- Нервная регуляция

Содержание
- 2. Гуморальная – регуляция с помощью биологически активных веществ. Она более древняя. Нервная – регуляция с помощью
- 3. Обратная связь Для большинства механизмов регуляции функций в их структуру включается обратная связь. С ее помощью
- 4. Точность «адресата». Рефлекторный принцип регуляции. Отличие нейронной регуляции от гуморальной.
- 5. Рефлекторный принцип организации нейронной регуляции Рефлексом называется стереотипная реакция организма или его отдельных органов на сенсорный
- 6. Обеспечивается точность регуляции, в основе которой лежит: а) получение информации от органа, б) ее анализ в
- 7. Сенсорные системы (органы чувств). Их, как минимум, 8! Начинаются они специализированными нервными окончаниями – рецепторами. Получение
- 8. Первичночувст-вующие рецепторы Это различного типа нервные окончания. В них под влиянием раздражителей возникает ПД. При возникновении
- 9. Вторично чувствующие рецепторы (ВЧР) ВЧР – более сложные структуры, которые непосредственно воспринимают действие раздражителя и трансформируют
- 10. Нейроны Нейрон имеет тело и различного типа отростки (аксон и дендриты). 1 - мультиполярный нейрон; 2
- 11. ПП – от –60 мВ до –90 мВ, что зависит величиной тела нейрона. Аксонный холмик (начало
- 12. Глиальные клетки 90% всех клеток ЦНС – глиальные клетки. Различаю 4 типа клеток (рис.) На рис.
- 13. Глиальные клетки: Астроциты Резорбция ряда медиаторов. Временное поглощение некоторых ионов (например, К+) из межклеточной жидкости в
- 14. Астроцит «Лапки» астроцитов охватывают кровеносные капилляры и нейроны. Астроцит создает преграду между нервом и кровеносным капилляром,
- 15. Физиология нейронов 1 – ядро, 2 – дендриты, 3 – тело, 4 – аксонный холмик, 5
- 16. Абсолютный рефрактерный период примерно такой же, как и длительность ПД. В нейроне абсолютный рефрактерный период около
- 17. Поверхность мембраны нервного волокна пропорциональна его диаметру, а поперечное сечение волокна возрастает пропорционально квадрату диаметра. При
- 18. Синапсы ЦНС В нервной системы нейроны за счет отростков контактируют между собой, образуя синапсы. На каждом
- 19. 1. Амины (ацетилхолин, норадреналин, адреналин, дофамин, серотонин). 2. Аминокислоты (глицин, глутамин, аспарагиновая, ГАМК и ряд др.).
- 20. В нервном окончании может выделяться один либо несколько медиаторов. Ионотропные медиаторы после взаимодействия с рецепторами постсинаптической
- 21. ВОЗБУЖДЕНИЕ В ЦНС при передачи возбуждения (для развития ПД) через синапсы происходит явление суммации. В результат
- 22. Виды суммации в ЦНС В ЦНС различают (см. рис.) два вида суммации: Временная суммация – как
- 23. Нексус Мембраны некоторых нейронов (их немного) соприкасаются и ПД передается от одного к другому без медиатора.
- 24. ТОРМОЖЕНИЕ В ЦНС большинство синапсов не передают возбуждение, а тормозят этот процесс. Различают два механизма торможения
- 25. Развитие гиперполяризации на постсинаптической мембране тормозного синапса Как правило, торможение обусловлено развитием не деполяризации, а гиперполяризации
- 26. Электро энцефалограмма (ЭЭГ) А - при открытых глазах (видны по преимуществу мелкой амплитуды β-волны); Б -
- 27. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ а – конвергенция (схождение), когда за счет тормозных нейронов (не показаны) количество возбужденных
- 28. При одновременном возбуждении нескольких нервных центров, один из очагов может стать доминантным, главенствующим. В результате к
- 29. Это системы нервных клеток, которые не выполняют специфические функции (рефлексы), они регулируют функцию самой нервной системы,
- 30. Ретикулярная формация (РФ) ствола мозга В стволе мозга разбросаны нейроны, которые объединяются в единую сеть (rete).
- 32. Скачать презентацию
 Сегментарное строение спинного мозга. Простая и сложная рефлекторные дуги
Сегментарное строение спинного мозга. Простая и сложная рефлекторные дуги Экстрагенитальды патология және жүктілік
Экстрагенитальды патология және жүктілік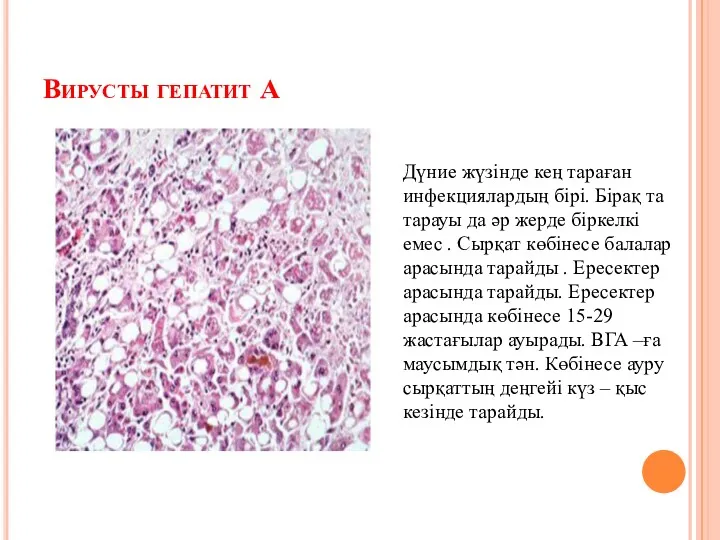 Вирусты гепатит А
Вирусты гепатит А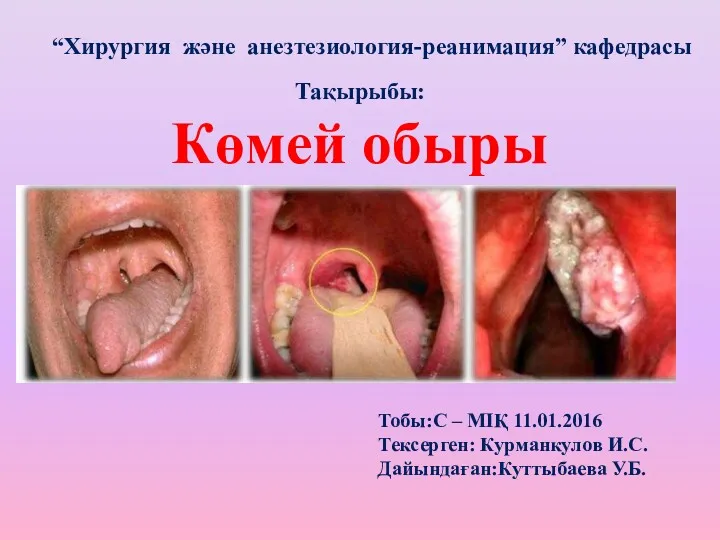 Көмей обыры
Көмей обыры Синдром длительного сдавления (СДС)
Синдром длительного сдавления (СДС) Практические подходы к применению лекарств при беременности
Практические подходы к применению лекарств при беременности Слайд-лекция №16. Противоаритмические средства
Слайд-лекция №16. Противоаритмические средства Аневризма сердца
Аневризма сердца Наименование вида ПАВ
Наименование вида ПАВ Физиотерапия при бронхолёгочной патологии
Физиотерапия при бронхолёгочной патологии Туберкулезный менингит
Туберкулезный менингит Что такое гепатит В?
Что такое гепатит В? АИВ инфекциясы
АИВ инфекциясы Медицина Западной Европы эпохи Возрождения (XV – XVII вв.). Лекция 5
Медицина Западной Европы эпохи Возрождения (XV – XVII вв.). Лекция 5 Некроз. Атрофия
Некроз. Атрофия Общая фармакология
Общая фармакология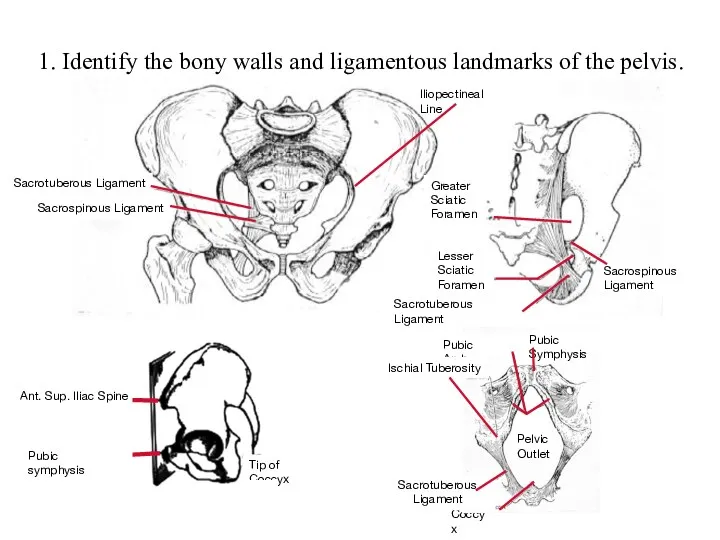 Identify the bony walls and ligamentous landmarks of the pelvis
Identify the bony walls and ligamentous landmarks of the pelvis Основы эхокардиографии
Основы эхокардиографии ҚР электронды денсаулық сақтау саласының 2013-2020 жылдарға арналған концепциясы концепциясы
ҚР электронды денсаулық сақтау саласының 2013-2020 жылдарға арналған концепциясы концепциясы Анафилаксия. Рекомендации ФАР
Анафилаксия. Рекомендации ФАР Лимфаайналының бұзылуы турлері. Лимфостаз. Лимфа тамырлардағы тробмоз және эмболия
Лимфаайналының бұзылуы турлері. Лимфостаз. Лимфа тамырлардағы тробмоз және эмболия Осанка – залог здоровья
Осанка – залог здоровья Organizacja pracy rejestracji medycznej
Organizacja pracy rejestracji medycznej Сердечно-легочная реанимация
Сердечно-легочная реанимация Роль структур ЦНС в регуляции физиологических функций
Роль структур ЦНС в регуляции физиологических функций Изменения СОПР при дерматозах
Изменения СОПР при дерматозах Первая помощь при растяжении связок, вывихах суставов, переломах костей
Первая помощь при растяжении связок, вывихах суставов, переломах костей Веселые предметы. Дыхательная гимнастика для детей
Веселые предметы. Дыхательная гимнастика для детей