- Микроағзалар генетикасы

Содержание
- 2. № 1 дәріс Микроағзалар генетикасына кіріспе Жоспары І. Микроағзалар генетикасының ерекшеліктері. ІІ. Микроағзаларды БТ қолдану артықшылықтары.
- 3. І. Микроағзалар генетикасының ерекшеліктері. Барлық организмдерде, сонымен қатар бактериялар мен вирустарда генетикалық қасиеттерді анықтайтын тұқымқуалаушылықтың материалдық
- 4. ІІ. Микроағзаларды БТ қолдану артықшылықтары. 1. Гаплоидтылық, яғни бір хромосоманың болуы, доминанттылық көрінісін болдырмайды. 2. Жоғары
- 5. ІІ. Микроағзаларды БТ қолдану артықшылықтары. Қазіргі молекулалық генетиканың жетістігі гендік инженерия тәсілдерінің жетілуімен байланысты – ол
- 6. ІІІ. Микроағзаларды БТ қолданатын салалары. Медицина. (Str. griseus. Bacillus brevis) Ауыл шаруашылығы. (Agrobacterium tumifaciens, Rizobium, Azotobacter)
- 7. ІІІ. Микроағзаларды БТ қолданатын салалары. Кейбір мысалдар: Медицина тарихында ерекше орын алатын ол антибиотиктер. Е. Хаувинк
- 8. Тағам өнеркәсібі. Ашыту кезінде биотехнологиялық өнім алынады. Клеткада энергетикалық зат алмасу және биосинтез бір мезгілде және
- 9. Биоремедация. Ксенобиотиктердің биодеградациясы, оның ішінде жылдар бойы қоқыстарда, суда және топырақтарда жиналатын пестицидтер мен токсикалық қосылыстар,
- 10. № 2 дәріс Бактерияларға ДНҚ-ны енгізу. 1. Микроағзаларда рекомбинация түрлері. 2. Трансформация арқылы тікелей енгізу. 3.
- 11. 1. Микроағзаларда рекомбинация түрлері. Бактерия донордан, бактерия рецепиентке тек бір бөлігі түседі (экзогенді ДНҚ) осы процестерді
- 12. 1. Микроағзаларда рекомбинация түрлері. 1. Жалпы немесе гомологиялық рекомбинация, әрекетесіп жатқан ДНҚ-дың бір-біріне гомологиялық үлескілері болса
- 14. 1. Микроағзаларда рекомбинация түрлері. 2. Заңды емес (Незаконная) рекомбинация ДНҚ-дың арасында гомолгиялық үлескілерді қажет етпейді. Заңды
- 15. 2. Трансформация арқылы тікелей енгізу. Трансформация бактерияларда табылған генетикалық алмасудың бірінші үдерісі болып есептеледі. 1928 ж
- 18. 2. Трансформация арқылы тікелей енгізу. 1944 ж. Освалбд Т. Эвери, Колин М. Маклеод және Маккарти әйгілі
- 19. 2. Трансформация арқылы тікелей енгізу. Құзыреттілік (компетентность) деп аталатын ДНҚ-ны сіңіру қабілеті әдетте белгілі бір жағдайларда
- 20. 2. Трансформация арқылы тікелей енгізу. Ішек таяқшасы жасушаларын ең бірінші О° С температурасында СаСl₂ өте жоғарғы
- 22. 3. Коньюгация арқылы енгізу. 1946 ж. Джошуа Ледерберг пен Эдвард Тайтэм бактериялар гендерінің конъюгациялық тасымалдауын ашты.
- 23. 1. Донор клетка жыныстық пилиді шығарады. 2. Пили рецепиент жасушаға бекінеді де екі жасушаны біріктіреді. 3.F-плазмида
- 24. 4. Бактериофаг ДНҚ-сын енгізу және трансдукция. Бактериофагтар бактерия жасушаларын зақымдаған кезде әрбір вирус сезімтал жасуша-иесінде адсорбцияланып,
- 25. Бірінші бактериофаг жасушаның бетінде адсорбцияланады. Содан кейін фагтың ДНҚ-сы цитоплазмаға енеді. Цитоплазмада фагтың ДНҚ-сы мен фаг
- 26. 4. Бактериофаг ДНҚ-сын енгізу және трансдукция. Фагтар бактерия жасушасында екі жолмен дамуы мүмкін: Лизистік (литический) -Бактерияға
- 27. № 3 дәріс Биопрепараттар өндірісі 1. Ішек микрофлорасы 2. Пробиотиктер 3. Пребиотиктер 4. Симбиотиктер 5. Пробиотиктер
- 28. 1. Ішек микрофлорасы Адам ішегінде 100 трилионға жуық микроорганизім өмір сүреді. Жалпы есепен алғанда 300 бен
- 29. 1. Ішек микрофлорасы Ішектегі микроорганизімдер түрі: анаэробты бактероидтар бифидобактериялар энтерококалар клостридиялар эубактериялар аэробты энтеробактериялар стрептококалар стафилококалар
- 30. 1. Ішек микрофлорасы Ішек микроорганизімдерінің қызметі: Ыдырату: көмірсуларды, майларды, ақуызды ыдырату. Имундық: Имуноглобулиндердің түзілуіне маңызды заттар
- 31. Ішектің астарында көптеген имундық жасушалар орналасқан (пейеровые бляшки)
- 32. 1. Ішек микрофлорасы Ішек микрофлорасын бұзатын факторлар: Антибиотиктер және басқада дәрі дәрмектер. Микробтық инфекциялар. Диета. Хроникалық
- 33. 1. Пробиотиктер Пробиотиктер - дәрілік препараттар немесе тағамға биологиялық активті қоспалар ретінде қолданылады, құрамы тірі микроорганизмдерден
- 34. 1. Пробиотиктер Пробиотиктер пайдасы: Сүт қышқылын өндіріеді, ол өз кезегінде ішіктін ph деңгейін дүрыстайды. Патагенді бактерияларға
- 35. 1. Пробиотиктер Пробиотиктер құрамы: Lactobacillus түрлері. L. acidophilus. L. plantarum. L. cased. L. brevis. L. bulgaricus.
- 36. 1. Пробиотиктер Құрғақ және сұйық пробиотиктер: Құрғақ пробиотиктер – олр лиофилизацияланған микроорганизмдерден тұрады, ол ұнтақ, капсула
- 37. 3. Пребиотиктер Пребиотиктер - асқазан-ішек жолының жоғары бөліктерінде қорытылмайтын және сіңірілмейтін, бірақ адамның тоқ ішегінің пайдалы
- 38. 3. Пребиотиктер Қызметі: Олар жуан ішекте өмір сүретін бактериялардың топтарының метоболитикалық активтілігін, стимуляциялануына байланысты денсаулықты жақсартады.
- 39. 3. Пребиотиктер Пребиотиктер, пробиотиктерге қарағанда, микрофлораға артығырақ ықпал ете алады, себебі: асқазан-ішек жолында бұзылмайды және антибиотиктердің
- 40. 3. Пребиотиктер
- 41. 4. Симбиотиктер Симбиотиктер – бұл пробиотиктердін және пребиотиктердің қосындысы.
- 43. 5. Пробиотиктер өндірісі. Ферментация: қоректік орта + микроорганизм Филтрация: қоректік ортаны. Лиофилизация: бактерия -18-17° С аралығында
- 44. № 4 дәріс Микробты инсектицидтер 1. Пестицидтер тарихы. 2. Химиялық пестицидтерді қолданудың кемшіліктері. 3. Микробты инсектицидтер
- 45. 1. Инсектицидтер тарихы. Ауылшаруашылық дақылдар үшін адам мен бунақденелілердің арасындағы бәсекелестіктің тарихы ауылшаруашылық сияқты өте бұрыннан
- 46. 2. Химиялық пестицидтерді қолданудың кемшіліктері. Химиялық пестицидтерді қолданудың келесі кемшіліктері бар: Бір химиялық қосылыстың кең қолданылуы,
- 47. 3. Микробты инсектицидтер Жоғарыда көрсетілген кемшіліктердің бірігіп әсер етуі зиянкестермен күресудің жаңа баламалы амалдарын іздеуге күшті
- 48. 3. Микробты инсектицидтер Осы пайдалы ерекшелікке қарамастан зиянкестермен күресудің микробты агенттері барлық сатылатын инсектицидтердің 1% -
- 49. № 5 дәріс Bacillus thuringiensis 1. Bacillus thuringiensis ашылу тарихы. 2. Bacillus thuringiensis-тің әсер ету механизымы.
- 50. 1. Bacillus thuringiensis ашылу тарихы. Шигатане Ишивате В. thuringiensis бактериясын ашты. Ол 1901 жылы Жапонияда фляшениямен
- 51. 1. Bacillus thuringiensis ашылу тарихы. Джо-эл Маргалит қос қанаттылардың бірінші патотипі В. thuringiensis var. Israelensis-тің ашылуы
- 52. Жіктелу және филогения Патшалық Тип Класс Отряд Тұқымдас Туыс Түр Жалпы келісім бойынша түр бойынша ғылыми
- 53. Жіктелу және филогения Санат Патшалық Тип Класс Отряд Тұқымдас Туыс Түр Архейлер Crenarchaeota Thermoprotei Sulfolobales Sulfolobaceae
- 54. 1. Bacillus thuringiensis ашылу тарихы. В. Thuringiensis не өлі ағзалардың ыдыраған органикалық заттармен, не тірі бунақденелілердің
- 55. 2. Bacillus thuringiensis-тің әсер ету механизымы. 1915 ж фляшениямен ауырған дернәсілдерін зерттегенде В.t. Споруляция кезінде уытты
- 56. 2. Bacillus thuringiensis-тің әсер ету механизымы. Осы протоксин дернәсіл ішегіне түскеннен кейін олардын ішектін сілтілі сөл
- 57. 3. Bacillus thuringiensis бөлетін δ-эндотоксин түрлері. B. thuringiensis кейбір штамдары тек қана бір δ-эндотоксинді өндірсе, басқалары
- 58. 3. Bacillus thuringiensis бөлетін δ-эндотоксин түрлері. Осы кристалдық нәруыздардың гендерің 2-і топқа жіктейді: Сry – ағза
- 59. 4. B. thuringiensis В-экзотоксині. Вегетацияның белсенді кезеңінде B. thuringiensis кейбір штамдары β-экзотоксиндер деп аталатын төмен молекулалық
- 60. 5. B. THURINGIENSIS ISRAELENSIS түршесі инсектицид ретінде. В. T var israelensis (Bti) түршесінің параспоралық қосындыларында қос
- 61. № 6 дәріс Бунақденелілерге төзімді трансгендік дақылдар. 1. Кіріспе. 2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы. 3.
- 62. 1. Кіріспе. Бүкіл әлемде 30 миллион акрдан көп жерде осы бунақденелерге төзімді дақылдар өсіріледі, осы дақылдарда
- 63. 2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы. Өсімдіктер жасушаларына бөгде ДНҚ енгізу үшін үш әдіс қолданылады: 1.
- 64. 2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы.
- 65. 3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы. 1992 жылы Монсанто қабыршақ қанатты бунақденелілердің
- 67. 3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы. Ori322/rop сайты. Инженерлік және PV-GHBK04 амплификациялық
- 68. 3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы. P-35S, nptII және NOS3’ осы үш
- 69. 3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы. Aad алтын түсті стафилококадан бөлініп алынған
- 70. 3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы. 7S3’ – Бұл сайт cry1Ac генін
- 71. № 7 дәріс Агробактерия плазмидаларын вектор ретінде қолдану. 1. Жалпы сипаттамасы. 2. Зақымдау процесі.
- 72. 1. Жалпы сипаттамасы. Agrobactera деген топырақта мекендейтін гарам теріс бактериялардың тобы. Олар өсімдіктерге жұғып тәж тәрізді
- 73. 1. Жалпы сипаттамасы. Ісік клеткаларында сау клеткалардан болмайтын опиндер деген жаңа класқа жататын химиялық заттар табылды.
- 74. Ті плазмиданың генетикалық картасы: Т-ДНҚ құрамына ауксин, цитокинин және опин гендері бар, олар тек өсімдік жасушаларында
- 75. 1. Жалпы сипаттамасы. Vir гендері. Vir A өсімдік заықмдалуына сезімтал. (Ацитсерингон) Vir B Тасымалдау үшін құрылым.
- 76. 2. Зақымдау процесі. Зақымдалған өсімдік ұлпалары ацитосерингонды бөліп шығарады. Оны A. tumefaciens сигнал ретінде қабылдайды. Осы
- 77. 2. Зақымдау процесі. Ары қарай Vir D1 кетіп Vir D2 қалып 5’ ұшында пилот қызметін атқарады,
- 79. Скачать презентацию

№ 1 дәріс
Микроағзалар генетикасына кіріспе
Жоспары
І. Микроағзалар генетикасының ерекшеліктері.
ІІ. Микроағзаларды БТ
№ 1 дәріс
Микроағзалар генетикасына кіріспе
Жоспары
І. Микроағзалар генетикасының ерекшеліктері.
ІІ. Микроағзаларды БТ

І. Микроағзалар генетикасының ерекшеліктері.
Барлық организмдерде, сонымен қатар бактериялар мен вирустарда
І. Микроағзалар генетикасының ерекшеліктері.
Барлық организмдерде, сонымен қатар бактериялар мен вирустарда

ІІ. Микроағзаларды БТ қолдану артықшылықтары.
1. Гаплоидтылық, яғни бір хромосоманың болуы, доминанттылық
ІІ. Микроағзаларды БТ қолдану артықшылықтары.
1. Гаплоидтылық, яғни бір хромосоманың болуы, доминанттылық

ІІ. Микроағзаларды БТ қолдану артықшылықтары.
Қазіргі молекулалық генетиканың жетістігі гендік инженерия тәсілдерінің
ІІ. Микроағзаларды БТ қолдану артықшылықтары.
Қазіргі молекулалық генетиканың жетістігі гендік инженерия тәсілдерінің

ІІІ. Микроағзаларды БТ қолданатын салалары.
Медицина. (Str. griseus. Bacillus brevis)
Ауыл шаруашылығы.
ІІІ. Микроағзаларды БТ қолданатын салалары.
Медицина. (Str. griseus. Bacillus brevis)
Ауыл шаруашылығы.

ІІІ. Микроағзаларды БТ қолданатын салалары.
Кейбір мысалдар:
Медицина тарихында ерекше орын алатын ол
ІІІ. Микроағзаларды БТ қолданатын салалары.
Кейбір мысалдар:
Медицина тарихында ерекше орын алатын ол

Тағам өнеркәсібі.
Ашыту кезінде биотехнологиялық өнім алынады. Клеткада энергетикалық зат алмасу және
Тағам өнеркәсібі.
Ашыту кезінде биотехнологиялық өнім алынады. Клеткада энергетикалық зат алмасу және

Биоремедация.
Ксенобиотиктердің биодеградациясы, оның ішінде жылдар бойы қоқыстарда, суда және топырақтарда жиналатын
Биоремедация.
Ксенобиотиктердің биодеградациясы, оның ішінде жылдар бойы қоқыстарда, суда және топырақтарда жиналатын

№ 2 дәріс
Бактерияларға ДНҚ-ны енгізу.
1. Микроағзаларда рекомбинация түрлері.
2. Трансформация арқылы тікелей
№ 2 дәріс
Бактерияларға ДНҚ-ны енгізу.
1. Микроағзаларда рекомбинация түрлері.
2. Трансформация арқылы тікелей

1. Микроағзаларда рекомбинация түрлері.
Бактерия донордан, бактерия рецепиентке тек бір бөлігі түседі
1. Микроағзаларда рекомбинация түрлері.
Бактерия донордан, бактерия рецепиентке тек бір бөлігі түседі

1. Микроағзаларда рекомбинация түрлері.
1. Жалпы немесе гомологиялық рекомбинация, әрекетесіп жатқан ДНҚ-дың
1. Микроағзаларда рекомбинация түрлері.
1. Жалпы немесе гомологиялық рекомбинация, әрекетесіп жатқан ДНҚ-дың
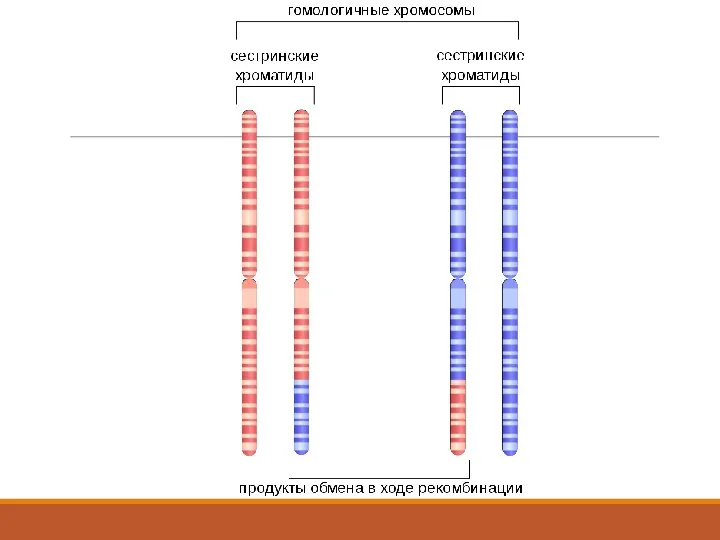

1. Микроағзаларда рекомбинация түрлері.
2. Заңды емес (Незаконная) рекомбинация ДНҚ-дың арасында гомолгиялық
1. Микроағзаларда рекомбинация түрлері.
2. Заңды емес (Незаконная) рекомбинация ДНҚ-дың арасында гомолгиялық

2. Трансформация арқылы тікелей енгізу.
Трансформация бактерияларда табылған генетикалық алмасудың бірінші үдерісі
2. Трансформация арқылы тікелей енгізу.
Трансформация бактерияларда табылған генетикалық алмасудың бірінші үдерісі
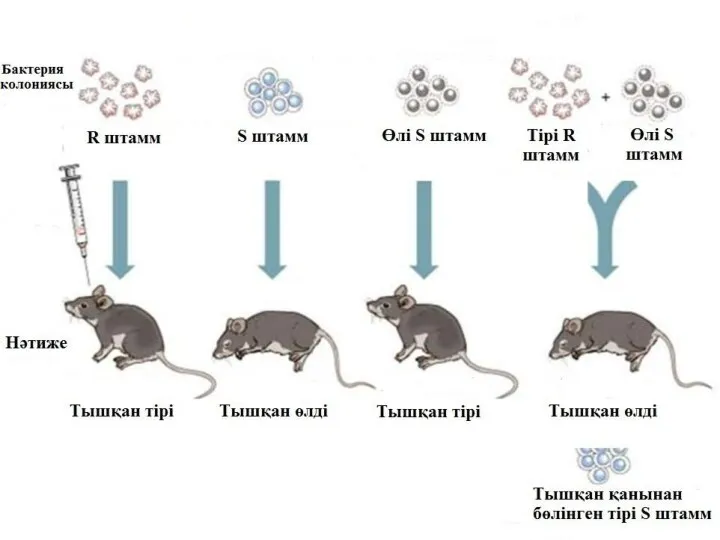
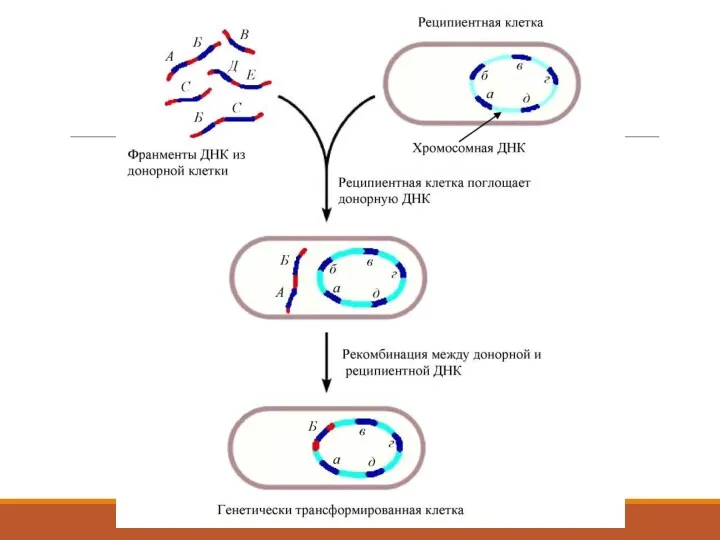

2. Трансформация арқылы тікелей енгізу.
1944 ж. Освалбд Т. Эвери, Колин М.
2. Трансформация арқылы тікелей енгізу.
1944 ж. Освалбд Т. Эвери, Колин М.

2. Трансформация арқылы тікелей енгізу.
Құзыреттілік (компетентность) деп аталатын ДНҚ-ны сіңіру қабілеті
2. Трансформация арқылы тікелей енгізу.
Құзыреттілік (компетентность) деп аталатын ДНҚ-ны сіңіру қабілеті

2. Трансформация арқылы тікелей енгізу.
Ішек таяқшасы жасушаларын ең бірінші О° С
2. Трансформация арқылы тікелей енгізу.
Ішек таяқшасы жасушаларын ең бірінші О° С
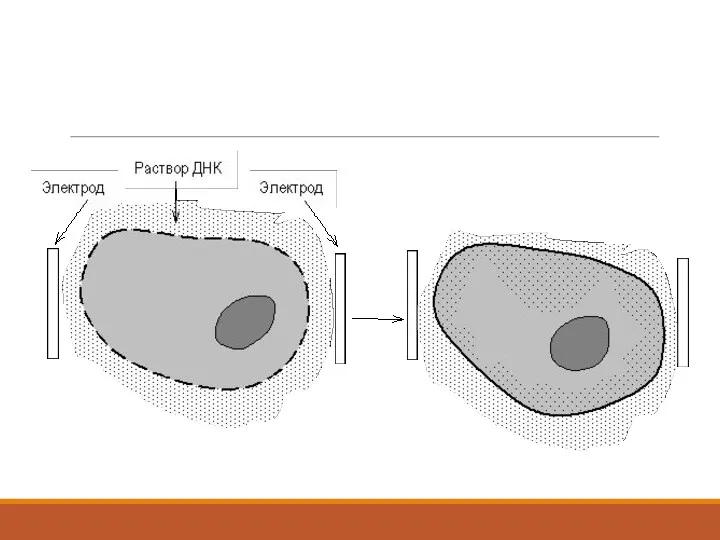

3. Коньюгация арқылы енгізу.
1946 ж. Джошуа Ледерберг пен Эдвард Тайтэм бактериялар
3. Коньюгация арқылы енгізу.
1946 ж. Джошуа Ледерберг пен Эдвард Тайтэм бактериялар
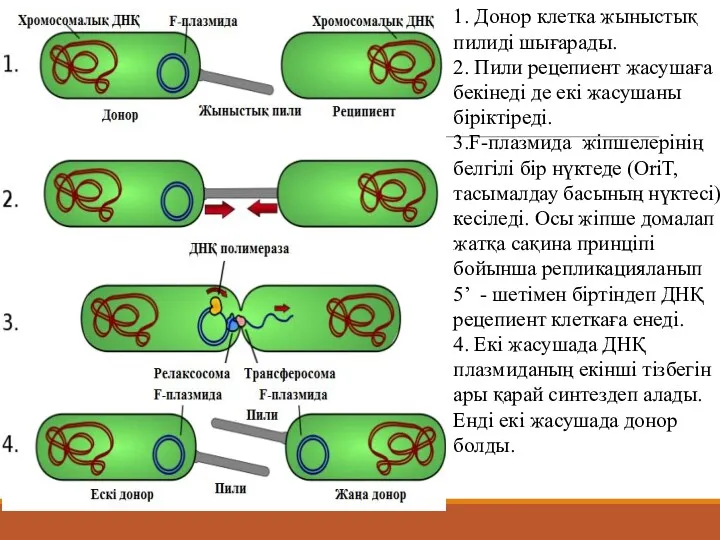
1. Донор клетка жыныстық пилиді шығарады.
2. Пили рецепиент жасушаға бекінеді де
1. Донор клетка жыныстық пилиді шығарады. 2. Пили рецепиент жасушаға бекінеді де

4. Бактериофаг ДНҚ-сын енгізу және трансдукция.
Бактериофагтар бактерия жасушаларын зақымдаған кезде әрбір
4. Бактериофаг ДНҚ-сын енгізу және трансдукция.
Бактериофагтар бактерия жасушаларын зақымдаған кезде әрбір

Бірінші бактериофаг жасушаның бетінде адсорбцияланады.
Содан кейін фагтың ДНҚ-сы цитоплазмаға енеді.
Бірінші бактериофаг жасушаның бетінде адсорбцияланады. Содан кейін фагтың ДНҚ-сы цитоплазмаға енеді.

4. Бактериофаг ДНҚ-сын енгізу және трансдукция.
Фагтар бактерия жасушасында екі жолмен дамуы
4. Бактериофаг ДНҚ-сын енгізу және трансдукция.
Фагтар бактерия жасушасында екі жолмен дамуы

№ 3 дәріс
Биопрепараттар өндірісі
1. Ішек микрофлорасы
2. Пробиотиктер
3. Пребиотиктер
4. Симбиотиктер
5. Пробиотиктер өндірісі.
№ 3 дәріс
Биопрепараттар өндірісі
1. Ішек микрофлорасы
2. Пробиотиктер
3. Пребиотиктер
4. Симбиотиктер
5. Пробиотиктер өндірісі.

1. Ішек микрофлорасы
Адам ішегінде 100 трилионға жуық микроорганизім өмір сүреді. Жалпы
1. Ішек микрофлорасы
Адам ішегінде 100 трилионға жуық микроорганизім өмір сүреді. Жалпы

1. Ішек микрофлорасы
Ішектегі микроорганизімдер түрі:
анаэробты
бактероидтар
бифидобактериялар
энтерококалар
клостридиялар
эубактериялар
аэробты
энтеробактериялар
стрептококалар
стафилококалар
лактобактериялар
санырауқұларқтар
1. Ішек микрофлорасы
Ішектегі микроорганизімдер түрі:
анаэробты
бактероидтар
бифидобактериялар
энтерококалар
клостридиялар
эубактериялар
аэробты
энтеробактериялар
стрептококалар
стафилококалар
лактобактериялар
санырауқұларқтар

1. Ішек микрофлорасы
Ішек микроорганизімдерінің қызметі:
Ыдырату: көмірсуларды, майларды, ақуызды ыдырату.
Имундық:
1. Ішек микрофлорасы
Ішек микроорганизімдерінің қызметі:
Ыдырату: көмірсуларды, майларды, ақуызды ыдырату.
Имундық:
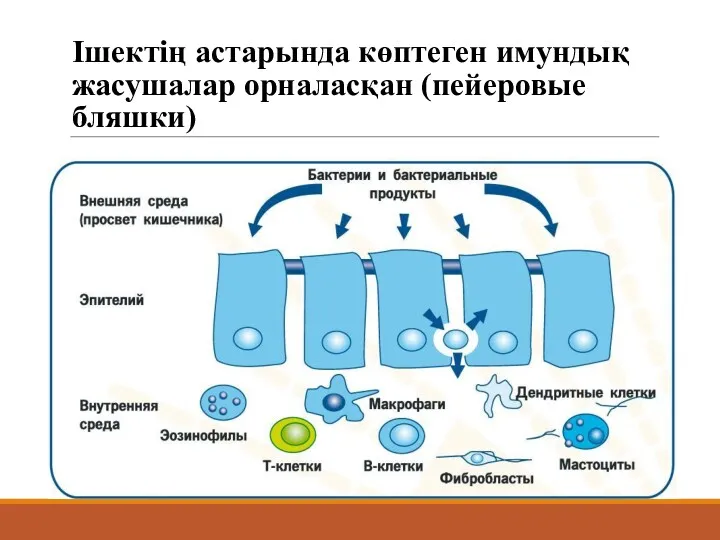
Ішектің астарында көптеген имундық жасушалар орналасқан (пейеровые бляшки)
Ішектің астарында көптеген имундық жасушалар орналасқан (пейеровые бляшки)

1. Ішек микрофлорасы
Ішек микрофлорасын бұзатын факторлар:
Антибиотиктер және басқада дәрі дәрмектер.
1. Ішек микрофлорасы
Ішек микрофлорасын бұзатын факторлар:
Антибиотиктер және басқада дәрі дәрмектер.

1. Пробиотиктер
Пробиотиктер - дәрілік препараттар немесе тағамға биологиялық активті қоспалар ретінде
1. Пробиотиктер
Пробиотиктер - дәрілік препараттар немесе тағамға биологиялық активті қоспалар ретінде

1. Пробиотиктер
Пробиотиктер пайдасы:
Сүт қышқылын өндіріеді, ол өз кезегінде ішіктін ph
1. Пробиотиктер
Пробиотиктер пайдасы:
Сүт қышқылын өндіріеді, ол өз кезегінде ішіктін ph

1. Пробиотиктер
Пробиотиктер құрамы:
Lactobacillus түрлері.
L. acidophilus.
L. plantarum.
L. cased.
L.
1. Пробиотиктер
Пробиотиктер құрамы:
Lactobacillus түрлері.
L. acidophilus.
L. plantarum.
L. cased.
L.

1. Пробиотиктер
Құрғақ және сұйық пробиотиктер:
Құрғақ пробиотиктер – олр лиофилизацияланған микроорганизмдерден тұрады,
1. Пробиотиктер
Құрғақ және сұйық пробиотиктер:
Құрғақ пробиотиктер – олр лиофилизацияланған микроорганизмдерден тұрады,

3. Пребиотиктер
Пребиотиктер - асқазан-ішек жолының жоғары бөліктерінде қорытылмайтын және сіңірілмейтін, бірақ
3. Пребиотиктер
Пребиотиктер - асқазан-ішек жолының жоғары бөліктерінде қорытылмайтын және сіңірілмейтін, бірақ

3. Пребиотиктер
Қызметі:
Олар жуан ішекте өмір сүретін бактериялардың топтарының метоболитикалық активтілігін,
3. Пребиотиктер
Қызметі:
Олар жуан ішекте өмір сүретін бактериялардың топтарының метоболитикалық активтілігін,

3. Пребиотиктер
Пребиотиктер, пробиотиктерге қарағанда, микрофлораға артығырақ ықпал ете алады, себебі:
асқазан-ішек жолында
3. Пребиотиктер
Пребиотиктер, пробиотиктерге қарағанда, микрофлораға артығырақ ықпал ете алады, себебі:
асқазан-ішек жолында

3. Пребиотиктер
3. Пребиотиктер

4. Симбиотиктер
Симбиотиктер – бұл пробиотиктердін және пребиотиктердің қосындысы.
4. Симбиотиктер
Симбиотиктер – бұл пробиотиктердін және пребиотиктердің қосындысы.


5. Пробиотиктер өндірісі.
Ферментация: қоректік орта + микроорганизм
Филтрация: қоректік ортаны.
Лиофилизация: бактерия -18-17°
5. Пробиотиктер өндірісі.
Ферментация: қоректік орта + микроорганизм
Филтрация: қоректік ортаны.
Лиофилизация: бактерия -18-17°

№ 4 дәріс
Микробты инсектицидтер
1. Пестицидтер тарихы.
2. Химиялық пестицидтерді қолданудың кемшіліктері.
3. Микробты
№ 4 дәріс
Микробты инсектицидтер
1. Пестицидтер тарихы.
2. Химиялық пестицидтерді қолданудың кемшіліктері.
3. Микробты

1. Инсектицидтер тарихы.
Ауылшаруашылық дақылдар үшін адам мен бунақденелілердің арасындағы бәсекелестіктің
1. Инсектицидтер тарихы.
Ауылшаруашылық дақылдар үшін адам мен бунақденелілердің арасындағы бәсекелестіктің

2. Химиялық пестицидтерді қолданудың кемшіліктері.
Химиялық пестицидтерді қолданудың келесі кемшіліктері бар:
Бір
2. Химиялық пестицидтерді қолданудың кемшіліктері.
Химиялық пестицидтерді қолданудың келесі кемшіліктері бар:
Бір

3. Микробты инсектицидтер
Жоғарыда көрсетілген кемшіліктердің бірігіп әсер етуі зиянкестермен күресудің жаңа
3. Микробты инсектицидтер
Жоғарыда көрсетілген кемшіліктердің бірігіп әсер етуі зиянкестермен күресудің жаңа

3. Микробты инсектицидтер
Осы пайдалы ерекшелікке қарамастан зиянкестермен күресудің микробты агенттері барлық
3. Микробты инсектицидтер
Осы пайдалы ерекшелікке қарамастан зиянкестермен күресудің микробты агенттері барлық

№ 5 дәріс
Bacillus thuringiensis
1. Bacillus thuringiensis ашылу тарихы.
2. Bacillus thuringiensis-тің әсер
№ 5 дәріс
Bacillus thuringiensis
1. Bacillus thuringiensis ашылу тарихы.
2. Bacillus thuringiensis-тің әсер

1. Bacillus thuringiensis ашылу тарихы.
Шигатане Ишивате В. thuringiensis бактериясын ашты. Ол
1. Bacillus thuringiensis ашылу тарихы.
Шигатане Ишивате В. thuringiensis бактериясын ашты. Ол

1. Bacillus thuringiensis ашылу тарихы.
Джо-эл Маргалит қос қанаттылардың бірінші патотипі В.
1. Bacillus thuringiensis ашылу тарихы.
Джо-эл Маргалит қос қанаттылардың бірінші патотипі В.

Жіктелу және филогения
Патшалық
Тип
Класс
Отряд
Тұқымдас
Туыс
Түр
Жалпы келісім бойынша түр бойынша ғылыми атаулар қиғаш жасумен
Жіктелу және филогения
Патшалық
Тип
Класс
Отряд
Тұқымдас
Туыс
Түр
Жалпы келісім бойынша түр бойынша ғылыми атаулар қиғаш жасумен

Жіктелу және филогения
Санат
Патшалық
Тип
Класс
Отряд
Тұқымдас
Туыс
Түр
Архейлер
Crenarchaeota
Thermoprotei
Sulfolobales
Sulfolobaceae
Sulfolobus
Sulfolobus acidocaldariaus
Жіктелу және филогения
Санат
Патшалық
Тип
Класс
Отряд
Тұқымдас
Туыс
Түр
Архейлер
Crenarchaeota
Thermoprotei
Sulfolobales
Sulfolobaceae
Sulfolobus
Sulfolobus acidocaldariaus

1. Bacillus thuringiensis ашылу тарихы.
В. Thuringiensis не өлі ағзалардың ыдыраған органикалық
1. Bacillus thuringiensis ашылу тарихы.
В. Thuringiensis не өлі ағзалардың ыдыраған органикалық

2. Bacillus thuringiensis-тің әсер ету механизымы.
1915 ж фляшениямен ауырған дернәсілдерін зерттегенде
2. Bacillus thuringiensis-тің әсер ету механизымы.
1915 ж фляшениямен ауырған дернәсілдерін зерттегенде

2. Bacillus thuringiensis-тің әсер ету механизымы.
Осы протоксин дернәсіл ішегіне түскеннен кейін
2. Bacillus thuringiensis-тің әсер ету механизымы.
Осы протоксин дернәсіл ішегіне түскеннен кейін

3. Bacillus thuringiensis бөлетін δ-эндотоксин түрлері.
B. thuringiensis кейбір штамдары тек қана
3. Bacillus thuringiensis бөлетін δ-эндотоксин түрлері.
B. thuringiensis кейбір штамдары тек қана

3. Bacillus thuringiensis бөлетін δ-эндотоксин түрлері.
Осы кристалдық нәруыздардың гендерің 2-і топқа
3. Bacillus thuringiensis бөлетін δ-эндотоксин түрлері.
Осы кристалдық нәруыздардың гендерің 2-і топқа

4. B. thuringiensis В-экзотоксині.
Вегетацияның белсенді кезеңінде B. thuringiensis кейбір штамдары β-экзотоксиндер
4. B. thuringiensis В-экзотоксині.
Вегетацияның белсенді кезеңінде B. thuringiensis кейбір штамдары β-экзотоксиндер

5. B. THURINGIENSIS ISRAELENSIS түршесі инсектицид ретінде.
В. T var israelensis
5. B. THURINGIENSIS ISRAELENSIS түршесі инсектицид ретінде.
В. T var israelensis

№ 6 дәріс
Бунақденелілерге төзімді трансгендік дақылдар.
1. Кіріспе.
2. Бунақденелілерге төзімді өсімдіктер линияларының
№ 6 дәріс
Бунақденелілерге төзімді трансгендік дақылдар.
1. Кіріспе.
2. Бунақденелілерге төзімді өсімдіктер линияларының

1. Кіріспе.
Бүкіл әлемде 30 миллион акрдан көп жерде осы бунақденелерге төзімді
1. Кіріспе.
Бүкіл әлемде 30 миллион акрдан көп жерде осы бунақденелерге төзімді

2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы.
Өсімдіктер жасушаларына бөгде ДНҚ енгізу үшін
2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы.
Өсімдіктер жасушаларына бөгде ДНҚ енгізу үшін
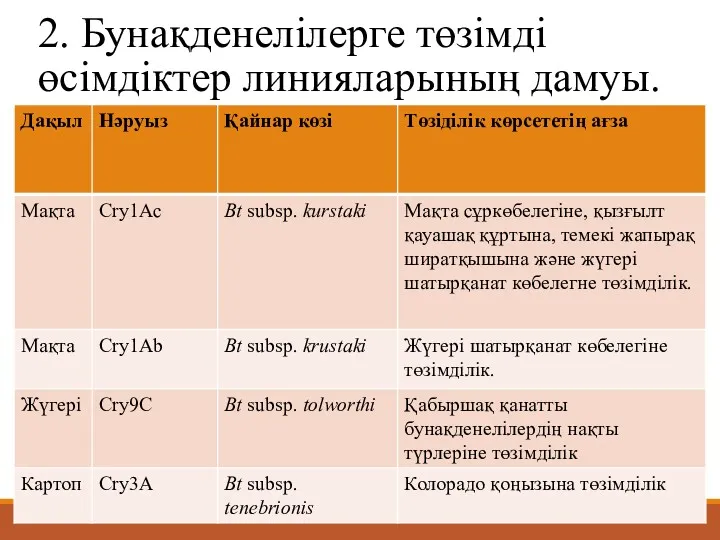
2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы.
2. Бунақденелілерге төзімді өсімдіктер линияларының дамуы.

3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
1992 жылы
3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
1992 жылы
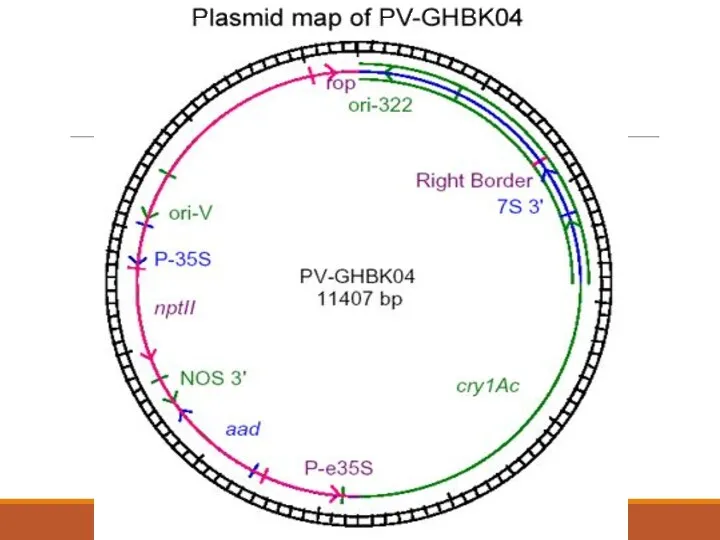

3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
Ori322/rop сайты.
3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
Ori322/rop сайты.

3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
P-35S, nptII
3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
P-35S, nptII

3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
Aad алтын
3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
Aad алтын

3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
7S3’ –
3. ГМ-Мақтасы 531 линиясының СRY1AC экспрессиясына сипаттама беру және дамуы.
7S3’ –

№ 7 дәріс
Агробактерия плазмидаларын вектор ретінде қолдану.
1. Жалпы сипаттамасы.
2. Зақымдау процесі.
№ 7 дәріс
Агробактерия плазмидаларын вектор ретінде қолдану.
1. Жалпы сипаттамасы.
2. Зақымдау процесі.

1. Жалпы сипаттамасы.
Agrobactera деген топырақта мекендейтін гарам теріс бактериялардың тобы. Олар
1. Жалпы сипаттамасы.
Agrobactera деген топырақта мекендейтін гарам теріс бактериялардың тобы. Олар

1. Жалпы сипаттамасы.
Ісік клеткаларында сау клеткалардан болмайтын опиндер деген жаңа класқа
1. Жалпы сипаттамасы.
Ісік клеткаларында сау клеткалардан болмайтын опиндер деген жаңа класқа
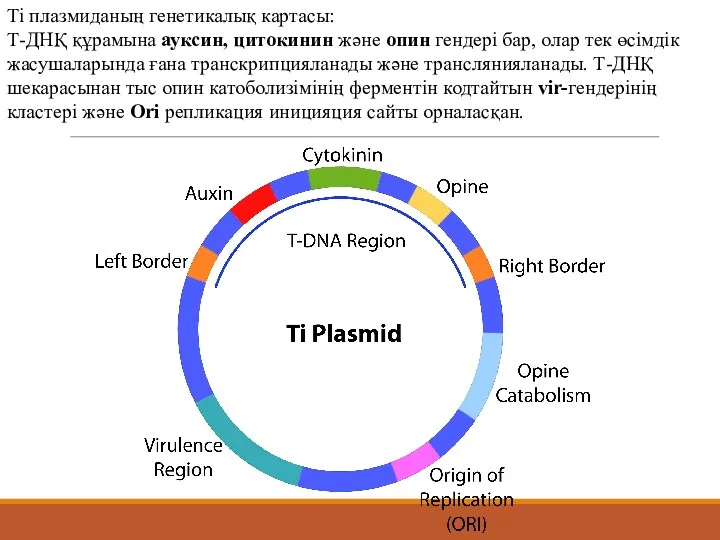
Ті плазмиданың генетикалық картасы:
Т-ДНҚ құрамына ауксин, цитокинин және опин гендері бар,
Ті плазмиданың генетикалық картасы:
Т-ДНҚ құрамына ауксин, цитокинин және опин гендері бар,

1. Жалпы сипаттамасы.
Vir гендері.
Vir A өсімдік заықмдалуына сезімтал. (Ацитсерингон)
Vir B Тасымалдау
1. Жалпы сипаттамасы.
Vir гендері.
Vir A өсімдік заықмдалуына сезімтал. (Ацитсерингон)
Vir B Тасымалдау

2. Зақымдау процесі.
Зақымдалған өсімдік ұлпалары ацитосерингонды бөліп шығарады. Оны A. tumefaciens
2. Зақымдау процесі.
Зақымдалған өсімдік ұлпалары ацитосерингонды бөліп шығарады. Оны A. tumefaciens

2. Зақымдау процесі.
Ары қарай Vir D1 кетіп Vir D2 қалып 5’
2. Зақымдау процесі.
Ары қарай Vir D1 кетіп Vir D2 қалып 5’
 Лимфатическая система человека
Лимфатическая система человека Введение в ортодонтию
Введение в ортодонтию Сбалансированное питание
Сбалансированное питание Тревожные расстройства
Тревожные расстройства Патофизиология экстремальных состояний
Патофизиология экстремальных состояний Жергілікті жансызданудың асқынулары
Жергілікті жансызданудың асқынулары Функциональный контроль при поражении спинного мозга, при детских церебральных параличах. (Лекция 4)
Функциональный контроль при поражении спинного мозга, при детских церебральных параличах. (Лекция 4) ГУЗ Сретенская ЦРБ. Информация о больнице
ГУЗ Сретенская ЦРБ. Информация о больнице Витамины. (Часть 2)
Витамины. (Часть 2) Кандидоз полости рта у детей
Кандидоз полости рта у детей Современные направления комплексного ухода за пациентами с язвенной болезнью 12-ти перстной кишки
Современные направления комплексного ухода за пациентами с язвенной болезнью 12-ти перстной кишки Пищевод. Рентгеноанатомия
Пищевод. Рентгеноанатомия Алгоритм диагностики, оказания скорой и неотложной медицинской помощи и порядок госпитализации пациентов с ОНМК
Алгоритм диагностики, оказания скорой и неотложной медицинской помощи и порядок госпитализации пациентов с ОНМК Первая помощь при отморожении
Первая помощь при отморожении Основы кожной пластики и кожные швы
Основы кожной пластики и кожные швы Антисептические и анальгетические лекарства
Антисептические и анальгетические лекарства Зубочелюстно-лицевая система – единый анатомо-функциональный комплекс. Особенности строения. Биодинамика движения нижней челюсти
Зубочелюстно-лицевая система – единый анатомо-функциональный комплекс. Особенности строения. Биодинамика движения нижней челюсти Ботулизм және тағамдық токсикоинфекция
Ботулизм және тағамдық токсикоинфекция Клинический случай. Хронический гломерулонефрит
Клинический случай. Хронический гломерулонефрит ВИЧ-инфекция. Основные моменты
ВИЧ-инфекция. Основные моменты Заболевания ЖКТ
Заболевания ЖКТ Ингибиторы контрольных точек
Ингибиторы контрольных точек Балалардағы туа пайда болған жүрек ақаулары
Балалардағы туа пайда болған жүрек ақаулары Этиопатогенетическая классификация дислалии
Этиопатогенетическая классификация дислалии Покрытие оболочками. Способ нанесения пленочных покрытий в дражировочном котле
Покрытие оболочками. Способ нанесения пленочных покрытий в дражировочном котле Стабилизация инъекционных растворов
Стабилизация инъекционных растворов Чесотка: клиника заболевания, диагностика, лечение
Чесотка: клиника заболевания, диагностика, лечение Амбулаториялық-емханалық мекеменің тоңазытқышында вакциналарды сақтаудың жалпы ережесі
Амбулаториялық-емханалық мекеменің тоңазытқышында вакциналарды сақтаудың жалпы ережесі