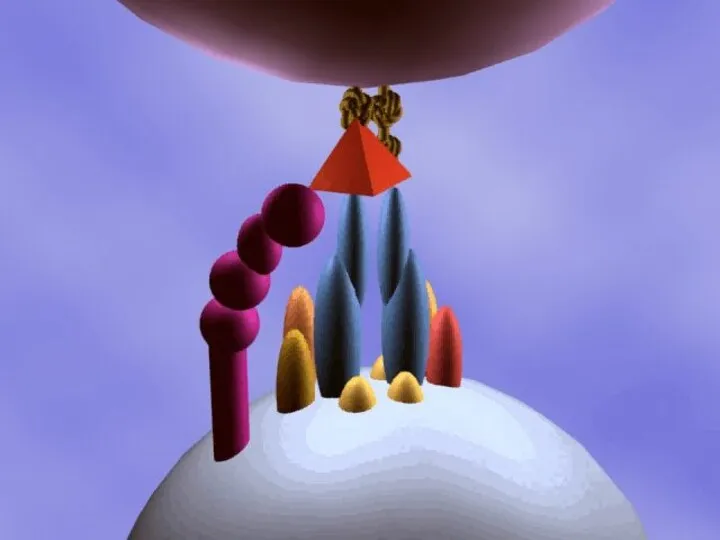
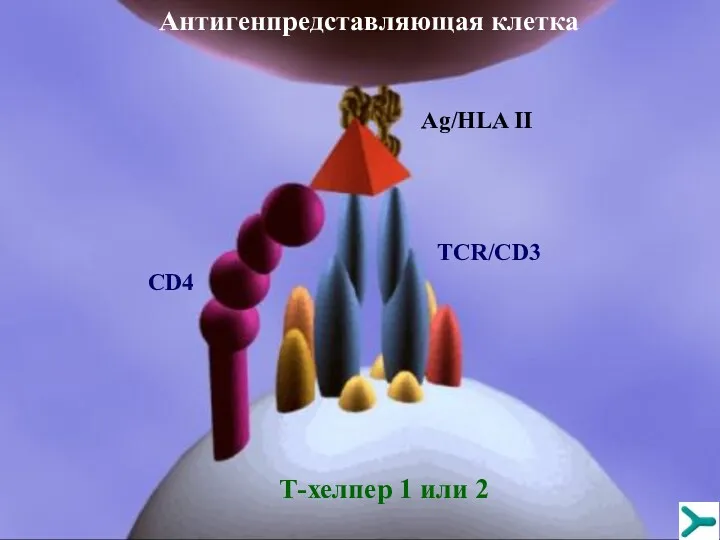
Антигенпредставляющая клетка
Т-хелпер 1 или 2
Ag/HLA II
TCR/CD3
CD4
Антигенпредставляющая клетка
Т-хелпер 1 или 2
Ag/HLA II
TCR/CD3
CD4

НОБЕЛЕВСКАЯ ПРЕМИЯ (1996)
P.C.DOHERTY
R.M.ZINKERNAGEL
Открытие «двойного распознавания»
НОБЕЛЕВСКАЯ ПРЕМИЯ (1996)
P.C.DOHERTY
R.M.ZINKERNAGEL
Открытие «двойного распознавания»
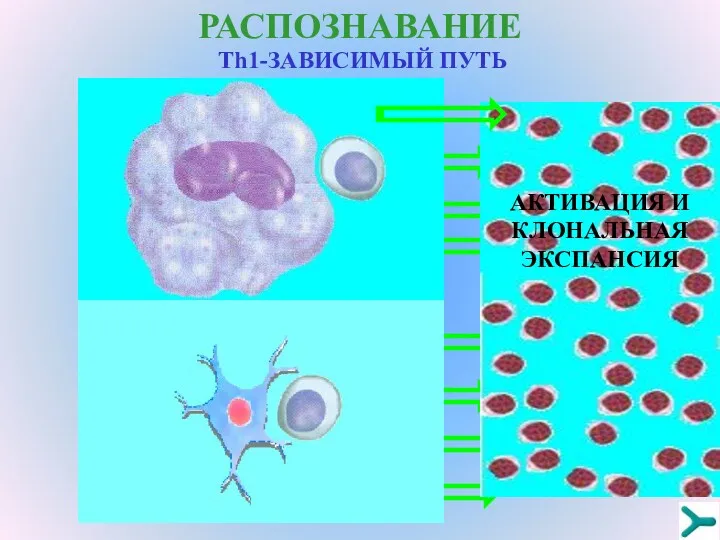
РАСПОЗНАВАНИЕ
Тh1-ЗАВИСИМЫЙ ПУТЬ
Специфический сигнал
Специфический сигнал
Неспецифический сигнал от костимулирующих молекул
Неспецифический сигнал
РАСПОЗНАВАНИЕ
Тh1-ЗАВИСИМЫЙ ПУТЬ
Специфический сигнал
Специфический сигнал
Неспецифический сигнал от костимулирующих молекул
Неспецифический сигнал

РАСПОЗНАВАНИЕ
Тh2-ЗАВИСИМЫЙ ПУТЬ
Специфический сигнал
Неспецифический сигнал от костимулирующих молекул
Неспецифический цитокиновый сигнал
Специфический сигнал был
РАСПОЗНАВАНИЕ
Тh2-ЗАВИСИМЫЙ ПУТЬ
Специфический сигнал
Неспецифический сигнал от костимулирующих молекул
Неспецифический цитокиновый сигнал
Специфический сигнал был

Костимулирующие молекулы семейства B7 (на антигенпредставляющих клетках), главным образом, B7-1 (CD80)
Костимулирующие молекулы семейства B7 (на антигенпредставляющих клетках), главным образом, B7-1 (CD80)
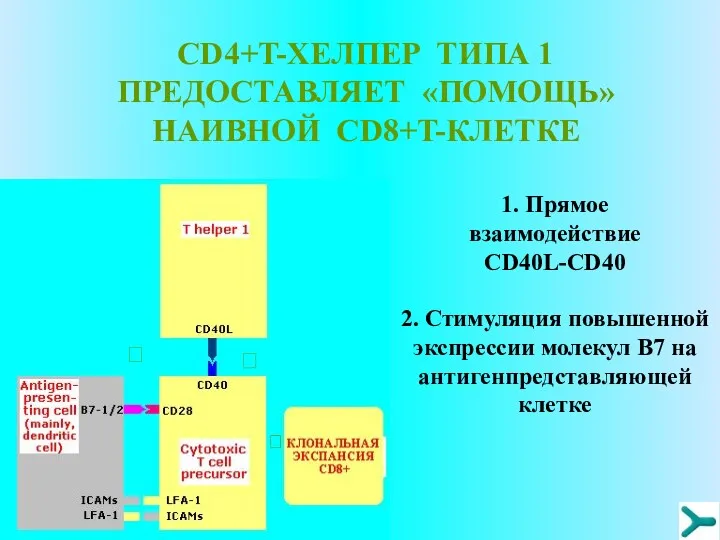
CD4+T-ХЕЛПЕР ТИПА 1 ПРЕДОСТАВЛЯЕТ «ПОМОЩЬ» НАИВНОЙ CD8+T-КЛЕТКЕ
1. Прямое взаимодействие CD40L-CD40
?
2.
CD4+T-ХЕЛПЕР ТИПА 1 ПРЕДОСТАВЛЯЕТ «ПОМОЩЬ» НАИВНОЙ CD8+T-КЛЕТКЕ
1. Прямое взаимодействие CD40L-CD40
?
2.

Т- и В-лимфоциты получают неспецифические цито-киновые сигналы не только от антигенпредстав-ляющих,
Т- и В-лимфоциты получают неспецифические цито-киновые сигналы не только от антигенпредстав-ляющих,

TРАНСДУКЦИЯ СИГНАЛА
Специфические сигналы, чтобы иметь последующую эффективность, должны быть преобразованы и
TРАНСДУКЦИЯ СИГНАЛА
Специфические сигналы, чтобы иметь последующую эффективность, должны быть преобразованы и

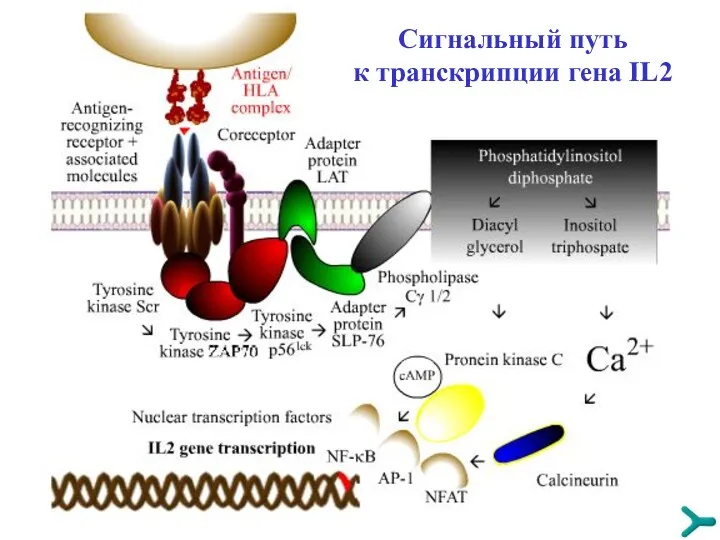
Сигнальный путь к транскрипции гена IL2
Сигнальный путь к транскрипции гена IL2
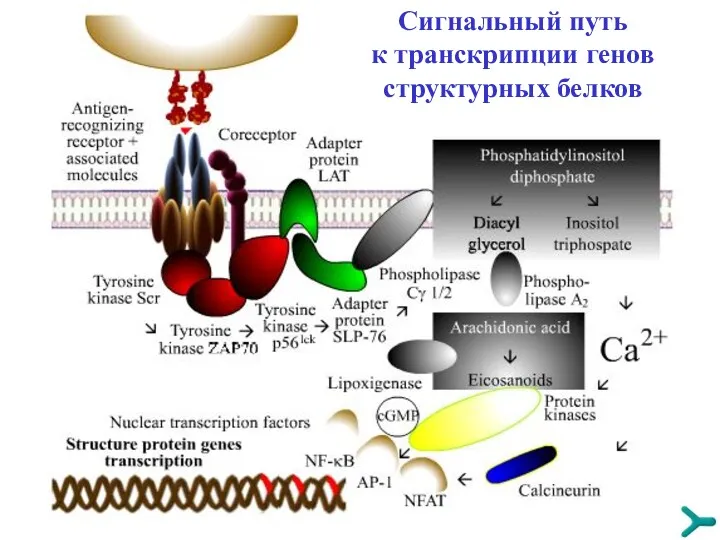
Сигнальный путь к транскрипции генов структурных белков
Сигнальный путь к транскрипции генов структурных белков

НОБЕЛЕВСКАЯ ПРЕМИЯ (1999)
G. BLOBEL
Работы в области сигнальной трансдукции
НОБЕЛЕВСКАЯ ПРЕМИЯ (1999)
G. BLOBEL
Работы в области сигнальной трансдукции

КЛОНАЛЬНАЯ ЭКСПАНСИЯ Т-КЛЕТОК
Клональная экспансия Т-лимфоцитов происходит в паракортикальных зонах лимфатических узлов
КЛОНАЛЬНАЯ ЭКСПАНСИЯ Т-КЛЕТОК
Клональная экспансия Т-лимфоцитов происходит в паракортикальных зонах лимфатических узлов

КЛОНАЛЬНАЯ ЭКСПАНСИЯ В-КЛЕТОК
Клональная экспансия В-лимфоцитов происходит в селезёнке и первичных фолликулах
КЛОНАЛЬНАЯ ЭКСПАНСИЯ В-КЛЕТОК
Клональная экспансия В-лимфоцитов происходит в селезёнке и первичных фолликулах
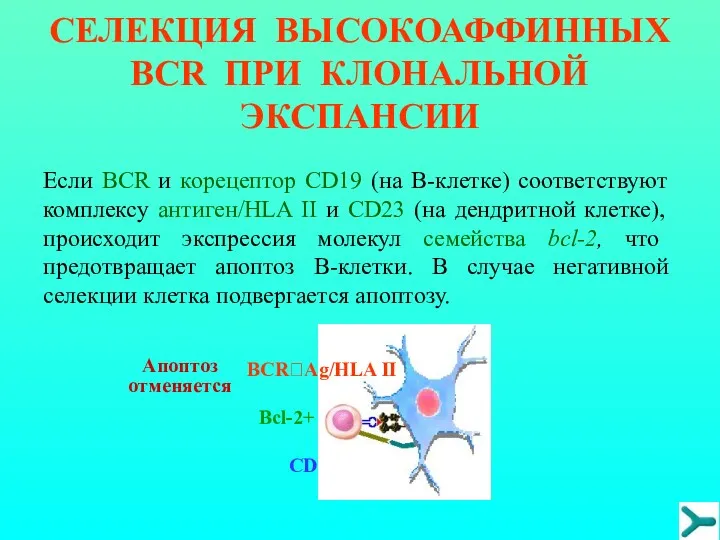
СЕЛЕКЦИЯ ВЫСОКОАФФИННЫХ BCR ПРИ КЛОНАЛЬНОЙ ЭКСПАНСИИ
CD19?CD23
Bcl-2+
Апоптоз отменяется
Если BCR и кoрецептор CD19
СЕЛЕКЦИЯ ВЫСОКОАФФИННЫХ BCR ПРИ КЛОНАЛЬНОЙ ЭКСПАНСИИ
CD19?CD23
Bcl-2+
Апоптоз отменяется
Если BCR и кoрецептор CD19

ДИФФЕРЕНЦИРОВКА Т-КЛЕТОК
Созревание Т-лимфоцитов происходит в пери-ферических органах иммунной системы, где пред-шественники
ДИФФЕРЕНЦИРОВКА Т-КЛЕТОК
Созревание Т-лимфоцитов происходит в пери-ферических органах иммунной системы, где пред-шественники

ДИФФЕРЕНЦИРОВКА В-КЛЕТОК
Созревание В-клеток начинается в лимфатических органах, а заканчивается в костном
ДИФФЕРЕНЦИРОВКА В-КЛЕТОК
Созревание В-клеток начинается в лимфатических органах, а заканчивается в костном

ПЕРЕКЛЮЧЕНИЕ ИЗОТИПОВ ИММУНОГЛОБУЛИНОВ
CD40L ? CD40
Острый инфекционный эпизод
1-й день
5-й день
9-й день
ПЕРЕКЛЮЧЕНИЕ ИЗОТИПОВ ИММУНОГЛОБУЛИНОВ
CD40L ? CD40
Острый инфекционный эпизод
1-й день
5-й день
9-й день
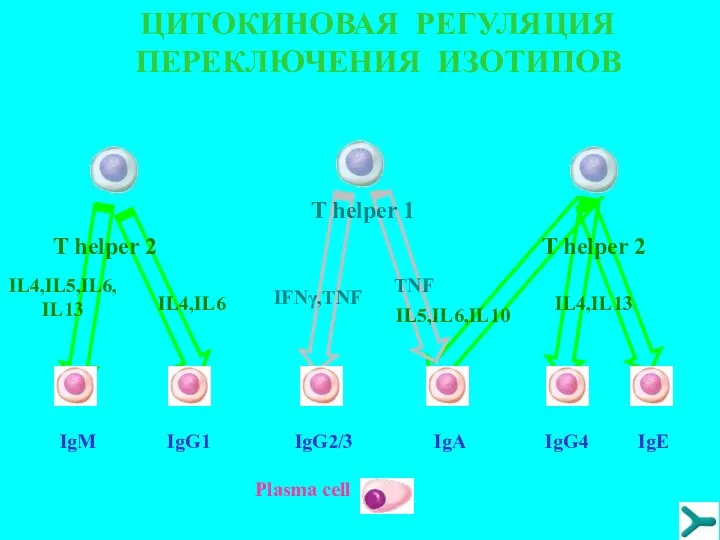
ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ ПЕРЕКЛЮЧЕНИЯ ИЗОТИПОВ
IgM
IgG1
IgG2/3
IgA
IgG4
IgE
T helper 1
Plasma cell
IL4,IL5,IL6,IL13
IL4,IL6
IL5,IL6,IL10
IL4,IL13
IFNγ,TNF
TNF
ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ ПЕРЕКЛЮЧЕНИЯ ИЗОТИПОВ
IgM
IgG1
IgG2/3
IgA
IgG4
IgE
T helper 1
Plasma cell
IL4,IL5,IL6,IL13
IL4,IL6
IL5,IL6,IL10
IL4,IL13
IFNγ,TNF
TNF
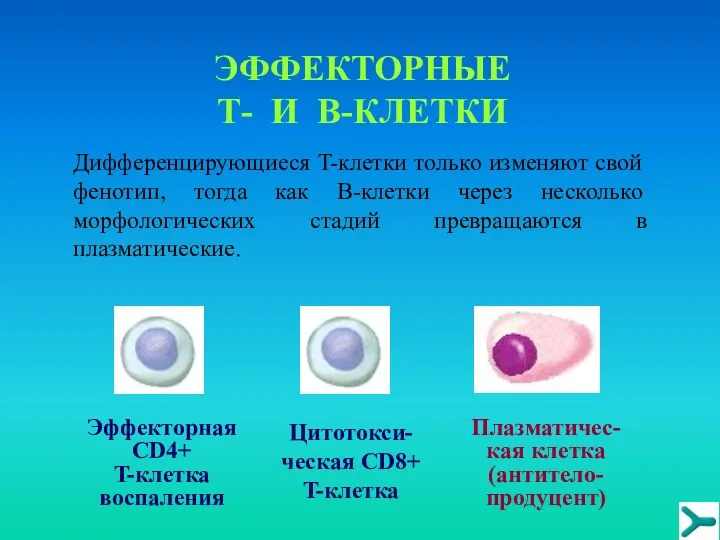
ЭФФЕКТОРНЫЕ Т- И В-КЛЕТКИ
Дифференцирующиеся T-клетки только изменяют свой фенотип, тогда как
ЭФФЕКТОРНЫЕ Т- И В-КЛЕТКИ
Дифференцирующиеся T-клетки только изменяют свой фенотип, тогда как

КЛЕТКИ ПАМЯТИ
T-клетки памяти имеют фенотип: CD45RO, VLA-4hi, CD44hi, LFA-1hi, CD2hi, LFA-3hi,
КЛЕТКИ ПАМЯТИ
T-клетки памяти имеют фенотип: CD45RO, VLA-4hi, CD44hi, LFA-1hi, CD2hi, LFA-3hi,
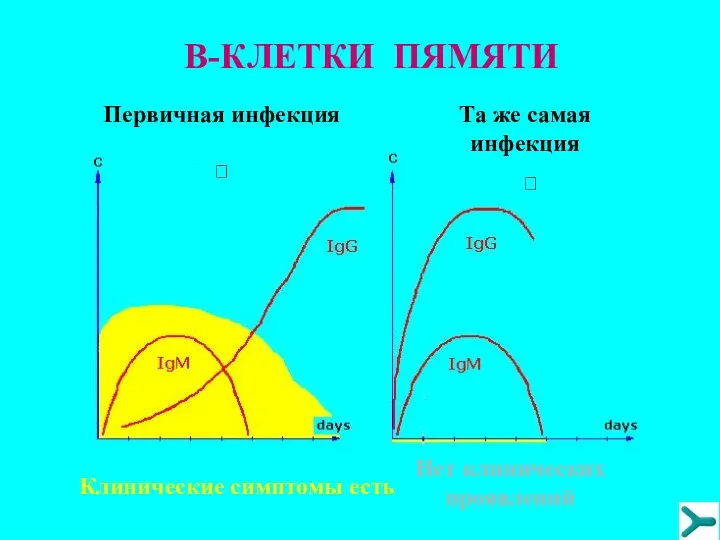
В-КЛЕТКИ ПЯМЯТИ
Первичная инфекция
?
Клинические симптомы есть
Tа же самая инфекция
Нет клинических проявлений
?
В-КЛЕТКИ ПЯМЯТИ
Первичная инфекция
?
Клинические симптомы есть
Tа же самая инфекция
Нет клинических проявлений
?
Похожие презентации
 Лимфоциты, как ключевые клетки адаптивного иммунитета
Лимфоциты, как ключевые клетки адаптивного иммунитета Применение нейронавигации в нейрохирургии
Применение нейронавигации в нейрохирургии Шизоидное и шизотипическое расстройства
Шизоидное и шизотипическое расстройства Тромбоцитопения. Классификация
Тромбоцитопения. Классификация Врожденные пороки сердца без цианоза
Врожденные пороки сердца без цианоза Острый тяжелый панкреатит. Современные принципы диагностики и лечения
Острый тяжелый панкреатит. Современные принципы диагностики и лечения Пломбировочный материал амальгама в зубоврачевании
Пломбировочный материал амальгама в зубоврачевании Т-клеточное звено иммунитета. Субпопуляции Т-лимфоцитов. Регуляторная роль CD4-лимфоцитов. Иммунный ответ по Тх1 пути
Т-клеточное звено иммунитета. Субпопуляции Т-лимфоцитов. Регуляторная роль CD4-лимфоцитов. Иммунный ответ по Тх1 пути Патофизиология печени. Печеночная недостаточность
Патофизиология печени. Печеночная недостаточность Работа сестринского персонала по обеспечению инфекционной безопасности в стоматологической клинике
Работа сестринского персонала по обеспечению инфекционной безопасности в стоматологической клинике Композиционные пломбировочные материалы при лечении кариеса и некариозных поражений твердых тканей зубов
Композиционные пломбировочные материалы при лечении кариеса и некариозных поражений твердых тканей зубов Анестезия при эндохирургических вмешательствах у детей
Анестезия при эндохирургических вмешательствах у детей Грудное вскармливание
Грудное вскармливание Transportnaya_immobilizatsia_2
Transportnaya_immobilizatsia_2 Гипер – IgE синдромы (Джоб синдромы)
Гипер – IgE синдромы (Джоб синдромы) Экстубация трахеи
Экстубация трахеи Пневмонии. Обструктивные и рестриктивные заболевания легких. Рак легких
Пневмонии. Обструктивные и рестриктивные заболевания легких. Рак легких Противоопухолевый иммунитет
Противоопухолевый иммунитет Қоршаған орта факторларының екіншілік ИТЖ дамуындағы маңызы
Қоршаған орта факторларының екіншілік ИТЖ дамуындағы маңызы Паралеллометрия әдісін жүргізу, паралеллометрдің құрылысы
Паралеллометрия әдісін жүргізу, паралеллометрдің құрылысы Хромосомные мутации
Хромосомные мутации
 Выпотной перикардит (экссудативный перикардит)
Выпотной перикардит (экссудативный перикардит) Неэпилептические пароксизмальные расстройства сознания
Неэпилептические пароксизмальные расстройства сознания Сравнительная характеристика уровня медицины воюющих государств в период Крымской войны
Сравнительная характеристика уровня медицины воюющих государств в период Крымской войны Сердечно-легочная реанимация
Сердечно-легочная реанимация Язвенно-некротический гингивит Венсана
Язвенно-некротический гингивит Венсана Анализ вспышки заболеваемости корью в г. Екатеринбург в 2018 году
Анализ вспышки заболеваемости корью в г. Екатеринбург в 2018 году