Т-клеточное звено иммунитета. Субпопуляции Т-лимфоцитов. Регуляторная роль CD4-лимфоцитов. Иммунный ответ по Тх1 пути презентация
- Т-клеточное звено иммунитета. Субпопуляции Т-лимфоцитов. Регуляторная роль CD4-лимфоцитов. Иммунный ответ по Тх1 пути

Содержание
- 2. Лимфоцит – центральная фигура в иммунной системе В организме человека содержится 1013 лимфоцитов (каждая 10 клетка
- 3. В развитии специфического иммунного ответа принимают участие три основных клеточных типа: В‑лимфоциты, Т‑лимфоциты антигенпрезентирующие клетки –
- 4. Т- и В-лимфоциты – истинные иммунокомпетентные клетки, способные: Распознавать АГ с помощью антигенспецифических рецепторов (TCR, BCR)
- 6. В развитии В- и Т-лимфоцитов выделяют 2 периода: антиген-зависимый (иммунопоэз) антигеннезависимый (лимфопоэз) Лимфопоэз - образование (из
- 8. В-лимфоциты – отвечают за гуморальный иммунный ответ Зрелые В-лимфоциты составляют 60-70% всех В-клеток крови. Срок жизни
- 10. Типы молекул на поверхности В-лимфоцитов
- 11. В-лимфоциты В1(CD5+) Врожденная субпопуляция В-лимфоцитов В2(CD5-) Зрелые В-лимфоциты Локализуются в лимфоидных скоплениях слизистых оболочек и кожи
- 12. Также известны В-клетки маргинальной зоны: Занимают промежуточное положение между В1- и В2-клетками Располагаются в маргинальной зоне
- 13. Из своего неактивного состояния В-клетки могут быть стимулированы: к росту, к развитию синтетического аппарата, к делению,
- 14. Активация В-клеток также подразумевает "переключение классов Ig“: первые антителопроизводящие В-клетки продуцируют IgM; позднее при первичном ответе,
- 15. Т-лимфоциты – главная популяция в развитии клеточного иммунного ответа Развитие Т-лф зависит от тимуса, в котором
- 16. Роль тимуса в Т-клеточной продукции Тимоциты (Т-клеточные предшественники из костного мозга) ТИМУС Зрелые (наивные) Т-лимфоциты CD4+
- 17. Типы молекул на поверхности Т-лимфоцитов
- 18. Стадии антиген-независимой дифференцировки Т-лимфоцитов протимоцит Носители CD2,CD7 Пролиферируют и развиваются под действием ИЛ-1,2,6,7. Мигрируют из КМ
- 19. Т-лимфоциты Периферические Т-лимфоциты-эффекторы Т-лимфоциты-регуляторы Т-лф -киллеры (CD8+) Распознают уникальные антигены, связанные с МСН-1 на поверхности любых
- 20. СD4+- Т-хелперы - функциональная субпопуляция Т-клеток Продуцируют различные цитокины Участвуют в распознавания АГ в комплексе с
- 21. Дифференцировка Т-хелперных клонов Тх0 Тх1 Тх2 Тх 17 Тх10 CD4+ CD25+ T-reg ИЛ-12 ИЛ-18 ИЛ-4 ИЛ-33
- 22. Тх1 Тх2 Тх17 CD4+ CD25+ T-reg МФ ЭОЗ НГ ДК Защита от внутриклеточных патогенов Защита от
- 23. Цитотоксические CD8+-(ЦТЛ, Т-киллеры) Распознают антигены в комплексе с молекулами HLA- II и поражают: инфицированные вирусом клетки-мишени
- 24. ХАРАКТЕРИСТИКА ТРЕХ ГЛАВНЫХ ТИПОВ Т-ЛИМФОЦИТОВ
- 25. Регуляторные CD4+CD25+FoxP3+ - T-лимфоциты (Treg) Естественные, развивающиеся в тимусе, вне АГ-стимуляции (Treg) Индуцированные на периферии из
- 26. ЗАБОЛЕВАНИЯ, ПАТОГЕНЕТИЧЕСКИ СВЯЗАННЫЕ С НАРУШЕНИЕМ ЕСТЕСТВЕННЫХ РЕГУЛЯТОРНЫХ Т-КЛЕТОК Содержание CD4+CD25+Foxp3+ Т-клеток часто понижено при атопическом дерматите;
- 27. NKT-лимфоциты Промежуточный вариант между клетками адаптивного и врожденного иммунитета Экспрессируют общую с NK-клетками молекулу СD161 и
- 28. ДИФФЕРЕНЦИРОВКА Т-ХЕЛПЕРОВ (ВЫБОР ПУТИ ИММУННОГО ОТВЕТА) Этот путь дифференцировки преобладает в слизистой кишечника Этот путь дифференцировки
- 29. Клеточный иммунный ответ (иммунный ответ по Тх-1 пути) Участвуют популяции Тх-1 CD4+ и цитотоксические Т-лимфоциты (CD8+)
- 30. Клеточный иммунный ответ (Тх1-путь)
- 31. ГУМОРАЛЬНЫЙ ИММУННЫЙ ОТВЕТ (Тх1/Тх2-путь иммунного ответа) Активация В-лимфоцитов (В2 CD5-) и их дифференцировка в антителообразующие (АОК)
- 32. Схема гуморального иммунного ответа
- 33. Противобактериальный иммунитет (иммунный ответ по Тх2-пути) Направлен против бактерий и их токсинов Бактерии и токсины нейтрализуются
- 34. Антибактериальный иммунный ответ
- 35. СРАВНИТЕЛЬНЫЕ ХАРАКТЕРИСТИКИ КЛЕТОЧНО-ОПОСРЕДОВАННОГО И ГУМОРАЛЬНОГО ИММУНИТЕТА
- 36. Иммунологическая память Способность иммунной системы (клеток памяти) отвечать ускоренно и усиленно на повторное введение антигена Основа
- 37. Т-клетки памяти могут формировать и при отсутствии контакта с чужеродными АГ: - У здорового новорожденного –
- 38. Формы клеточной гибели Апоптоз Аутофагия Некроз Митотическая катастрофа Ороговение и др. Классификация по данным Комитета по
- 39. Апоптоз Запрограммированная смерть клетки - процесс, посредством которого внутренние или внешние факторы, активируя генетическую программу, приводят
- 41. Последовательность ультраструктурных изменений при апоптозе (справа) и некрозе (слева) 1 – нормальная клетка; 2 – начало
- 42. Апоптоз необходим для элиминации из организма клеток: пораженных различными патогенами, старых клеток, прекращающих выполнять свои функции,
- 44. Скачать презентацию

Лимфоцит – центральная фигура в иммунной системе
В организме человека содержится 1013
Лимфоцит – центральная фигура в иммунной системе
В организме человека содержится 1013
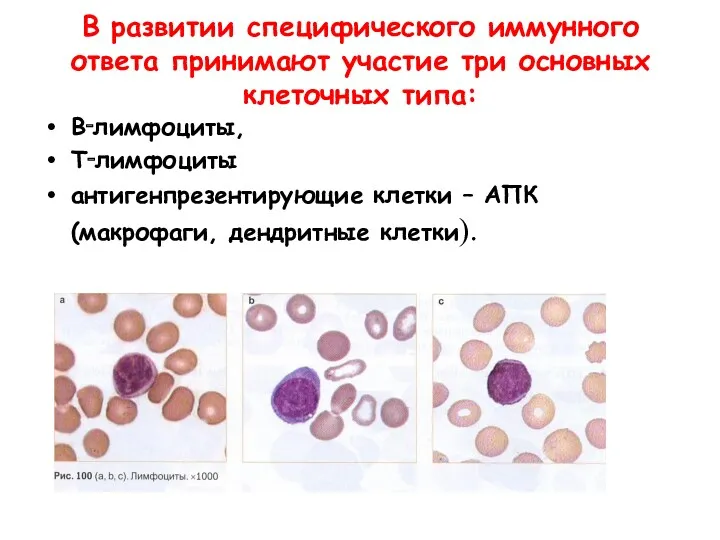
В развитии специфического иммунного ответа принимают участие три основных клеточных типа:
В развитии специфического иммунного ответа принимают участие три основных клеточных типа:

Т- и В-лимфоциты – истинные иммунокомпетентные клетки, способные:
Распознавать АГ с помощью
Т- и В-лимфоциты – истинные иммунокомпетентные клетки, способные:
Распознавать АГ с помощью
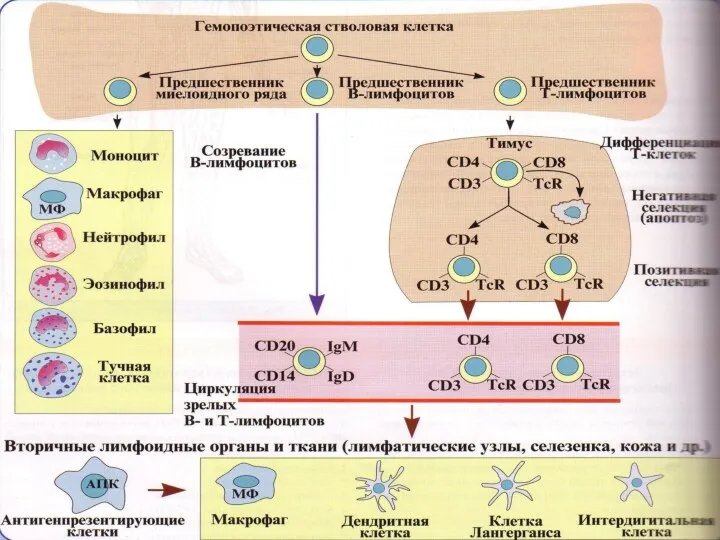
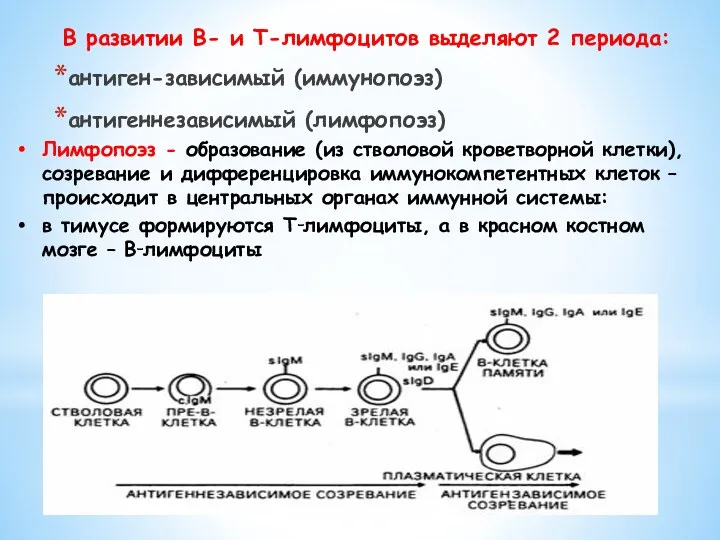
В развитии В- и Т-лимфоцитов выделяют 2 периода:
антиген-зависимый (иммунопоэз)
антигеннезависимый (лимфопоэз)
Лимфопоэз
В развитии В- и Т-лимфоцитов выделяют 2 периода:
антиген-зависимый (иммунопоэз)
антигеннезависимый (лимфопоэз)
Лимфопоэз
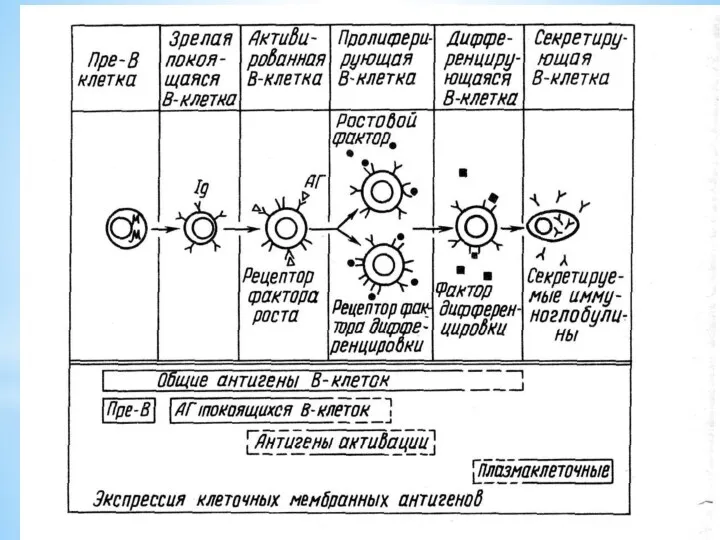

В-лимфоциты – отвечают за гуморальный иммунный ответ
Зрелые В-лимфоциты составляют 60-70%
В-лимфоциты – отвечают за гуморальный иммунный ответ
Зрелые В-лимфоциты составляют 60-70%

Типы молекул на поверхности В-лимфоцитов
Типы молекул на поверхности В-лимфоцитов
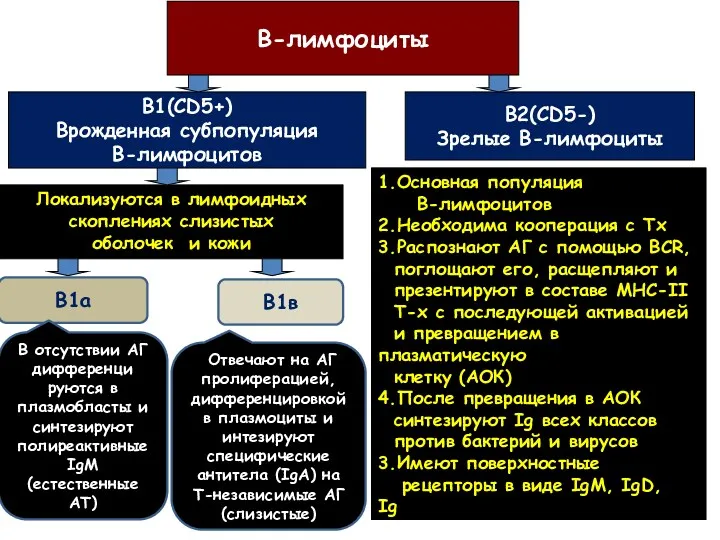
В-лимфоциты
В1(CD5+)
Врожденная субпопуляция
В-лимфоцитов
В2(CD5-)
Зрелые В-лимфоциты
Локализуются в лимфоидных
скоплениях слизистых
оболочек и кожи
1.Основная популяция
В-лимфоциты
В1(CD5+)
Врожденная субпопуляция
В-лимфоцитов
В2(CD5-)
Зрелые В-лимфоциты
Локализуются в лимфоидных
скоплениях слизистых
оболочек и кожи
1.Основная популяция

Также известны В-клетки маргинальной зоны:
Занимают промежуточное положение между В1- и
Также известны В-клетки маргинальной зоны:
Занимают промежуточное положение между В1- и
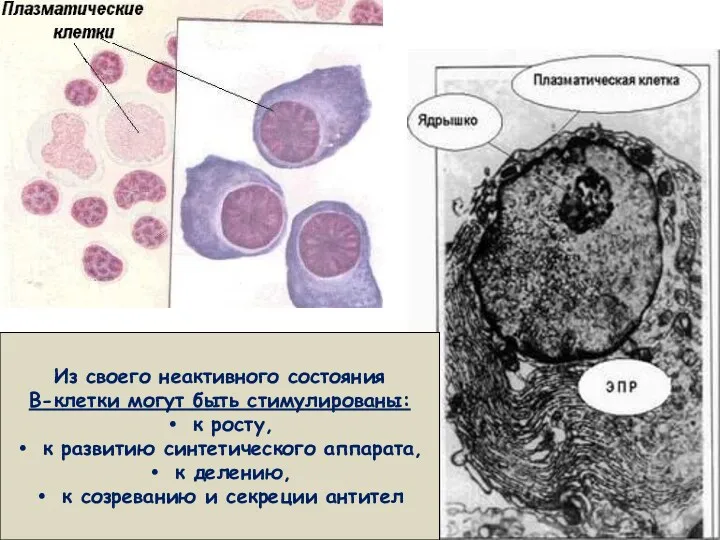
Из своего неактивного состояния
В-клетки могут быть стимулированы:
к росту,
к развитию
Из своего неактивного состояния
В-клетки могут быть стимулированы:
к росту,
к развитию

Активация В-клеток также подразумевает "переключение классов Ig“:
первые антителопроизводящие В-клетки продуцируют IgM;
Активация В-клеток также подразумевает "переключение классов Ig“:
первые антителопроизводящие В-клетки продуцируют IgM;

Т-лимфоциты – главная популяция в развитии клеточного иммунного ответа
Развитие Т-лф
Т-лимфоциты – главная популяция в развитии клеточного иммунного ответа
Развитие Т-лф
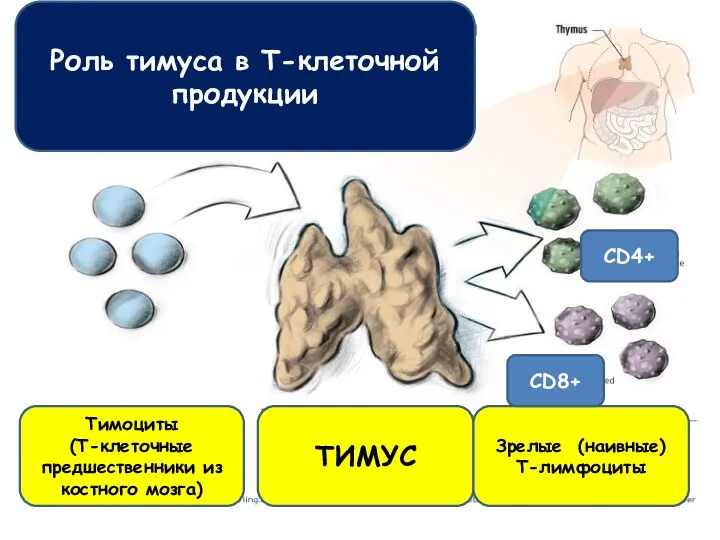
Роль тимуса в Т-клеточной продукции
Тимоциты
(Т-клеточные предшественники из костного мозга)
ТИМУС
Зрелые (наивные)
Т-лимфоциты
CD4+
CD8+
Роль тимуса в Т-клеточной продукции
Тимоциты
(Т-клеточные предшественники из костного мозга)
ТИМУС
Зрелые (наивные)
Т-лимфоциты
CD4+
CD8+

Типы молекул на поверхности Т-лимфоцитов
Типы молекул на поверхности Т-лимфоцитов
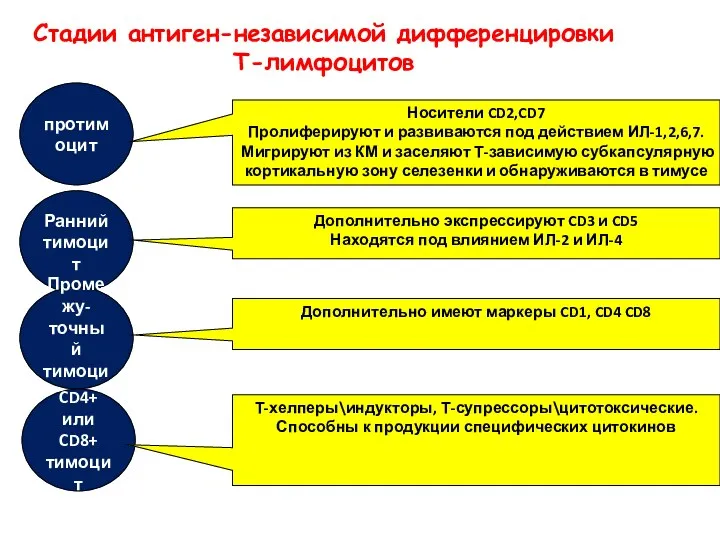
Стадии антиген-независимой дифференцировки
Т-лимфоцитов
протимоцит
Носители CD2,CD7
Пролиферируют и развиваются под действием ИЛ-1,2,6,7.
Стадии антиген-независимой дифференцировки
Т-лимфоцитов
протимоцит
Носители CD2,CD7
Пролиферируют и развиваются под действием ИЛ-1,2,6,7.
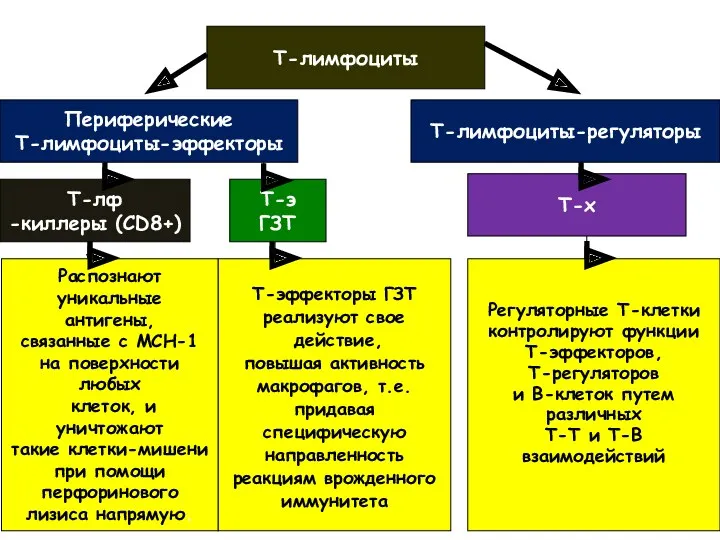
Т-лимфоциты
Периферические
Т-лимфоциты-эффекторы
Т-лимфоциты-регуляторы
Т-лф
-киллеры (CD8+)
Распознают
уникальные антигены,
связанные с МСН-1
на поверхности
любых
клеток, и
Т-лимфоциты
Периферические
Т-лимфоциты-эффекторы
Т-лимфоциты-регуляторы
Т-лф
-киллеры (CD8+)
Распознают
уникальные антигены,
связанные с МСН-1
на поверхности
любых
клеток, и
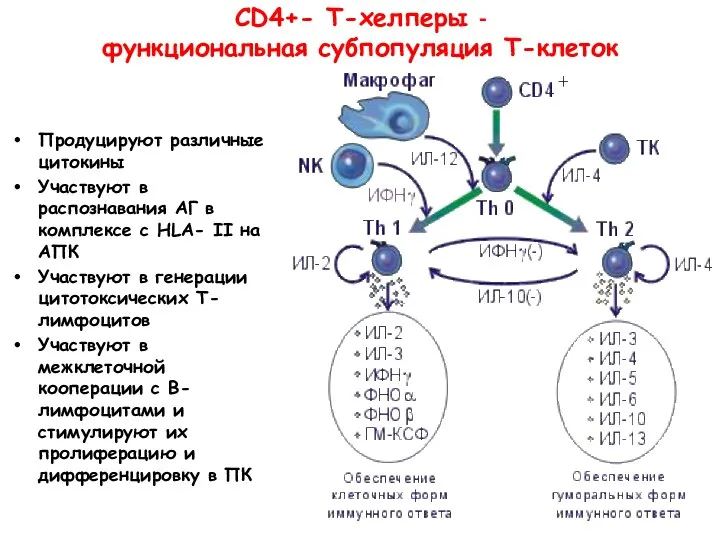
СD4+- Т-хелперы -
функциональная субпопуляция Т-клеток
Продуцируют различные цитокины
Участвуют в распознавания АГ
СD4+- Т-хелперы -
функциональная субпопуляция Т-клеток
Продуцируют различные цитокины
Участвуют в распознавания АГ
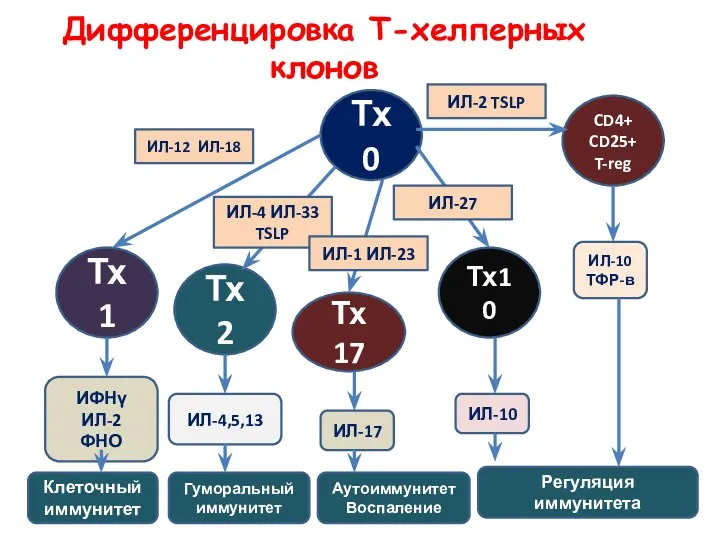
Дифференцировка Т-хелперных клонов
Тх0
Тх1
Тх2
Тх
17
Тх10
CD4+
CD25+
T-reg
ИЛ-12 ИЛ-18
ИЛ-4 ИЛ-33
TSLP
ИЛ-1 ИЛ-23
ИЛ-27
ИЛ-2 TSLP
ИФНγ
ИЛ-2
ФНО
ИЛ-4,5,13
ИЛ-17
ИЛ-10
ИЛ-10
ТФР-в
Клеточный иммунитет
Гуморальный иммунитет
Аутоиммунитет
Воспаление
Регуляция иммунитета
Дифференцировка Т-хелперных клонов
Тх0
Тх1
Тх2
Тх
17
Тх10
CD4+
CD25+
T-reg
ИЛ-12 ИЛ-18
ИЛ-4 ИЛ-33
TSLP
ИЛ-1 ИЛ-23
ИЛ-27
ИЛ-2 TSLP
ИФНγ
ИЛ-2
ФНО
ИЛ-4,5,13
ИЛ-17
ИЛ-10
ИЛ-10
ТФР-в
Клеточный иммунитет
Гуморальный иммунитет
Аутоиммунитет
Воспаление
Регуляция иммунитета
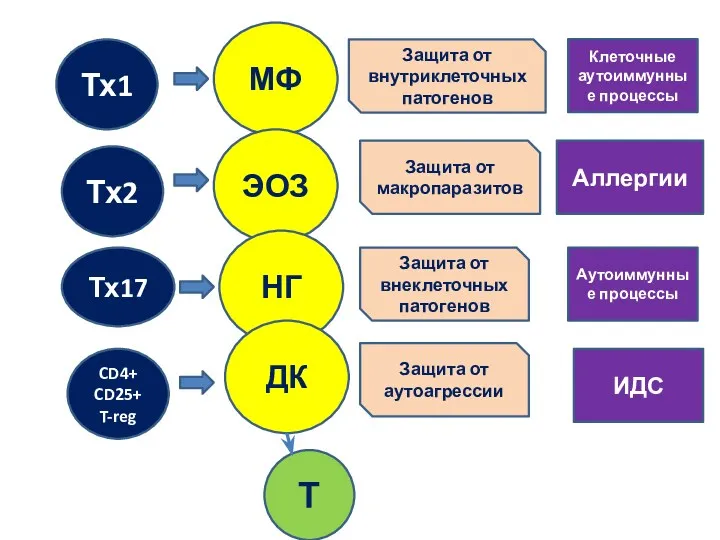
Тх1
Тх2
Тх17
CD4+
CD25+
T-reg
МФ
ЭОЗ
НГ
ДК
Защита от внутриклеточных патогенов
Защита от макропаразитов
Защита от внеклеточных патогенов
Защита от аутоагрессии
Т
ИДС
Аутоиммунные
Тх1
Тх2
Тх17
CD4+
CD25+
T-reg
МФ
ЭОЗ
НГ
ДК
Защита от внутриклеточных патогенов
Защита от макропаразитов
Защита от внеклеточных патогенов
Защита от аутоагрессии
Т
ИДС
Аутоиммунные
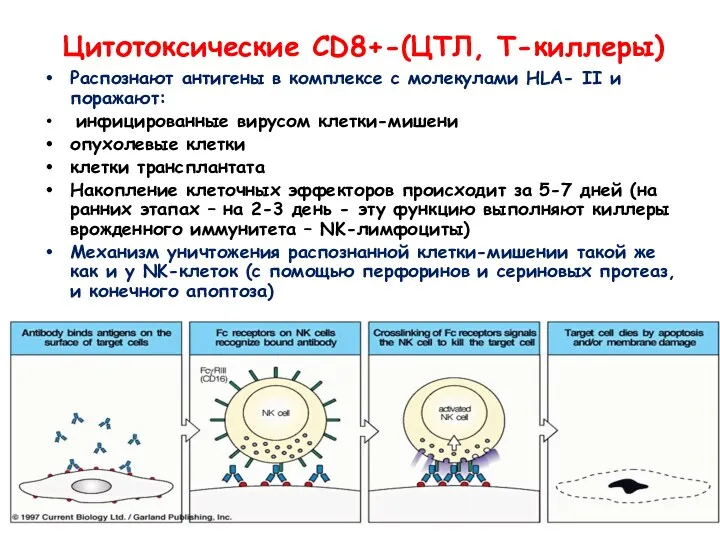
Цитотоксические CD8+-(ЦТЛ, Т-киллеры)
Распознают антигены в комплексе с молекулами HLA- II и
Цитотоксические CD8+-(ЦТЛ, Т-киллеры)
Распознают антигены в комплексе с молекулами HLA- II и
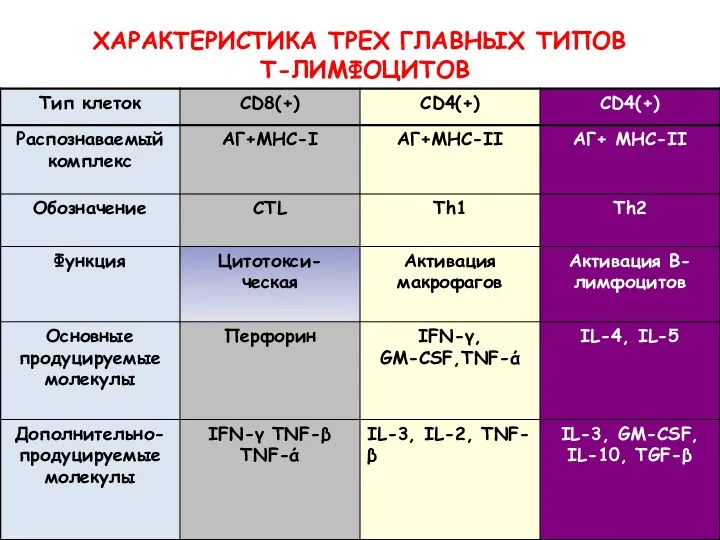
ХАРАКТЕРИСТИКА ТРЕХ ГЛАВНЫХ ТИПОВ
Т-ЛИМФОЦИТОВ
ХАРАКТЕРИСТИКА ТРЕХ ГЛАВНЫХ ТИПОВ
Т-ЛИМФОЦИТОВ
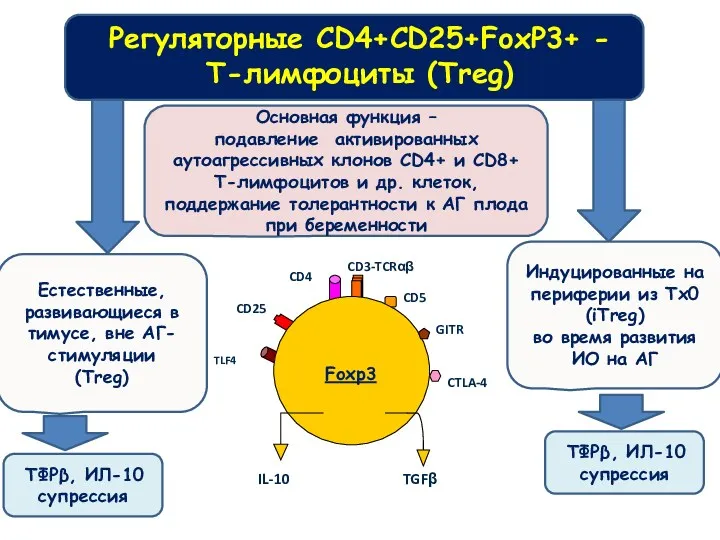
Регуляторные CD4+CD25+FoxP3+ -
T-лимфоциты (Treg)
Естественные, развивающиеся в тимусе, вне АГ-стимуляции
(Treg)
Индуцированные
Регуляторные CD4+CD25+FoxP3+ -
T-лимфоциты (Treg)
Естественные, развивающиеся в тимусе, вне АГ-стимуляции
(Treg)
Индуцированные

ЗАБОЛЕВАНИЯ, ПАТОГЕНЕТИЧЕСКИ СВЯЗАННЫЕ
С НАРУШЕНИЕМ ЕСТЕСТВЕННЫХ РЕГУЛЯТОРНЫХ
Т-КЛЕТОК
Содержание CD4+CD25+Foxp3+ Т-клеток часто
ЗАБОЛЕВАНИЯ, ПАТОГЕНЕТИЧЕСКИ СВЯЗАННЫЕ
С НАРУШЕНИЕМ ЕСТЕСТВЕННЫХ РЕГУЛЯТОРНЫХ
Т-КЛЕТОК
Содержание CD4+CD25+Foxp3+ Т-клеток часто
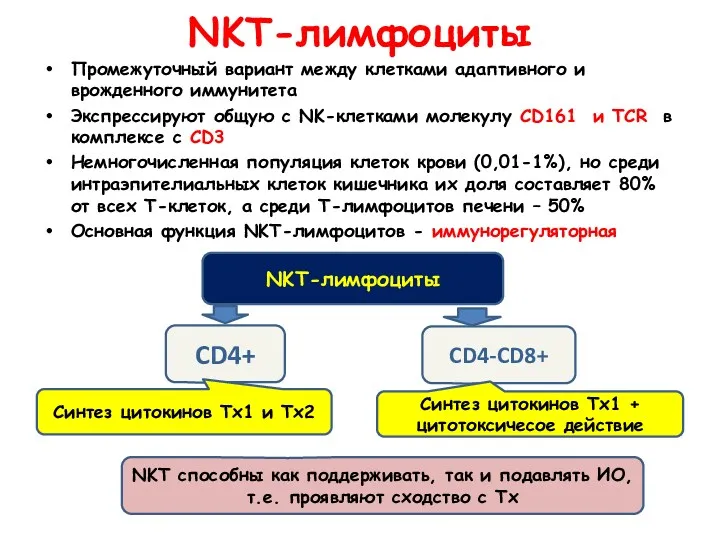
NKT-лимфоциты
Промежуточный вариант между клетками адаптивного и врожденного иммунитета
Экспрессируют общую с NK-клетками
NKT-лимфоциты
Промежуточный вариант между клетками адаптивного и врожденного иммунитета
Экспрессируют общую с NK-клетками
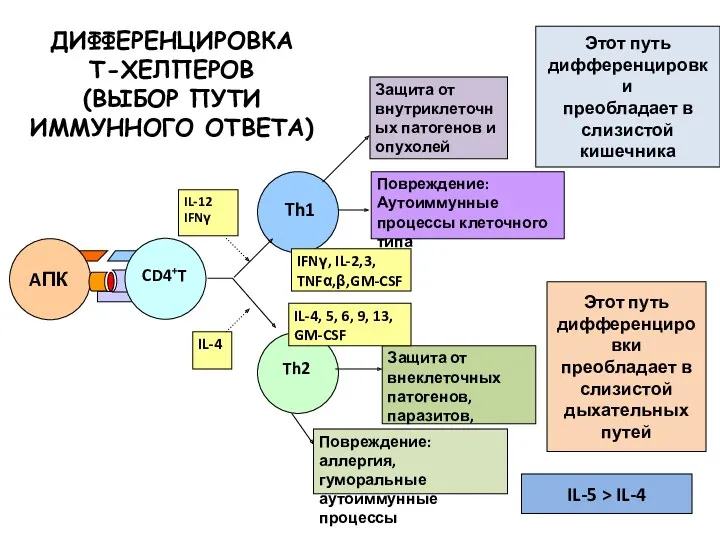
ДИФФЕРЕНЦИРОВКА
Т-ХЕЛПЕРОВ
(ВЫБОР ПУТИ
ИММУННОГО ОТВЕТА)
Этот путь
дифференцировки
преобладает в
слизистой кишечника
Этот
ДИФФЕРЕНЦИРОВКА
Т-ХЕЛПЕРОВ
(ВЫБОР ПУТИ
ИММУННОГО ОТВЕТА)
Этот путь
дифференцировки
преобладает в
слизистой кишечника
Этот

Клеточный иммунный ответ
(иммунный ответ по Тх-1 пути)
Участвуют популяции Тх-1 CD4+ и
Клеточный иммунный ответ
(иммунный ответ по Тх-1 пути)
Участвуют популяции Тх-1 CD4+ и
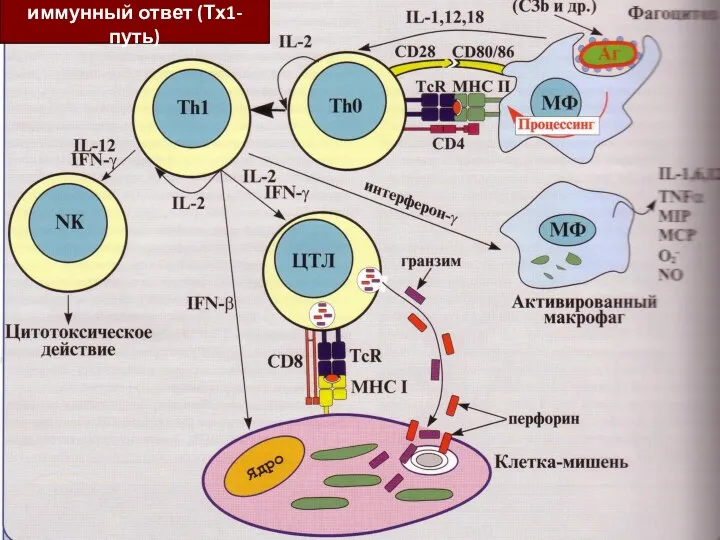
Клеточный
иммунный ответ (Тх1-путь)
Клеточный
иммунный ответ (Тх1-путь)

ГУМОРАЛЬНЫЙ ИММУННЫЙ ОТВЕТ (Тх1/Тх2-путь иммунного ответа)
Активация В-лимфоцитов (В2 CD5-) и их
ГУМОРАЛЬНЫЙ ИММУННЫЙ ОТВЕТ (Тх1/Тх2-путь иммунного ответа)
Активация В-лимфоцитов (В2 CD5-) и их
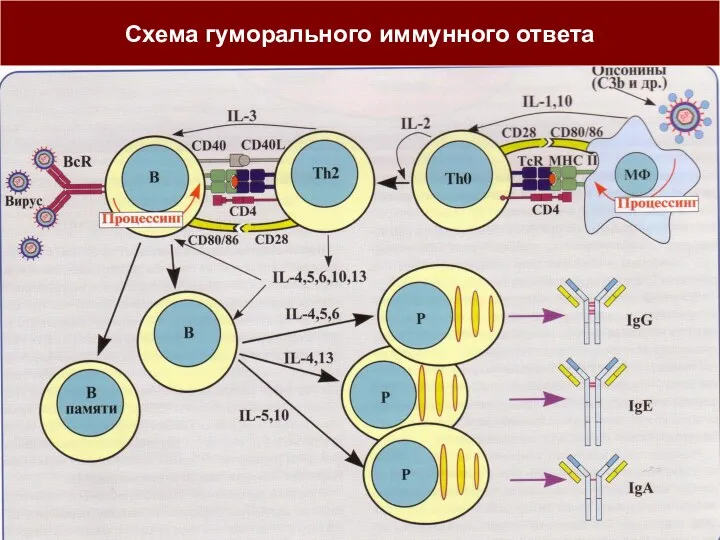
Схема гуморального иммунного ответа
Схема гуморального иммунного ответа

Противобактериальный иммунитет (иммунный ответ по Тх2-пути)
Направлен против бактерий и их токсинов
Бактерии
Противобактериальный иммунитет (иммунный ответ по Тх2-пути)
Направлен против бактерий и их токсинов
Бактерии
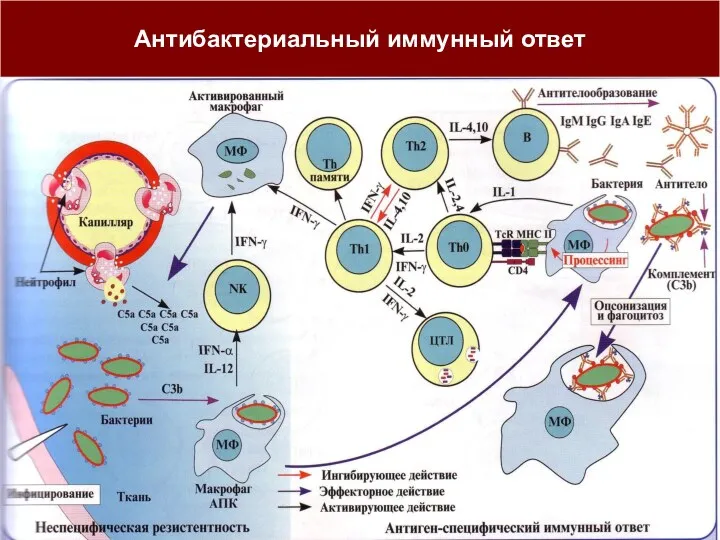
Антибактериальный иммунный ответ
Антибактериальный иммунный ответ
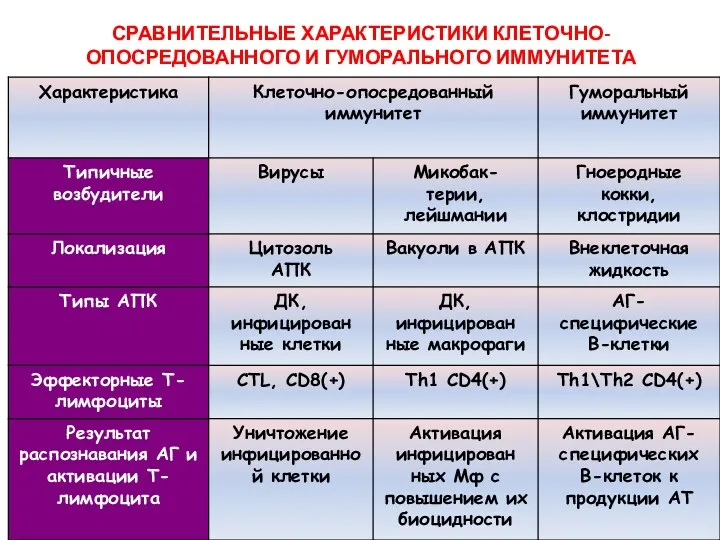
СРАВНИТЕЛЬНЫЕ ХАРАКТЕРИСТИКИ КЛЕТОЧНО-ОПОСРЕДОВАННОГО И ГУМОРАЛЬНОГО ИММУНИТЕТА
СРАВНИТЕЛЬНЫЕ ХАРАКТЕРИСТИКИ КЛЕТОЧНО-ОПОСРЕДОВАННОГО И ГУМОРАЛЬНОГО ИММУНИТЕТА
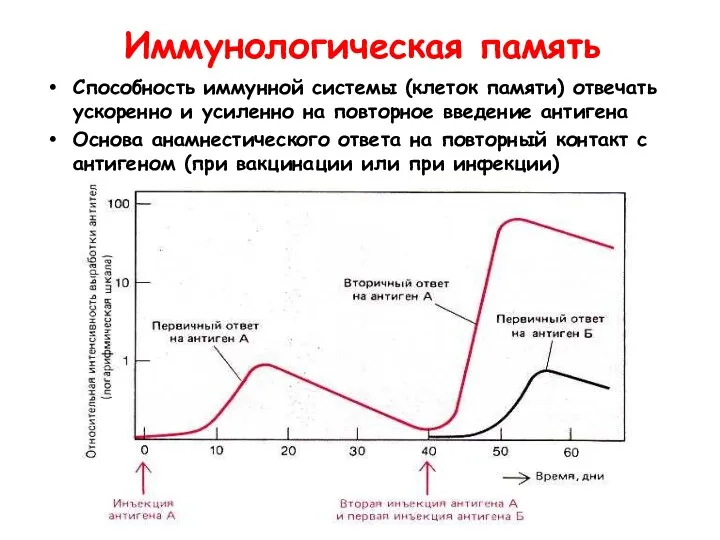
Иммунологическая память
Способность иммунной системы (клеток памяти) отвечать ускоренно и усиленно на
Иммунологическая память
Способность иммунной системы (клеток памяти) отвечать ускоренно и усиленно на

Т-клетки памяти могут формировать и при отсутствии контакта с чужеродными АГ:
Т-клетки памяти могут формировать и при отсутствии контакта с чужеродными АГ:

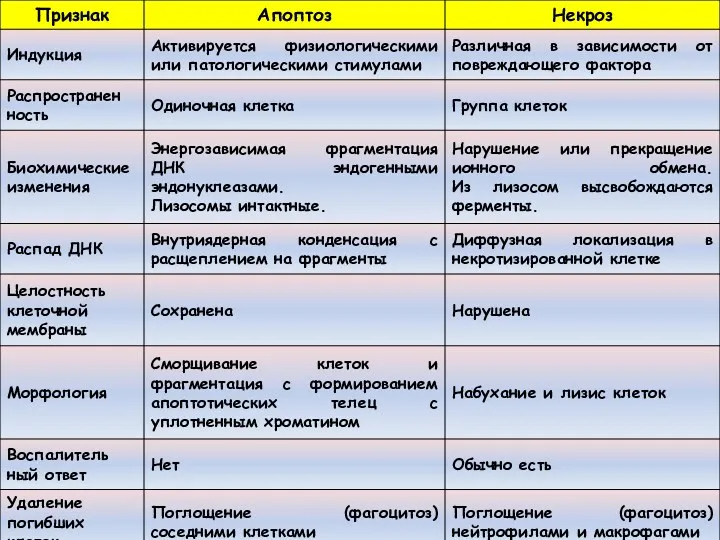
Формы клеточной гибели
Апоптоз
Аутофагия
Некроз
Митотическая катастрофа
Ороговение и др.
Классификация по данным Комитета по номенклатуре
Формы клеточной гибели
Апоптоз
Аутофагия
Некроз
Митотическая катастрофа
Ороговение и др.
Классификация по данным Комитета по номенклатуре

Апоптоз
Запрограммированная смерть клетки - процесс, посредством которого внутренние или внешние факторы,
Апоптоз
Запрограммированная смерть клетки - процесс, посредством которого внутренние или внешние факторы,
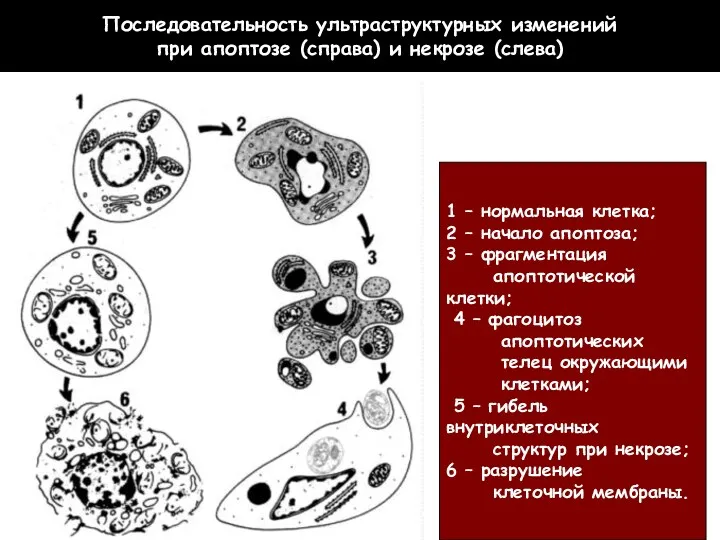
Последовательность ультраструктурных изменений
при апоптозе (справа) и некрозе (слева)
1 –
при апоптозе (справа) и некрозе (слева)
1 –
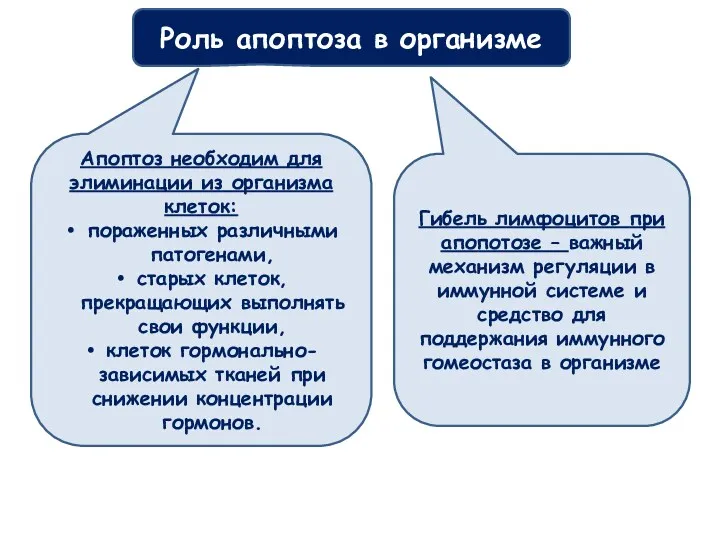
Апоптоз необходим для элиминации из организма клеток:
пораженных различными патогенами,
старых клеток,
Апоптоз необходим для элиминации из организма клеток:
пораженных различными патогенами,
старых клеток,
 Патологиялық анатомия
Патологиялық анатомия Оба қоздырғышы
Оба қоздырғышы Microscopic colitis
Microscopic colitis Сосудистые анастомозы РУС
Сосудистые анастомозы РУС Нарушение половых функций. Климакс
Нарушение половых функций. Климакс Мeningeal a syndrome in clinic of infectious diseases
Мeningeal a syndrome in clinic of infectious diseases Медсестринський процес – історичний нарис. Професійні організації медичних сестер
Медсестринський процес – історичний нарис. Професійні організації медичних сестер Оксигенотерапия. Виды оксигенотерапии
Оксигенотерапия. Виды оксигенотерапии Потребность в нормальном дыхании
Потребность в нормальном дыхании Cardiovascular system
Cardiovascular system Первая помощь детям при травмах и несчастных случаях. Общие советы для детей и взрослых
Первая помощь детям при травмах и несчастных случаях. Общие советы для детей и взрослых Медикаментозды остеопороз: алдын алудағы және түзетуге негізгі ұстанымдар
Медикаментозды остеопороз: алдын алудағы және түзетуге негізгі ұстанымдар Первая помощь при отморожении, общем охлаждении организма
Первая помощь при отморожении, общем охлаждении организма Обследование зубных рядов, зубов и периодонта в детском возрасте
Обследование зубных рядов, зубов и периодонта в детском возрасте Терапиялық стоматология клиникасындағы ауру сезімі және жансыздандыру
Терапиялық стоматология клиникасындағы ауру сезімі және жансыздандыру Общие вопросы лучевой диагностики. Ультразвуковое исследование
Общие вопросы лучевой диагностики. Ультразвуковое исследование Концепция обеспечения качества лекарственных средств
Концепция обеспечения качества лекарственных средств Клиническая симптоматология рака и абсцесса легких. Синдром полости в легком. (Тема 21)
Клиническая симптоматология рака и абсцесса легких. Синдром полости в легком. (Тема 21) Острая почечная и острая печеночная недостаточность
Острая почечная и острая печеночная недостаточность Ранний детский аутизм
Ранний детский аутизм Технические регламенты и национальные стандарты оказания оздоровительных услуг гостиничными организациями. (Лекция 6.1)
Технические регламенты и национальные стандарты оказания оздоровительных услуг гостиничными организациями. (Лекция 6.1) Уход за больными хирургического профиля
Уход за больными хирургического профиля Требования к организации и проведению профилактической дератизации и дезинсекции
Требования к организации и проведению профилактической дератизации и дезинсекции Периферические венозные катетеры
Периферические венозные катетеры Ортаңғы және ішкі құлақ аурулары. Саңыраулық және мылқаулық орта кұлактың жедел және созылмалы кабынуы мастоидит
Ортаңғы және ішкі құлақ аурулары. Саңыраулық және мылқаулық орта кұлактың жедел және созылмалы кабынуы мастоидит Нейропротезирование. История нейропротезирования
Нейропротезирование. История нейропротезирования Синдромы при заболеваниях дыхательной системы
Синдромы при заболеваниях дыхательной системы Первая медицинская помощь
Первая медицинская помощь