- Иммуногенетика. Открытие МНС

Содержание
- 2. План 1. Функции МНС 2.Характеристика МНС 3.Генетическая карта МНС 4.Наследование МНС 5.Строение классических МНС 6.МНС 1
- 3. Иммуногенетика – раздел иммунологии, изучающий генетические основы становления и развития иммунных реакций
- 4. МНС (Major Histocompatibility Complex) – главный комплекс гистосовместимости – система генов, кодирующих антигены, определяющих функционирование иммунной
- 5. Актуальность
- 6. Первый антиген гистосовместимости — антиген Мас — открыл в 1958 г. французский исследователь Ж. Доссе. Компьютерный
- 7. B этом же году впервые было сформулировано предположение об аллельном полиморфизме генов системы Hu-I. Исследования Ван
- 8. Открытие МНС. Нобелевская премия 1980 г. Жан Доссе Открыл первый антиген гистосовместимости человека (HLA) Барух Бенацерраф
- 9. ФУНКЦИИ МНС
- 10. Биологическая роль МНС Распознавание «свой – чужой» – реакция отторжения трансплантата, РТПХ (реакция трансплантат против хозяина)
- 11. ХАРАКТЕРИСТИКИ МНС
- 12. Гены комплекса MHC (в отличие от генов TCR и Ig) не подвергаются рекомбинации. Механизм их приспособления
- 13. Характеристики комплекса: ПОЛИМОРФИЗМ существование большого количества различных специфичностей HLA-генов в пределах каждого локуса. Гены отличаются между
- 14. Характеристики комплекса: ПОЛИГЕННОСТЬ наличие нескольких неаллельных близкосцепленных генов, белковые продукты которых сходны в структурном отношении и
- 15. Характеристики комплекса: ПОЛИГЕННОСТЬ и ПОЛИМОРФИЗМ Система HLA, включает гены 1 класса: А, В, С 2 класса:
- 16. ГЕНЕТИЧЕСКАЯ КАРТА МНС
- 17. Гены MHC делятся на три группы. Каждая группа включает гены, контролирующие синтез полипептидов одного из трех
- 18. MHC-I класс Гены групп HLA-A, HLA-B и HLA-C кодируют молекулы MHC класса I. MHC-II класс Гены
- 20. НАСЛЕДОВАНИЕ МНС
- 21. Экспрессия на мембране клетки Гены MHC кодоминантны, т.е. одновременно экспрессируются гены материнской и отцовской хромосом. Генов
- 22. СТРОЕНИЕ классических МНС
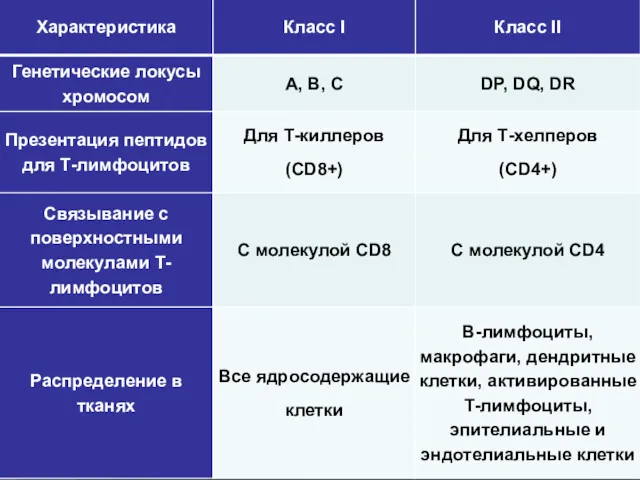
- 23. Класс I Класс II ЛОКУСЫ А, В, С ЛОКУСЫ DP, DQ, DR Классификация генов и их
- 24. МНС I класса
- 25. Молекула I класса состоит из 2-х цепей. Тяжелой α-цепи и легкой β2-микроглобулина
- 26. α-цепь, включает три фрагмента: внеклеточный, трансмембранный и цитоплазматический. Внеклеточный содержит 3 домена - α1, α2 и
- 27. Экзонная организация генов, кодирующих α-цепь молекул I класса 1 экзон, кодирующий сигнальный пептид, 4 экзона, кодирующие
- 28. Экспрессия и функции МНС 1 класса Экспрессия антигены представлены на всех клетках, тканях и органах, поэтому
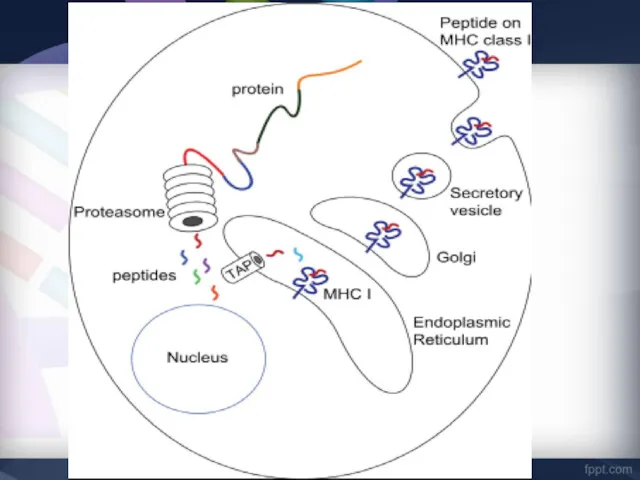
- 29. MHC-I «обслуживают» зону цитозоля, сообщающегося через ядерные поры с содержимым ядра. Здесь происходит фолдинг синтезированных белковых
- 30. Этапы подготовки вирусных белков к взаимодействию с молекулами I класса главного комплекса гистосовместимости.
- 31. I этап - разрушение вирусных белков, находящихся в цитозоле, с помощью протеазного комплекса - протеосомы.
- 32. II этап - транспорт образовавшихся пептидов во внутреннее пространство эндоплазматического ретикулума с помощью ТАР-1 и ТАР-2,
- 33. III этап - встреча транспортируемых пептидов с молекулами I класса МНС. Взаимодействие пептида с молекулой I
- 34. IV этап - комплекс через аппарат Гольджи транспортируется к клеточной поверхности, вирусный пептид в комплексе с
- 36. МНС II класса
- 37. Молекула II класса
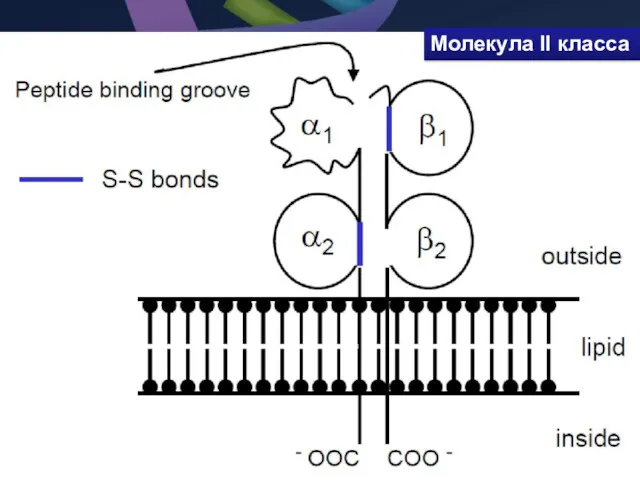
- 38. Молекула II класса гетеродимер из двух нековалентно связанных цепей α и β, каждая из которых включает
- 39. Экзонная организация генов, кодирующих α и β-цепи молекул II класса 1 экзон кодирует лидерную последовательность. 2
- 40. Экспрессия и функции МНС II класса Экспрессия антигены представлены на макрофагах, В-лимфоцитах и активированных Т-лимфоцитах. Функции
- 41. MHC-II. Зона «обслуживания» связана с внеклеточной средой и с клеточными органоидами (аппарат Гольджи, ЭПС, лизосомы, эндосомы
- 42. Этапы подготовки вирусных белков к взаимодействию с молекулами II класса главного комплекса гистосовместимости.
- 43. I этап - поглощение бактерий или их токсинов фагоцитирующей, способной к презентации антигена клеткой и разрушение
- 44. II этап - во внутреннем пространстве ЭПР происходит сборка молекул II класса, которые до встречи с
- 45. III этап - вакуоль, содержащая комплекс молекулы II класса с Ii, сливается с фаголизосомой. Протеазы разрушают
- 49. Заключение Из представленных данных литературы можно сделать вывод о том, что генетически детерминированные различия в силе
- 51. Скачать презентацию

План
1. Функции МНС
2.Характеристика МНС
3.Генетическая карта МНС
4.Наследование МНС
5.Строение классических МНС
6.МНС 1
План
1. Функции МНС
2.Характеристика МНС
3.Генетическая карта МНС
4.Наследование МНС
5.Строение классических МНС
6.МНС 1

Иммуногенетика – раздел иммунологии, изучающий генетические основы становления и развития иммунных
Иммуногенетика – раздел иммунологии, изучающий генетические основы становления и развития иммунных

МНС (Major Histocompatibility Complex) – главный комплекс гистосовместимости – система генов,
МНС (Major Histocompatibility Complex) – главный комплекс гистосовместимости – система генов,

Актуальность
Актуальность

Первый антиген гистосовместимости — антиген Мас — открыл в 1958 г.
Первый антиген гистосовместимости — антиген Мас — открыл в 1958 г.

B этом же году впервые было сформулировано предположение об аллельном полиморфизме
B этом же году впервые было сформулировано предположение об аллельном полиморфизме

Открытие МНС.
Нобелевская премия 1980 г.
Жан Доссе
Открыл первый антиген гистосовместимости человека
Открытие МНС.
Нобелевская премия 1980 г.
Жан Доссе
Открыл первый антиген гистосовместимости человека

ФУНКЦИИ МНС
ФУНКЦИИ МНС

Биологическая роль МНС
Распознавание «свой – чужой»
– реакция отторжения трансплантата, РТПХ
Биологическая роль МНС
Распознавание «свой – чужой»
– реакция отторжения трансплантата, РТПХ

ХАРАКТЕРИСТИКИ МНС
ХАРАКТЕРИСТИКИ МНС
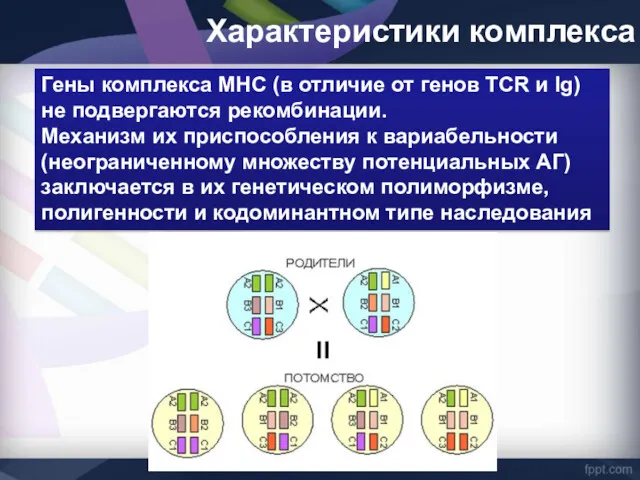
Гены комплекса MHC (в отличие от генов TCR и Ig) не
Гены комплекса MHC (в отличие от генов TCR и Ig) не

Характеристики комплекса: ПОЛИМОРФИЗМ
существование большого количества различных специфичностей HLA-генов в пределах каждого
Характеристики комплекса: ПОЛИМОРФИЗМ
существование большого количества различных специфичностей HLA-генов в пределах каждого

Характеристики комплекса: ПОЛИГЕННОСТЬ
наличие нескольких неаллельных близкосцепленных генов, белковые продукты которых сходны
Характеристики комплекса: ПОЛИГЕННОСТЬ
наличие нескольких неаллельных близкосцепленных генов, белковые продукты которых сходны
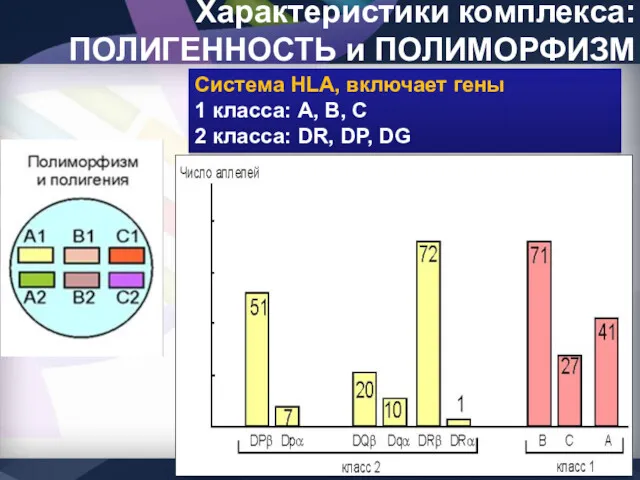
Характеристики комплекса: ПОЛИГЕННОСТЬ и ПОЛИМОРФИЗМ
Система HLA, включает гены
1 класса: А, В,
Характеристики комплекса: ПОЛИГЕННОСТЬ и ПОЛИМОРФИЗМ
Система HLA, включает гены
1 класса: А, В,

ГЕНЕТИЧЕСКАЯ КАРТА МНС
ГЕНЕТИЧЕСКАЯ КАРТА МНС
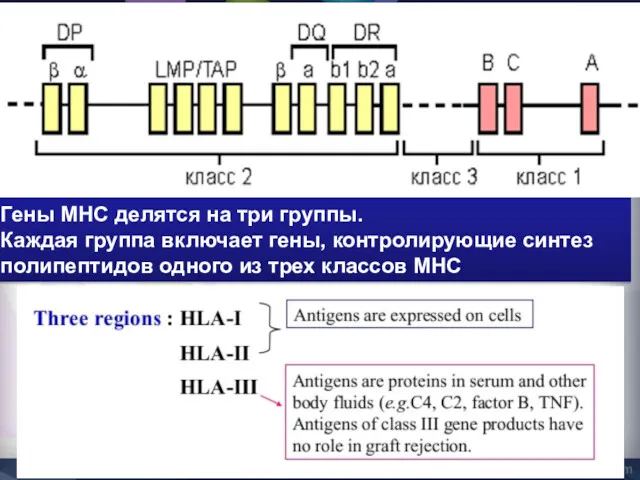
Гены MHC делятся на три группы.
Каждая группа включает гены, контролирующие
Гены MHC делятся на три группы.
Каждая группа включает гены, контролирующие

MHC-I класс
Гены групп HLA-A, HLA-B и HLA-C кодируют молекулы MHC
MHC-I класс
Гены групп HLA-A, HLA-B и HLA-C кодируют молекулы MHC
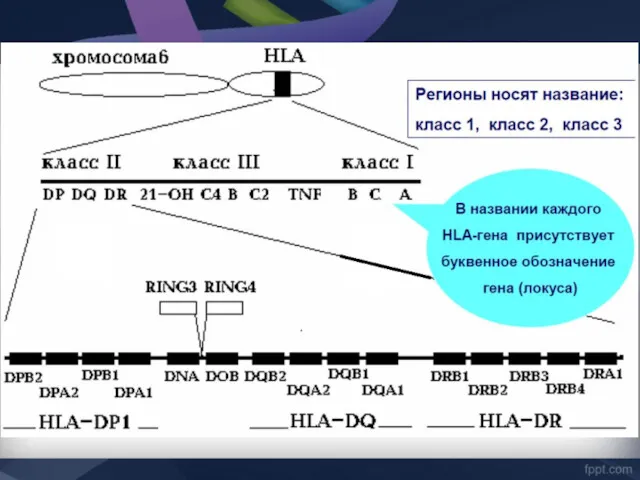

НАСЛЕДОВАНИЕ МНС
НАСЛЕДОВАНИЕ МНС

Экспрессия на мембране клетки
Гены MHC кодоминантны, т.е. одновременно экспрессируются гены материнской
Экспрессия на мембране клетки
Гены MHC кодоминантны, т.е. одновременно экспрессируются гены материнской

СТРОЕНИЕ
классических МНС
СТРОЕНИЕ
классических МНС
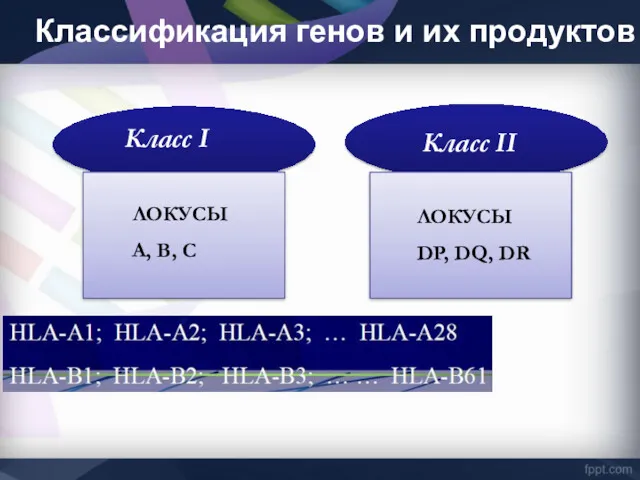
Класс I
Класс II
ЛОКУСЫ
А, В, С
ЛОКУСЫ
DP, DQ, DR
Классификация генов и их продуктов
Класс I
Класс II
ЛОКУСЫ
А, В, С
ЛОКУСЫ
DP, DQ, DR
Классификация генов и их продуктов

МНС I класса
МНС I класса
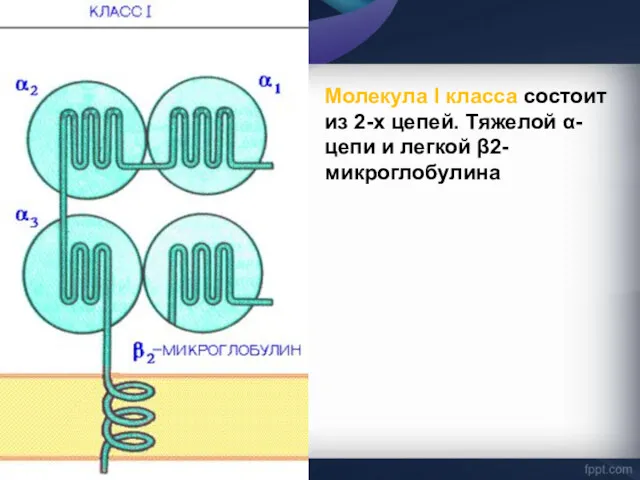
Молекула I класса состоит из 2-х цепей. Тяжелой α-цепи и легкой
Молекула I класса состоит из 2-х цепей. Тяжелой α-цепи и легкой

α-цепь, включает три фрагмента: внеклеточный, трансмембранный и цитоплазматический.
Внеклеточный содержит 3 домена
α-цепь, включает три фрагмента: внеклеточный, трансмембранный и цитоплазматический.
Внеклеточный содержит 3 домена
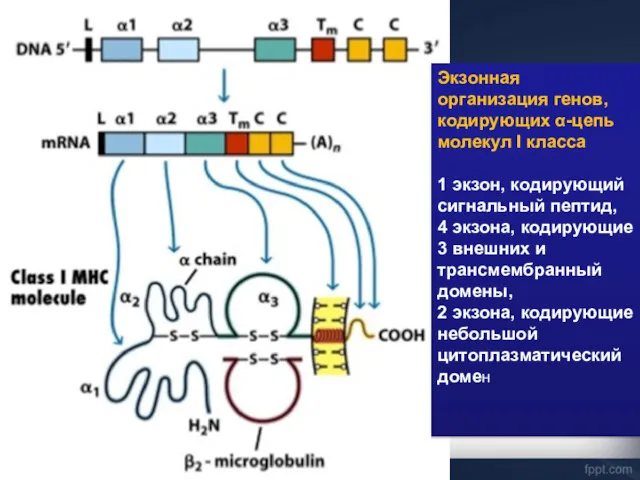
Экзонная организация генов, кодирующих α-цепь молекул I класса
1 экзон, кодирующий сигнальный
Экзонная организация генов, кодирующих α-цепь молекул I класса
1 экзон, кодирующий сигнальный
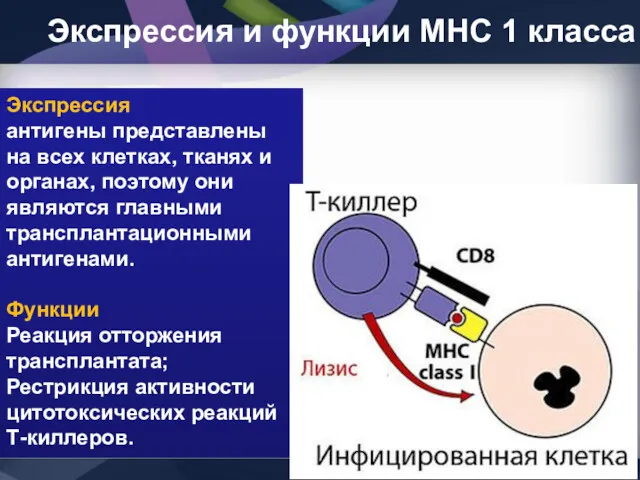
Экспрессия и функции МНС 1 класса
Экспрессия
антигены представлены на всех клетках,
Экспрессия и функции МНС 1 класса
Экспрессия
антигены представлены на всех клетках,

MHC-I «обслуживают» зону цитозоля, сообщающегося через ядерные поры с содержимым ядра.
MHC-I «обслуживают» зону цитозоля, сообщающегося через ядерные поры с содержимым ядра.
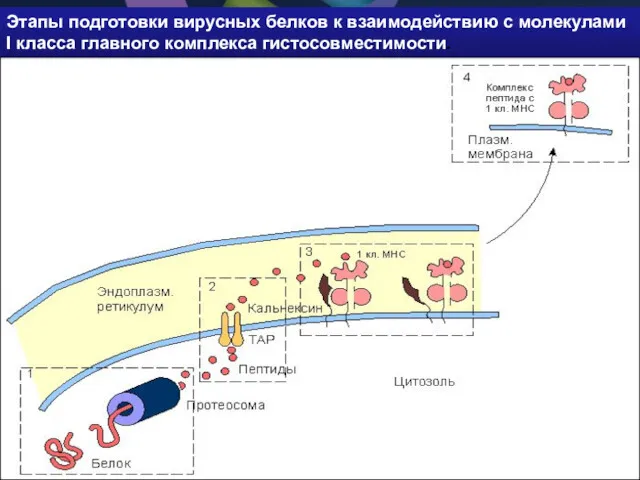
Этапы подготовки вирусных белков к взаимодействию с молекулами I класса главного
Этапы подготовки вирусных белков к взаимодействию с молекулами I класса главного
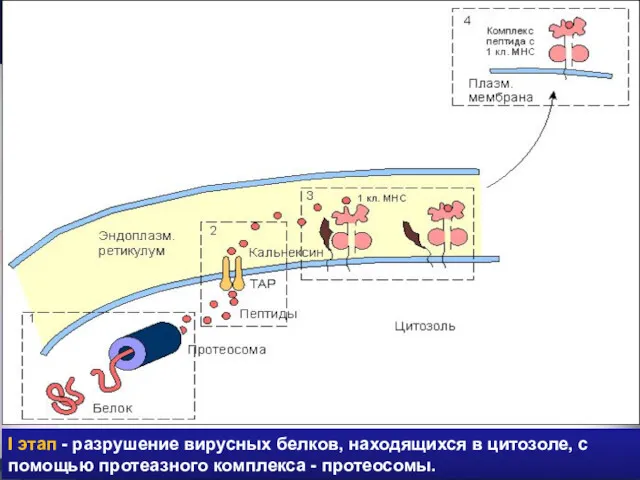
I этап - разрушение вирусных белков, находящихся в цитозоле, с помощью
I этап - разрушение вирусных белков, находящихся в цитозоле, с помощью
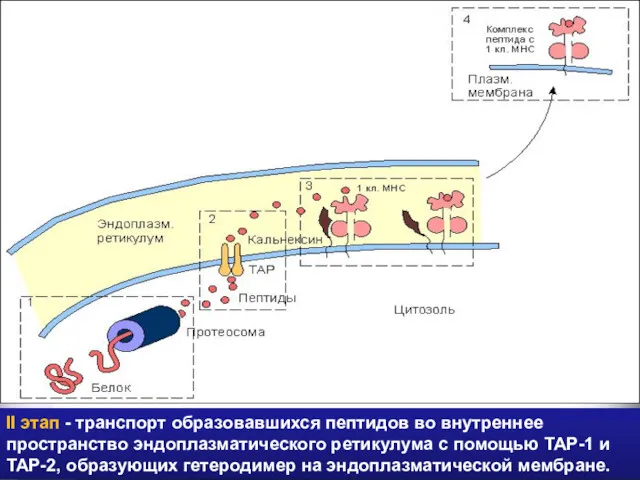
II этап - транспорт образовавшихся пептидов во внутреннее пространство эндоплазматического ретикулума
II этап - транспорт образовавшихся пептидов во внутреннее пространство эндоплазматического ретикулума
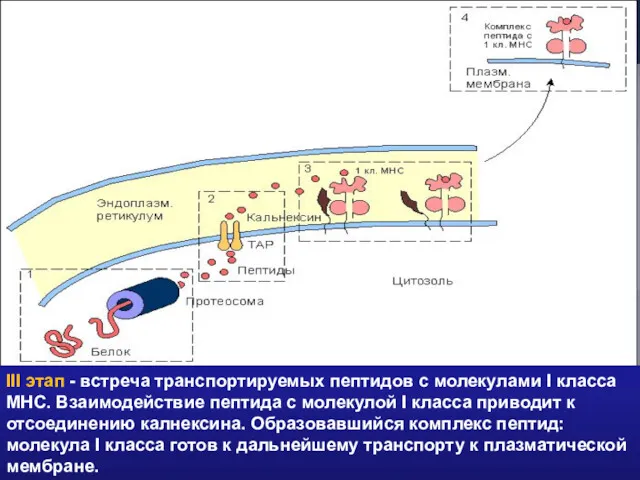
III этап - встреча транспортируемых пептидов с молекулами I класса МНС.
III этап - встреча транспортируемых пептидов с молекулами I класса МНС.
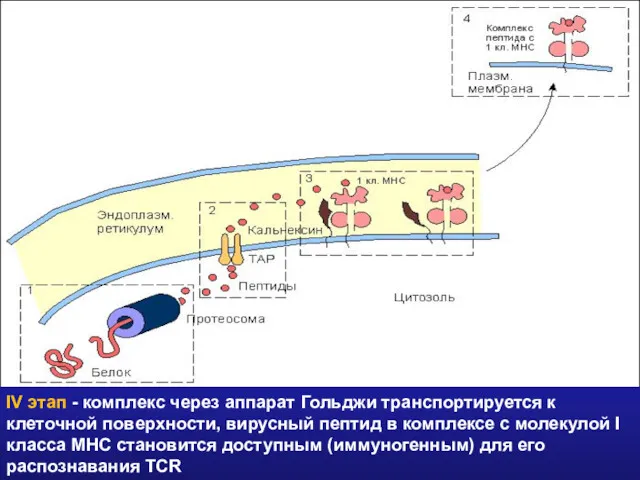
IV этап - комплекс через аппарат Гольджи транспортируется к клеточной поверхности,
IV этап - комплекс через аппарат Гольджи транспортируется к клеточной поверхности,

МНС II класса
МНС II класса
Молекула II класса
Молекула II класса
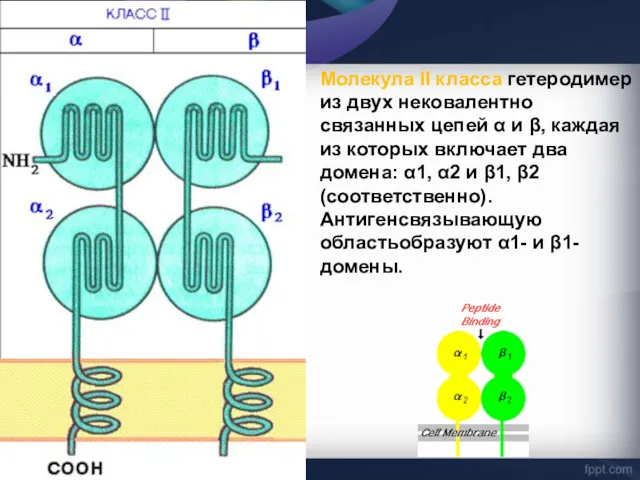
Молекула II класса гетеродимер из двух нековалентно связанных цепей α и
Молекула II класса гетеродимер из двух нековалентно связанных цепей α и
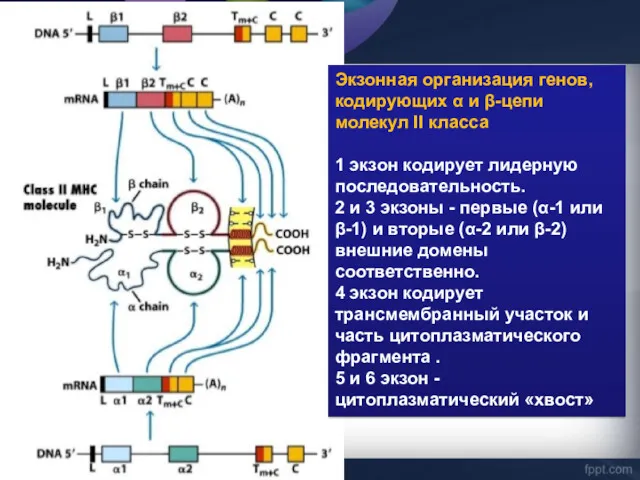
Экзонная организация генов, кодирующих α и β-цепи молекул II класса
1 экзон
Экзонная организация генов, кодирующих α и β-цепи молекул II класса
1 экзон
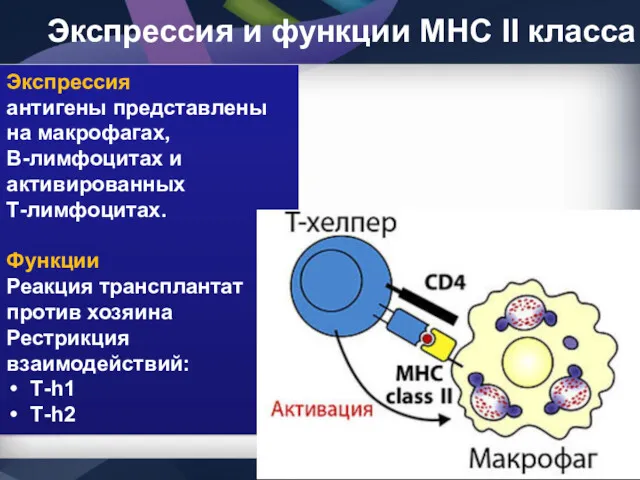
Экспрессия и функции МНС II класса
Экспрессия
антигены представлены на макрофагах, В-лимфоцитах
Экспрессия и функции МНС II класса
Экспрессия
антигены представлены на макрофагах, В-лимфоцитах

MHC-II. Зона «обслуживания» связана с внеклеточной средой и с клеточными органоидами
MHC-II. Зона «обслуживания» связана с внеклеточной средой и с клеточными органоидами
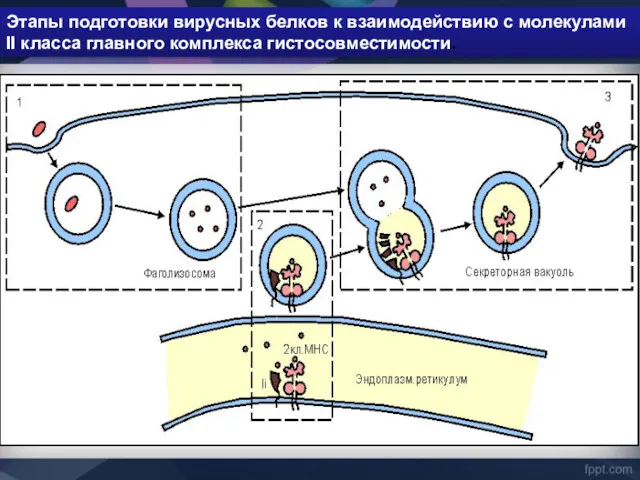
Этапы подготовки вирусных белков к взаимодействию с молекулами II класса главного
Этапы подготовки вирусных белков к взаимодействию с молекулами II класса главного
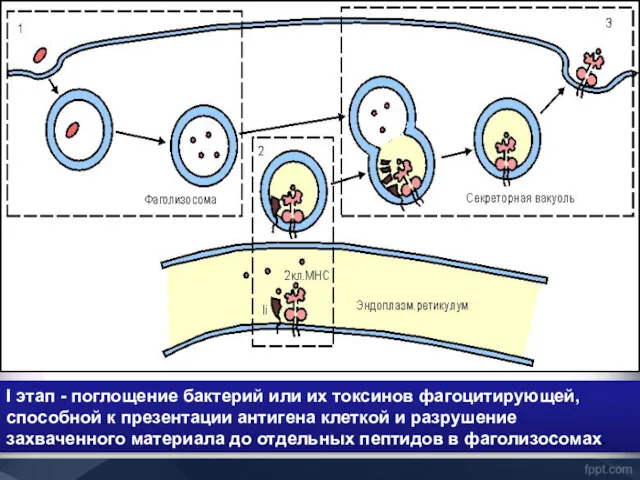
I этап - поглощение бактерий или их токсинов фагоцитирующей, способной к
I этап - поглощение бактерий или их токсинов фагоцитирующей, способной к
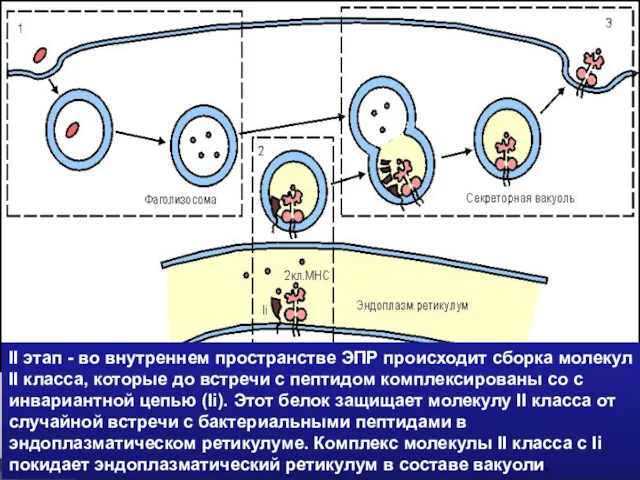
II этап - во внутреннем пространстве ЭПР происходит сборка молекул II
II этап - во внутреннем пространстве ЭПР происходит сборка молекул II
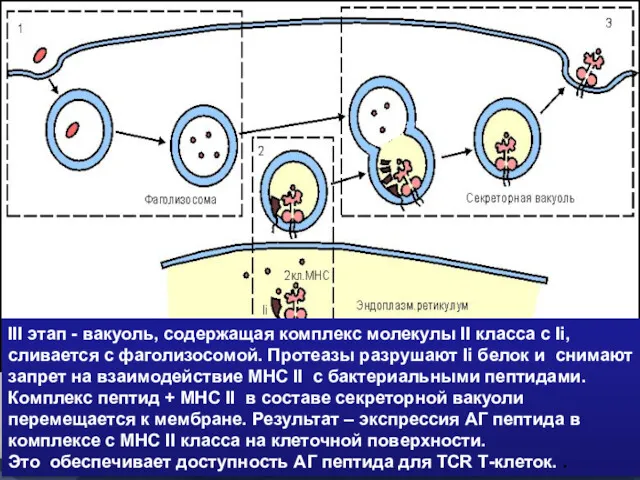
III этап - вакуоль, содержащая комплекс молекулы II класса с Ii,
III этап - вакуоль, содержащая комплекс молекулы II класса с Ii,
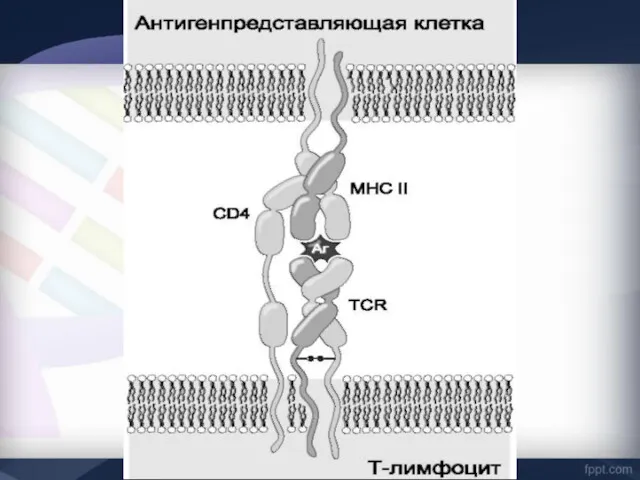
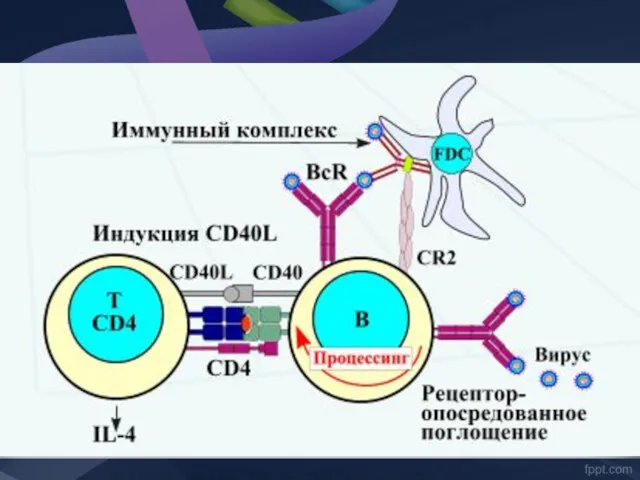

Заключение
Из представленных данных литературы можно сделать вывод о том, что генетически
Заключение
Из представленных данных литературы можно сделать вывод о том, что генетически
 Стоматологиядағы кірісулер кезіндегі шұғыл көмек
Стоматологиядағы кірісулер кезіндегі шұғыл көмек Первая помощь при инсульте
Первая помощь при инсульте Первая помощь при повреждениях головы и позвоночника
Первая помощь при повреждениях головы и позвоночника Ауыл тұрғындарына ,бастапқы медико – санитарлық көмекті ұйымдастыру жане шешу жолдары
Ауыл тұрғындарына ,бастапқы медико – санитарлық көмекті ұйымдастыру жане шешу жолдары Су-Джок терапия
Су-Джок терапия Первая доврачебная помощь при ДТП
Первая доврачебная помощь при ДТП Инфекционная безопасность в учреждениях здравоохранения
Инфекционная безопасность в учреждениях здравоохранения Первая медицинская помощь при поражении отравляющими веществами
Первая медицинская помощь при поражении отравляющими веществами Современный взгляд на терапию гиперфосфатемии при ХБП. Гиперкалиемия, профилактика и коррекция
Современный взгляд на терапию гиперфосфатемии при ХБП. Гиперкалиемия, профилактика и коррекция Жасөспірім шақтағы семіздіктің алдын алу
Жасөспірім шақтағы семіздіктің алдын алу Конкурентные преимущества биологически активных комплексов Арт Лайф
Конкурентные преимущества биологически активных комплексов Арт Лайф Черепно-мозговая травма
Черепно-мозговая травма Экстрагенитальная патология и беременность (заболевания почек, эндокринных органов, острые хирургические заболевания, инфекции)
Экстрагенитальная патология и беременность (заболевания почек, эндокринных органов, острые хирургические заболевания, инфекции) Аномалии конституции у детей. Диатезы
Аномалии конституции у детей. Диатезы Жапырақтары дәрілік өсімдмік шикізатын дайындау ерекшеліктері. Жинау, кептіру, сақтау ерекшеліктері
Жапырақтары дәрілік өсімдмік шикізатын дайындау ерекшеліктері. Жинау, кептіру, сақтау ерекшеліктері Микобактерия туберкулеза. Микробиология
Микобактерия туберкулеза. Микробиология Болезнь Верльгофа (хроническая иммунопатологическая тромбоцитопеническая пурпура)
Болезнь Верльгофа (хроническая иммунопатологическая тромбоцитопеническая пурпура) Сечовидільна система
Сечовидільна система Дәрілік өсімдіктерді халық медицинасында қолдану тарихы
Дәрілік өсімдіктерді халық медицинасында қолдану тарихы Аллергиялық реакциялар
Аллергиялық реакциялар Буллезные дерматозы у детей
Буллезные дерматозы у детей Көпұлтты Қазақстан жағдайында тұлғаны
Көпұлтты Қазақстан жағдайында тұлғаны Нейроофтальмологические расстройства при ВИЧ-инфекции
Нейроофтальмологические расстройства при ВИЧ-инфекции Препарат Диспорт. Комплекс из ботулинического токсина типа А и другого протеина, нетоксичного и неактивного
Препарат Диспорт. Комплекс из ботулинического токсина типа А и другого протеина, нетоксичного и неактивного Joint Commission International. Стандарты JCI
Joint Commission International. Стандарты JCI Методологические предпосылки и основные понятия теории системной динамической локализации высших психических функций
Методологические предпосылки и основные понятия теории системной динамической локализации высших психических функций Философия сестринского дела
Философия сестринского дела Составление индивидуального календаря прививок с фоновыми заболеваниями
Составление индивидуального календаря прививок с фоновыми заболеваниями