- Physiology of kidneys

Содержание
- 2. The Nephron Is the Functional Unit of the Kidney Each kidney in the human contains about
- 3. Each nephron contains (1) a tuft of glomerular capillaries called the glomerulus, through which large amounts
- 5. The macula densa plays an important role in controlling nephron function. Beyond the macula densa, fluid
- 6. Renal Blood Supply Blood flow to the two kidneys is normally about 22 per cent of
- 8. PHYSIOLOGIC CONTROL OF GLOMERULAR FILTRATION AND RENAL BLOOD FLOW The determinants of GFR that are most
- 9. Sympathetic Nervous System Activation Decreases GFR Strong activation of the renal sympathetic nerves can constrict the
- 10. Hormonal and Autacoid Control of Renal Circulation Norepinephrine, Epinephrine, and Endothelin Constrict Renal Blood Vessels and
- 11. Angiotensin II Constricts Efferent Arterioles A powerful renal vasoconstrictor, angiotensin II, can be considered as a
- 13. Endothelial-Derived Nitric Oxide Decreases Renal Vascular Resistance and Increases GFR A basal level of nitric oxide
- 14. Prostaglandins and Bradykinin Tend to Increase GFR Hormones and autacoids that cause vasodilation and increased renal
- 15. Function of nephrone Video
- 16. AUTOREGULATION OF GFR AND RENAL BLOOD FLOW Feedback mechanisms intrinsic to the kidneys normally keep the
- 17. Myogenic Autoregulation of Renal Blood Flow and GFR A second mechanism that contributes to the maintenance
- 18. URINE FORMATION The rates at which different substances are excreted in the urine represent the sum
- 19. Urine formation begins with filtration from the glomerular capillaries into Bowman's capsule of a large amount
- 20. Why Are Large Amounts of Solutes Filtered and Then Reabsorbed by the Kidneys? One advantage of
- 21. Glomerular Capillary Membrane The glomerular capillary membrane is similar to that of other capillaries, except that
- 22. Glomerular Capillary Membrane Although the fenestrations are relatively large, endothelial cells are richly endowed with fixed
- 23. Podocytes The final part of the glomerular membrane is a layer of epithelial cells (podocytes) that
- 25. Three basic renal processes The substance is freely filtered but is also partly reabsorbed from the
- 26. Filtration, Reabsorption, and Secretion of Different Substances In general, tubular, reabsorption is quantitatively more important than
- 28. Filtration, Reabsorption, and Secretion of Different Substances Nutritional substances, such as amino acids and glucose, are
- 29. Tubular reabsorption
- 30. Tubular secretion
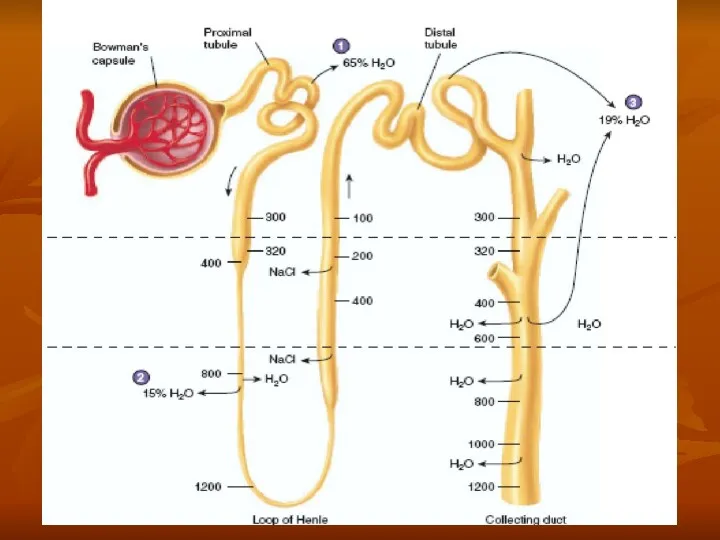
- 31. Countercurrent mecanism and concentration of urine
- 34. MULTIPLE FUNCTIONS OF THE KIDNEYS IN HOMEOSTASIS Excretion of metabolic waste products and foreign chemicals Regulation
- 35. Excretion of Metabolic Waste Products, Foreign Chemicals, Drugs, and Hormone Metabolites The kidneys are the primary
- 36. Regulation of Water and Electrolyte Balances For maintenance of homeostasis, excretion of water and electrolytes must
- 37. Regulation of Arterial Pressure In addition, the kidneys contribute to short-term arterial pressure regulation by secreting
- 39. Regulation of Acid-Base Balance The kidneys contribute to acid-base regulation, along with the lungs and body
- 40. Regulation of 1,25-Dihydroxy Vitamin D 3 Production The kidneys produce the active form of vitamin D,
- 41. Glucose Synthesis The kidneys synthesize glucose from amino acids and other precursors during prolonged fasting, a
- 42. BASIC PRINCIPLES OF OSMOSIS AND OSMOTIC PRESSURE Osmosis is' the net diffusion of water across a
- 43. Isosmotic, Hyperosmotic, and Hypo-osmotic Fluids Solutions with an osmolarity the same as the cell are called
- 44. OSMORECEPTOR-ADH FEEDBACK SYSTEM 1. An increase in extracellular fluid osmolarity causes the special nerve cells called
- 46. ADH Synthesis in Supraoptic and Paraventricular Nuclei of the Hypothalamus and ADH Release from the Posterior
- 47. A second neuronal area A second neuronal area important in controlling osmolarity and ADH secretion is
- 48. ROLE OF THIRST IN CONTROLLING EXTRACELLULAR FLUID OSMOLARITY AND SODIUM CONCENTRATION The kidneys minimize fluid loss
- 49. Central Nervous System Centers for Thirst Located anterolaterally in the preoptic nucleus is another small area
- 50. Stimuli for Thirst One of the most important is increased extracellular fluid osmolarity, which causes intracellular
- 51. Stimuli for Thirst These regions are outside the blood-brain barrier, and peptides such as angiotensin II
- 52. Threshold for Osmolar Stimulus of Drinking The kidneys must continually excrete at least some fluid, even
- 53. Cardiovascular Reflex Stimulation of ADH Release by Decreased Arterial Pressure and/or Decreased Blood Volume ADH release
- 55. Role of Angiotensin II and Aldosterone in Controlling Extracellular Fluid Osmolarity and Sodium Concentration Both angiotensin
- 57. SALT-APPETITE MECHANISM FOR CONTROLLING EXTRACELLULAR FLUID SODIUM CONCENTRATION AND VOLUME Maintenance of normal extracellular fluid volume
- 59. Скачать презентацию
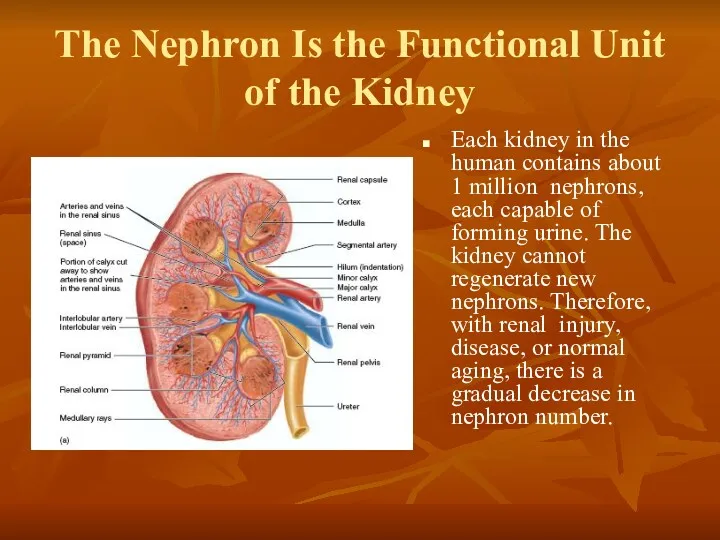
The Nephron Is the Functional Unit
of the Kidney
Each kidney in
The Nephron Is the Functional Unit
of the Kidney
Each kidney in
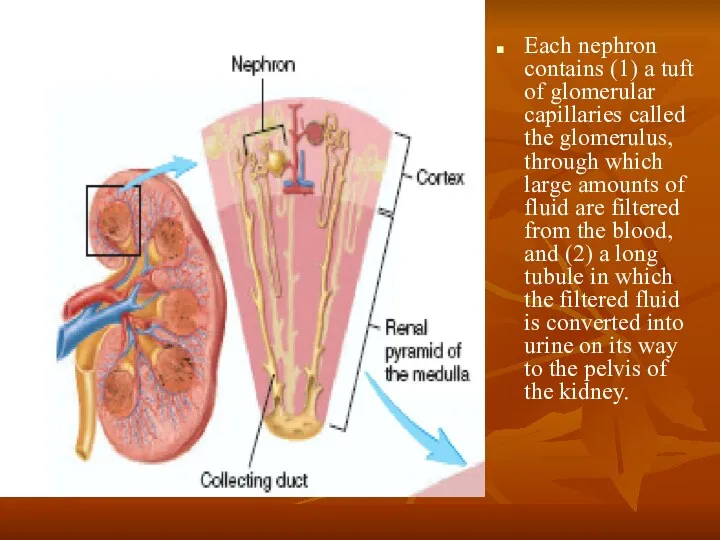
Each nephron contains (1) a tuft of glomerular capillaries called the
Each nephron contains (1) a tuft of glomerular capillaries called the
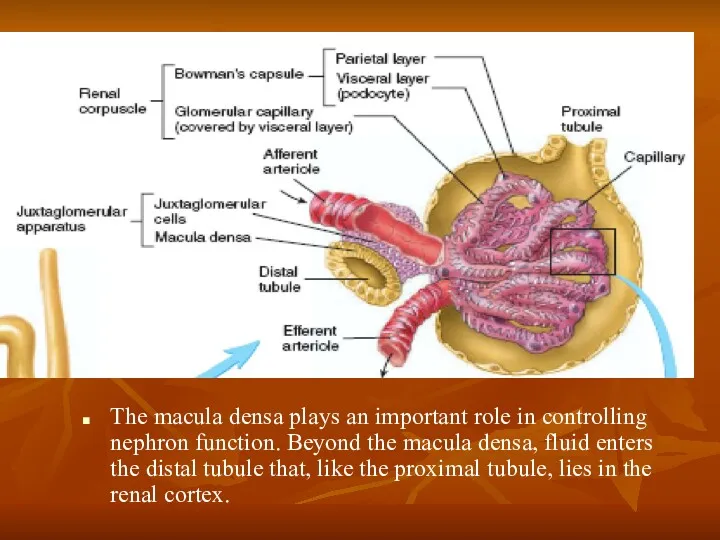
The macula densa plays an important role in controlling nephron function.
The macula densa plays an important role in controlling nephron function.

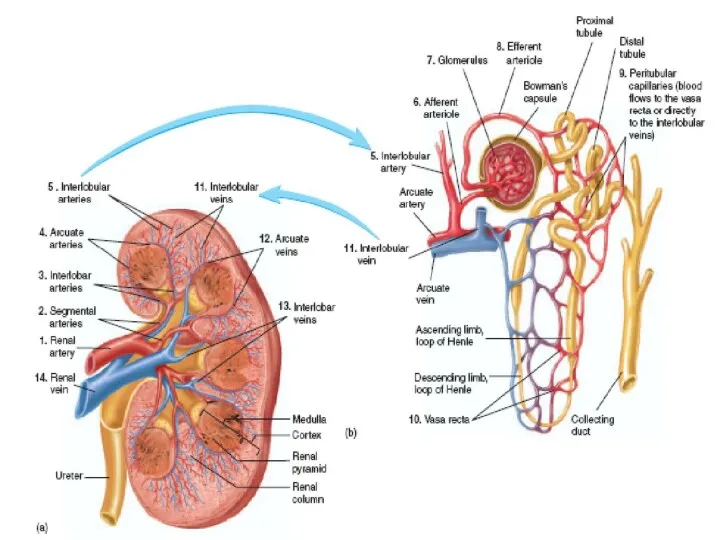
Renal Blood Supply
Blood flow to the two kidneys is normally about
Renal Blood Supply
Blood flow to the two kidneys is normally about
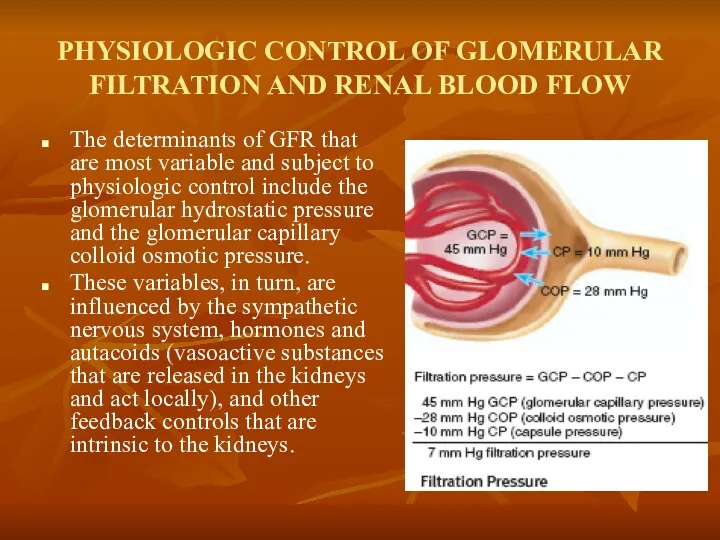
PHYSIOLOGIC CONTROL OF GLOMERULAR FILTRATION AND RENAL BLOOD FLOW
The determinants of
PHYSIOLOGIC CONTROL OF GLOMERULAR FILTRATION AND RENAL BLOOD FLOW
The determinants of

Sympathetic Nervous System Activation Decreases GFR
Strong activation of the renal sympathetic
Sympathetic Nervous System Activation Decreases GFR
Strong activation of the renal sympathetic

Hormonal and Autacoid Control of Renal Circulation
Norepinephrine, Epinephrine, and Endothelin Constrict
Hormonal and Autacoid Control of Renal Circulation
Norepinephrine, Epinephrine, and Endothelin Constrict

Angiotensin II Constricts Efferent Arterioles
A powerful renal vasoconstrictor, angiotensin II, can
Angiotensin II Constricts Efferent Arterioles
A powerful renal vasoconstrictor, angiotensin II, can
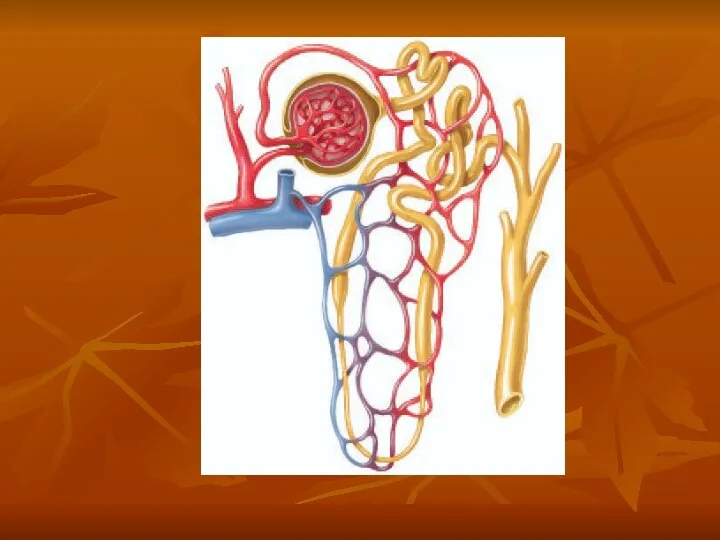

Endothelial-Derived Nitric Oxide Decreases Renal Vascular Resistance and Increases GFR
A basal
Endothelial-Derived Nitric Oxide Decreases Renal Vascular Resistance and Increases GFR
A basal

Prostaglandins and Bradykinin Tend to Increase GFR
Hormones and autacoids that cause
Prostaglandins and Bradykinin Tend to Increase GFR
Hormones and autacoids that cause
Function of nephrone Video
Function of nephrone Video

AUTOREGULATION OF GFR AND RENAL BLOOD FLOW
Feedback mechanisms intrinsic to the
AUTOREGULATION OF GFR AND RENAL BLOOD FLOW
Feedback mechanisms intrinsic to the

Myogenic Autoregulation of Renal Blood Flow and GFR
A second mechanism that
Myogenic Autoregulation of Renal Blood Flow and GFR
A second mechanism that
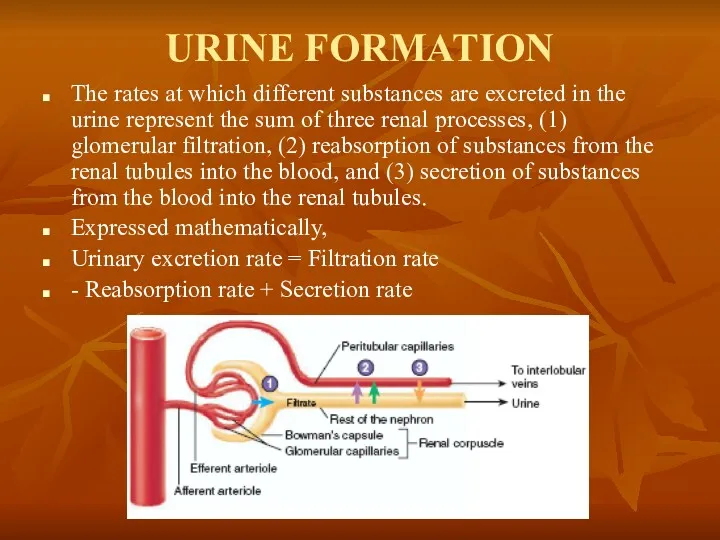
URINE FORMATION
The rates at which different substances are excreted in the
URINE FORMATION
The rates at which different substances are excreted in the
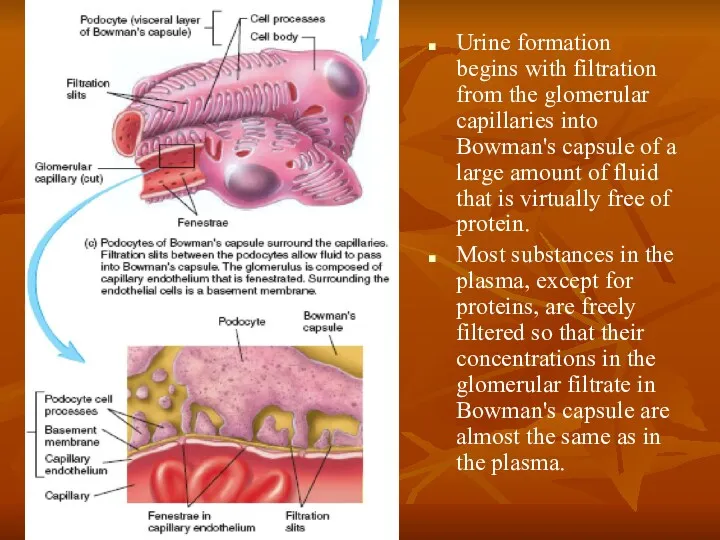
Urine formation begins with filtration from the glomerular capillaries into Bowman's
Urine formation begins with filtration from the glomerular capillaries into Bowman's

Why Are Large Amounts of Solutes Filtered and Then Reabsorbed by
Why Are Large Amounts of Solutes Filtered and Then Reabsorbed by

Glomerular Capillary Membrane
The glomerular capillary membrane is similar to that of
Glomerular Capillary Membrane
The glomerular capillary membrane is similar to that of

Glomerular Capillary Membrane
Although the fenestrations are relatively large, endothelial cells are
Glomerular Capillary Membrane
Although the fenestrations are relatively large, endothelial cells are
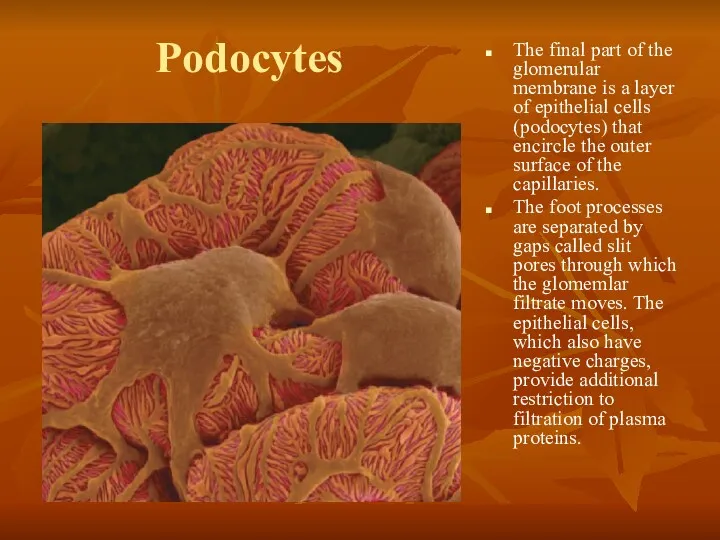
Podocytes
The final part of the glomerular membrane is a layer of
Podocytes
The final part of the glomerular membrane is a layer of
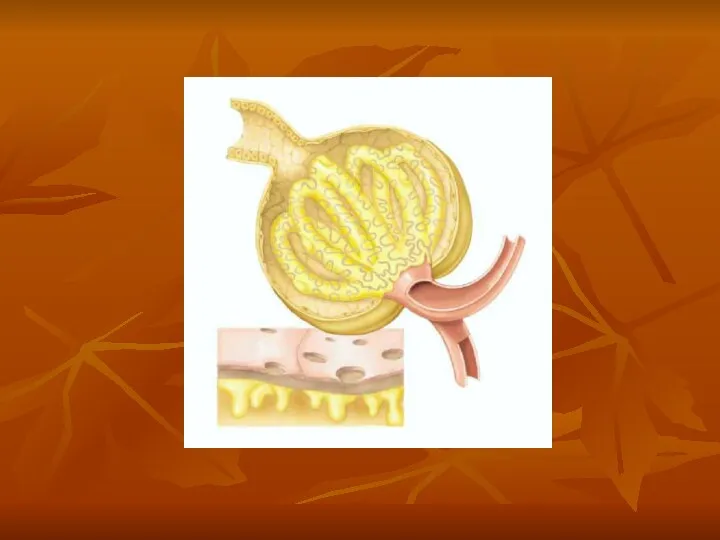

Three basic renal processes
The substance is freely filtered but is also
Three basic renal processes
The substance is freely filtered but is also

Filtration, Reabsorption, and Secretion of Different Substances
In general, tubular, reabsorption is
Filtration, Reabsorption, and Secretion of Different Substances
In general, tubular, reabsorption is
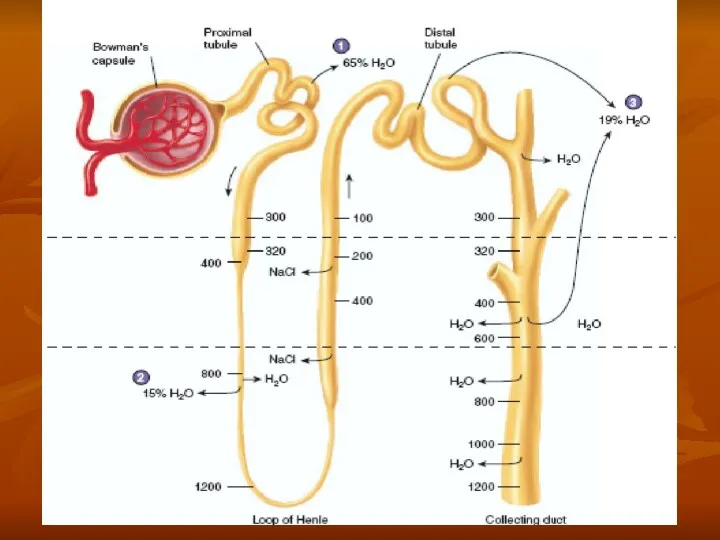

Filtration, Reabsorption, and Secretion of Different Substances
Nutritional substances, such as amino
Filtration, Reabsorption, and Secretion of Different Substances
Nutritional substances, such as amino
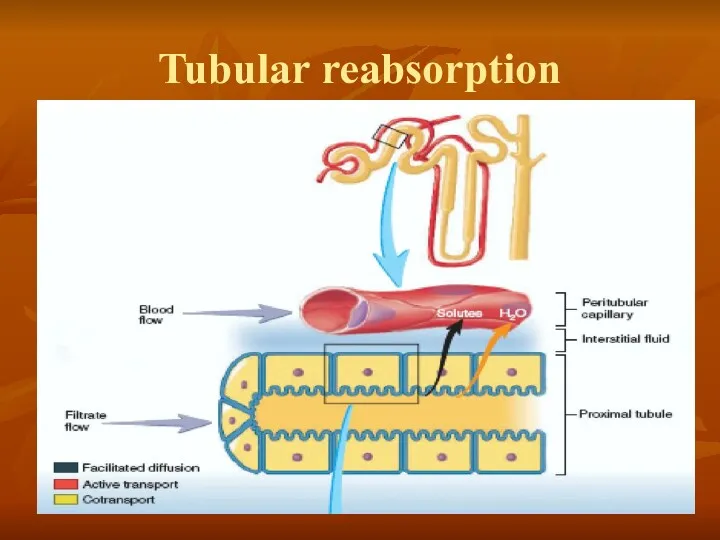
Tubular reabsorption
Tubular reabsorption
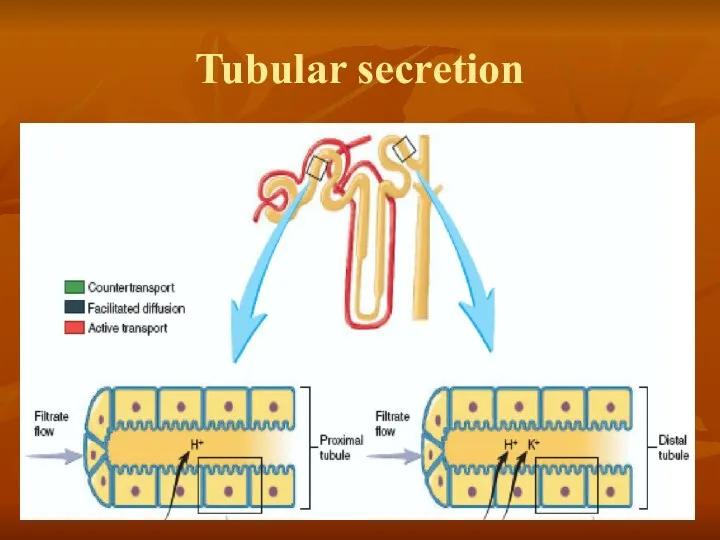
Tubular secretion
Tubular secretion
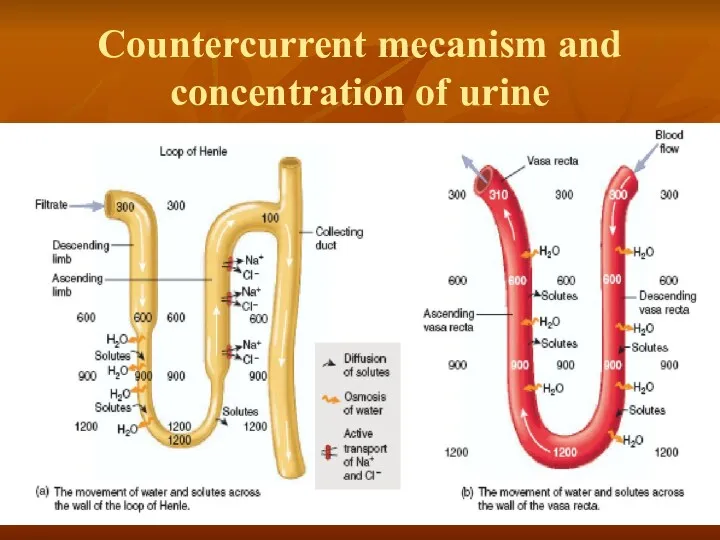
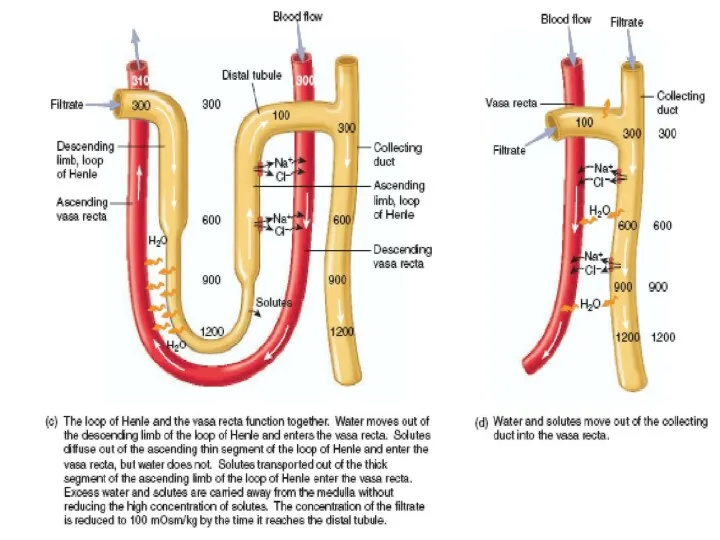
Countercurrent mecanism and concentration of urine
Countercurrent mecanism and concentration of urine
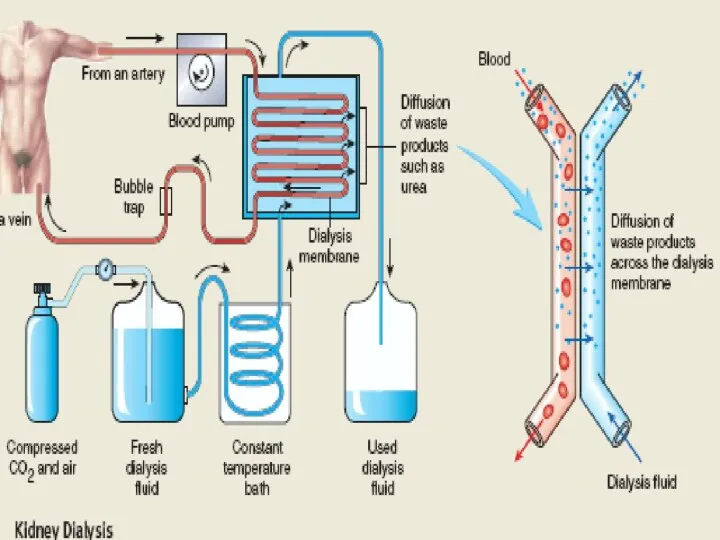

MULTIPLE FUNCTIONS OF THE KIDNEYS IN HOMEOSTASIS
Excretion of metabolic waste products
MULTIPLE FUNCTIONS OF THE KIDNEYS IN HOMEOSTASIS
Excretion of metabolic waste products

Excretion of Metabolic Waste Products, Foreign Chemicals, Drugs, and Hormone Metabolites
The
Excretion of Metabolic Waste Products, Foreign Chemicals, Drugs, and Hormone Metabolites
The

Regulation of Water and Electrolyte Balances
For maintenance of homeostasis, excretion of
Regulation of Water and Electrolyte Balances
For maintenance of homeostasis, excretion of
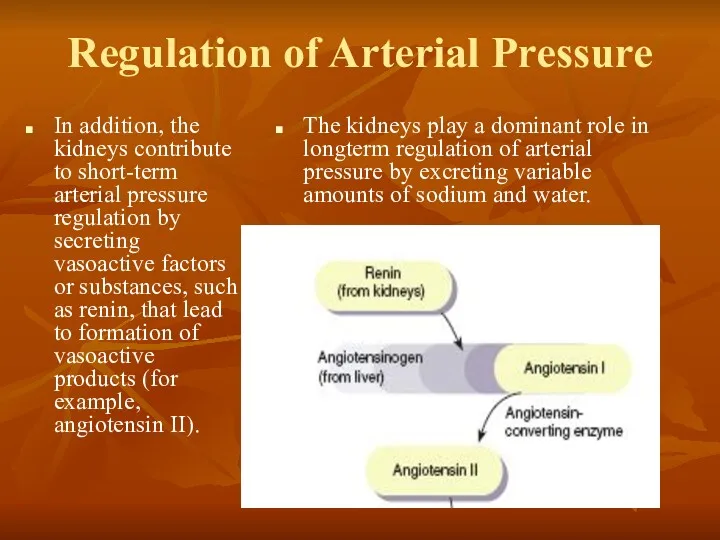
Regulation of Arterial Pressure
In addition, the kidneys contribute to short-term arterial
Regulation of Arterial Pressure
In addition, the kidneys contribute to short-term arterial
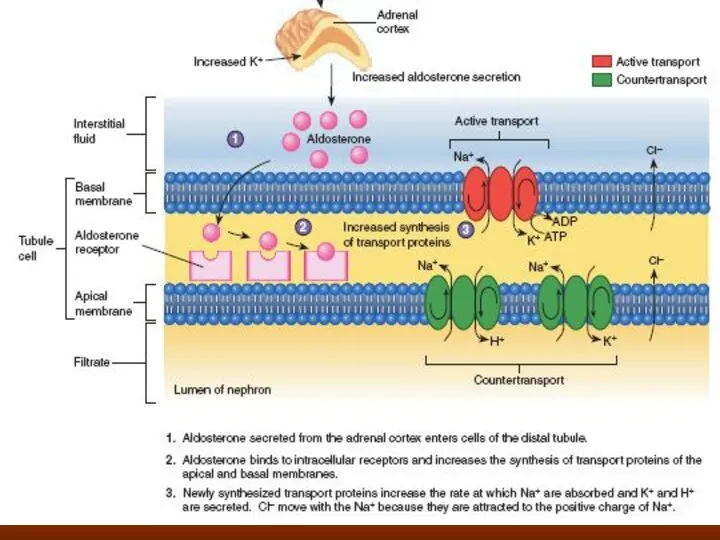

Regulation of Acid-Base Balance
The kidneys contribute to acid-base regulation, along with
Regulation of Acid-Base Balance
The kidneys contribute to acid-base regulation, along with

Regulation of 1,25-Dihydroxy Vitamin D 3 Production
The kidneys produce the active
Regulation of 1,25-Dihydroxy Vitamin D 3 Production
The kidneys produce the active

Glucose Synthesis
The kidneys synthesize glucose from amino acids and other precursors
Glucose Synthesis
The kidneys synthesize glucose from amino acids and other precursors

BASIC PRINCIPLES OF OSMOSIS AND OSMOTIC PRESSURE
Osmosis is' the net diffusion
BASIC PRINCIPLES OF OSMOSIS AND OSMOTIC PRESSURE
Osmosis is' the net diffusion

Isosmotic, Hyperosmotic, and Hypo-osmotic Fluids
Solutions with an osmolarity the same as
Isosmotic, Hyperosmotic, and Hypo-osmotic Fluids
Solutions with an osmolarity the same as

OSMORECEPTOR-ADH FEEDBACK SYSTEM
1. An increase in extracellular fluid osmolarity causes the
OSMORECEPTOR-ADH FEEDBACK SYSTEM
1. An increase in extracellular fluid osmolarity causes the
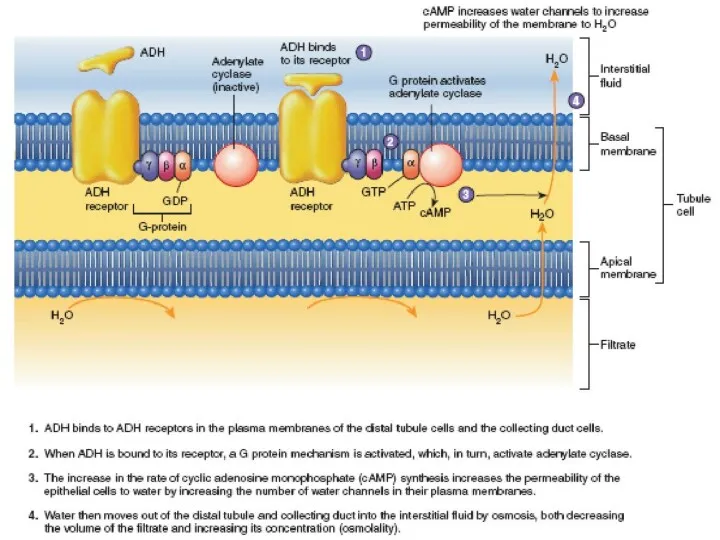

ADH Synthesis in Supraoptic and Paraventricular Nuclei of the Hypothalamus and
ADH Synthesis in Supraoptic and Paraventricular Nuclei of the Hypothalamus and
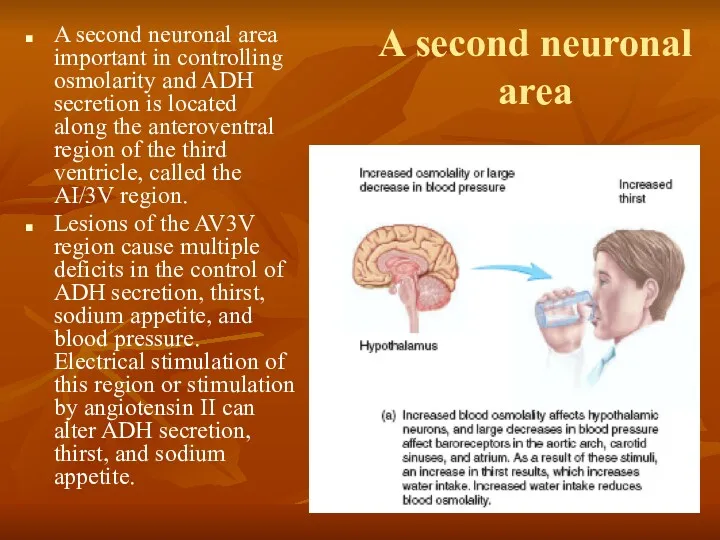
A second neuronal area
A second neuronal area important in controlling osmolarity
A second neuronal area
A second neuronal area important in controlling osmolarity

ROLE OF THIRST IN CONTROLLING EXTRACELLULAR FLUID OSMOLARITY AND
SODIUM CONCENTRATION
The
ROLE OF THIRST IN CONTROLLING EXTRACELLULAR FLUID OSMOLARITY AND
SODIUM CONCENTRATION
The

Central Nervous System Centers for Thirst
Located anterolaterally in the preoptic nucleus
Central Nervous System Centers for Thirst
Located anterolaterally in the preoptic nucleus

Stimuli for Thirst
One of the most important is increased extracellular fluid
Stimuli for Thirst
One of the most important is increased extracellular fluid

Stimuli for Thirst
These regions are outside the blood-brain barrier, and peptides
Stimuli for Thirst
These regions are outside the blood-brain barrier, and peptides

Threshold for Osmolar Stimulus of Drinking
The kidneys must continually excrete at
Threshold for Osmolar Stimulus of Drinking
The kidneys must continually excrete at
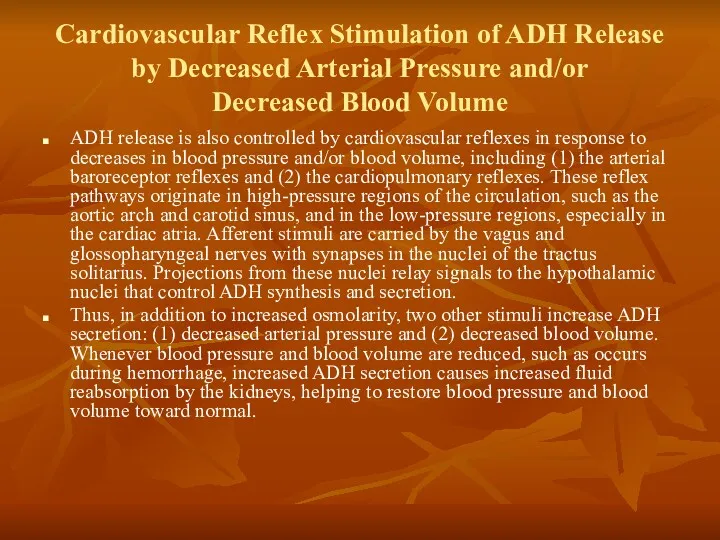
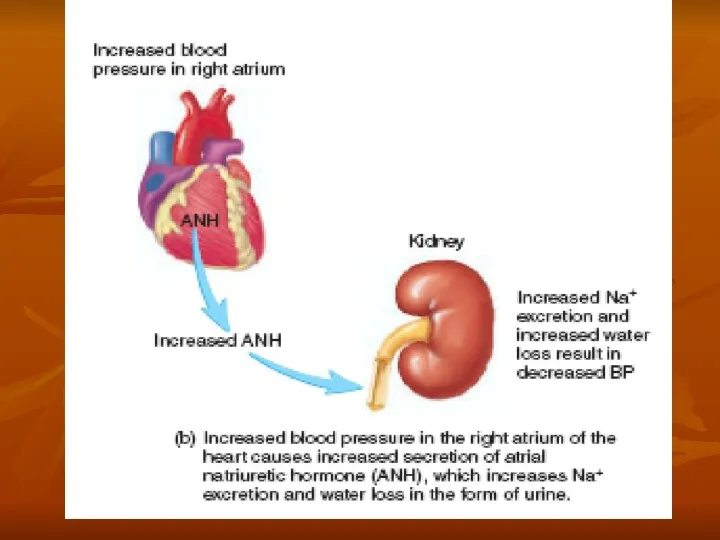
Cardiovascular Reflex Stimulation of ADH Release by Decreased Arterial Pressure and/or
Cardiovascular Reflex Stimulation of ADH Release by Decreased Arterial Pressure and/or

Role of Angiotensin II and Aldosterone in Controlling Extracellular Fluid Osmolarity
Role of Angiotensin II and Aldosterone in Controlling Extracellular Fluid Osmolarity
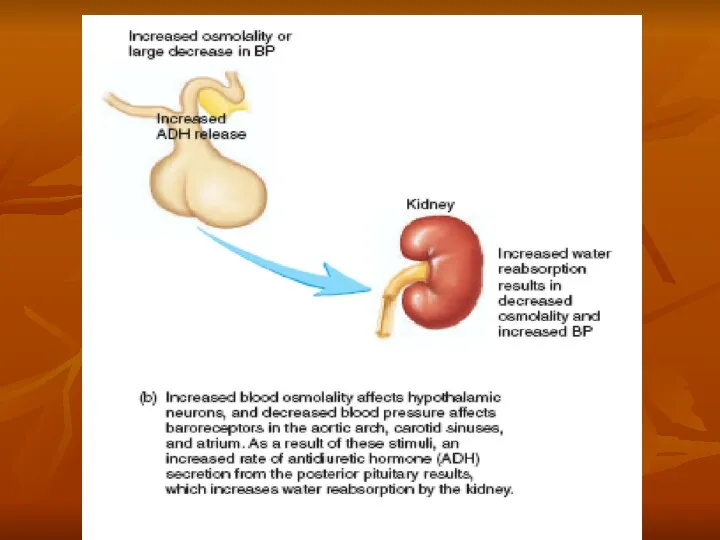

SALT-APPETITE MECHANISM FOR CONTROLLING EXTRACELLULAR FLUID SODIUM CONCENTRATION AND VOLUME
Maintenance of
SALT-APPETITE MECHANISM FOR CONTROLLING EXTRACELLULAR FLUID SODIUM CONCENTRATION AND VOLUME
Maintenance of
 Санитариялық-гигиеналық зертхана жұмысын ұйымдастыру
Санитариялық-гигиеналық зертхана жұмысын ұйымдастыру Отит у детей
Отит у детей Грыжи брюшной стенки
Грыжи брюшной стенки Функциональная недостаточность мышц челюстно-лицевой области как фактор развития аномалий
Функциональная недостаточность мышц челюстно-лицевой области как фактор развития аномалий Центральные органы кроветворения. Красный костный мозг, тимус
Центральные органы кроветворения. Красный костный мозг, тимус Денситометрия и остеопороз
Денситометрия и остеопороз Балалардағы асқазан мен он екі елі ішектің жара аурулары. Синдромдық негізде диагностика, ем және профилактика жүргізу
Балалардағы асқазан мен он екі елі ішектің жара аурулары. Синдромдық негізде диагностика, ем және профилактика жүргізу Cеборея. Акне
Cеборея. Акне Заболевания, диагностируемые неонатальным скринингом
Заболевания, диагностируемые неонатальным скринингом Лекарственные средства, влияющие на систему крови
Лекарственные средства, влияющие на систему крови Система гемостаза
Система гемостаза Өсіргіш заттар. Өсіргіш заттардың өсімдіктерге әсер ету механизмі. Ауксин әсері
Өсіргіш заттар. Өсіргіш заттардың өсімдіктерге әсер ету механизмі. Ауксин әсері Фармакокинетика лекарственных средств: пути введения, всасывание, превращение лекарств и выведение их из организма
Фармакокинетика лекарственных средств: пути введения, всасывание, превращение лекарств и выведение их из организма Лечебные и профилактические зубные пасты
Лечебные и профилактические зубные пасты Биохимические исследования. Занятие 12
Биохимические исследования. Занятие 12 Клиника интеллектуальных нарушений при олигофрении
Клиника интеллектуальных нарушений при олигофрении Комлексное лечение скелетной формы мезиальной окклюзии
Комлексное лечение скелетной формы мезиальной окклюзии Послеродовый период и его патология
Послеродовый период и его патология Регуляция иммунного ответа. Гормональная регуляция иммунных реакций. Система цитокинов. Интерфероны. Факторы роста
Регуляция иммунного ответа. Гормональная регуляция иммунных реакций. Система цитокинов. Интерфероны. Факторы роста Лейкоплакия полости рта
Лейкоплакия полости рта Острый и хронический бронхит. Хроническая обструктивная легочная болезнь. Эмфизема. Астма
Острый и хронический бронхит. Хроническая обструктивная легочная болезнь. Эмфизема. Астма Эпидемиологическая ситуация по особо опасным инфекциям, в т.ч. лихорадке Зика в мире и Российской Федерации
Эпидемиологическая ситуация по особо опасным инфекциям, в т.ч. лихорадке Зика в мире и Российской Федерации Желудочно-кишечные кровотечения
Желудочно-кишечные кровотечения Коррекция эмоциональных нарушений у детей и подростков с детским церебральным параличом
Коррекция эмоциональных нарушений у детей и подростков с детским церебральным параличом Спонтанды пневмоторакс
Спонтанды пневмоторакс Травматизм. Первичная и вторичная профилактика. Первая помощь при травмах
Травматизм. Первичная и вторичная профилактика. Первая помощь при травмах Медицинская этика, понятия, принципы, значимость в медицинской практики. Этические аспекты взаимоотношений врача и пациента
Медицинская этика, понятия, принципы, значимость в медицинской практики. Этические аспекты взаимоотношений врача и пациента История оториноларингологии. Анатомо-физиологические особенности носа и околоносовых пазух. Заболевания носа и ОНП
История оториноларингологии. Анатомо-физиологические особенности носа и околоносовых пазух. Заболевания носа и ОНП