- Онкогенні папіломавіруси

Содержание
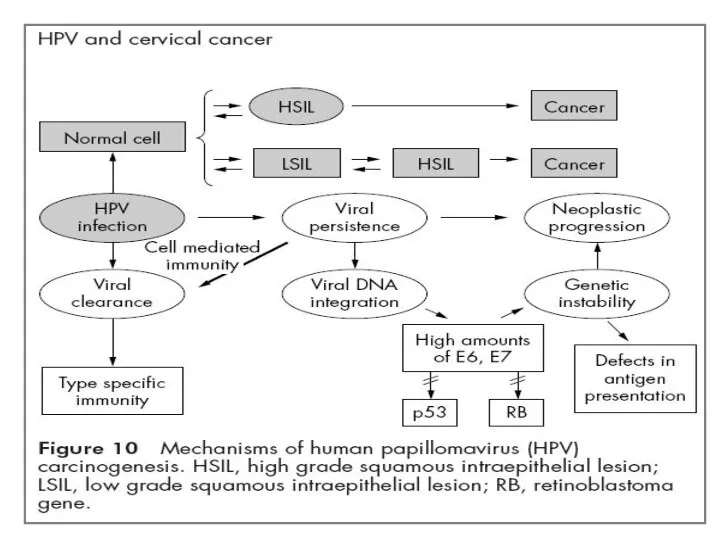
- 2. . Контроль клеточного цикла и дифференцировка клеток осуществляется белками Е6 и Е7 посредством их взаимодействия и
- 3. Роль гормональних факторів у розвитку раку шийки матки Відома роль естрогенів в розвитку неопластичних процесів в
- 4. р53 Установлено, что онкобелок Е6, кодируемый вирусами ВПЧ 18 и ВПЧ 16, может взаимодействовать с белком
- 5. HARALD ZUR HAUSEN У 1974–1976 рр. вперше висунув припущення про можливу участь Human Papilloma Virus, HPV
- 6. Папілома- віруси Руки, ноги, гортань або статевий тракт Зараження епітелію Локальне розмноження бородавка Відшарування (латенція) Клітинна
- 7. Папіломи — посилене розмноження клітин базального шару (бородавки або папіломи на шкірі обличчя, кінцівках, статевих органів)
- 8. ВПЧ инфицирует эпителий и реплицируется в нем, что подтвержда"ется обнаружением эписомального вирусного генома в клетках базального
- 9. HPV класифікація: онкогенний ризик Онкогенний ризик базується на потенціалі Е білків різних штамів Низький ? знаходять
- 10. Common warts with thrombosed vessels (black dots). (From Medical Microbiology, 4th ed., Murray, Rosenthal, Kobayashi &
- 12. Рак шкіри Epidermodysplasia verruciformis Рідке генетичне захворювання ДНК репарація та імунна відповідь Може бути уражено все
- 13. Генітиальний тракт та “слизові” HPV (From Fields Virology, 4th ed, Knipe & Howley, eds, Lippincott Williams
- 14. Respiratory Papillomas HPv - типи 6 тa 11 3-7/100,000 Хірургічне втручання Малігнізація 1- 3% хворих Відомий
- 15. Генітальна інфекція HPV переважно: 30% жінок 15-40 років Малігнізація - Можливі ко-фактори: куріння,екстроген, HLA тип типи
- 16. Поширення ВПЛ Пік ВПІ - 18–25 років та знижується після 30 р., суттєво зростає частота дисплазій
- 17. HPV високого ризику - фактор виникнення РШМ Присутність всіх типів HPV показано ПЦР у 95% РШМ:
- 18. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings. На основі
- 19. HPV16 genome Genomic map of HPV-16. The genome is a double-stranded circular DNA molecule of 7904
- 20. Функції генів папіломавірусів (From Fields Virology, 4th ed, Knipe & Howley, eds, Lippincott Williams & Wilkins,
- 21. ДНК HPV можуть виявляти на відстані до 1 см від «кордонів» пухлини
- 22. Експресія HPV mРНК Базальна мембрана Surface
- 23. Схема реплікації HPV Продуктивна інфекція залежить від процесу диференціації, а не від ступеня зрілості клітин Абортивна
- 24. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings.
- 25. Інтеграція неспецифічна, в різні ділянки геному, часто в область підвищеної “хрупкості” хромосом. Інтеграція в-ДНК індукує декілька
- 26. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings.
- 27. Трансформуючі властивості папіломавірусів забезпечується функціонуванням генів Е5, Е6 та Е7 Продукт гену Е5 важливий на ранніх
- 28. Клітинна сигналізація
- 29. Papilloma E5 гомолог EGFR ліганду Ліганд-зв”я-зуючий домен Кіназний домен Опосередкована EGFR димерізація BPV E5 -ліганд -незалежна
- 30. EGFR та сигнальні шляхи ras raf MEK 1,2 MAPK-P PI-3-K PI-3,4,5-P AKT -P PIP2 PTEN PKC
- 31. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings.
- 32. HPV високого ризику онкобілки: Hausen, 2000.
- 33. Ранні гени та точки рестрикції клітинного циклу E6 та E7 взаємодіють з ключовимим білками проходження клітинного
- 34. Білки-мішені для Е7 АР1 (члени родини) - інгібування ІРФ-1 транскрипційної активності С-mус - посилення с-mус індукованої
- 35. E7 плеотропний білок Інактивація p21CIP-1 таp27KIP-1 (cdk -інгібітор) призводить до посилення росту інфікованих клітин Е7-залежна регуляція
- 36. E7 дезактивує IRF-1 Можливість пояснення механізми імуно-резистентності HPV-інфікованих цервікальних клітин IRF-1 aктивується впродовж встановлення вірусної інфекції,
- 37. E6 плеотропний білок Bak, Bax Анти-апоптичний ефект СВР/р300 Дерегуляція р53 залежну транскрипції С-mус Запобігання апоптозу, підвищення
- 38. Стимуляція експресії транскрипційного факторуr HIF-1α Прогностичний маркер: високий рівень експресії HIF-1α на ранніх етапах інвазивного раку
- 40. Е6 Онкобелок Е6 онкогенных типов ВПЧ (типов 16 и 18) способен вызвать и ммортализацию эпителиальных клеток
- 41. Процесинг антигену TAP-1 та протеосоми критична ланка для MHC I презентації E7 звязує обох,інактивуючи ATP’азну активність
- 42. E6 & E7 in Cervical Cancer Progression Furumoto et al., 2002.
- 43. From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006 Pearson Benjamin Cummings.
- 44. ІМУНОПАТОГЕНЕЗ HPV І Вірус не уражує антиген-презентуючі клітини (АПК) – уникнення прямого шляху активації імунітету Ранні
- 45. Імунологічна чутливість до інфекції HPV генетично детермінована та важлива для прогресії CIN та РШМ. Жінки з
- 46. Фактори ризику розвитку цервікального раку Фактори ризику персистенції інфекції HPV Кількість сексуальних партнерів впродовж життя Вік
- 47. Багатостадійність РШМ
- 48. Карцинома — вірус інтегрований в геном клітини –поява змінених «атипових» клітин, що свідчить про злоякісність процесу
- 49. Дисплазія (неоплазія) — вірус інтегрований в геном клітини — відбуваються зміни в структурі клітин, койлоцитоз —
- 50. Високий ризик HPV -16 та 18, зустрічається 70 % випадків.
- 51. Cervical squamous carcinoma precursors. Schematic representation of cervical cancer precursors and the different terminologies that have
- 52. Стадії CIN Cervical intraepithelial neoplasia (CIN) 1. CIN I Кількість та глибина аномальних клітин низька 2.
- 53. Кореляція між типами HPV, інтеграцією та гістологією (From Cullen, A.P., Reid, R., Campion, M., Lorincz, A.T
- 54. Вимоги для трансформації Персистуюча інфекція – реплікація вірусу Експресія вірусних білків Мутації в клітині Пeрсистуюча інфекція
- 55. Клітинні фактори та злоякісність HPV Невеликий % розвитку раку Тривалий латентний період Стадії прогресії захворювання Моноклональність
- 57. Головна відмінність HPV від інших вірусів –можливість організму позбавитись від нього самостійно. УСПІХ звільнення від папіломи
- 58. Шляхи встановлення та прогресії малігнізації інфекція Зміни в клітинній регуляції Зміни в кл.генах ↑ HPV експресія
- 59. Механізм інактивації Rb E2F Rb E1A T ag E7 E2F Rb E1A Транскрипція E2F відповідних генів
- 60. Механізми інактивації p53 p53 T ag p53 Tag Stabilizes p53 in an inactive state p53 E6
- 61. ПІДСУМОК: HPV Структура Невеликий (8 kb) циркулярний длДНК геном, голий капсид Хвороби Бородавки шкіри, генітальний тракт
- 62. Structure of the single copy of HPV-16 DNA integrated into the SiHa cell line derived from
- 64. Скачать презентацию
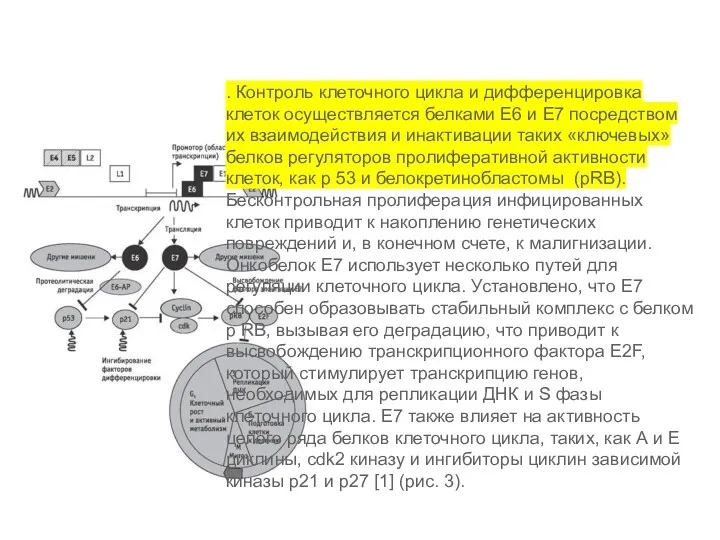
. Контроль клеточного цикла и дифференцировка клеток осуществляется белками Е6 и
. Контроль клеточного цикла и дифференцировка клеток осуществляется белками Е6 и

Роль гормональних факторів у розвитку раку шийки матки
Відома роль естрогенів в
Роль гормональних факторів у розвитку раку шийки матки
Відома роль естрогенів в

р53
Установлено, что онкобелок Е6, кодируемый вирусами ВПЧ 18 и ВПЧ
р53
Установлено, что онкобелок Е6, кодируемый вирусами ВПЧ 18 и ВПЧ

HARALD ZUR HAUSEN
У 1974–1976 рр. вперше висунув припущення про можливу
HARALD ZUR HAUSEN
У 1974–1976 рр. вперше висунув припущення про можливу
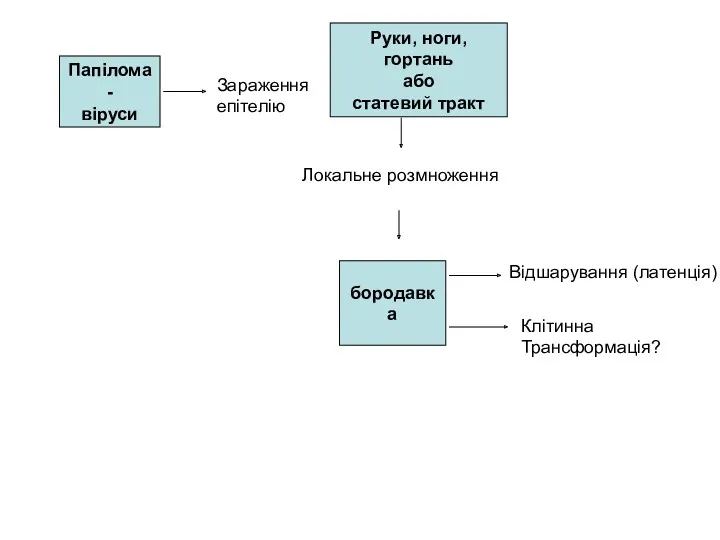
Папілома-
віруси
Руки, ноги,
гортань
або
статевий тракт
Зараження
епітелію
Локальне розмноження
бородавка
Відшарування (латенція)
Клітинна
Трансформація?
Папілома-
віруси
Руки, ноги,
гортань
або
статевий тракт
Зараження
епітелію
Локальне розмноження
бородавка
Відшарування (латенція)
Клітинна
Трансформація?

Папіломи — посилене розмноження клітин базального шару (бородавки або папіломи на
Папіломи — посилене розмноження клітин базального шару (бородавки або папіломи на

ВПЧ инфицирует эпителий и реплицируется в нем, что подтвержда"ется обнаружением эписомального
ВПЧ инфицирует эпителий и реплицируется в нем, что подтвержда"ется обнаружением эписомального

HPV класифікація:
онкогенний ризик
Онкогенний ризик базується на потенціалі Е білків різних
HPV класифікація:
онкогенний ризик
Онкогенний ризик базується на потенціалі Е білків різних
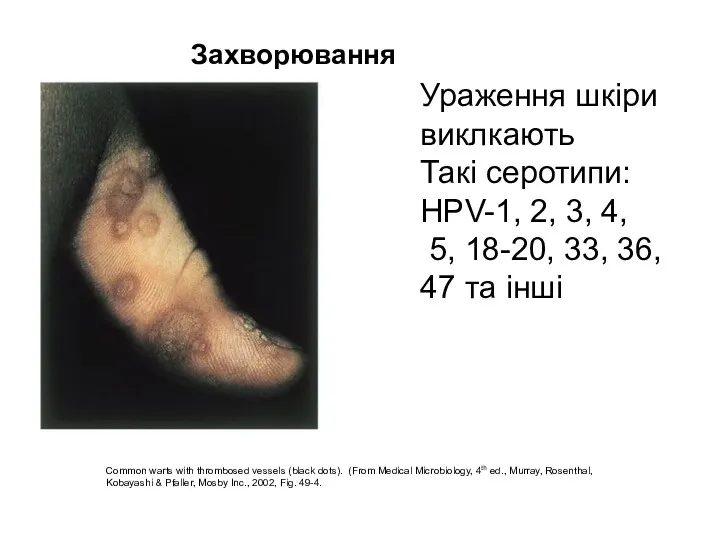
Common warts with thrombosed vessels (black dots). (From Medical Microbiology, 4th
Common warts with thrombosed vessels (black dots). (From Medical Microbiology, 4th
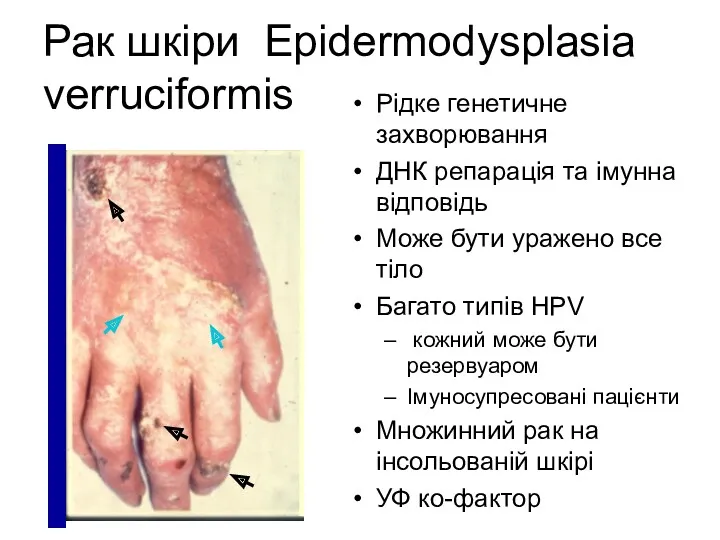
Рак шкіри Epidermodysplasia verruciformis
Рідке генетичне захворювання
ДНК репарація та імунна відповідь
Може бути
Рак шкіри Epidermodysplasia verruciformis
Рідке генетичне захворювання
ДНК репарація та імунна відповідь
Може бути

Генітиальний тракт та “слизові” HPV
(From Fields Virology, 4th ed, Knipe &
Генітиальний тракт та “слизові” HPV
(From Fields Virology, 4th ed, Knipe &
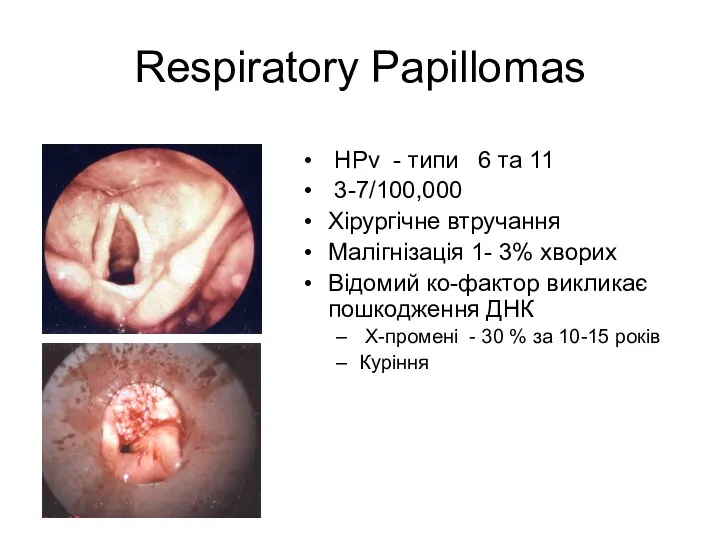
Respiratory Papillomas
HPv - типи 6 тa 11
3-7/100,000
Хірургічне втручання
Малігнізація 1-
Respiratory Papillomas
HPv - типи 6 тa 11
3-7/100,000
Хірургічне втручання
Малігнізація 1-
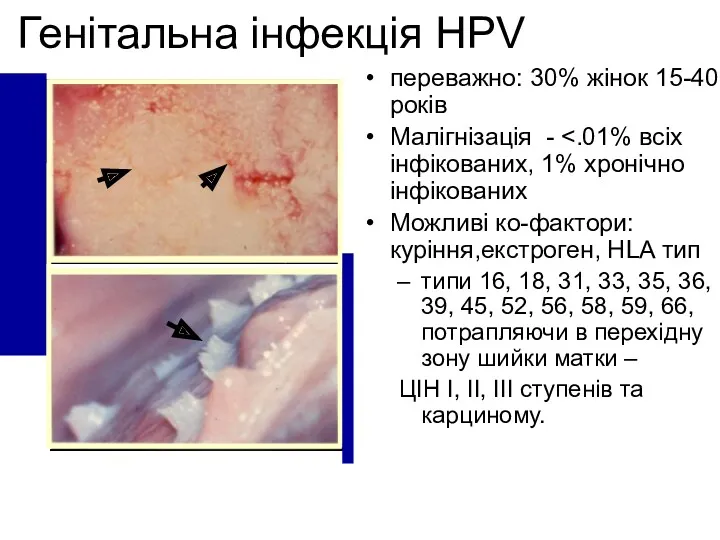
Генітальна інфекція HPV
переважно: 30% жінок 15-40 років
Малігнізація - <.01% всіх інфікованих,
Генітальна інфекція HPV
переважно: 30% жінок 15-40 років
Малігнізація - <.01% всіх інфікованих,

Поширення ВПЛ
Пік ВПІ - 18–25 років та знижується після 30 р.,
Поширення ВПЛ
Пік ВПІ - 18–25 років та знижується після 30 р.,

HPV високого ризику - фактор виникнення РШМ
Присутність всіх типів HPV показано
HPV високого ризику - фактор виникнення РШМ
Присутність всіх типів HPV показано
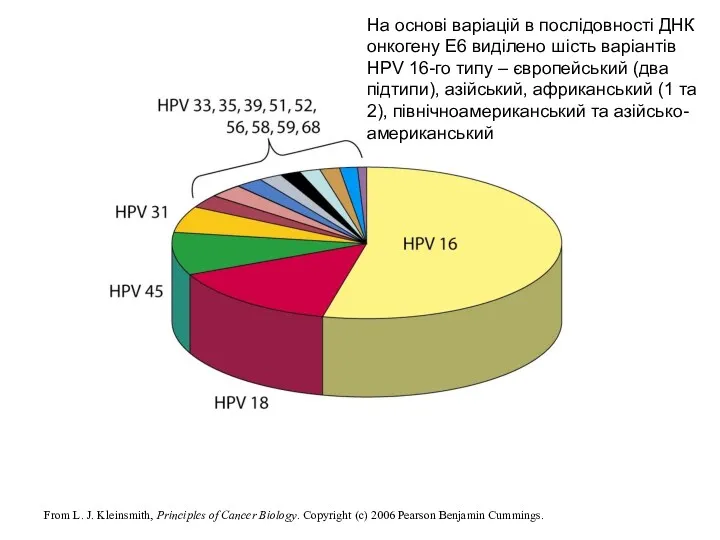
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
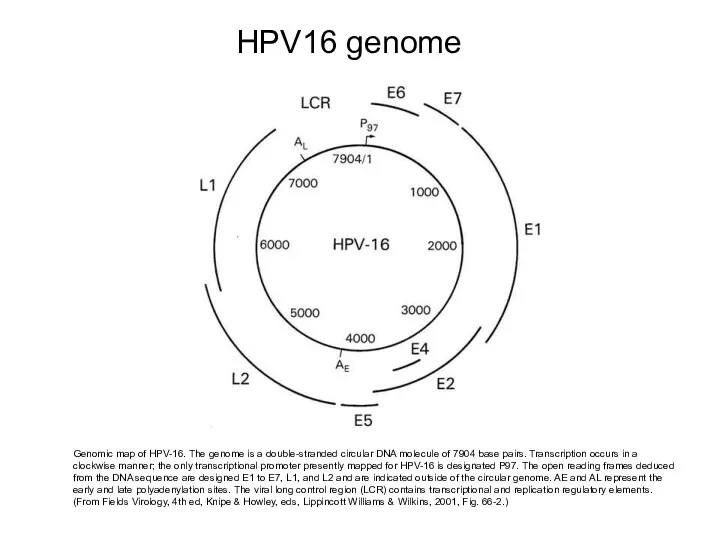
HPV16 genome
Genomic map of HPV-16. The genome is a double-stranded circular
HPV16 genome
Genomic map of HPV-16. The genome is a double-stranded circular

Функції генів папіломавірусів
(From Fields Virology, 4th ed, Knipe & Howley, eds,
Функції генів папіломавірусів
(From Fields Virology, 4th ed, Knipe & Howley, eds,
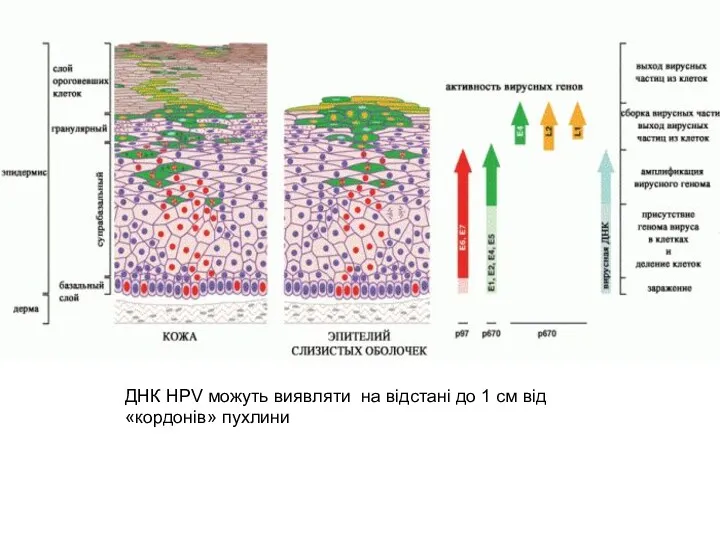
ДНК HPV можуть виявляти на відстані до 1 см від
«кордонів»
ДНК HPV можуть виявляти на відстані до 1 см від
«кордонів»
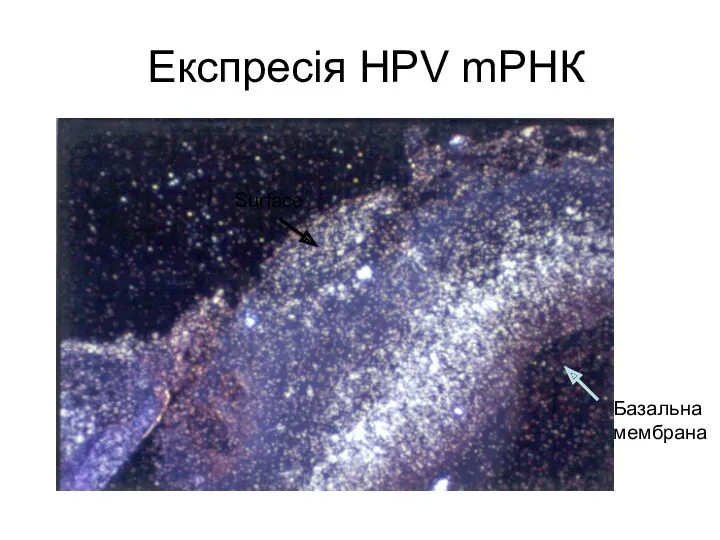
Експресія HPV mРНК
Базальна
мембрана
Surface
Експресія HPV mРНК
Базальна
мембрана
Surface
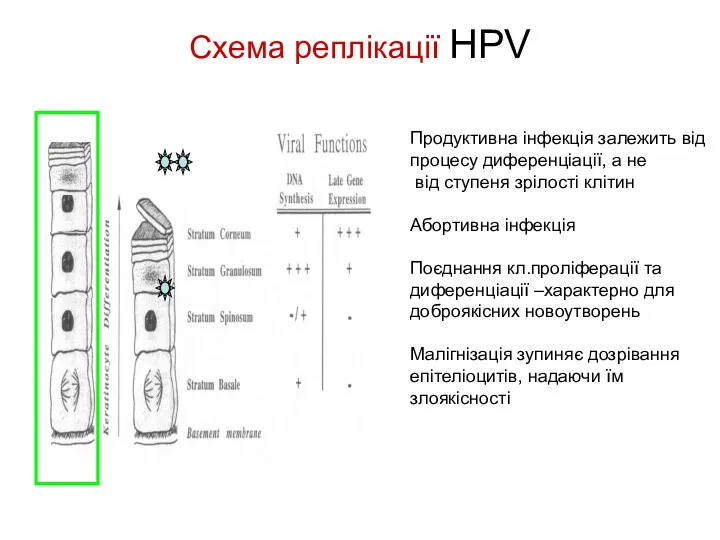
Схема реплікації HPV
Продуктивна інфекція залежить від процесу диференціації, а не
Схема реплікації HPV
Продуктивна інфекція залежить від процесу диференціації, а не
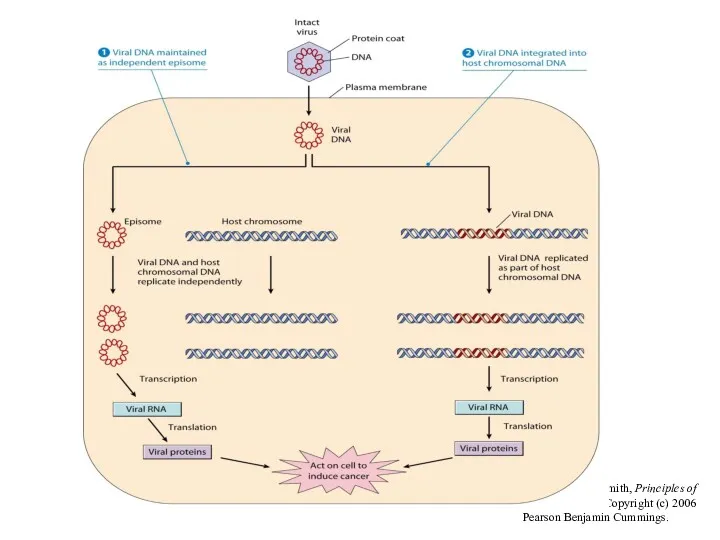
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
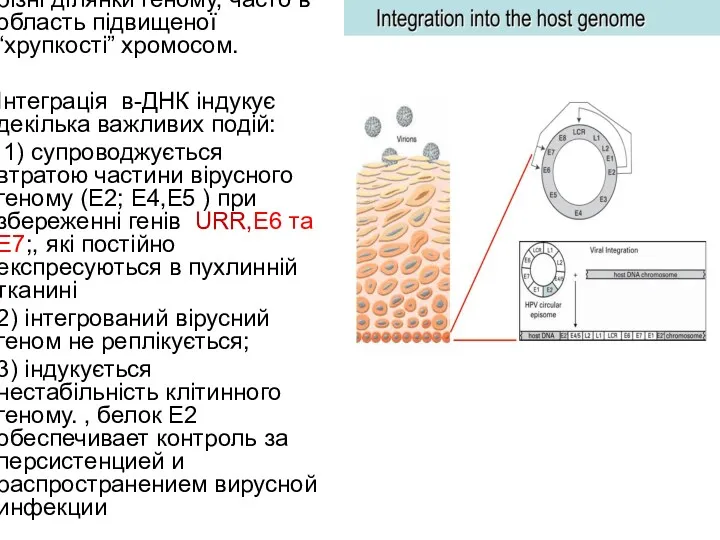
Інтеграція неспецифічна, в різні ділянки геному, часто в область підвищеної “хрупкості”
Інтеграція неспецифічна, в різні ділянки геному, часто в область підвищеної “хрупкості”
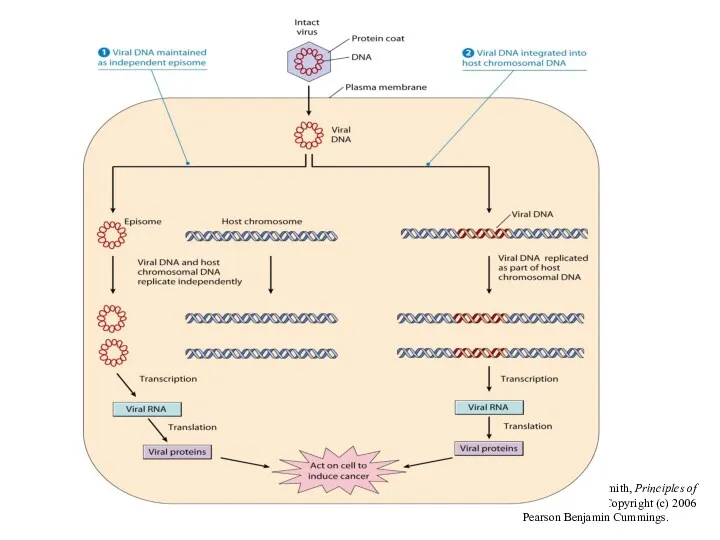
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006

Трансформуючі властивості папіломавірусів забезпечується функціонуванням генів Е5, Е6 та Е7
Продукт
Трансформуючі властивості папіломавірусів забезпечується функціонуванням генів Е5, Е6 та Е7
Продукт
Клітинна сигналізація
Клітинна сигналізація
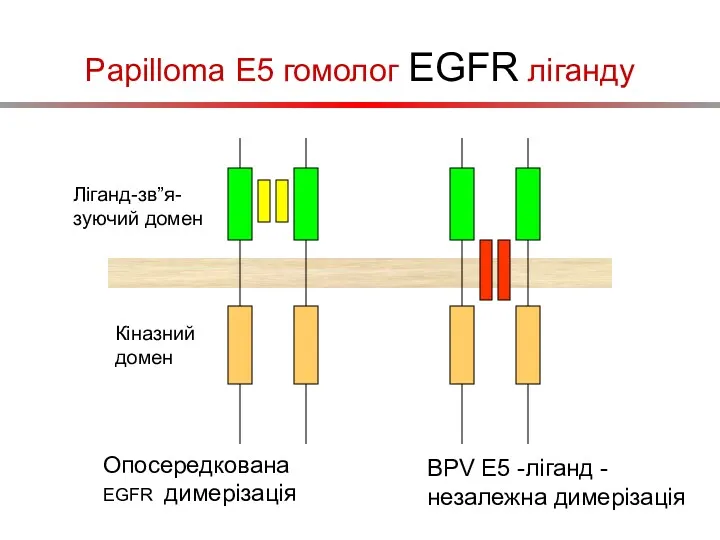
Papilloma E5 гомолог EGFR ліганду
Ліганд-зв”я-зуючий домен
Кіназний домен
Опосередкована EGFR димерізація
BPV E5
Papilloma E5 гомолог EGFR ліганду
Ліганд-зв”я-зуючий домен
Кіназний домен
Опосередкована EGFR димерізація
BPV E5
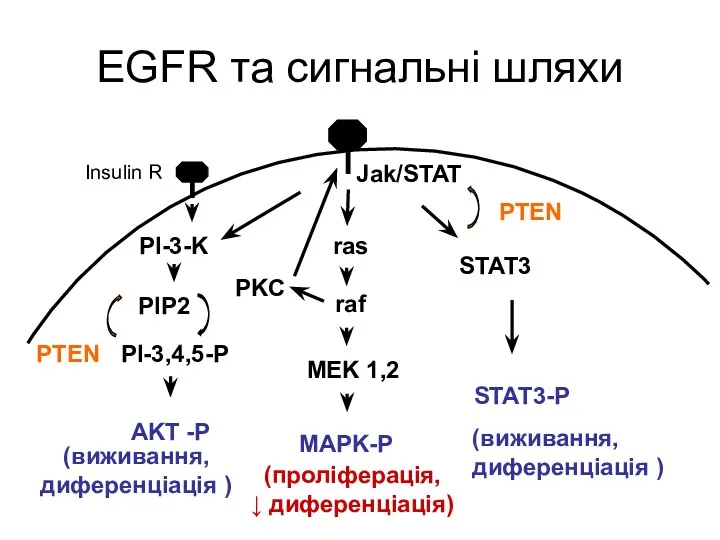
EGFR та сигнальні шляхи
ras
raf
MEK 1,2
MAPK-P
PI-3-K
PI-3,4,5-P
AKT -P
PIP2
PTEN
PKC
Jak/STAT
STAT3-P
STAT3
(виживання,
диференціація )
(виживання,
диференціація )
PTEN
Insulin R
(проліферація,
↓ диференціація)
EGFR та сигнальні шляхи
ras
raf
MEK 1,2
MAPK-P
PI-3-K
PI-3,4,5-P
AKT -P
PIP2
PTEN
PKC
Jak/STAT
STAT3-P
STAT3
(виживання,
диференціація )
(виживання,
диференціація )
PTEN
Insulin R
(проліферація,
↓ диференціація)
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006

HPV високого ризику онкобілки:
Hausen, 2000.
HPV високого ризику онкобілки:
Hausen, 2000.

Ранні гени та точки рестрикції клітинного циклу
E6 та E7 взаємодіють
Ранні гени та точки рестрикції клітинного циклу
E6 та E7 взаємодіють

Білки-мішені для Е7
АР1 (члени родини) - інгібування ІРФ-1 транскрипційної активності
С-mус -
Білки-мішені для Е7
АР1 (члени родини) - інгібування ІРФ-1 транскрипційної активності
С-mус -

E7 плеотропний білок
Інактивація p21CIP-1 таp27KIP-1 (cdk -інгібітор) призводить до посилення росту
E7 плеотропний білок
Інактивація p21CIP-1 таp27KIP-1 (cdk -інгібітор) призводить до посилення росту

E7 дезактивує IRF-1
Можливість пояснення механізми імуно-резистентності HPV-інфікованих цервікальних клітин
IRF-1 aктивується впродовж
E7 дезактивує IRF-1
Можливість пояснення механізми імуно-резистентності HPV-інфікованих цервікальних клітин
IRF-1 aктивується впродовж

E6 плеотропний білок
Bak, Bax Анти-апоптичний ефект
СВР/р300 Дерегуляція р53 залежну транскрипції
С-mус Запобігання
E6 плеотропний білок
Bak, Bax Анти-апоптичний ефект
СВР/р300 Дерегуляція р53 залежну транскрипції
С-mус Запобігання

Стимуляція експресії транскрипційного факторуr HIF-1α
Прогностичний маркер: високий рівень експресії HIF-1α на
Стимуляція експресії транскрипційного факторуr HIF-1α
Прогностичний маркер: високий рівень експресії HIF-1α на


Е6 Онкобелок Е6 онкогенных типов ВПЧ (типов 16 и 18) способен
Е6 Онкобелок Е6 онкогенных типов ВПЧ (типов 16 и 18) способен
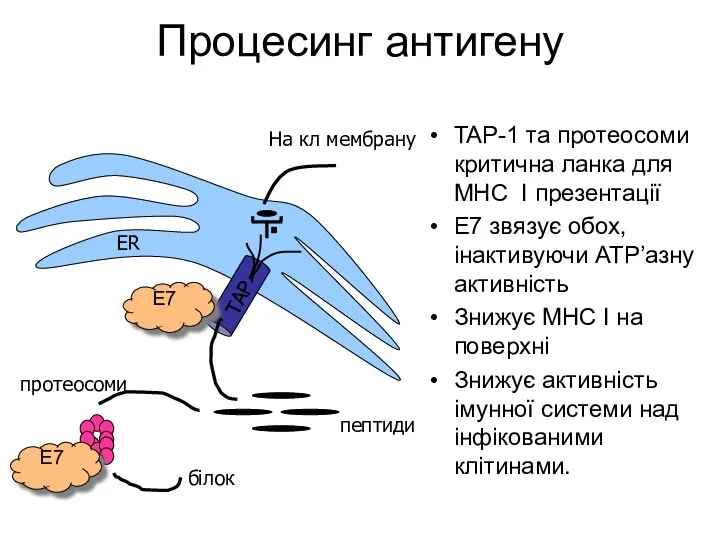
Процесинг антигену
TAP-1 та протеосоми критична ланка для MHC I презентації
E7
Процесинг антигену
TAP-1 та протеосоми критична ланка для MHC I презентації
E7
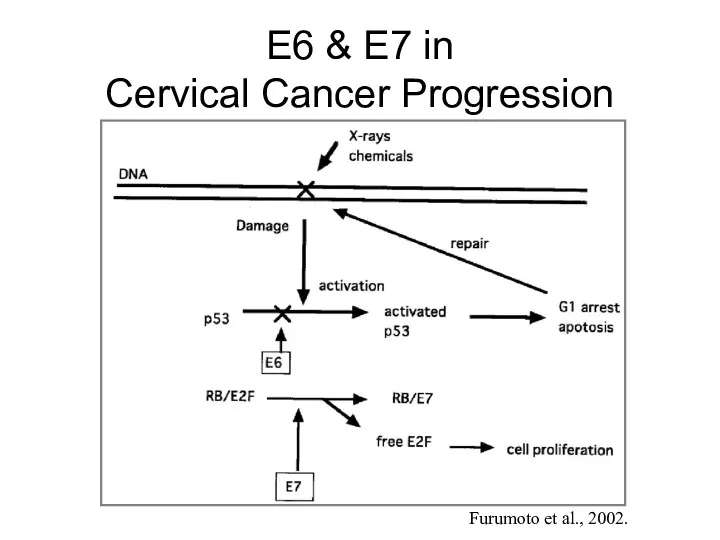
E6 & E7 in
Cervical Cancer Progression
Furumoto et al., 2002.
E6 & E7 in
Cervical Cancer Progression
Furumoto et al., 2002.
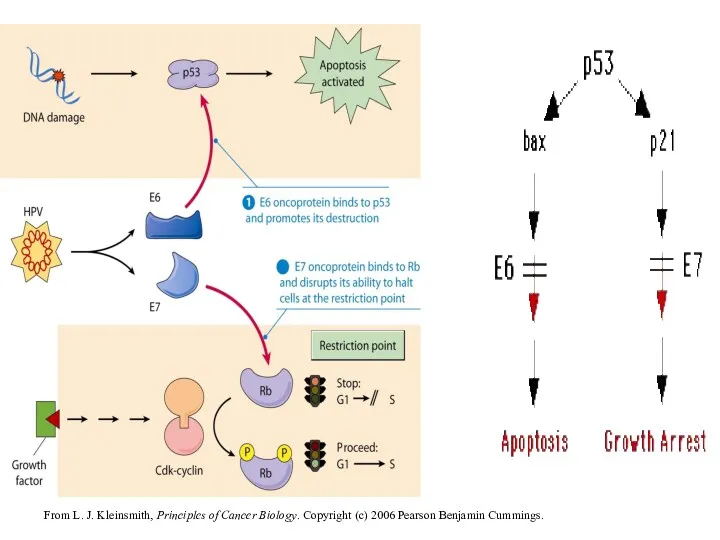
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006
From L. J. Kleinsmith, Principles of Cancer Biology. Copyright (c) 2006

ІМУНОПАТОГЕНЕЗ HPV І
Вірус не уражує антиген-презентуючі клітини (АПК) – уникнення прямого
ІМУНОПАТОГЕНЕЗ HPV І
Вірус не уражує антиген-презентуючі клітини (АПК) – уникнення прямого

Імунологічна чутливість до інфекції HPV генетично детермінована та важлива для прогресії
Імунологічна чутливість до інфекції HPV генетично детермінована та важлива для прогресії

Фактори ризику розвитку цервікального раку
Фактори ризику персистенції інфекції HPV
Кількість сексуальних партнерів
Фактори ризику розвитку цервікального раку
Фактори ризику персистенції інфекції HPV
Кількість сексуальних партнерів

Багатостадійність РШМ
Багатостадійність РШМ

Карцинома — вірус інтегрований в геном клітини –поява змінених «атипових» клітин,
Карцинома — вірус інтегрований в геном клітини –поява змінених «атипових» клітин,
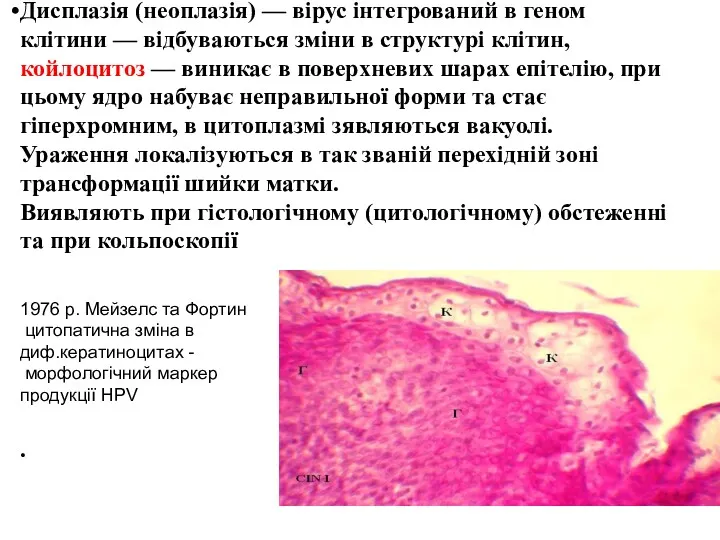
Дисплазія (неоплазія) — вірус інтегрований в геном клітини — відбуваються зміни
Дисплазія (неоплазія) — вірус інтегрований в геном клітини — відбуваються зміни
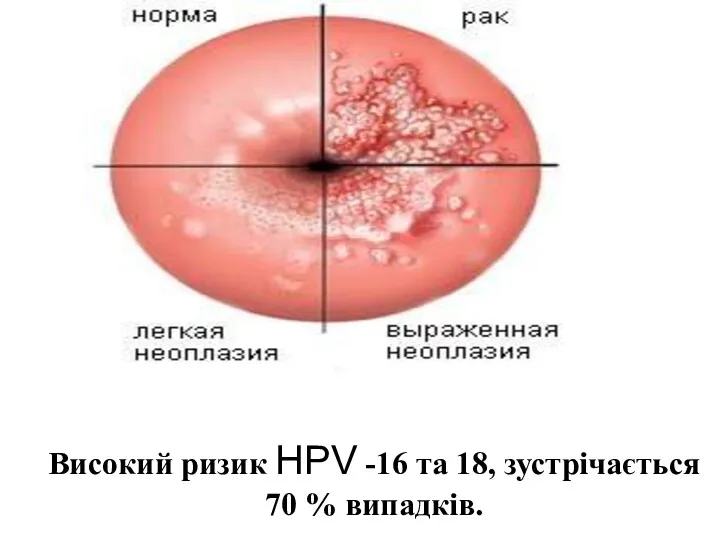
Високий ризик HPV -16 та 18, зустрічається 70 % випадків.
Високий ризик HPV -16 та 18, зустрічається 70 % випадків.
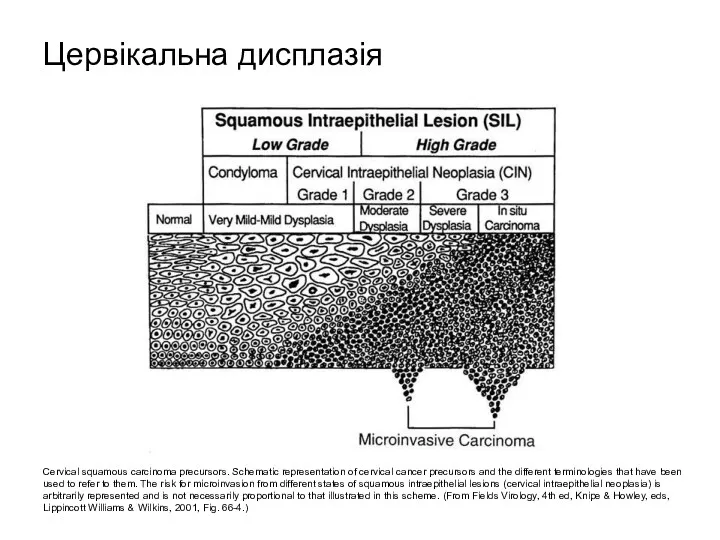
Cervical squamous carcinoma precursors. Schematic representation of cervical cancer precursors and
Cervical squamous carcinoma precursors. Schematic representation of cervical cancer precursors and
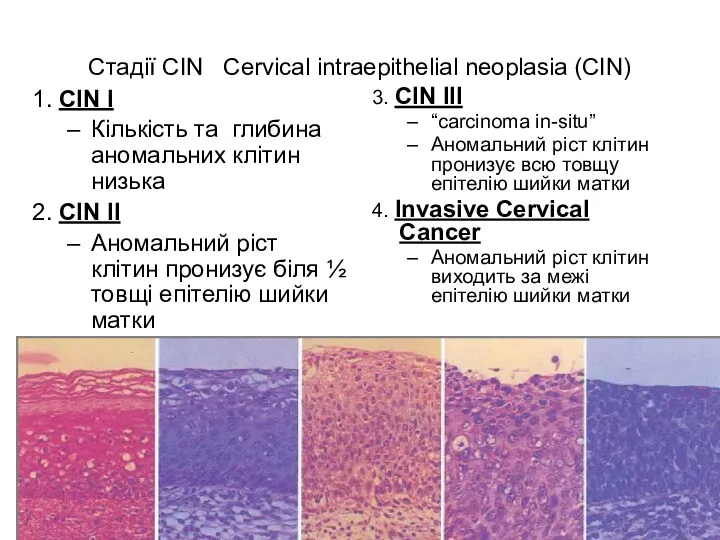
Стадії CIN Cervical intraepithelial neoplasia (CIN)
1. CIN I
Кількість та глибина аномальних
Стадії CIN Cervical intraepithelial neoplasia (CIN)
1. CIN I
Кількість та глибина аномальних
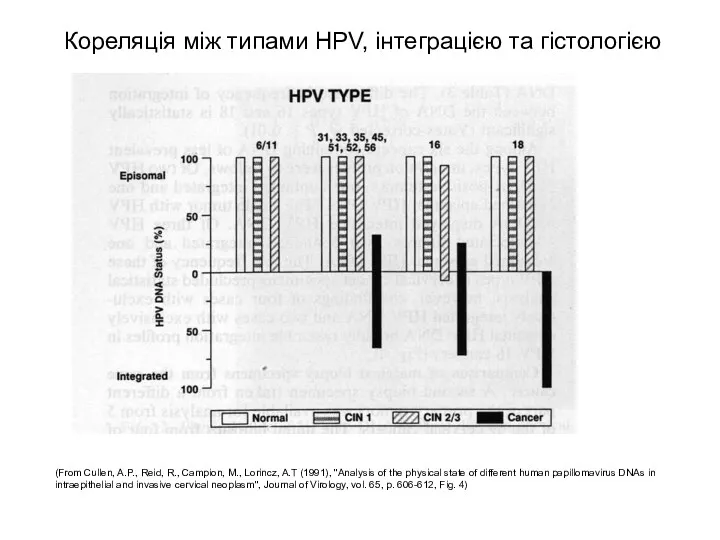
Кореляція між типами HPV, інтеграцією та гістологією
(From Cullen, A.P., Reid, R.,
Кореляція між типами HPV, інтеграцією та гістологією
(From Cullen, A.P., Reid, R.,

Вимоги для трансформації
Персистуюча інфекція – реплікація вірусу
Експресія вірусних білків
Мутації в клітині
Пeрсистуюча
Вимоги для трансформації
Персистуюча інфекція – реплікація вірусу
Експресія вірусних білків
Мутації в клітині
Пeрсистуюча

Клітинні фактори та злоякісність HPV
Невеликий % розвитку раку
Тривалий латентний період
Стадії прогресії
Клітинні фактори та злоякісність HPV
Невеликий % розвитку раку
Тривалий латентний період
Стадії прогресії

Головна відмінність HPV від інших вірусів –можливість організму позбавитись від нього
Головна відмінність HPV від інших вірусів –можливість організму позбавитись від нього
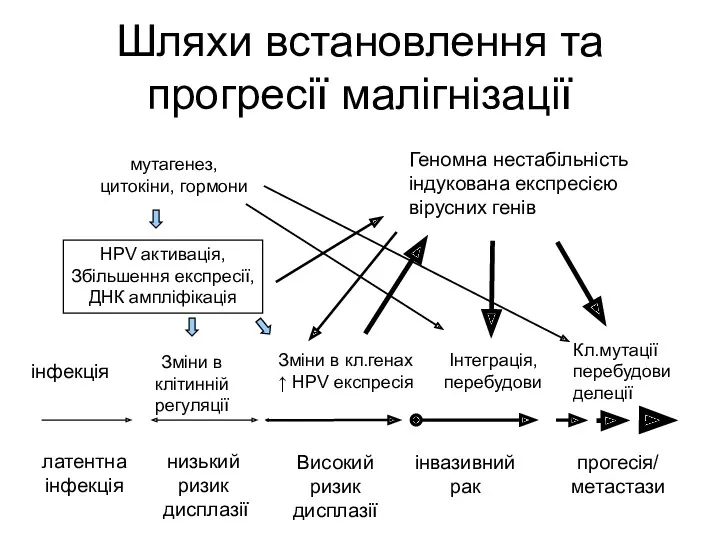
Шляхи встановлення та прогресії малігнізації
інфекція
Зміни в клітинній регуляції
Зміни в кл.генах
Шляхи встановлення та прогресії малігнізації
інфекція
Зміни в клітинній регуляції
Зміни в кл.генах
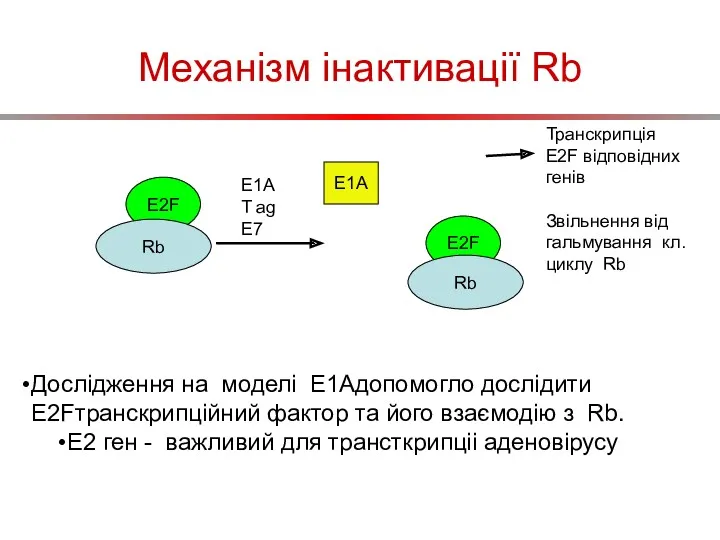
Механізм інактивації Rb
E2F
Rb
E1A
T ag
E7
E2F
Rb
E1A
Транскрипція E2F відповідних генів
Звільнення від гальмування кл.циклу
Механізм інактивації Rb
E2F
Rb
E1A
T ag
E7
E2F
Rb
E1A
Транскрипція E2F відповідних генів
Звільнення від гальмування кл.циклу
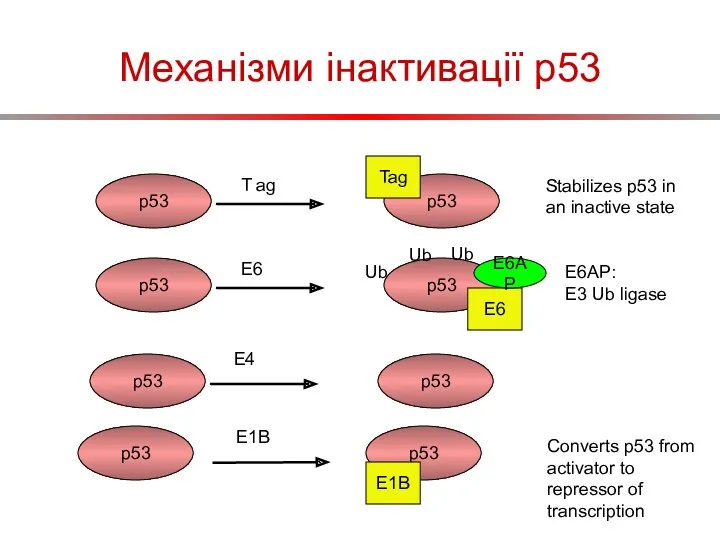
Механізми інактивації p53
p53
T ag
p53
Tag
Stabilizes p53 in an inactive state
p53
E6
p53
E6
E6AP
Ub
Ub
Ub
E6AP:
E3 Ub
Механізми інактивації p53
p53
T ag
p53
Tag
Stabilizes p53 in an inactive state
p53
E6
p53
E6
E6AP
Ub
Ub
Ub
E6AP:
E3 Ub

ПІДСУМОК: HPV
Структура
Невеликий (8 kb) циркулярний длДНК геном, голий капсид
Хвороби
Бородавки
ПІДСУМОК: HPV
Структура
Невеликий (8 kb) циркулярний длДНК геном, голий капсид
Хвороби
Бородавки
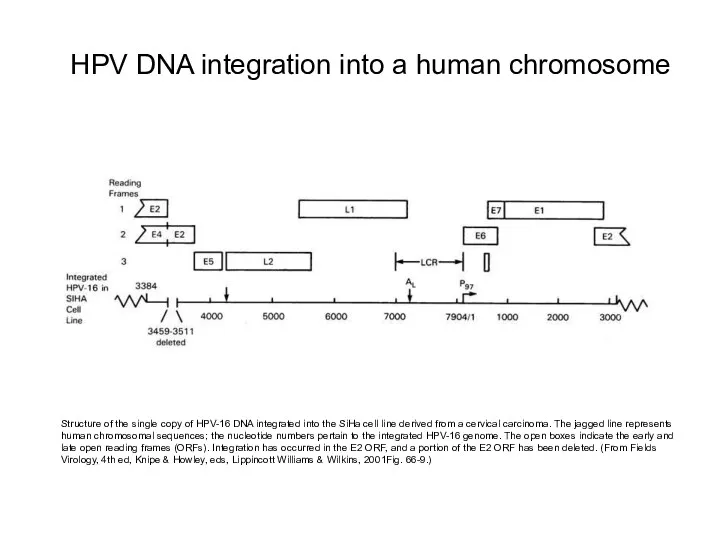
Structure of the single copy of HPV-16 DNA integrated into the
Structure of the single copy of HPV-16 DNA integrated into the
 Мышцы верхней части спины, области плечевого сустава и плеча
Мышцы верхней части спины, области плечевого сустава и плеча Введение в фармакогнозию. Лекция №1
Введение в фармакогнозию. Лекция №1 Antibiotics having a β-lactam ring
Antibiotics having a β-lactam ring Балалардағы тыныс алу жолдарының бөгде заттары
Балалардағы тыныс алу жолдарының бөгде заттары Вирусные инфекции. Задачи
Вирусные инфекции. Задачи Дезинфекция. Понятия, виды, методы дезинфекции
Дезинфекция. Понятия, виды, методы дезинфекции Регуляция кровообращения
Регуляция кровообращения Лазерная терапия
Лазерная терапия Дифференциальная диагностика воспалительных миопатий
Дифференциальная диагностика воспалительных миопатий Период младенчества (грудного возраста). Теоретическое занятие
Период младенчества (грудного возраста). Теоретическое занятие Плоскостопие у детей и эффективные методы его устранения на уроках физической культуры
Плоскостопие у детей и эффективные методы его устранения на уроках физической культуры Синдром гипотиреоза
Синдром гипотиреоза Статеві хвороби та їх запобігання
Статеві хвороби та їх запобігання Кровотечения из пищеварительного тракта
Кровотечения из пищеварительного тракта Фозинприл вива фарм 20мг таблеткаларының өндірісін ұйымдастыру
Фозинприл вива фарм 20мг таблеткаларының өндірісін ұйымдастыру Генетически обусловленные формы умственной отсталости
Генетически обусловленные формы умственной отсталости Лекарственные растения в огороде. 4 класс
Лекарственные растения в огороде. 4 класс Аккредитация
Аккредитация Психодисциплины. Психиатрия. Психотерапия. Медицинская психология
Психодисциплины. Психиатрия. Психотерапия. Медицинская психология Гіповітаміноз. Рахіт. Гіпервітаміноз вітаміну Д. Гіпокальціємічний синдром. Аномалії конституції
Гіповітаміноз. Рахіт. Гіпервітаміноз вітаміну Д. Гіпокальціємічний синдром. Аномалії конституції Острые отравления парацетамолом
Острые отравления парацетамолом Первая медицинская помощь (дидактический материал для подготовки спасателей)
Первая медицинская помощь (дидактический материал для подготовки спасателей) Малоинвазивное лечение лимфаденитов
Малоинвазивное лечение лимфаденитов Средства, действующие на адренергические синапсы
Средства, действующие на адренергические синапсы Нарушения памяти в неврологической практике
Нарушения памяти в неврологической практике Грипп. Возбудители гриппа. Пути передачи. Виды профилактики
Грипп. Возбудители гриппа. Пути передачи. Виды профилактики Домедична допомога в екстремальних умовах. Травми грудної клітки. Пневмоторакс. Гемоторакс. Переломи ребер
Домедична допомога в екстремальних умовах. Травми грудної клітки. Пневмоторакс. Гемоторакс. Переломи ребер Кровь – жизненно важная система организма ( функции)
Кровь – жизненно важная система организма ( функции)