- Механизмы действия сигнальных соединений. Ядерные рецепторы

Содержание
- 2. Этапы инициации эффектов сигнальных соединений Селективная генерация межклеточного сигнала (эндокринная клетка/железа) Клеточное узнавание сигнала (рецепторы) Внутриклеточная
- 3. Классификация гормонкомпетентных тканей Ткани Мишени (рецепторы есть) Немишени (рецепторов нет) Гормонзависимые Гормончувствительные Селективные эффекты Общетрофические эффекты+селективные
- 4. Ядерные рецепторы Тирозинкиназы Серин/треонинкиназы Гуанилатциклазы Транскрипционные факторы Фосфатазы Ферменты Сопряженные с тирозинкиназами Сопряженные с G-белками Сопряженные
- 5. Модель структурно-функциональной организации рецептора Гормонсвязывающий домен Сопрягающий домен Эффекторный домен
- 6. Уни- и мультирецепторные модели действия рецепторов Рецептор 1 Рецептор 1 Рецептор 2 Рецептор 3 Эффекты 1
- 7. R2 R1 H1 H2 Эффектор A Эффектор B Эффектор C Эффектор D Сигнальные соединения могут усиливать
- 8. kass H + R ↔ HR; kass/kdiss = Ka = 1/Kd kdiss R R R R
- 9. Концепции индуцированного лигандом конформационного перехода рецепторов А: Связывание лиганда смещает равновесие между активной (R*) и неактивной
- 10. H ½ HRE ½ HRE H H R R ДНК А Б Коактиватор R R R
- 11. Эффекты Десенситизация R R’ H Индуцируемые лигандом конформационные изменения рецептора обеспечивают параллельно проведение сигнала и десенситизацию
- 12. Ядерные рецепторы и системы передачи сигнала
- 13. Специфические факторы транскрипции Конститутивные факторы транскрипции = факторы транскрипции базального транскрипционого комплекса: Обеспечение инициации транскрипции Обеспечение
- 14. Способ передачи: на входе: Взаимодействие с гормоном Взаимодействие с другим сигнальным соединением Фосфорилирование На выходе: Изменение
- 15. Корегуляторы ядерных рецепторов: 1. Корепрессоры Помогают подавлять действие рецептора без лиганда Связывание лиганда ведет к диссоциации
- 16. Регуляция экспрессии гормонзависимого гена млекопитающих Корегулятор мРНК Активация транскрипции ЯР ЯР ГЧЭ Базальный транскрип-ционный комплекс Гормон
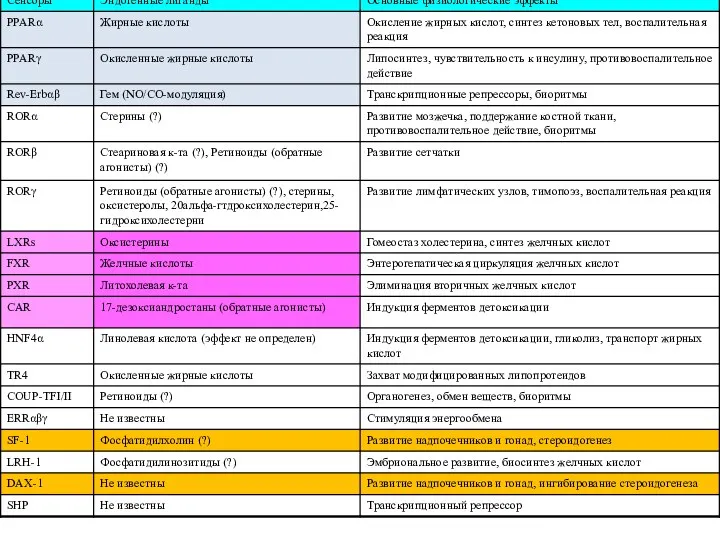
- 17. Лиганды ядерных рецепторов Гормоны (стероидные, тиреоидоные) Гормональные формы жирорастворимых витаминов (Д и А) Сиротские: Сенсоры липидов
- 18. Производные холестерина Гем
- 19. Сродство к лиганду Соединения с исключительно сигнальной функцией (гормоны) Обычные метаболиты Минорные метаболиты Рецепторы Сенсоры Рецепторы
- 21. Активность ядерных рецепторов в отсутствие лиганда Транскрипционно неактивные без лиганда : некоторые рецепторы стероидов 2. Оказывающие
- 22. Снятие репрессорного действия на ген белка Hairless при инактивирующей мутации рецептора гормональной формы витамина D Репрессорное
- 23. Инактивирующая мутация рецептора гормональной формы витамина Д
- 24. N Связывание ДНК Связывание лиганда и коактиватора CoR AF-2 CTE Zn2+ AF-1 F E D C
- 25. Домен NTD (A/B) (модуляторный): Наиболее вариабельный по длине и послед-ти а/к Сайты фосфорилирования MAP-киназами и другими
- 26. Структура ДНК-связывающего домена ядерных рецепторов Контакт с сахаро-фосфатным остовом ДНК
- 27. Гормончувствительные элементы (ГЧЭ) ДНК Особенности взаимодействия ГЧЭ с ядерными рецепторами (ЯР): Полусайты –связывание мономера ЯР Сдвоенные
- 28. Функции C-концевого расширения ДНК-связывающего домена (СТЕ) во взаимодействии ядерных рецепторов с гормончувствительными элементами: 1. Повышение прочности
- 29. Размер спейсера между полусайтами гормончувствительных элементов дискриминирует взаимодействие с ДНК димерных ядерных рецепторов. Размер спейсера узнается
- 30. Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их локализации относительно старта транскрипции регулируемых гормоном генов
- 31. Шарнирный домен D Вариабелен по размеру и последовательности а/к Гибкий (поворот на 180° по отношению к
- 32. HRE NTD DBD D LBD ДНК Шарнирный домен D обеспечивает поворот лигандсвязывающего домена относительно ДНК-связывающего домена
- 33. Умеренно консервативен Лигандсвязывающий карман: Связывание лиганда Участие в димеризации Взаимодействие с белками теплового шока Сигнал ядерной
- 34. Работа гормонзависимой активаторной функции AF2 AF2 AF2 Лигандсвязывающий карман Гормон Появление сайта взаимодействия с коактиватором AF2
- 35. Связывание лиганда-агониста (H) меняет ориентацию α-спирали H12, что препятствует взаимодействию ядерного рецептора (NR) с корепрессором (CoR)
- 36. Роль димеризации
- 37. Два домена взаимодействия с ядерными рецепторами (ID-N и ID-C) корепрессора NCoR обеспечивают его связывание одновременно с
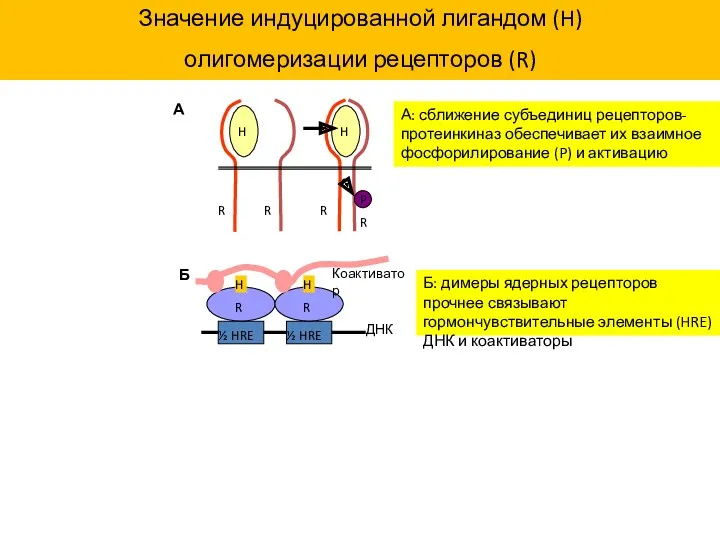
- 38. ½ HRE ½ HRE H H R R ДНК Значение индуцированной лигандом (H) олигомеризации рецепторов (R):
- 39. Роль N/C взаимодействия и димеризации в активации рецептора N/C взаимодействие может быть внутри- и межмолекулярным
- 40. Партнеры по димеризации В зависимости от партнера по димеризации чаще узнаются разные типы ГЧЭ Способствуют удержанию
- 41. Связывание ДНК Гомо-димеризация Связывание шаперонов Связывание корегуляторов H Связывание лиганда Лиганд влияет на взаимодействия рецептора с
- 42. Роль комплексирования рецепторов стероидов с белками теплового шока (hsp90) В цитоплазме гетеродимеры: 1 молекула ЯР+2 молекулы
- 43. гормон ядро цитоплазма А NR NR гормон Стероидные гормоны индуцируют транслокацию своих рецепторов (SR) из цитоплазмы
- 44. В отсутствие лиганда-агониста ядерные рецепторы (NR) рекрутируют многокомпонентный комплекс, включающий первичные (NCoR) и вторичные (Sin3B) корепрессоры
- 45. Рекрутируемые ядерными рецепторами (NR) в присутствии лиганда-агониста (черные треугольники) многокомпонентные комплексы коактиваторов (SRC), коинтеграторов (CBP) и
- 46. Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их локализации относительно старта транскрипции регулируемых гормоном генов Роль
- 47. Позиционирование нуклеосомы может усиливать стимулирующее действие ядерных рецепторов на транскрипцию (А) и Придавать тканеспецифичность эффектам гормонов
- 49. Скачать презентацию
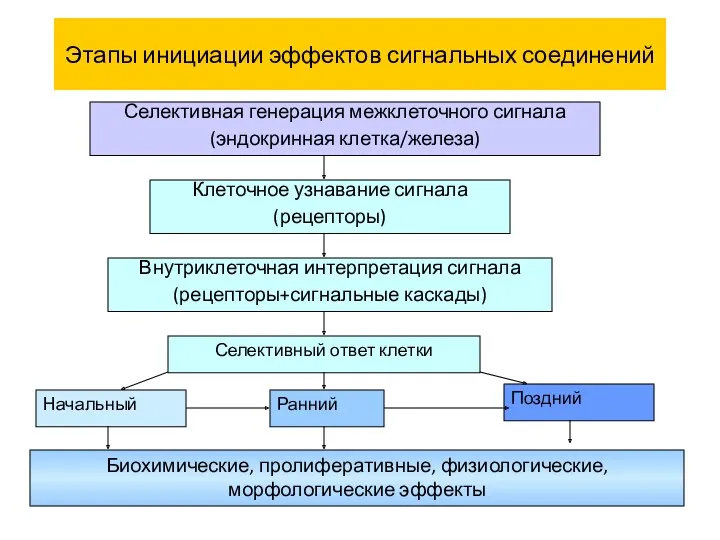
Этапы инициации эффектов сигнальных соединений
Селективная генерация межклеточного сигнала
(эндокринная клетка/железа)
Клеточное узнавание
Этапы инициации эффектов сигнальных соединений
Селективная генерация межклеточного сигнала
(эндокринная клетка/железа)
Клеточное узнавание
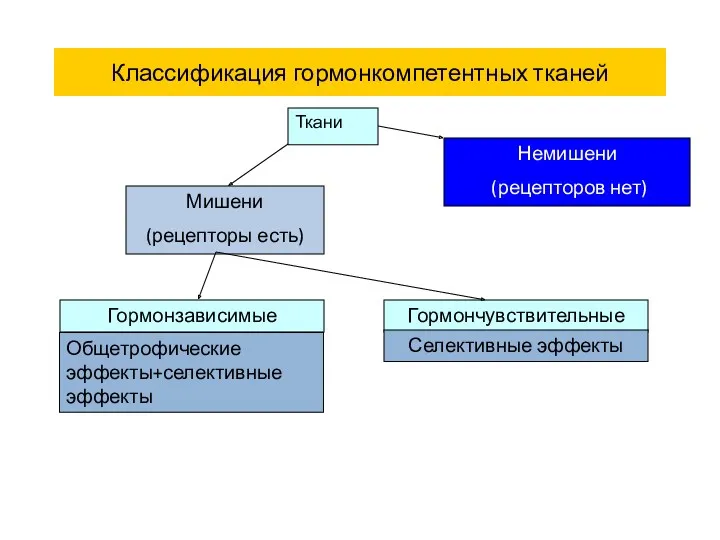
Классификация гормонкомпетентных тканей
Ткани
Мишени
(рецепторы есть)
Немишени
(рецепторов нет)
Гормонзависимые
Гормончувствительные
Селективные эффекты
Общетрофические эффекты+селективные эффекты
Классификация гормонкомпетентных тканей
Ткани
Мишени
(рецепторы есть)
Немишени
(рецепторов нет)
Гормонзависимые
Гормончувствительные
Селективные эффекты
Общетрофические эффекты+селективные эффекты
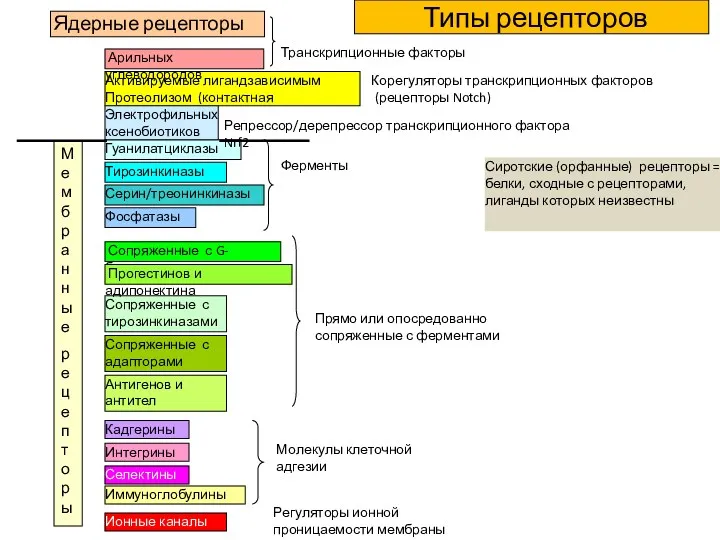
Ядерные рецепторы
Тирозинкиназы
Серин/треонинкиназы
Гуанилатциклазы
Транскрипционные факторы
Фосфатазы
Ферменты
Сопряженные с тирозинкиназами
Сопряженные с G-белками
Сопряженные с адапторами
Прямо
Ядерные рецепторы
Тирозинкиназы
Серин/треонинкиназы
Гуанилатциклазы
Транскрипционные факторы
Фосфатазы
Ферменты
Сопряженные с тирозинкиназами
Сопряженные с G-белками
Сопряженные с адапторами
Прямо
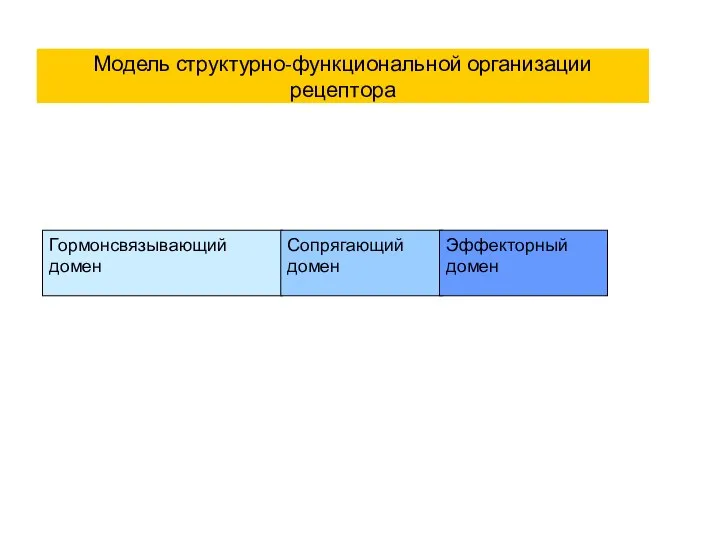
Модель структурно-функциональной организации рецептора
Гормонсвязывающий домен
Сопрягающий домен
Эффекторный домен
Модель структурно-функциональной организации рецептора
Гормонсвязывающий домен
Сопрягающий домен
Эффекторный домен
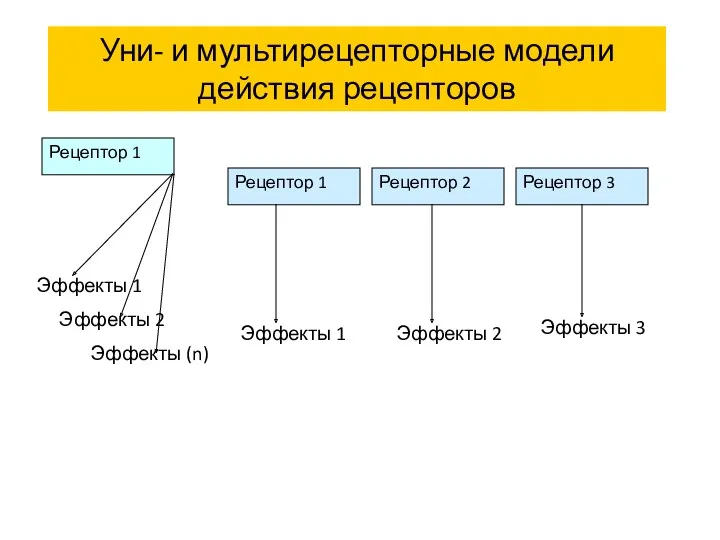
Уни- и мультирецепторные модели действия рецепторов
Рецептор 1
Рецептор 1
Рецептор 2
Рецептор 3
Эффекты 1
Уни- и мультирецепторные модели действия рецепторов
Рецептор 1
Рецептор 1
Рецептор 2
Рецептор 3
Эффекты 1
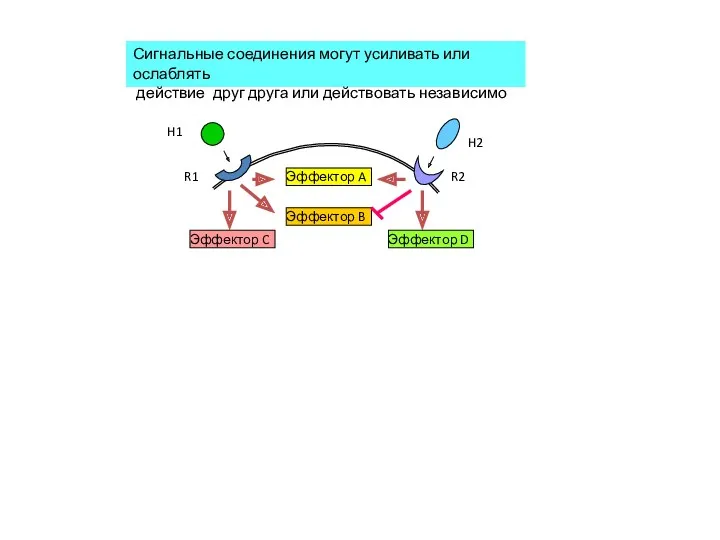
R2
R1
H1
H2
Эффектор A
Эффектор B
Эффектор C
Эффектор D
Сигнальные соединения могут усиливать или ослаблять
действие
R2
R1
H1
H2
Эффектор A
Эффектор B
Эффектор C
Эффектор D
Сигнальные соединения могут усиливать или ослаблять
действие
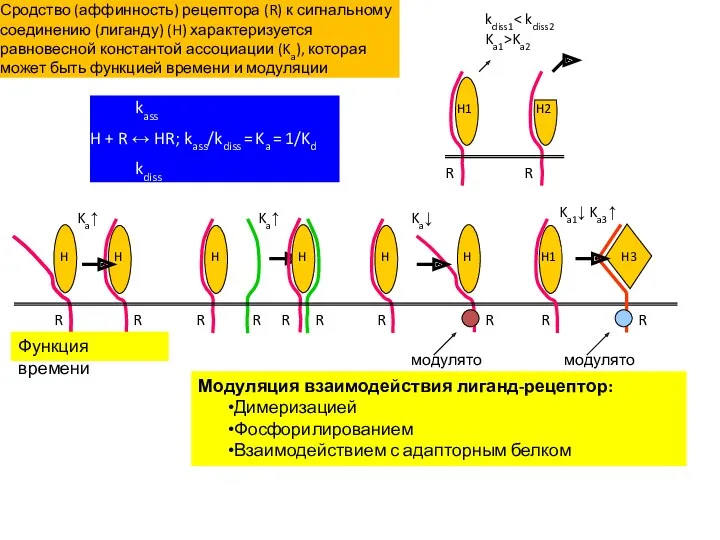
kass
H + R ↔ HR; kass/kdiss = Ka = 1/Kd
kass
H + R ↔ HR; kass/kdiss = Ka = 1/Kd
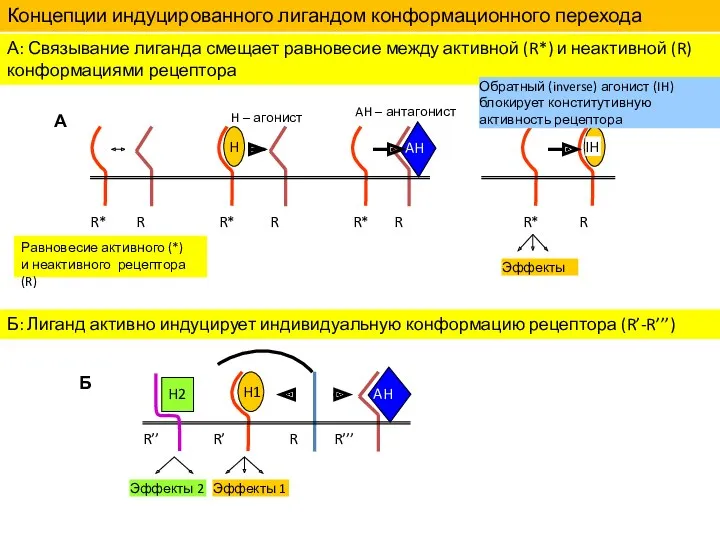
Концепции индуцированного лигандом конформационного перехода рецепторов
А: Связывание лиганда смещает равновесие между
Концепции индуцированного лигандом конформационного перехода рецепторов
А: Связывание лиганда смещает равновесие между
H
½ HRE
½ HRE
H
H
R
R
ДНК
А
Б
Коактиватор
R
R
R
R
Значение индуцированной лигандом (H)
олигомеризации рецепторов (R)
А: сближение субъединиц
H
½ HRE
½ HRE
H
H
R
R
ДНК
А
Б
Коактиватор
R
R
R
R
Значение индуцированной лигандом (H)
олигомеризации рецепторов (R)
А: сближение субъединиц
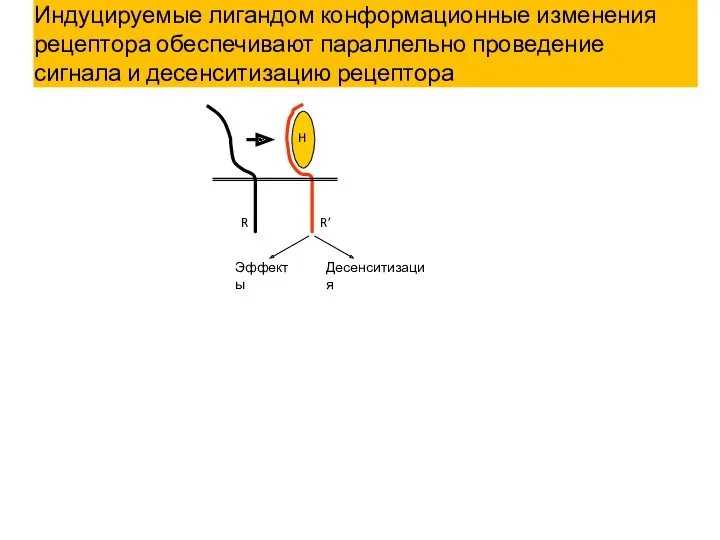
Эффекты
Десенситизация
R
R’
H
Индуцируемые лигандом конформационные изменения рецептора обеспечивают параллельно проведение сигнала и десенситизацию
Эффекты
Десенситизация
R
R’
H
Индуцируемые лигандом конформационные изменения рецептора обеспечивают параллельно проведение сигнала и десенситизацию

Ядерные рецепторы и
системы передачи сигнала
Ядерные рецепторы и
системы передачи сигнала

Специфические факторы транскрипции
Конститутивные факторы транскрипции =
факторы транскрипции базального транскрипционого комплекса:
Специфические факторы транскрипции
Конститутивные факторы транскрипции =
факторы транскрипции базального транскрипционого комплекса:

Способ передачи:
на входе:
Взаимодействие с гормоном
Взаимодействие с другим сигнальным соединением
Фосфорилирование
На
Способ передачи:
на входе:
Взаимодействие с гормоном
Взаимодействие с другим сигнальным соединением
Фосфорилирование
На

Корегуляторы ядерных рецепторов:
1. Корепрессоры
Помогают подавлять действие рецептора без
Корегуляторы ядерных рецепторов:
1. Корепрессоры
Помогают подавлять действие рецептора без
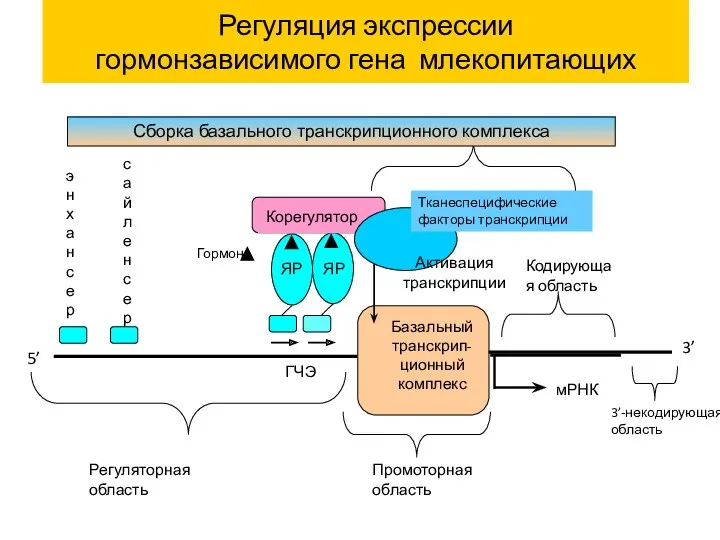
Регуляция экспрессии
гормонзависимого гена млекопитающих
Корегулятор
мРНК
Активация
транскрипции
ЯР
ЯР
ГЧЭ
Базальный
транскрип-ционный
комплекс
Гормон
Промоторная область
5’
3’
Кодирующая область
Тканеспецифические факторы
Регуляция экспрессии
гормонзависимого гена млекопитающих
Корегулятор
мРНК
Активация
транскрипции
ЯР
ЯР
ГЧЭ
Базальный
транскрип-ционный
комплекс
Гормон
Промоторная область
5’
3’
Кодирующая область
Тканеспецифические факторы

Лиганды ядерных рецепторов
Гормоны (стероидные, тиреоидоные)
Гормональные формы жирорастворимых витаминов (Д и А)
Сиротские:
Лиганды ядерных рецепторов
Гормоны (стероидные, тиреоидоные)
Гормональные формы жирорастворимых витаминов (Д и А)
Сиротские:
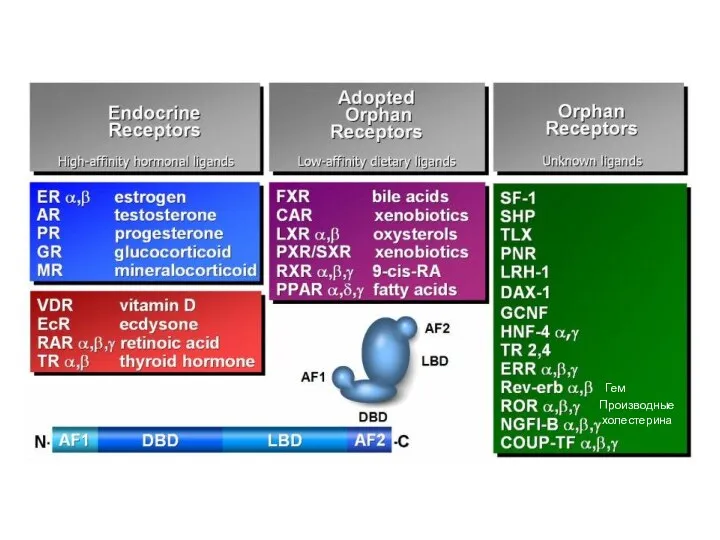
Производные
холестерина
Гем
Производные
холестерина
Гем
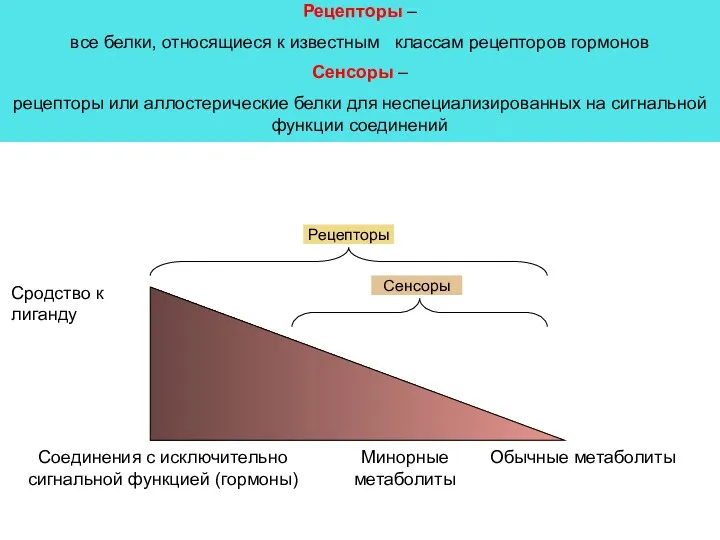
Сродство к лиганду
Соединения с исключительно сигнальной функцией (гормоны)
Обычные метаболиты
Минорные метаболиты
Рецепторы
Сенсоры
Рецепторы –
Сродство к лиганду
Соединения с исключительно сигнальной функцией (гормоны)
Обычные метаболиты
Минорные метаболиты
Рецепторы
Сенсоры
Рецепторы –

Активность ядерных рецепторов в отсутствие лиганда
Транскрипционно неактивные без лиганда : некоторые
Активность ядерных рецепторов в отсутствие лиганда
Транскрипционно неактивные без лиганда : некоторые
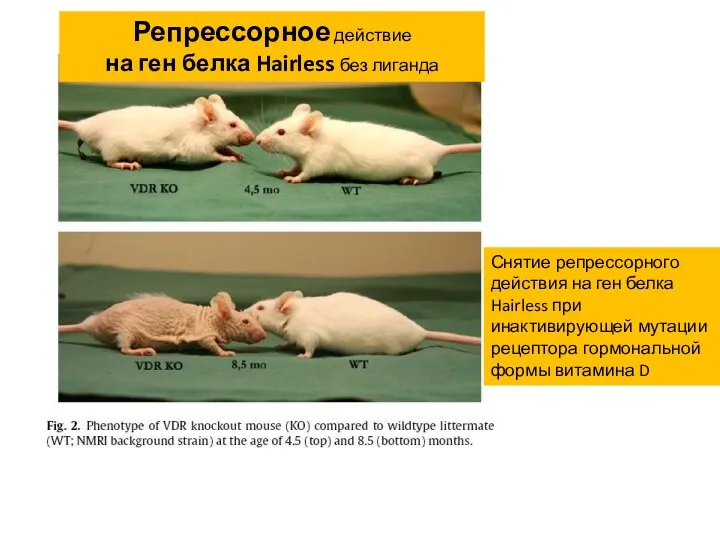
Снятие репрессорного действия на ген белка Hairless при инактивирующей мутации рецептора
Снятие репрессорного действия на ген белка Hairless при инактивирующей мутации рецептора
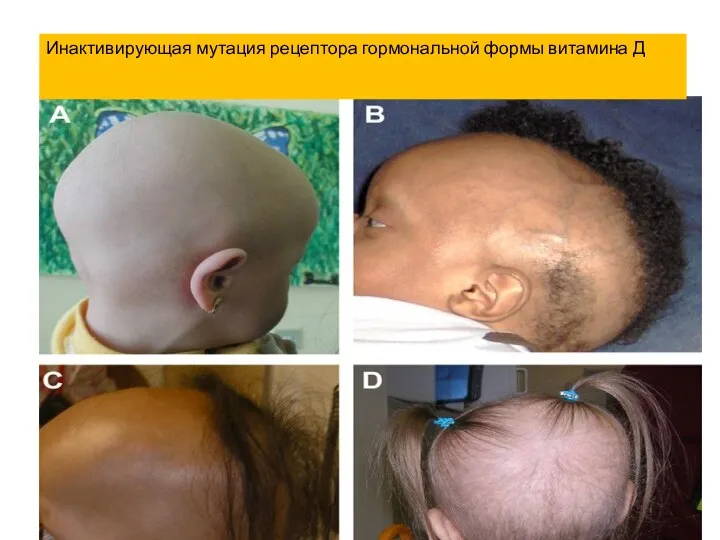
Инактивирующая мутация рецептора гормональной формы витамина Д
Инактивирующая мутация рецептора гормональной формы витамина Д
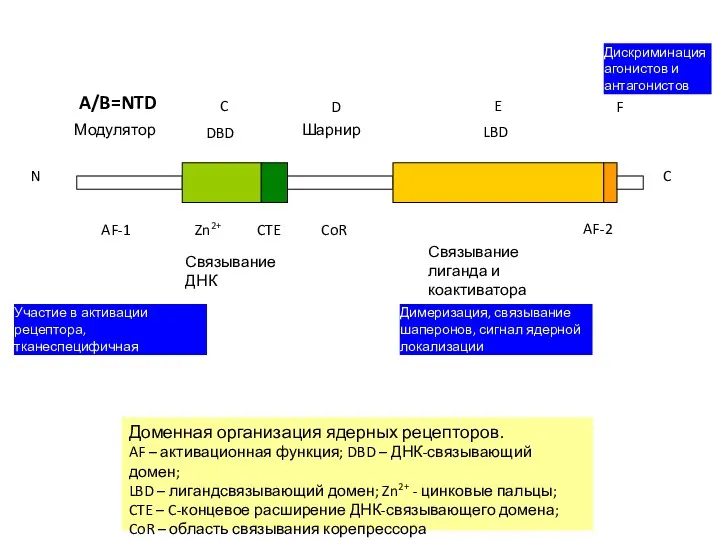
N
Связывание
ДНК
Связывание
лиганда и коактиватора
CoR
AF-2
CTE
Zn2+
AF-1
F
E
D
C
A/B=NTD
Модулятор
LBD
Шарнир
DBD
Доменная организация ядерных рецепторов.
AF – активационная функция; DBD –
N
Связывание
ДНК
Связывание
лиганда и коактиватора
CoR
AF-2
CTE
Zn2+
AF-1
F
E
D
C
A/B=NTD
Модулятор
LBD
Шарнир
DBD
Доменная организация ядерных рецепторов.
AF – активационная функция; DBD –

Домен NTD (A/B) (модуляторный):
Наиболее вариабельный по длине и послед-ти а/к
Сайты фосфорилирования
Домен NTD (A/B) (модуляторный):
Наиболее вариабельный по длине и послед-ти а/к
Сайты фосфорилирования
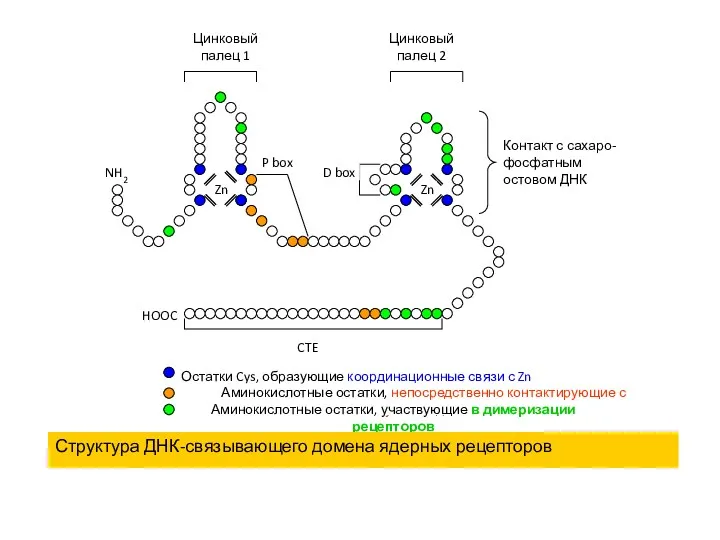
Структура ДНК-связывающего домена ядерных рецепторов
Контакт с сахаро-фосфатным остовом ДНК
Структура ДНК-связывающего домена ядерных рецепторов
Контакт с сахаро-фосфатным остовом ДНК

Гормончувствительные элементы (ГЧЭ) ДНК
Особенности взаимодействия ГЧЭ с ядерными рецепторами (ЯР):
Полусайты –связывание
Гормончувствительные элементы (ГЧЭ) ДНК
Особенности взаимодействия ГЧЭ с ядерными рецепторами (ЯР):
Полусайты –связывание
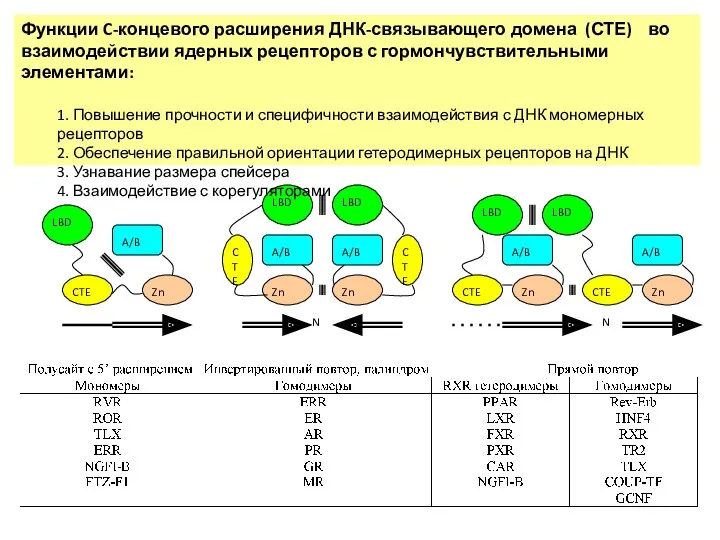
Функции C-концевого расширения ДНК-связывающего домена (СТЕ) во взаимодействии ядерных рецепторов с
Функции C-концевого расширения ДНК-связывающего домена (СТЕ) во взаимодействии ядерных рецепторов с
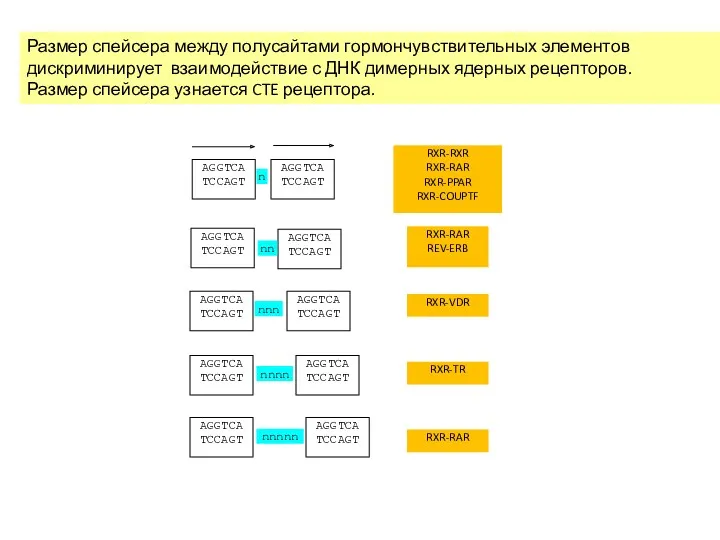
Размер спейсера между полусайтами гормончувствительных элементов дискриминирует взаимодействие с ДНК димерных
Размер спейсера между полусайтами гормончувствительных элементов дискриминирует взаимодействие с ДНК димерных
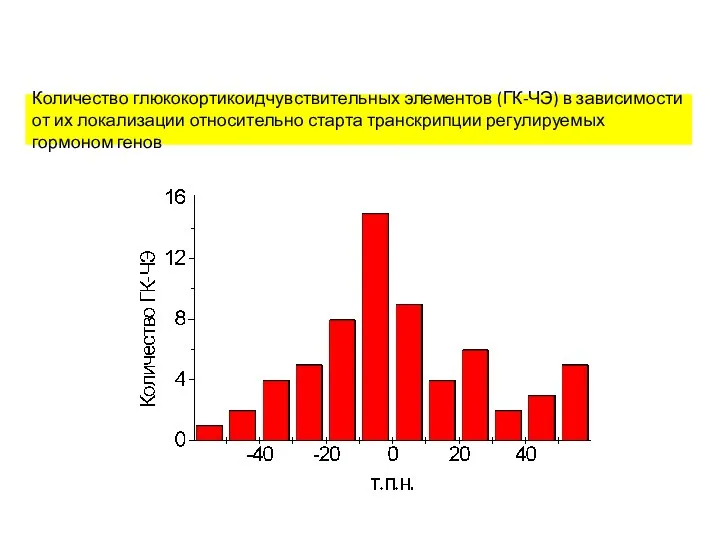
Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их локализации относительно старта
Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их локализации относительно старта

Шарнирный домен D
Вариабелен по размеру и последовательности а/к
Гибкий (поворот на 180°
Шарнирный домен D
Вариабелен по размеру и последовательности а/к
Гибкий (поворот на 180°
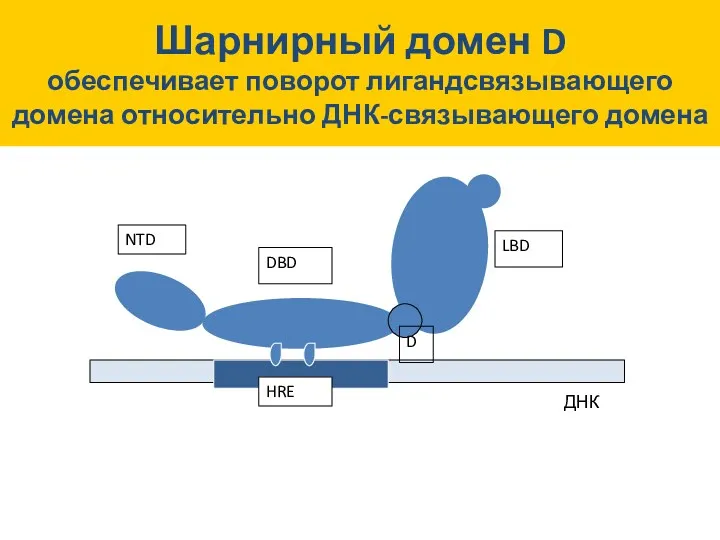
HRE
NTD
DBD
D
LBD
ДНК
Шарнирный домен D
обеспечивает поворот лигандсвязывающего домена относительно ДНК-связывающего домена
HRE
NTD
DBD
D
LBD
ДНК
Шарнирный домен D
обеспечивает поворот лигандсвязывающего домена относительно ДНК-связывающего домена

Умеренно консервативен
Лигандсвязывающий карман:
Связывание лиганда
Участие в димеризации
Взаимодействие с белками теплового шока
Сигнал ядерной
Умеренно консервативен
Лигандсвязывающий карман:
Связывание лиганда
Участие в димеризации
Взаимодействие с белками теплового шока
Сигнал ядерной
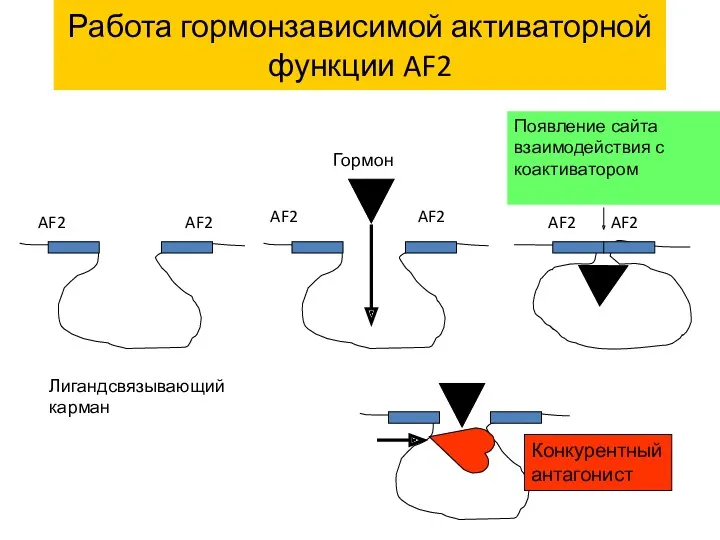
Работа гормонзависимой активаторной функции AF2
AF2
AF2
Лигандсвязывающий карман
Гормон
Появление сайта взаимодействия с коактиватором
AF2
AF2
AF2
AF2
Конкурентный антагонист
Работа гормонзависимой активаторной функции AF2
AF2
AF2
Лигандсвязывающий карман
Гормон
Появление сайта взаимодействия с коактиватором
AF2
AF2
AF2
AF2
Конкурентный антагонист
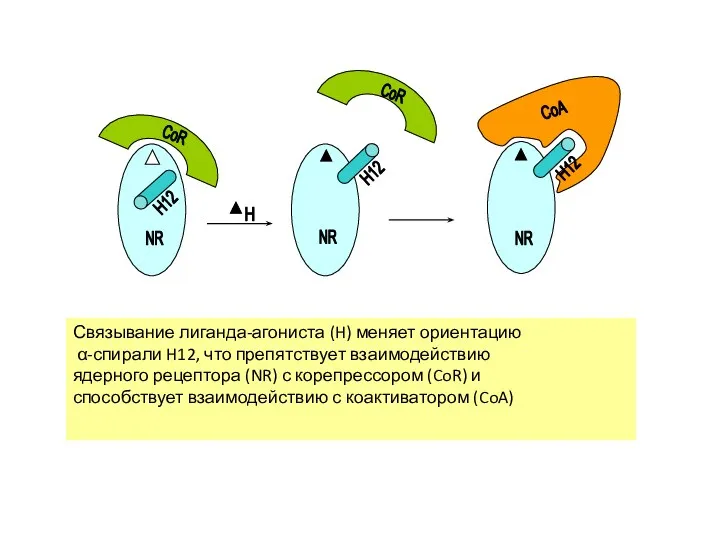
Связывание лиганда-агониста (H) меняет ориентацию
α-спирали H12, что препятствует взаимодействию
ядерного
Связывание лиганда-агониста (H) меняет ориентацию
α-спирали H12, что препятствует взаимодействию
ядерного

Роль димеризации
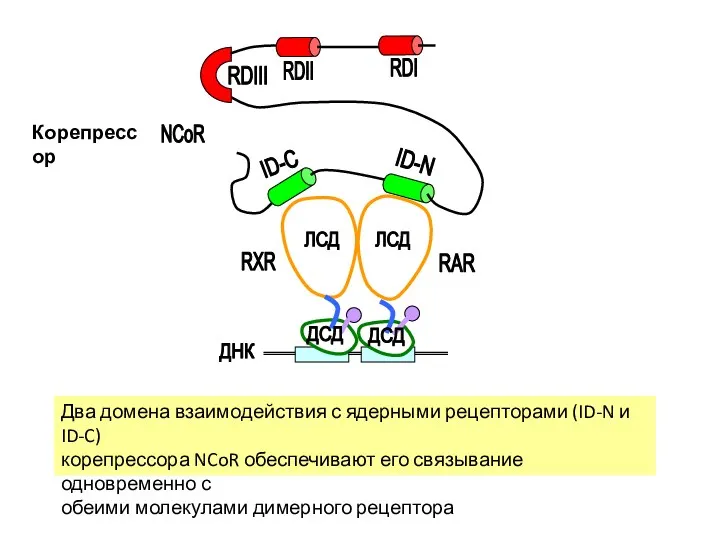
Два домена взаимодействия с ядерными рецепторами (ID-N и ID-C)
корепрессора NCoR
Два домена взаимодействия с ядерными рецепторами (ID-N и ID-C)
корепрессора NCoR
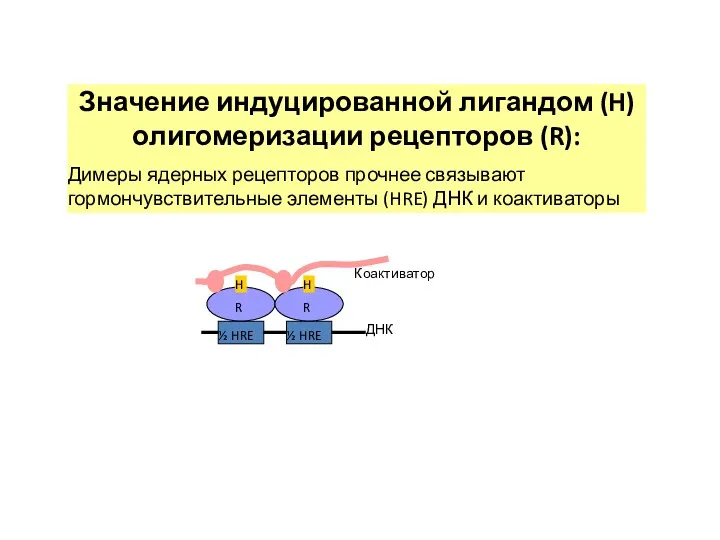
½ HRE
½ HRE
H
H
R
R
ДНК
Значение индуцированной лигандом (H) олигомеризации рецепторов (R):
Димеры ядерных рецепторов
½ HRE
½ HRE
H
H
R
R
ДНК
Значение индуцированной лигандом (H) олигомеризации рецепторов (R):
Димеры ядерных рецепторов
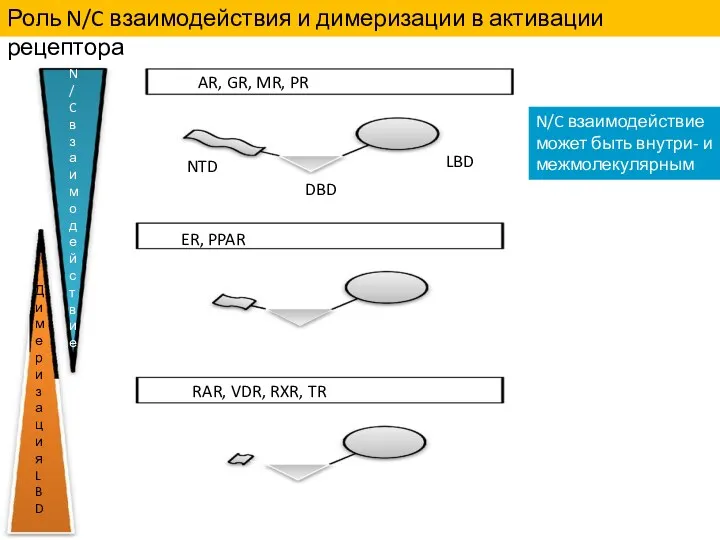
Роль N/C взаимодействия и димеризации в активации рецептора
N/C взаимодействие может быть
Роль N/C взаимодействия и димеризации в активации рецептора
N/C взаимодействие может быть

Партнеры по димеризации
В зависимости от партнера по димеризации чаще узнаются разные
Партнеры по димеризации
В зависимости от партнера по димеризации чаще узнаются разные
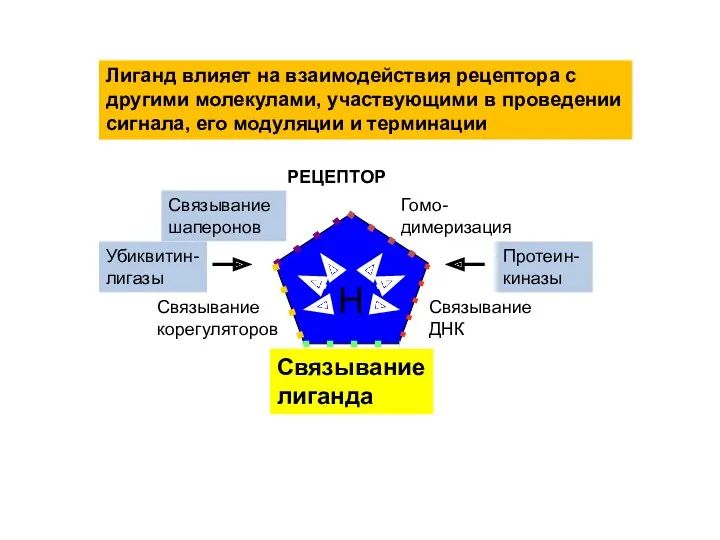
Связывание ДНК
Гомо-димеризация
Связывание шаперонов
Связывание корегуляторов
H
Связывание лиганда
Лиганд влияет на взаимодействия рецептора с другими
Связывание ДНК
Гомо-димеризация
Связывание шаперонов
Связывание корегуляторов
H
Связывание лиганда
Лиганд влияет на взаимодействия рецептора с другими

Роль комплексирования рецепторов стероидов с белками теплового шока (hsp90)
В цитоплазме гетеродимеры:
Роль комплексирования рецепторов стероидов с белками теплового шока (hsp90)
В цитоплазме гетеродимеры:
гормон
ядро
цитоплазма
А
NR
NR
гормон
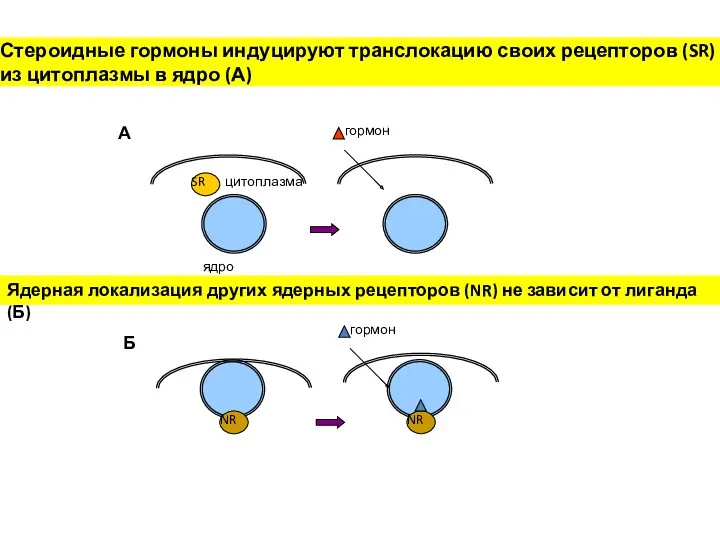
Стероидные гормоны индуцируют транслокацию своих рецепторов (SR) из цитоплазмы в ядро
гормон
ядро
цитоплазма
А
NR
NR
гормон
Стероидные гормоны индуцируют транслокацию своих рецепторов (SR) из цитоплазмы в ядро
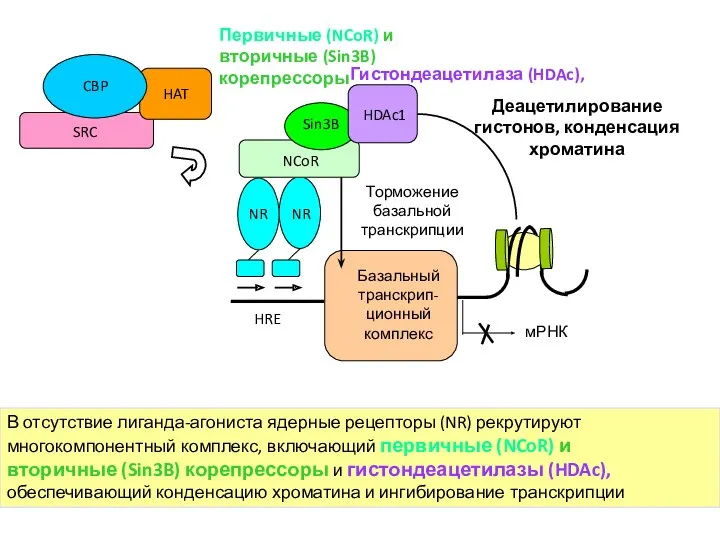
В отсутствие лиганда-агониста ядерные рецепторы (NR) рекрутируют
многокомпонентный комплекс, включающий первичные
В отсутствие лиганда-агониста ядерные рецепторы (NR) рекрутируют
многокомпонентный комплекс, включающий первичные
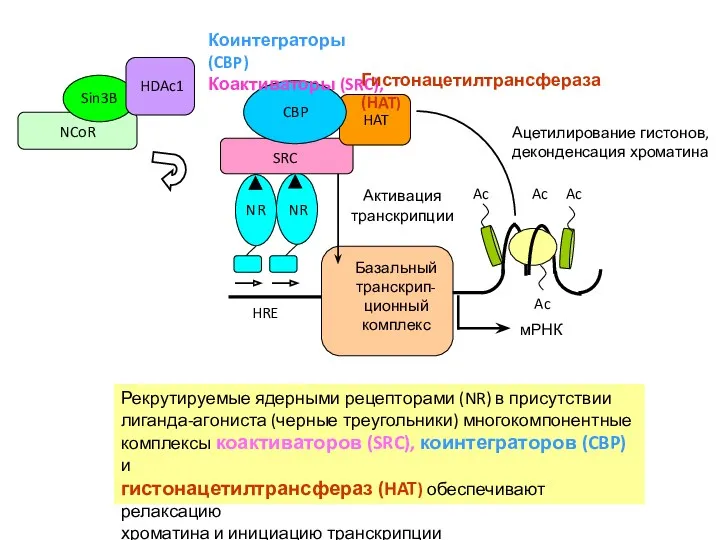
Рекрутируемые ядерными рецепторами (NR) в присутствии
лиганда-агониста (черные треугольники) многокомпонентные
комплексы
Рекрутируемые ядерными рецепторами (NR) в присутствии
лиганда-агониста (черные треугольники) многокомпонентные
комплексы
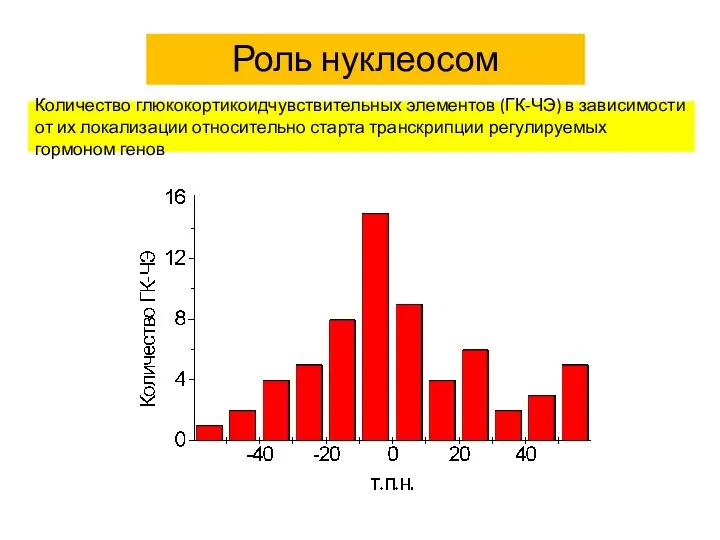
Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их локализации относительно старта
Количество глюкокортикоидчувствительных элементов (ГК-ЧЭ) в зависимости от их локализации относительно старта
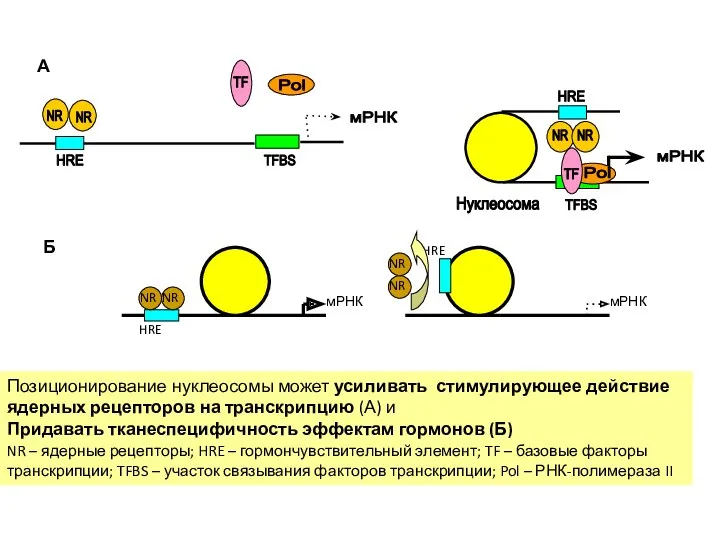
Позиционирование нуклеосомы может усиливать стимулирующее действие ядерных рецепторов на транскрипцию (А)
Позиционирование нуклеосомы может усиливать стимулирующее действие ядерных рецепторов на транскрипцию (А)
 Акушерство. История развития акушерства
Акушерство. История развития акушерства Первичный и вторичный гуморальный иммунный ответ. Переключение изотипов антител. Клетки памяти. Аффинность и авидность антител
Первичный и вторичный гуморальный иммунный ответ. Переключение изотипов антител. Клетки памяти. Аффинность и авидность антител Қазақстан Республикасындағы медицина ғылымы мен денсаулық сақтауды дамытудың концепциясы. Қ.Р денсаулық сақтаудың даму кезендері
Қазақстан Республикасындағы медицина ғылымы мен денсаулық сақтауды дамытудың концепциясы. Қ.Р денсаулық сақтаудың даму кезендері Острый живот в гинекологии (внематочная беременность, апоплексия яичника, перекрут ножки опухоли яичника)
Острый живот в гинекологии (внематочная беременность, апоплексия яичника, перекрут ножки опухоли яичника) Инфекционный эндокардит
Инфекционный эндокардит Плоскостопие у детей и эффективные методы его устранения на уроках физической культуры
Плоскостопие у детей и эффективные методы его устранения на уроках физической культуры Коарктация аорты
Коарктация аорты Школа бронхиальной астмы
Школа бронхиальной астмы Карбоновый пилинг
Карбоновый пилинг Антисептика. История развития
Антисептика. История развития Сестринский уход при ревматоидном полиартрите и деформирующем остеоартрозе
Сестринский уход при ревматоидном полиартрите и деформирующем остеоартрозе Иммунитет и проблемы питания жителей современного города
Иммунитет и проблемы питания жителей современного города Артикуляционно-акустическая дисграфия
Артикуляционно-акустическая дисграфия Коллекция от Askona
Коллекция от Askona Технология лекарств как наука. Задачи технологии лекарств
Технология лекарств как наука. Задачи технологии лекарств Развитие эмоциональной саморегуляции у больных, перенесших инсульт
Развитие эмоциональной саморегуляции у больных, перенесших инсульт Эндогенная профилактика кариеса зубов у детей
Эндогенная профилактика кариеса зубов у детей Голотопия желудка
Голотопия желудка Противовирусная терапия при невропатии лицевого нерва, ассоциированной с вирусом простого герпеса 1-го типа
Противовирусная терапия при невропатии лицевого нерва, ассоциированной с вирусом простого герпеса 1-го типа Стероидные противовоспалительные средства
Стероидные противовоспалительные средства Рентгенодиагностика туберкулеза
Рентгенодиагностика туберкулеза Дәрігердің басшылық қасиеттері және олардың қызметте алатын орны
Дәрігердің басшылық қасиеттері және олардың қызметте алатын орны 2 типті қант диабеті кезінде глюкофаж + инсулинді қолдану
2 типті қант диабеті кезінде глюкофаж + инсулинді қолдану Эндопротез тазобедренного сустава
Эндопротез тазобедренного сустава Введение в биологию
Введение в биологию Повышенная частота сердечных сокращений
Повышенная частота сердечных сокращений Зақымдаушы әсерлерге жасушалардың реакциясы. Жасушаның қартаюы және өлуі
Зақымдаушы әсерлерге жасушалардың реакциясы. Жасушаның қартаюы және өлуі Советы бобра Суперзуба. Как правильно чистить зубы?
Советы бобра Суперзуба. Как правильно чистить зубы?