- Роль структур ЦНС в регуляции физиологических функций

Содержание
- 2. Спинной мозг Спинной мозг - низший и филогенетически самый древний отдел центральной нервной системы. Центральная нервная
- 3. Спинной мозг со спинномозговыми нервами
- 4. Задние корешки образованы аксонами афферентных нейронов, тела которых лежат вне ЦНС, а дендриты этих нейронов связаны
- 5. Спинной мозг осуществляет помимо головного мозга собственную интегративную функцию, поскольку участвует в сложных процессах координации функций
- 7. Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки (промежуточные нейроны), выполняющие связующую функцию
- 8. Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга и являются фоновоактивными. Проводящие пути
- 9. Нисходящие пути связывают отделы головного мозга с моторными или вегетативными эфферентными нейронами. Цереброспинальные нисходящие пути начинаются
- 10. Спинной мозг имеет двустороннюю связь с головным мозгом, и поэтому кроме рефлекторной функции выполняет проводниковую, благодаря
- 12. Подобная организация позволяет реализовывать все двигательные рефлексы тела, диафрагмы, мочеполовой системы и прямой кишки, терморегуляции, сосудистые
- 13. Сложной формой рефлекторной деятельности спинного мозга является рефлекс, реализующий произвольное движение. В основе реализации произвольного движения
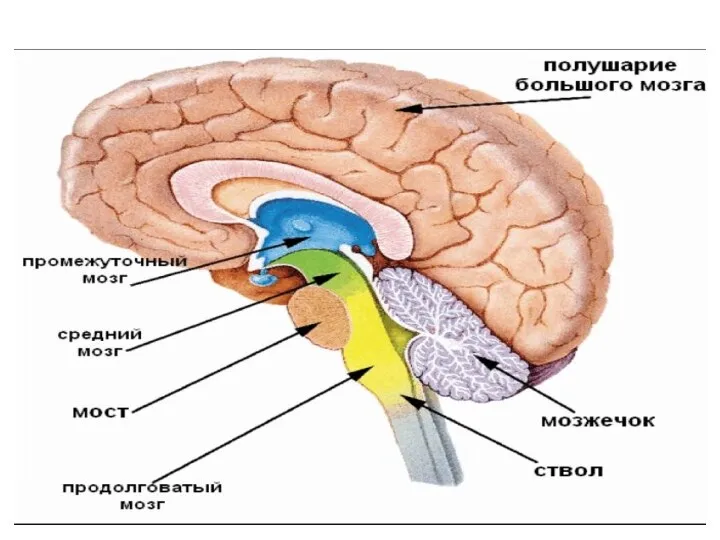
- 14. Головной мозг 1. Передний мозг: - Кора больших полушарий; - Базальные ганглии: - система бледного шара
- 19. В функциональном отношении мозг можно разделить на несколько отделов: 1) передний мозг, состоящий из конечного мозга
- 20. Продолговатый мозг и мост Продолговатый мозг- непосредственное продолжение вверх спинного мозга, одновременно является частью ствола мозга
- 22. Продолговатый мозг выполняет две функции (рефлекторную и проводниковую). Рефлекторная функция осуществляется за счет: 1 - простых
- 23. Проводниковые функции п.м. и моста проходят восх. и нисх. пути с.м.: пирамидный, руброспинальный, спиноталамический, заканчиваются пути
- 24. Собственные функции п.м. и ствола В продолговатом мозге находятся рефлекторные центры: дыхательный , сердечно - сосудистый
- 25. Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов
- 27. Опыт об участии ствола мозга в обменных процессах Опыт К.Бернара – при локальном уколе в дно
- 28. Состояние тонуса мышц бульбарного животного и схема Функции продолговатого мозга были изучены на бульбарных животных, у
- 29. Структура среднего мозга 1. Крышка мозга (дорзальный отдел); 2. Ножки мозга (вентральный отдел). Животные с перерезкой
- 30. Ядра нижних холмиков играют существенную роль в реализации функции слуха и формировании сложных рефлексов в ответ
- 31. Средний мозг выполняет следующие функции: центр ориентировочного рефлекса центр позы центр обработки первичной информации (зрение, слух)
- 32. Проводникова функция с.м. осущ. через ножки мозга, по которым в восходящем направлении несут импульсы волокна спиноталамического
- 33. Статические –определяют поддержание удобной позы в пространстве при лежании, сидении и стоянии животных за счет мышечного
- 34. Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и ядра черепных (глазодвигательного и блокового)
- 35. Рефлексы позы Роль заднего мозга регуляции двигательных функций: 1) анализ сенсорной информации от рецепторов кожи и
- 36. Ориентировочный рефлекс Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. За счет нейронов ядер верхнего и нижнего
- 37. Свободное падение кошки из положения на спину (по Магнусу, 1924). Описанные выше установочные рефлексы легко наблюдать
- 38. Выпрямительный рефлекс Выпрямительные рефлексы можно разделить на несколько групп: Лабиринтные реакции Рефлексы с туловища на голову
- 39. Мезенцефальный организм (нарушение связи между средним и промежуточным мозгом) имеет нормальный мышечный тонус, сохраняются все тонические
- 40. Функции черной субстанции Черная субстанция ограничивает влияние норадренергической системы, активирует продукцию оксида азота в головном мозге,
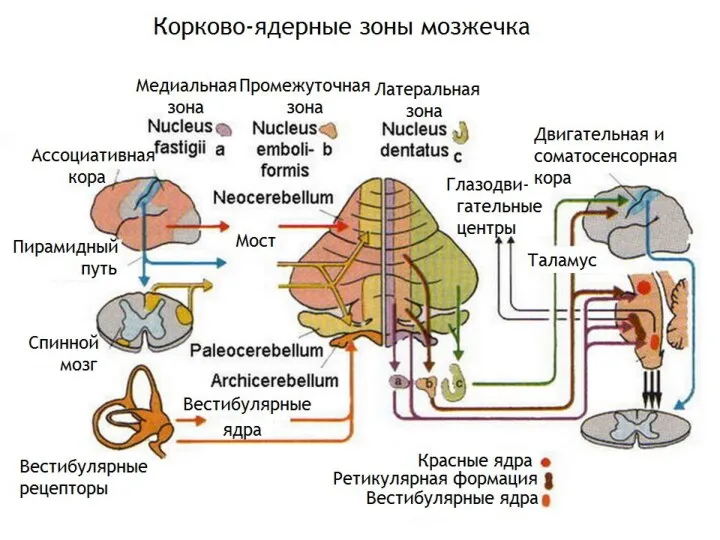
- 41. Мозжечок Состоит из двух полушарий, червя (между ними), трех пар ножек, образованных пучками нервных волокон. Выделяют
- 44. Строение трех слоев коры мозжечка и их нейронных связей
- 46. Функции мозжечка связаны с организацией двигательных актов и регуляцией вегетативных функций. При осуществлении двигательных актов перемещающиеся
- 47. Последствия нарушения функций мозжечка: атония (нарушения тонуса склетных мышц); астения (повышения утомляемости); астазия (качание, дрожжение); асинергия
- 48. ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА ТРИАДА ЛЮЧИАНИ: атония, астазия, астения ТРИАДА ШАРКО: нистагм, тремор, скандированная речь АТАКСИЯ (пьяная
- 49. Взаимодействие мозжечка и коры большого мозга. Это взаимодействие организовано соматотопически. Функционально мозжечок может оказывать облегчающее, тормозящее
- 50. Функции мозжечка Статические и статокинетические рефлексы; Регулирует вегетативные функции.
- 53. Роль мозжечка в регуляции тонуса мышц Регуляция мышечного тонуса, обеспечивающего поддержание естественной позы (равновесия) осуществляется преимущественно
- 54. передний мозг- состоит из конечного мозга (больших полушарий) и промежуточного мозга
- 55. Промежуточный мозг - часть переднего отдела ствола мозга. Основными образованиями промежуточного мозга являются зрительные бугры (таламус)
- 56. Ядра таламуса и гипоталамуса
- 58. Переключательную функцию в сенсорных ядрах обеспечивают таламокортикальные (релейные) нейроны, длинный аксон которых непосредственно иннервируют нейроны сенсорной
- 59. Неспецифические ядра таламуса – более древняя область, включающая интраламинарную ядерную группу и ретикулярное ядро. Неспецифические ядра
- 60. Гипоталамус. Содержит 50 пар ядер, которые объединены в следующие группы: Преоптическую. Образуют функционально-гипофизарную область, их нейроны
- 61. Функциональное значение гипоталамуса. 1. Высший подкорковый автономный центр. 2. Регулирует биоритмы - благодаря центрам сна и
- 62. Участие гипоталамуса в регуляции поведенческих реакций . Экспериментальные исследования , в которых использовались раздражения определенных зон
- 63. По А. Р. Лурия, реальным анатомическим и функциональным образованием, включенным в реализацию двигательного акта, помимо собственно
- 65. Пирамидная система, пирамидный путь, система нервных структур, участвующих в сложной и тонкой координации двигательных актов. У
- 66. Экстрапирамидная система осуществляет непроизвольную регуляции и координацию движений, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений
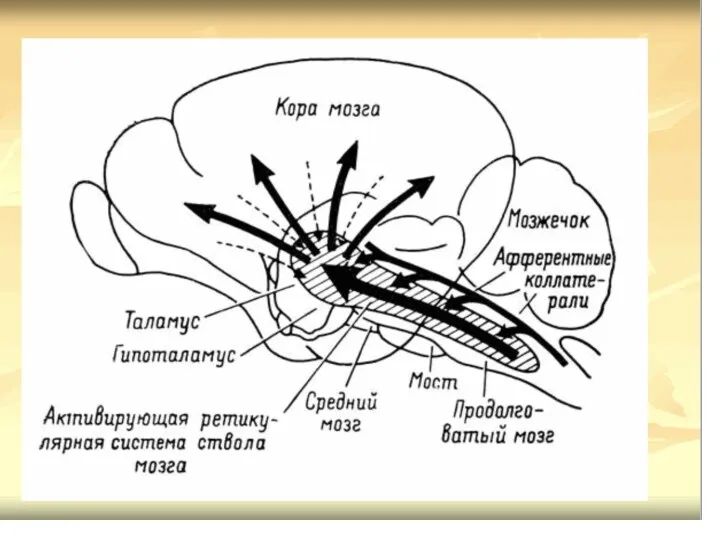
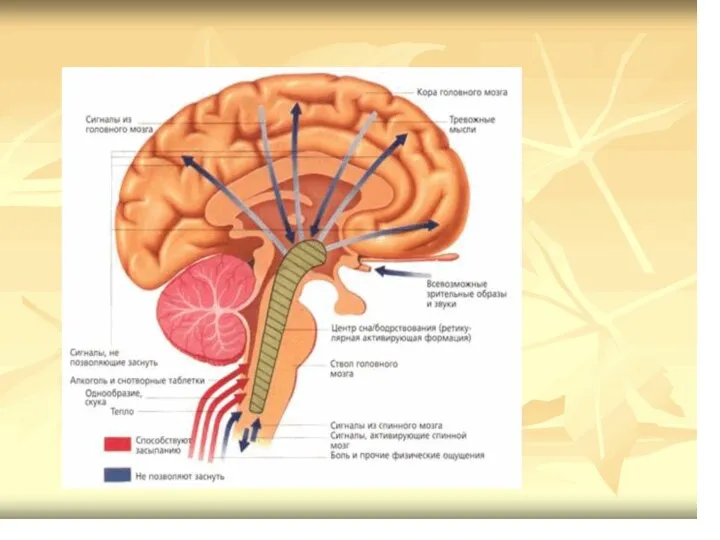
- 69. РФ включает: Латеральные ретикулярные ядра; Ретикулярные вентральные ядра; Ядра покрышки (Бехтерева); Парамедиальные ретикулярные ядра; ретикулярные гигантоклеточные
- 70. Ретикулярная формация (РФ) Функциональные особенности ретикулярных нейронов. Полисенсорная конвергенция: принимают коллатерали от нескольких сенсорных путей, идущих
- 71. Проводниковые функции: Афф.пути представлены спиноретикулярным, несущим импульсы от температурных, болевых рецепторов; Церебеллоретикулярный корково-подкорково-ретикулярный путь( от коры,
- 72. Восходящие и нисходящие влияния ретикулярной формации
- 73. РФ восходящее влияние Импульсы ретикулярных нейронов продолговатого мозга (гигантеклеточное, латеральное и вентральное ретикулярные ядра), моста (особенно
- 74. РФ нисходящее влияние Структуры р.ф. ствола мозга оказывают облегчающее и тормозное влияние на рефлекторные реакции спинного
- 78. Нисходящие влияние на тонус мышц и формирование позы РФ оказывает через латеральный и медиальный ретикулоспинальные тракты.
- 80. Лимбическая система Лимбическая система – совокупность структур промежуточного, конечного и среднего мозга, обеспечивающая эмоционально – мотивационные
- 81. Функции лимбической системы. Получая инфу о внешней среде и внутренней среде организма, лимбическая система после сравнения
- 82. Круг Пайпетца (гиппокамп - сосцевидные тела - передние ядра таламуса - кора поясной извилины - парагиппокампова
- 83. Поддерживает гомеостаз и регулирует работу внутренних органов. При поражении - нарушение обменных процессов в миокарде, нарушение
- 84. Функции новой коры Новая кора – слой серого вещества общей площадью 1100 – 1900 см2, покрывающий
- 85. Слои коры больших полушарий Полушария головного мозга покрыты серым веществом или корой, толщина которой в разных
- 86. Функциональная организация коры Функциональная единица коры - вертикальная колонка диаметром около 500 мкм - макромодуль Колонка
- 87. Физиологическое значение коры: 1. Обеспечение высшей нервной деятельности. 2. Обработка сенсорной информации. 3. Формирование двигательных команд.
- 88. Функции новой коры Новая кора – слой серого вещества общей площадью 1100 – 1900 см2, покрывающий
- 89. Функции клеток коры больших полушарий Основная информация в кору поступает по специфическим афферентным проводящим путям, заканчивающимся
- 91. Кортикальные связи 1 – ассоциативные (связывают зоны коры в пределах одного полушария); 2 – комиссуральные (связывают
- 92. Вторичная соматосенсорная зона расположена в стенке боковой борозды, на границе ее пересечения с центральной бороздой. Ассоциативные
- 94. Карта локализации функций в коре головного мозга по Бродману Вкусовой центр Чувствительные центры Зрительные центры Слуховые
- 95. Чувствительные центры коры располагаются в области задней, центральной извилины (поля 1, 3, 5, 7 по Бродману),
- 96. Корковый зрительный центр находится в затылочной доле (поля 17, 18, 19), воспринимает и анализирует информацию от
- 97. Проекции частей тела в соматосенсорной зоне коры больших полушарий
- 98. Корковый обонятельный центр локализован в крючковидной извилине и в аммоновом роге. Корковый вкусовой центр находится в
- 99. Ассоциативные зоны коры Верхнетеменная область связана с восприятием сложных форм тактильной чувствительности и имеет отношение к
- 100. Корковые двигательные центры (в области передней центральной извилины) содержит в 5 слое коры пирамидные клетки Беца,
- 101. ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ Управление врожденными поведенческими реакциями при помощи накопленного опыта Согласование внешних и внутренних мотиваций
- 102. Центр проприоцептивного гнозиса (в верхней теменной дольке) обеспечивает восприятие Локализация функций в коре БП импульсов от
- 103. Функциональная асимметрия мозга
- 104. Принципы межполушарных взаимодействий Параллельная деятельность – информация параллельно обрабатывается в обоих полушариях с использованием всех механизмов.
- 105. Функции полушарий мозга Левое полушарие Специализируется на выполнении вербальных символических функций, формировании абстрактного мышления, способности к
- 106. Функции полушарий мозга Правое полушарие Быстрее обрабатывает информацию, участвует в формировании конкретного мышления, отрицательных эмоций, различение
- 107. Функции полушарий мозга
- 108. Межполушарные различия ЛЕВОЕ ПОЛУШАРИЕ ПРАВОЕ ПОЛУШАРИЕ Лучше узнаются стимулы Словесные Несловесные Легко различимые Трудно различимые Знакомые
- 109. Понятия «леворукость» и «левшество» синонимами не являются. Леворукость – это термин, отражающий предпочтение, активное использование левой
- 110. Генетическое левшество. Ученые Оксфордского университета обнаружили ген ЛРРТМ1, устанавливающий «правила» работы правого и левого полушарий мозга.
- 111. «Компенсаторное" левшество связанно с каким-либо поражением мозга, чаще - его левого полушария. Поскольку деятельность правой руки
- 112. Псевдолеворукость К определенному возрасту (окончательно примерно к 5 годам) у ребенка какое-то из полушарий формируется как
- 113. Скрытое левшество Наблюдается при смене доминирующего полушария. Момент смены является тем критическим периодом, когда основные функции
- 114. Электроэнцефалография. Электроэнцефалография - это метод регистрации суммарной биоэлектрической активности мозга с поверхности головы. Электрическая активность является
- 117. Роль спинного мозга в регуляции функций организма: вегетативные и соматические центры и их значение. Соматические нейроны
- 119. Скачать презентацию

Спинной мозг
Спинной мозг - низший и филогенетически самый древний отдел центральной
Спинной мозг
Спинной мозг - низший и филогенетически самый древний отдел центральной
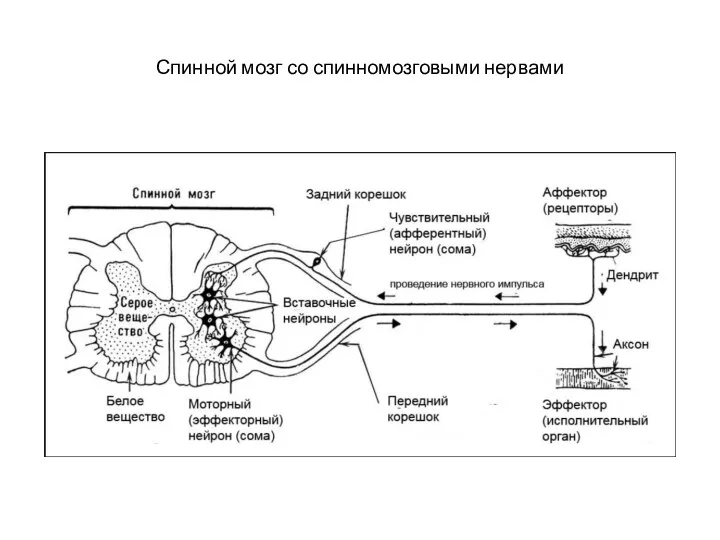
Спинной мозг со спинномозговыми нервами
Спинной мозг со спинномозговыми нервами

Задние корешки образованы аксонами афферентных нейронов, тела которых лежат вне ЦНС,
Задние корешки образованы аксонами афферентных нейронов, тела которых лежат вне ЦНС,

Спинной мозг осуществляет помимо головного мозга собственную интегративную функцию, поскольку участвует
Спинной мозг осуществляет помимо головного мозга собственную интегративную функцию, поскольку участвует

Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки
Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки

Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга
Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга

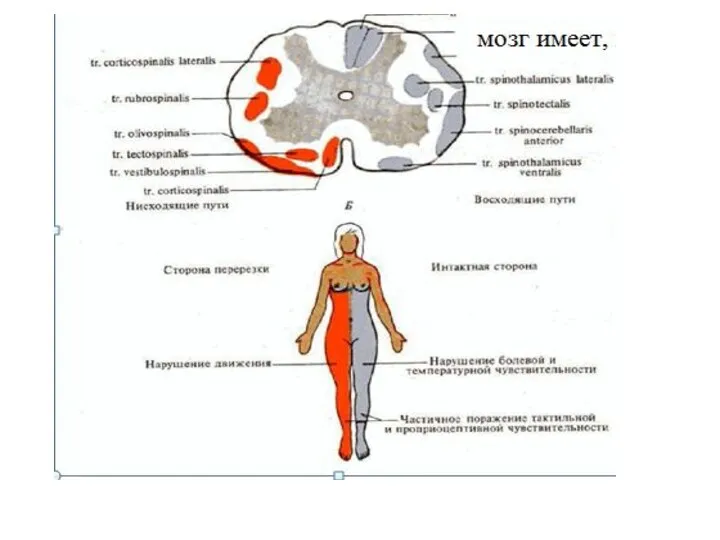
Нисходящие пути связывают отделы головного мозга с моторными или вегетативными эфферентными
Нисходящие пути связывают отделы головного мозга с моторными или вегетативными эфферентными

Спинной мозг имеет двустороннюю связь с головным мозгом, и поэтому кроме
Спинной мозг имеет двустороннюю связь с головным мозгом, и поэтому кроме
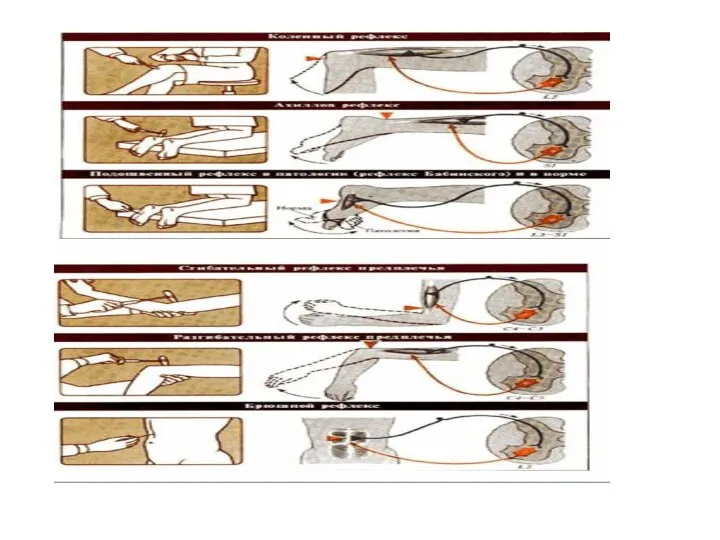

Подобная организация позволяет реализовывать все двигательные рефлексы тела, диафрагмы, мочеполовой системы
Подобная организация позволяет реализовывать все двигательные рефлексы тела, диафрагмы, мочеполовой системы

Сложной формой рефлекторной деятельности спинного мозга является рефлекс, реализующий произвольное движение.
Сложной формой рефлекторной деятельности спинного мозга является рефлекс, реализующий произвольное движение.

Головной мозг
1. Передний мозг:
- Кора больших полушарий;
- Базальные ганглии:
Головной мозг
1. Передний мозг:
- Кора больших полушарий;
- Базальные ганглии:

В функциональном отношении мозг можно разделить на несколько отделов:
1) передний мозг, состоящий
В функциональном отношении мозг можно разделить на несколько отделов:
1) передний мозг, состоящий

Продолговатый мозг и мост
Продолговатый мозг- непосредственное продолжение вверх спинного мозга, одновременно
Продолговатый мозг и мост
Продолговатый мозг- непосредственное продолжение вверх спинного мозга, одновременно
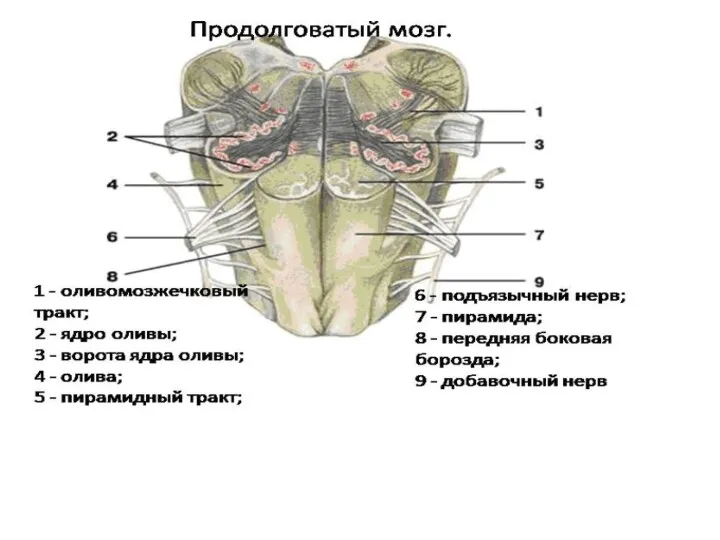

Продолговатый мозг выполняет две функции (рефлекторную и проводниковую). Рефлекторная функция осуществляется
Продолговатый мозг выполняет две функции (рефлекторную и проводниковую). Рефлекторная функция осуществляется

Проводниковые функции п.м. и моста
проходят восх. и нисх. пути с.м.: пирамидный,
Проводниковые функции п.м. и моста
проходят восх. и нисх. пути с.м.: пирамидный,

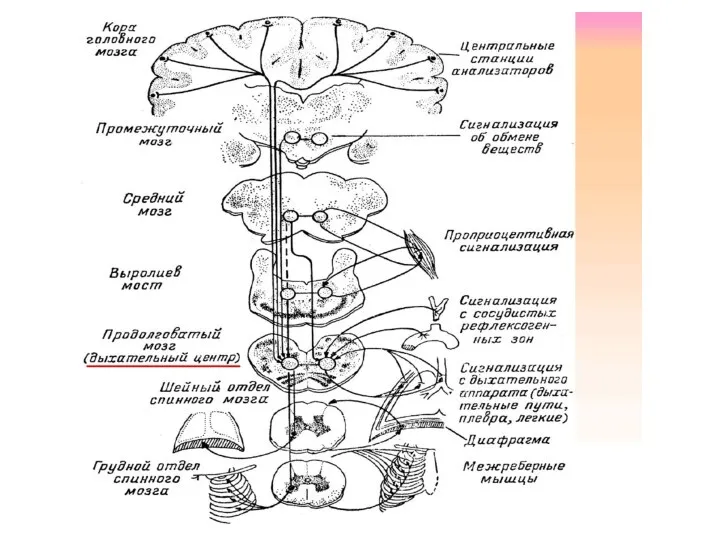
Собственные функции п.м. и ствола
В продолговатом мозге находятся рефлекторные центры: дыхательный
Собственные функции п.м. и ствола
В продолговатом мозге находятся рефлекторные центры: дыхательный

Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются
Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются

Опыт об участии ствола мозга в обменных процессах
Опыт К.Бернара – при
Опыт об участии ствола мозга в обменных процессах
Опыт К.Бернара – при
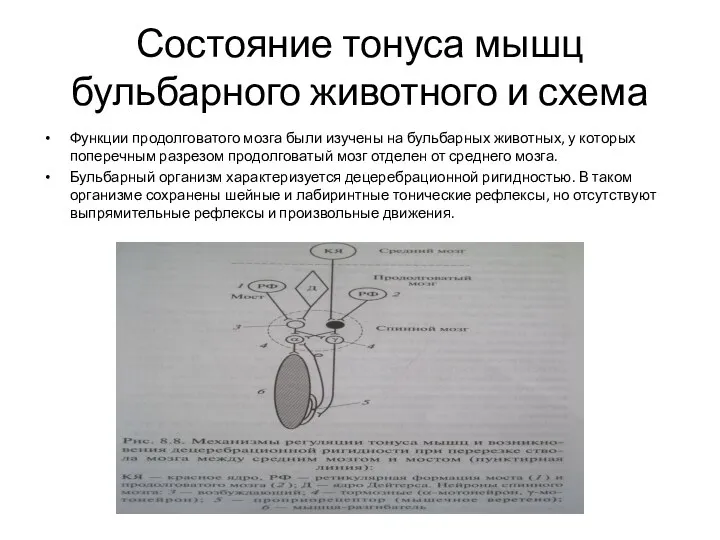
Состояние тонуса мышц бульбарного животного и схема
Функции продолговатого мозга были изучены
Состояние тонуса мышц бульбарного животного и схема
Функции продолговатого мозга были изучены

Структура среднего мозга
1. Крышка мозга (дорзальный отдел);
2. Ножки мозга (вентральный отдел).
Структура среднего мозга
1. Крышка мозга (дорзальный отдел);
2. Ножки мозга (вентральный отдел).

Ядра нижних холмиков играют существенную роль в реализации функции слуха и
Ядра нижних холмиков играют существенную роль в реализации функции слуха и

Средний мозг выполняет следующие функции:
центр ориентировочного рефлекса
центр позы
центр обработки первичной информации
Средний мозг выполняет следующие функции:
центр ориентировочного рефлекса
центр позы
центр обработки первичной информации

Проводникова функция с.м. осущ. через ножки мозга, по которым в восходящем
Проводникова функция с.м. осущ. через ножки мозга, по которым в восходящем

Статические –определяют поддержание удобной позы в пространстве при лежании, сидении и
Статические –определяют поддержание удобной позы в пространстве при лежании, сидении и

Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и
Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и
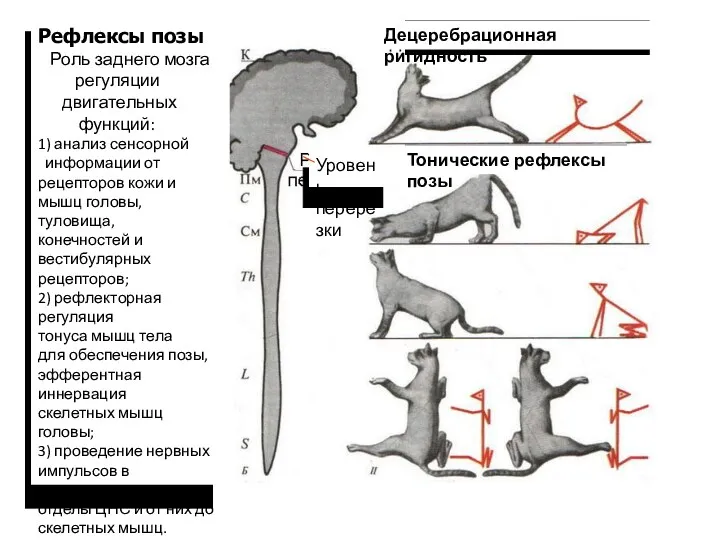
Рефлексы позы
Роль заднего мозга
регуляции
двигательных
функций:
1) анализ сенсорной
информации
Рефлексы позы
Роль заднего мозга
регуляции
двигательных
функций:
1) анализ сенсорной
информации

Ориентировочный рефлекс
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. За счет нейронов
Ориентировочный рефлекс
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. За счет нейронов
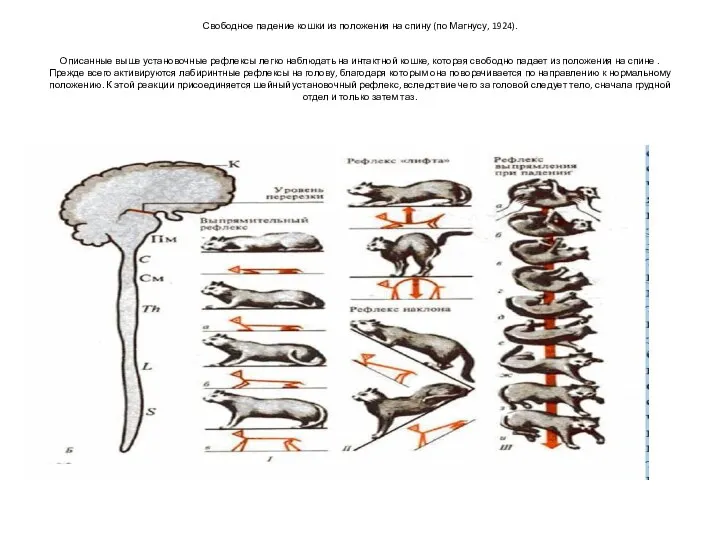
Свободное падение кошки из положения на спину (по Магнусу, 1924).
Описанные выше
Свободное падение кошки из положения на спину (по Магнусу, 1924). Описанные выше

Выпрямительный рефлекс
Выпрямительные рефлексы можно разделить на несколько групп:
Лабиринтные реакции
Рефлексы с туловища
Выпрямительный рефлекс
Выпрямительные рефлексы можно разделить на несколько групп:
Лабиринтные реакции
Рефлексы с туловища

Мезенцефальный организм (нарушение связи между средним и промежуточным мозгом) имеет нормальный
Мезенцефальный организм (нарушение связи между средним и промежуточным мозгом) имеет нормальный

Функции черной субстанции
Черная субстанция ограничивает влияние норадренергической системы, активирует продукцию оксида
Функции черной субстанции
Черная субстанция ограничивает влияние норадренергической системы, активирует продукцию оксида

Мозжечок
Состоит из двух полушарий, червя (между ними), трех пар ножек, образованных
Мозжечок
Состоит из двух полушарий, червя (между ними), трех пар ножек, образованных
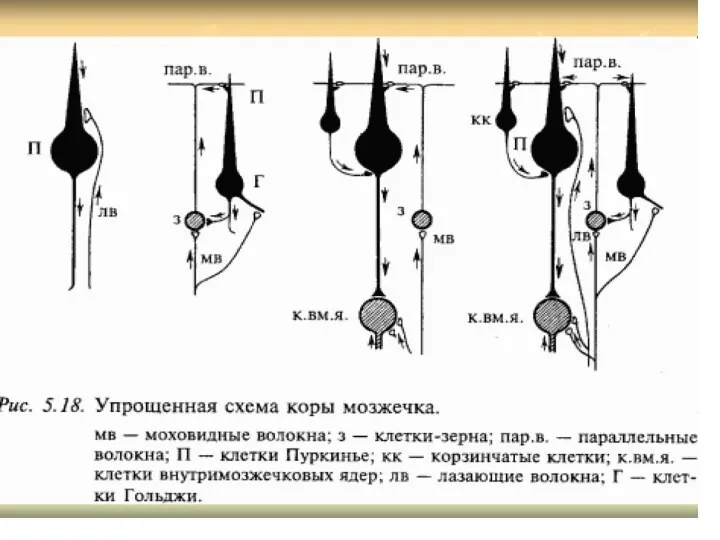
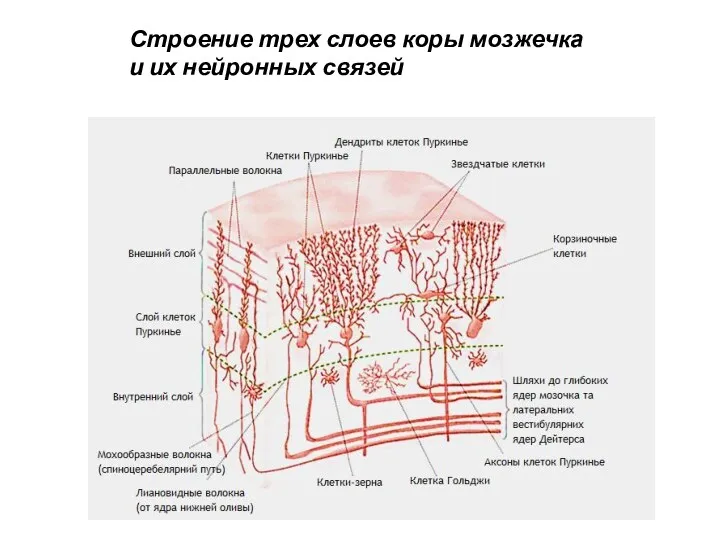
Строение трех слоев коры мозжечка и их нейронных связей
Строение трех слоев коры мозжечка и их нейронных связей


Функции мозжечка связаны с организацией двигательных актов и регуляцией вегетативных функций.
Функции мозжечка связаны с организацией двигательных актов и регуляцией вегетативных функций.

Последствия нарушения функций мозжечка:
атония (нарушения тонуса склетных мышц);
астения (повышения утомляемости);
астазия (качание,
Последствия нарушения функций мозжечка:
атония (нарушения тонуса склетных мышц);
астения (повышения утомляемости);
астазия (качание,

ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
ТРИАДА ЛЮЧИАНИ: атония,
астазия,
астения
ТРИАДА ШАРКО: нистагм,
ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
ТРИАДА ЛЮЧИАНИ: атония,
астазия,
астения
ТРИАДА ШАРКО: нистагм,

Взаимодействие мозжечка и коры большого мозга. Это взаимодействие организовано соматотопически. Функционально
Взаимодействие мозжечка и коры большого мозга. Это взаимодействие организовано соматотопически. Функционально

Функции мозжечка
Статические и статокинетические рефлексы;
Регулирует вегетативные функции.
Функции мозжечка
Статические и статокинетические рефлексы;
Регулирует вегетативные функции.
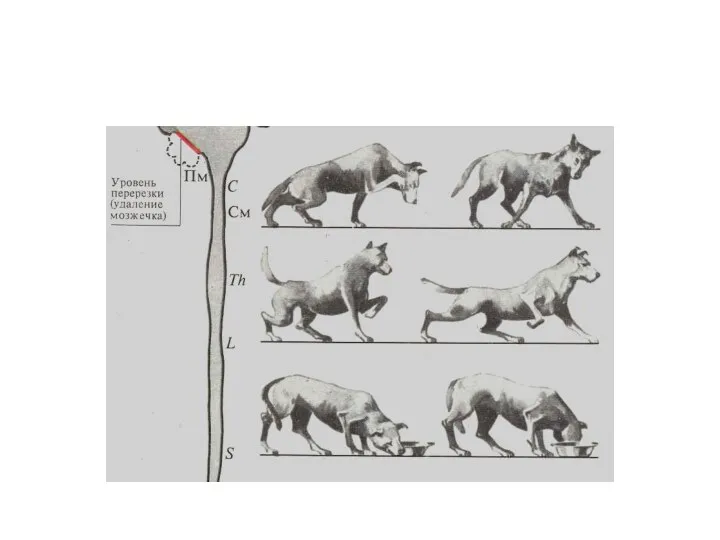


Роль мозжечка в регуляции тонуса мышц
Регуляция мышечного тонуса, обеспечивающего поддержание естественной
Роль мозжечка в регуляции тонуса мышц
Регуляция мышечного тонуса, обеспечивающего поддержание естественной

передний мозг- состоит из конечного мозга (больших полушарий) и промежуточного мозга
передний мозг- состоит из конечного мозга (больших полушарий) и промежуточного мозга

Промежуточный мозг
- часть переднего отдела ствола мозга. Основными образованиями промежуточного
Промежуточный мозг - часть переднего отдела ствола мозга. Основными образованиями промежуточного
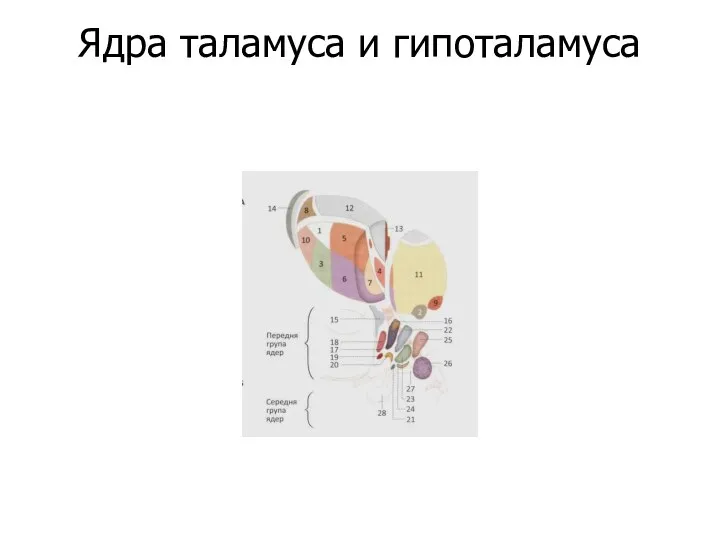
Ядра таламуса и гипоталамуса
Ядра таламуса и гипоталамуса
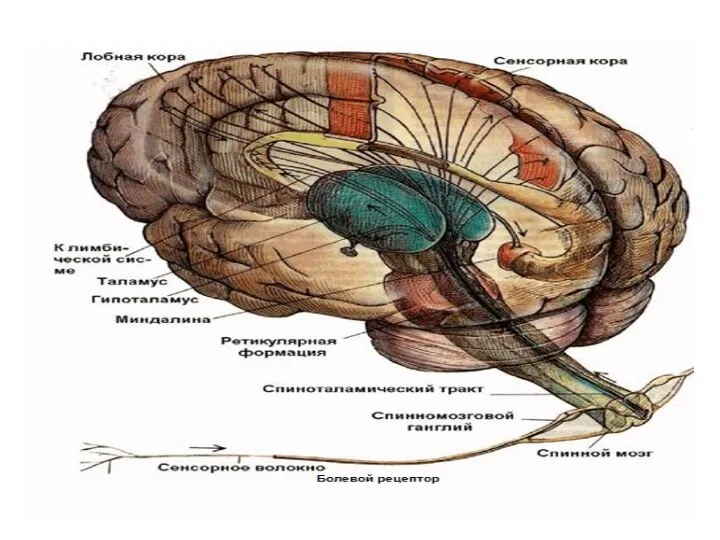

Переключательную функцию в сенсорных ядрах обеспечивают таламокортикальные (релейные) нейроны, длинный аксон
Переключательную функцию в сенсорных ядрах обеспечивают таламокортикальные (релейные) нейроны, длинный аксон

Неспецифические ядра таламуса – более древняя область, включающая интраламинарную ядерную группу
Неспецифические ядра таламуса – более древняя область, включающая интраламинарную ядерную группу

Гипоталамус.
Содержит 50 пар ядер, которые объединены в следующие группы:
Преоптическую. Образуют функционально-гипофизарную
Гипоталамус.
Содержит 50 пар ядер, которые объединены в следующие группы:
Преоптическую. Образуют функционально-гипофизарную

Функциональное значение гипоталамуса.
1. Высший подкорковый автономный центр.
2. Регулирует биоритмы - благодаря
Функциональное значение гипоталамуса.
1. Высший подкорковый автономный центр.
2. Регулирует биоритмы - благодаря

Участие гипоталамуса в регуляции поведенческих реакций .
Экспериментальные исследования , в которых
Участие гипоталамуса в регуляции поведенческих реакций .
Экспериментальные исследования , в которых

По А. Р. Лурия, реальным анатомическим и функциональным образованием, включенным в
По А. Р. Лурия, реальным анатомическим и функциональным образованием, включенным в
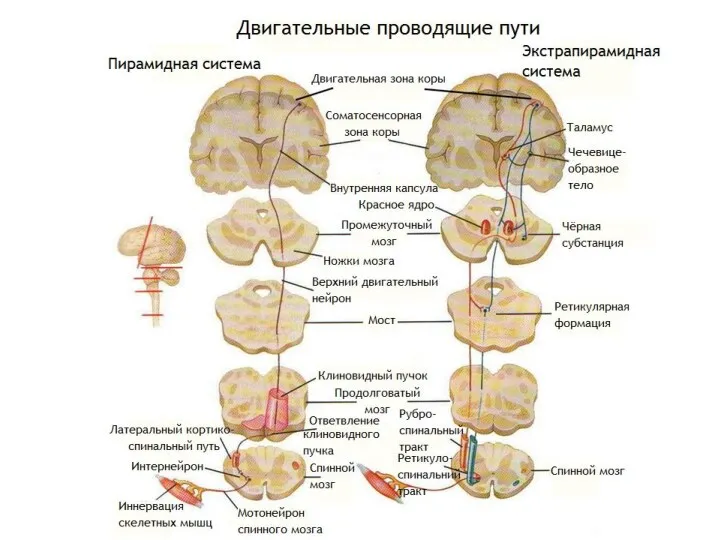

Пирамидная система, пирамидный путь, система нервных структур, участвующих в сложной и
Пирамидная система, пирамидный путь, система нервных структур, участвующих в сложной и

Экстрапирамидная система осуществляет непроизвольную регуляции и координацию движений, регуляцию мышечного тонуса,
Экстрапирамидная система осуществляет непроизвольную регуляции и координацию движений, регуляцию мышечного тонуса,



РФ включает:
Латеральные ретикулярные ядра;
Ретикулярные вентральные ядра;
Ядра покрышки (Бехтерева);
Парамедиальные ретикулярные ядра;
РФ включает:
Латеральные ретикулярные ядра;
Ретикулярные вентральные ядра;
Ядра покрышки (Бехтерева);
Парамедиальные ретикулярные ядра;

Ретикулярная формация (РФ)
Функциональные особенности ретикулярных нейронов.
Полисенсорная конвергенция: принимают коллатерали от нескольких
Ретикулярная формация (РФ)
Функциональные особенности ретикулярных нейронов.
Полисенсорная конвергенция: принимают коллатерали от нескольких

Проводниковые функции:
Афф.пути представлены спиноретикулярным, несущим импульсы от температурных, болевых рецепторов; Церебеллоретикулярный
Проводниковые функции:
Афф.пути представлены спиноретикулярным, несущим импульсы от температурных, болевых рецепторов; Церебеллоретикулярный
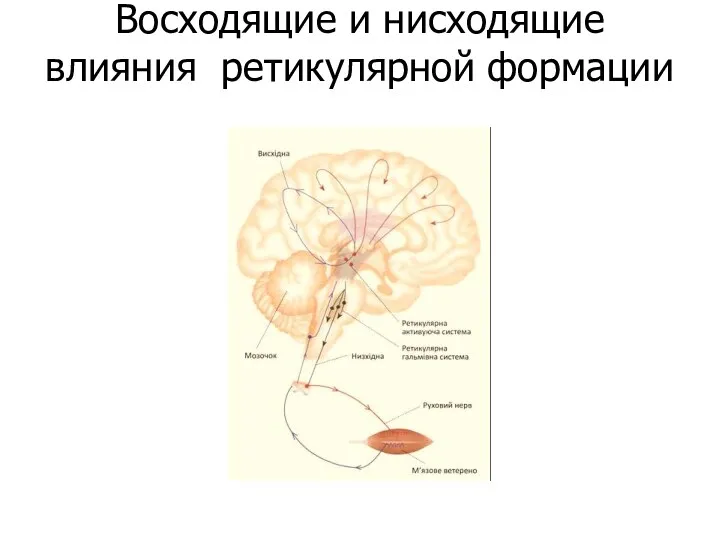
Восходящие и нисходящие влияния ретикулярной формации
Восходящие и нисходящие влияния ретикулярной формации

РФ восходящее влияние
Импульсы ретикулярных нейронов продолговатого мозга (гигантеклеточное, латеральное и вентральное
РФ восходящее влияние
Импульсы ретикулярных нейронов продолговатого мозга (гигантеклеточное, латеральное и вентральное

РФ нисходящее влияние
Структуры р.ф. ствола мозга оказывают облегчающее и тормозное влияние
РФ нисходящее влияние
Структуры р.ф. ствола мозга оказывают облегчающее и тормозное влияние
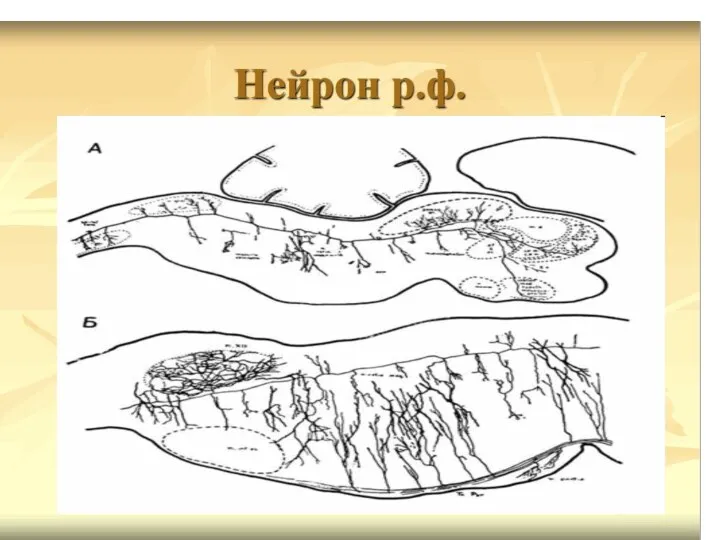
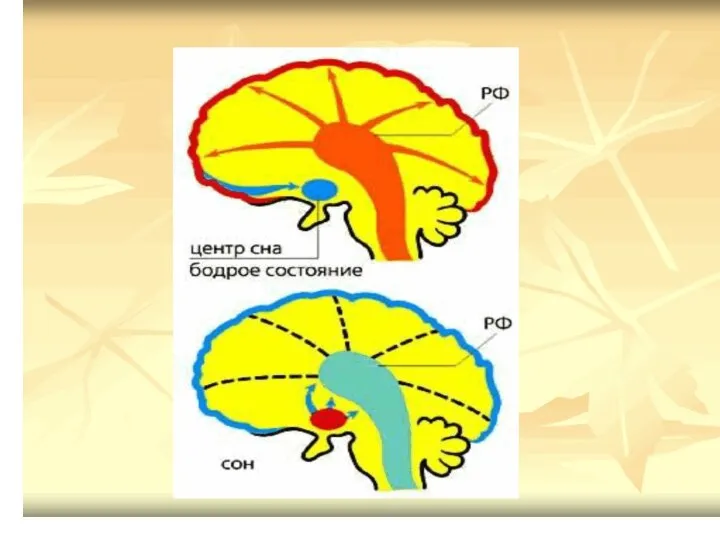


Нисходящие влияние на тонус мышц и формирование позы РФ оказывает через
Нисходящие влияние на тонус мышц и формирование позы РФ оказывает через


Лимбическая система
Лимбическая система – совокупность структур промежуточного, конечного и среднего мозга,
Лимбическая система
Лимбическая система – совокупность структур промежуточного, конечного и среднего мозга,

Функции лимбической системы. Получая инфу о внешней среде и внутренней среде
Функции лимбической системы. Получая инфу о внешней среде и внутренней среде
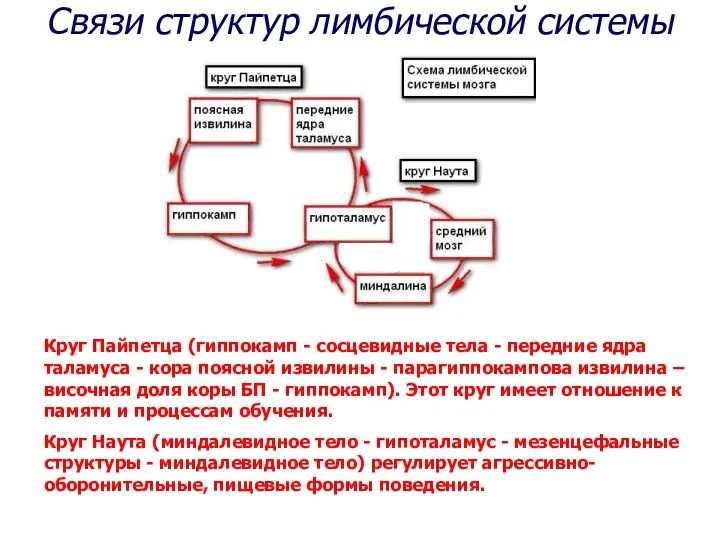
Круг Пайпетца (гиппокамп - сосцевидные тела - передние ядра таламуса -
Круг Пайпетца (гиппокамп - сосцевидные тела - передние ядра таламуса -

Поддерживает гомеостаз и регулирует работу внутренних органов.
При поражении - нарушение
Поддерживает гомеостаз и регулирует работу внутренних органов.
При поражении - нарушение

Функции новой коры
Новая кора – слой серого вещества общей площадью 1100
Функции новой коры
Новая кора – слой серого вещества общей площадью 1100

Слои коры больших полушарий
Полушария головного мозга покрыты серым веществом или корой,
Слои коры больших полушарий
Полушария головного мозга покрыты серым веществом или корой,
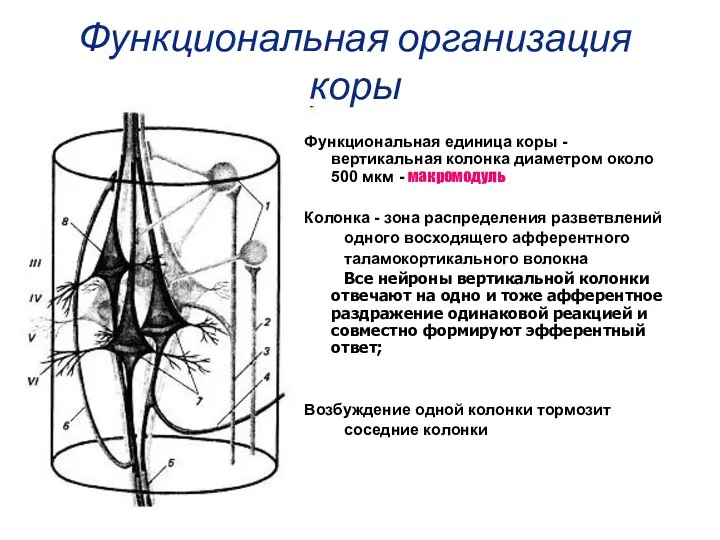
Функциональная организация коры
Функциональная единица коры - вертикальная колонка диаметром около 500
Функциональная организация коры
Функциональная единица коры - вертикальная колонка диаметром около 500

Физиологическое значение коры:
1. Обеспечение высшей нервной деятельности.
2. Обработка сенсорной информации.
3. Формирование
Физиологическое значение коры:
1. Обеспечение высшей нервной деятельности.
2. Обработка сенсорной информации.
3. Формирование

Функции новой коры
Новая кора – слой серого вещества общей площадью 1100
Функции новой коры
Новая кора – слой серого вещества общей площадью 1100

Функции клеток коры больших полушарий
Основная информация в кору поступает по специфическим
Функции клеток коры больших полушарий
Основная информация в кору поступает по специфическим

Кортикальные связи
1 – ассоциативные (связывают зоны коры в пределах одного полушария);
2
Кортикальные связи
1 – ассоциативные (связывают зоны коры в пределах одного полушария);
2

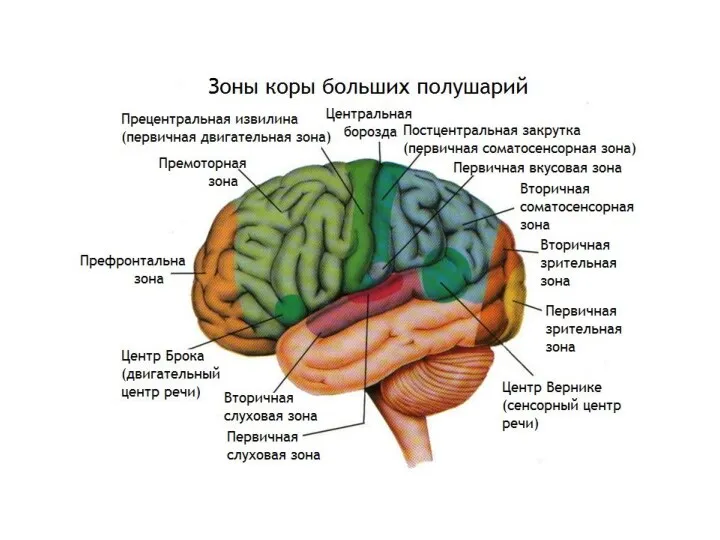
Вторичная соматосенсорная зона расположена в стенке боковой борозды, на границе ее
Вторичная соматосенсорная зона расположена в стенке боковой борозды, на границе ее
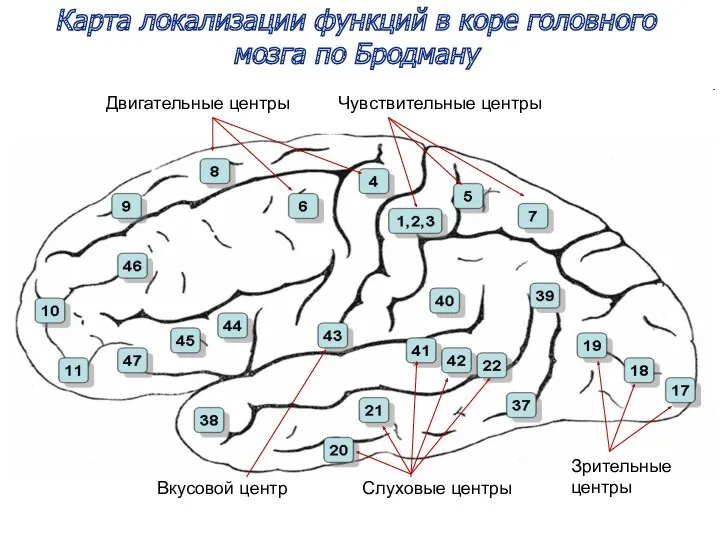
Карта локализации функций в коре головного мозга по Бродману
Вкусовой центр
Чувствительные
Карта локализации функций в коре головного мозга по Бродману
Вкусовой центр
Чувствительные
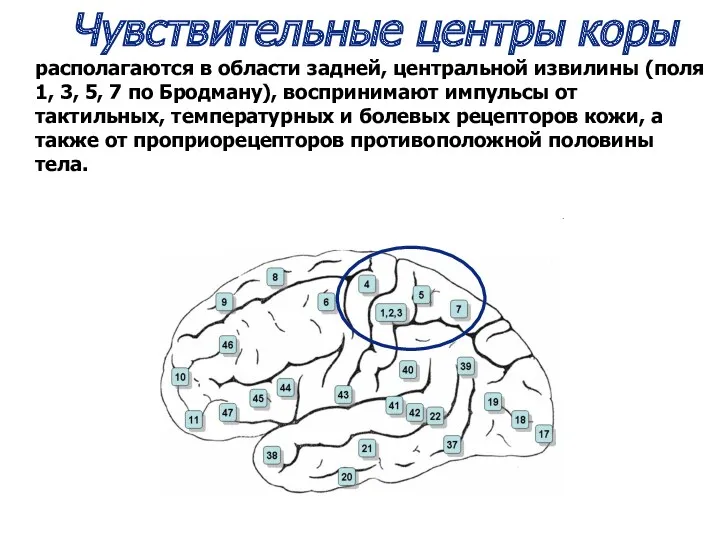
Чувствительные центры коры
располагаются в области задней, центральной извилины (поля 1,
Чувствительные центры коры
располагаются в области задней, центральной извилины (поля 1,
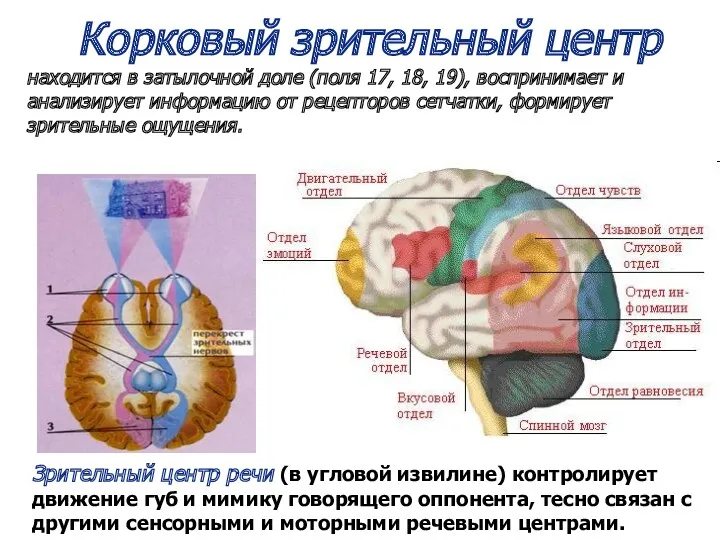
Корковый зрительный центр
находится в затылочной доле (поля 17, 18, 19),
Корковый зрительный центр
находится в затылочной доле (поля 17, 18, 19),
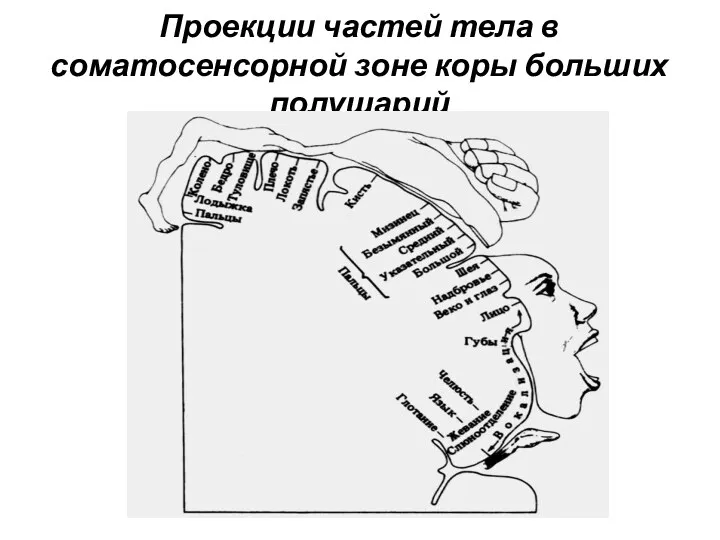
Проекции частей тела в соматосенсорной зоне коры больших полушарий
Проекции частей тела в соматосенсорной зоне коры больших полушарий
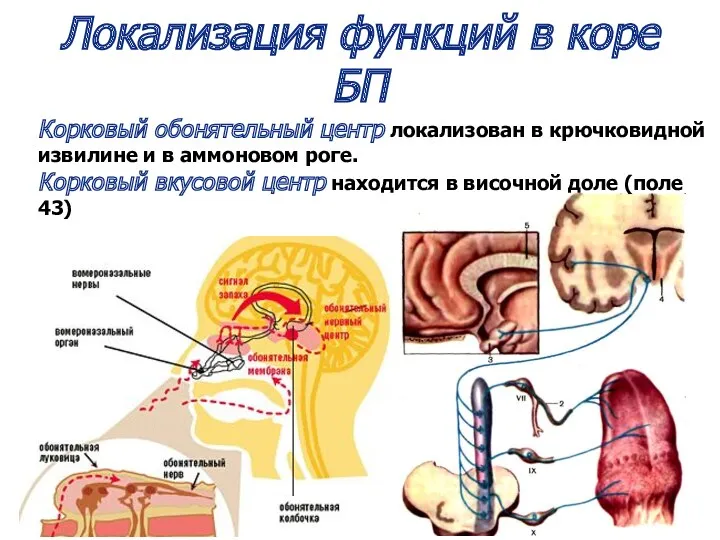
Корковый обонятельный центр локализован в крючковидной извилине и в аммоновом роге.
Корковый
Корковый обонятельный центр локализован в крючковидной извилине и в аммоновом роге.
Корковый
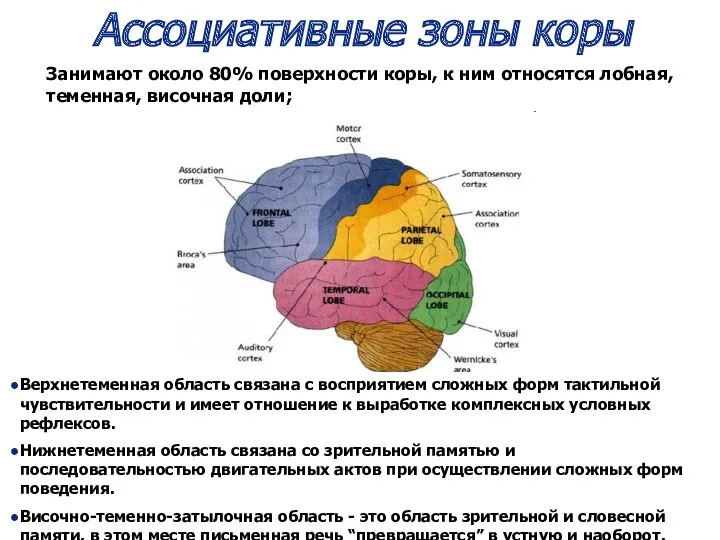
Ассоциативные зоны коры
Верхнетеменная область связана с восприятием сложных форм тактильной
Ассоциативные зоны коры
Верхнетеменная область связана с восприятием сложных форм тактильной
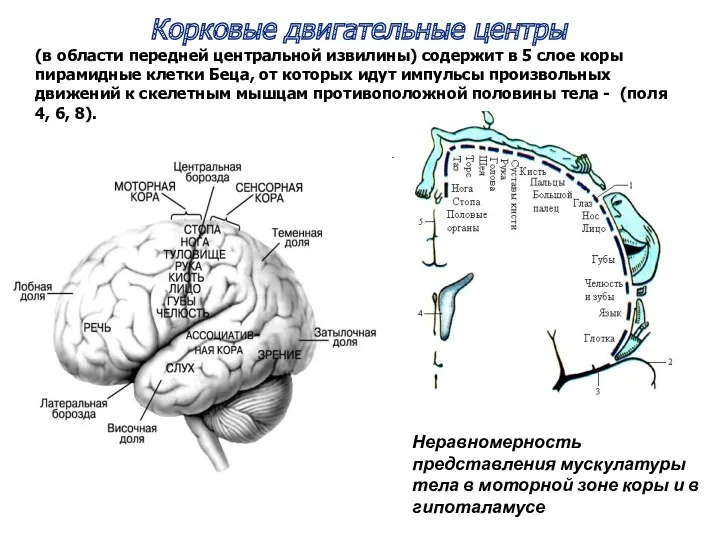
Корковые двигательные центры
(в области передней центральной извилины) содержит в 5
Корковые двигательные центры
(в области передней центральной извилины) содержит в 5
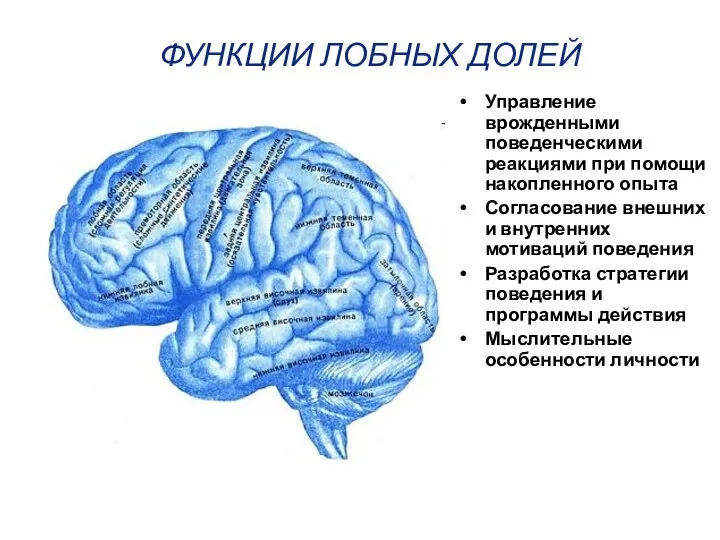
ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
Управление врожденными поведенческими реакциями при помощи накопленного опыта
Согласование внешних
ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
Управление врожденными поведенческими реакциями при помощи накопленного опыта
Согласование внешних

Центр проприоцептивного гнозиса (в верхней теменной дольке) обеспечивает восприятие
Локализация функций в
Центр проприоцептивного гнозиса (в верхней теменной дольке) обеспечивает восприятие
Локализация функций в
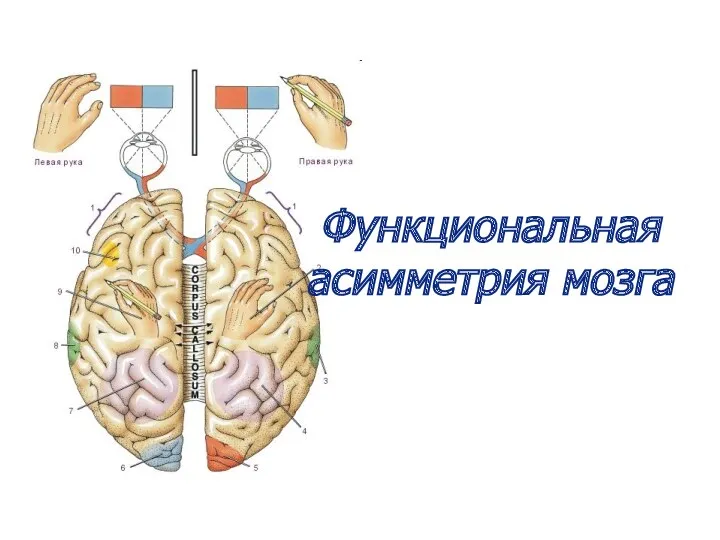
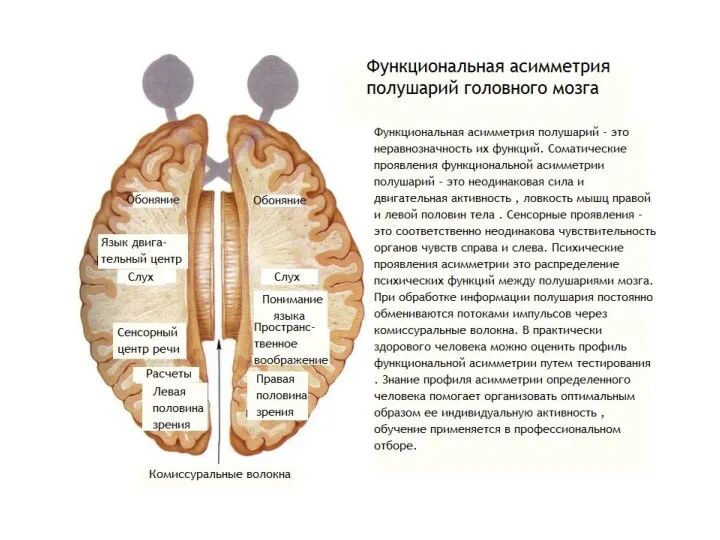
Функциональная асимметрия мозга
Функциональная асимметрия мозга

Принципы межполушарных взаимодействий
Параллельная деятельность – информация параллельно обрабатывается в обоих полушариях
Принципы межполушарных взаимодействий
Параллельная деятельность – информация параллельно обрабатывается в обоих полушариях

Функции полушарий мозга
Левое полушарие
Специализируется на выполнении вербальных символических функций, формировании абстрактного
Функции полушарий мозга
Левое полушарие
Специализируется на выполнении вербальных символических функций, формировании абстрактного

Функции полушарий мозга
Правое полушарие
Быстрее обрабатывает информацию, участвует в формировании конкретного мышления,
Функции полушарий мозга
Правое полушарие
Быстрее обрабатывает информацию, участвует в формировании конкретного мышления,
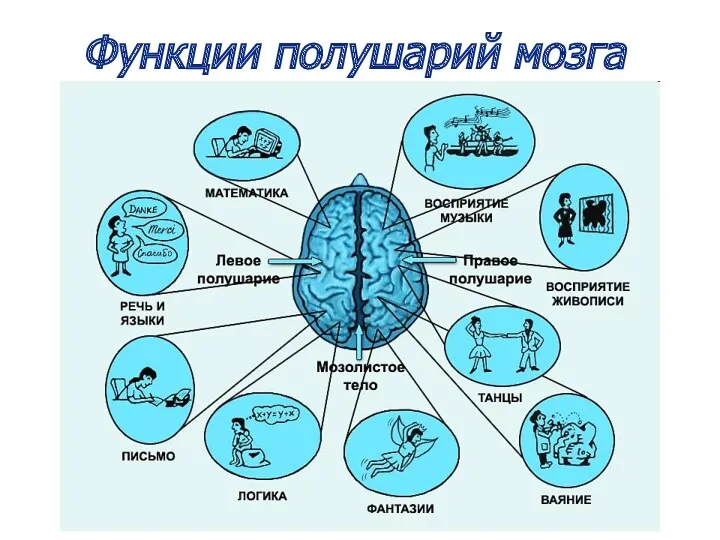
Функции полушарий мозга
Функции полушарий мозга

Межполушарные различия
ЛЕВОЕ ПОЛУШАРИЕ ПРАВОЕ ПОЛУШАРИЕ
Лучше узнаются стимулы
Словесные Несловесные
Легко
Межполушарные различия
ЛЕВОЕ ПОЛУШАРИЕ ПРАВОЕ ПОЛУШАРИЕ
Лучше узнаются стимулы
Словесные Несловесные
Легко

Понятия «леворукость» и «левшество»
синонимами не являются.
Леворукость – это термин, отражающий
Понятия «леворукость» и «левшество»
синонимами не являются.
Леворукость – это термин, отражающий

Генетическое левшество.
Ученые Оксфордского университета обнаружили ген ЛРРТМ1, устанавливающий «правила» работы
Генетическое левшество.
Ученые Оксфордского университета обнаружили ген ЛРРТМ1, устанавливающий «правила» работы

«Компенсаторное" левшество
связанно с каким-либо поражением мозга, чаще - его левого полушария.
«Компенсаторное" левшество
связанно с каким-либо поражением мозга, чаще - его левого полушария.

Псевдолеворукость
К определенному возрасту (окончательно примерно к 5 годам) у ребенка какое-то
Псевдолеворукость
К определенному возрасту (окончательно примерно к 5 годам) у ребенка какое-то

Скрытое левшество
Наблюдается при смене доминирующего полушария. Момент смены является тем критическим
Скрытое левшество
Наблюдается при смене доминирующего полушария. Момент смены является тем критическим

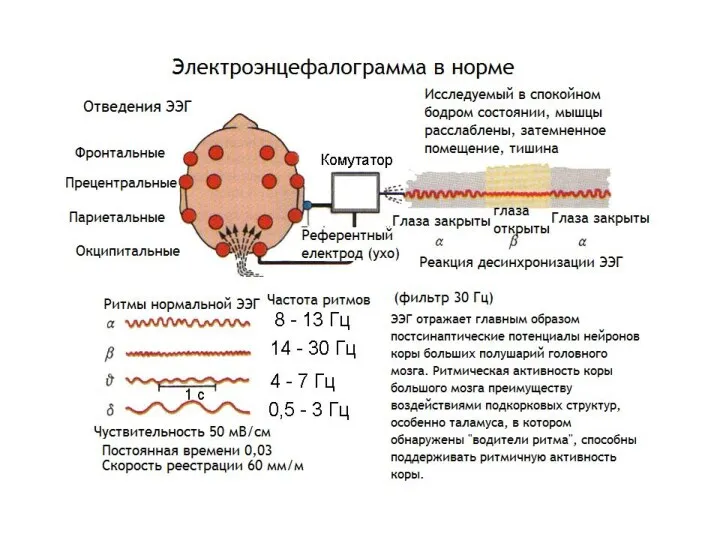
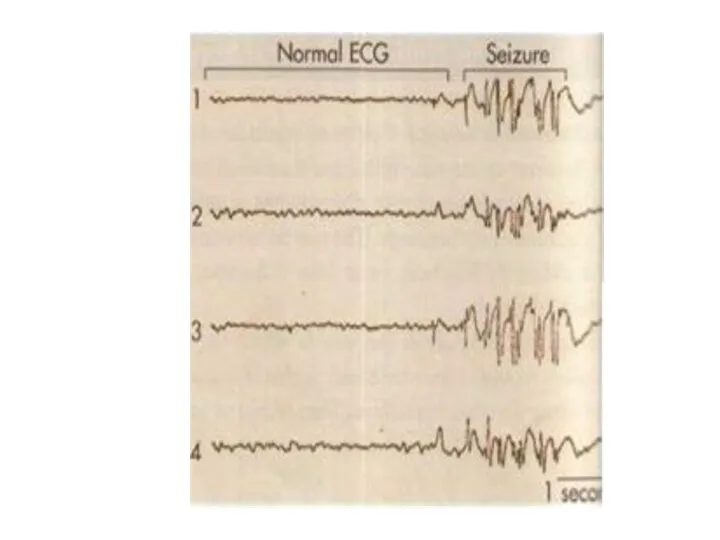
Электроэнцефалография.
Электроэнцефалография - это метод регистрации суммарной биоэлектрической активности мозга с поверхности
Электроэнцефалография.
Электроэнцефалография - это метод регистрации суммарной биоэлектрической активности мозга с поверхности

Роль спинного мозга в регуляции функций организма: вегетативные и соматические центры
Роль спинного мозга в регуляции функций организма: вегетативные и соматические центры
 Лейкокория у детей
Лейкокория у детей Методика исследования периферических и внутригрудных лимфатических узлов
Методика исследования периферических и внутригрудных лимфатических узлов Инфекционные болезни - чума
Инфекционные болезни - чума Наложение шин на руку
Наложение шин на руку Сердечно-легочная реанимация
Сердечно-легочная реанимация Жүктілікпен байланысты гипертензиясы Ерте токсикоздар
Жүктілікпен байланысты гипертензиясы Ерте токсикоздар Воздействие на поведение водителя психотропных, наркотических веществ, алкоголя и медицинских препаратов
Воздействие на поведение водителя психотропных, наркотических веществ, алкоголя и медицинских препаратов Косметические средства ухода за кожей детей при аномалиях конституции
Косметические средства ухода за кожей детей при аномалиях конституции Diet and teen health
Diet and teen health Зрительный нерв. Заболевания зрительного нерва
Зрительный нерв. Заболевания зрительного нерва Реанимационная помощь
Реанимационная помощь Дәрілік заттардың әсерінің ағза қасиеттеріне тәуелділігі. Фармакологиялық әсердің дәрілік заттардың қасиеттері мен қолдану
Дәрілік заттардың әсерінің ағза қасиеттеріне тәуелділігі. Фармакологиялық әсердің дәрілік заттардың қасиеттері мен қолдану Antimicrobial drugs
Antimicrobial drugs Снотворные и противосудорожные средства. Противопаркинсонические средства
Снотворные и противосудорожные средства. Противопаркинсонические средства Предмет и задачи медицинской микробиологии. История микробиологии. Медицинская микробиология XXI века
Предмет и задачи медицинской микробиологии. История микробиологии. Медицинская микробиология XXI века Ошибки диагностики и лечения при сочетанной травме
Ошибки диагностики и лечения при сочетанной травме Профилактика стоматологических заболеваний в раннем детском возрасте
Профилактика стоматологических заболеваний в раннем детском возрасте Вскармливание детей грудного возраста. Раздел 1.Лекция 6
Вскармливание детей грудного возраста. Раздел 1.Лекция 6 Современные направления пластической хирургии в акушерстве и гинекологии
Современные направления пластической хирургии в акушерстве и гинекологии Местные анестетики
Местные анестетики Всероссийский день трезвости
Всероссийский день трезвости ВКР: Анализ работы медсестры процедурного кабинета хирургического отделения стационара
ВКР: Анализ работы медсестры процедурного кабинета хирургического отделения стационара Врожденные аномалии грудной клетки
Врожденные аномалии грудной клетки Вреден ли фаст-фуд для нашего здоровья
Вреден ли фаст-фуд для нашего здоровья Здоровый образ жизни и профилактика основных неинфекционных заболеваний
Здоровый образ жизни и профилактика основных неинфекционных заболеваний Қазіргі заманғы денсаулық сақтау, әлемдік тенденция, дамыған елдер
Қазіргі заманғы денсаулық сақтау, әлемдік тенденция, дамыған елдер Иммуногенез бұзылуының морфологиясы. Тимустың, шеткері лимфалық тіндердің иммунгенез бұзылғандағы өзгерістері
Иммуногенез бұзылуының морфологиясы. Тимустың, шеткері лимфалық тіндердің иммунгенез бұзылғандағы өзгерістері Жүкті студенттердің медико-әлеуметтік аспектілері
Жүкті студенттердің медико-әлеуметтік аспектілері