- Подавление программируемой клеточной смерти в опухолевых клетках. Лекция 4

Содержание
- 2. Нестабильность генома (мутации и эпигенетические изменения) Нечувствительность к рост-ингибирующим сигналам (приобретение бессмертия и др.) Постоянная инициация
- 3. ‘Accidental cell death’ (ACD) – «смерть от несчастного случая» (химические, физические или механические повреждения) ‘Regulated cell
- 4. Способы программируемой гибели клеток: Классификация Номенклатурного Комитета (2012) Nomenclature Committee on Cell Death 2012. Cell Death
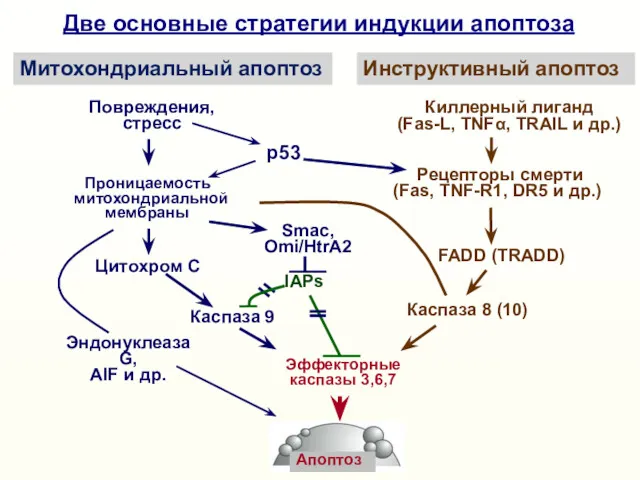
- 5. Для неопластических клеток характерно подавление большинства типов программируемой гибели клеток: Апоптоз Митотическая катастрофа 3. Аутофагия 4.
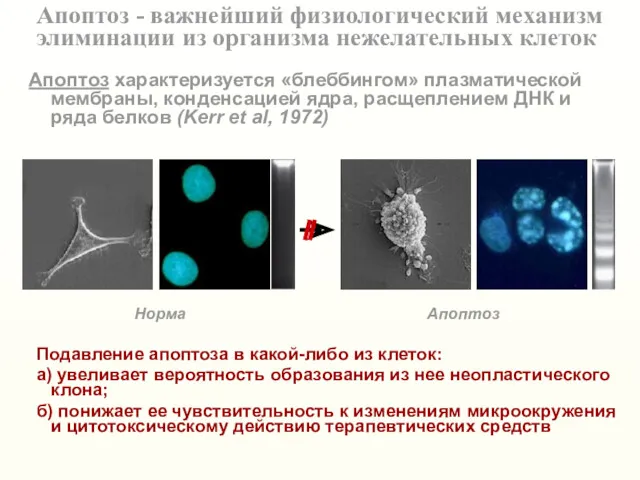
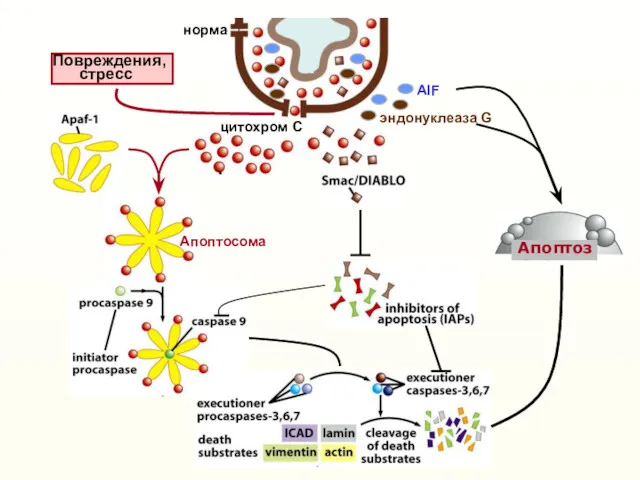
- 6. Апоптоз - важнейший физиологический механизм элиминации из организма нежелательных клеток Норма Апоптоз Апоптоз характеризуется «блеббингом» плазматической
- 7. Погибающие клетки дают сигналы «Найди меня» (лизофосфатидилхолин и др.) и «Съешь меня» (опсонины, связанные с фосфатидилсерином)
- 8. Существенные отличия апоптоза от некроза: а) содержимое погибшей клетки не выходит в межклеточное пространство; апоптотические клетки,
- 9. Апоптоз - важнейший физиологический механизм элиминации из организма нежелательных клеток Подавление апоптоза в какой-либо из клеток:
- 10. Активный тетрамер каспазы Сaйт расщепления Asp-X Расщепление Активный caйт p20 p10 p20 p10 p10 p20 Ключевую
- 11. Эффекторные каспазы 3, 6, 7 «Инструктивный» (extrinsic) апоптоз вызывается активацией «рецепторов смерти» (Death receptors) Киллерный лиганд
- 12. Прокаспаза-8 FADD Fas
- 13. 2 варианта запуска инструктивного пути – рецепторы смерти или рецепторы зависимости Nomenclature Committee on Cell Death
- 14. Эффекторные каспазы 3,6,7 Киллерный лиганд (Fas-L, TNFα, TRAIL и др.) Рецепторы смерти (Fas, TNF-R1, DR5 и
- 16. Nomenclature Committee on Cell Death 2012. Cell Death and Differentiation (2012) 19, 107–120 Intrinsic (mitochondrial) apoptosis
- 17. нечувствительность к апоптогенному действию Fas-L (потеря экспрессии или мутации рецептора смерти Fas, гиперэкспрессия рецептора-ловушки DCR3); потеря
- 18. Онкогены и опухолевые супрессоры регулируют: a) проницаемость митохондриальной мембраны cем. Bcl2 (защитники) Bcl2, Bcl-XL, Bcl-W, Mcl1,
- 19. Онкогены и опухолевые супрессоры регулируют: a) проницаемость митохондриальной мембраны cем. Bcl2 (защитники) Bcl2, Bcl-XL, Bcl-W, Mcl1,
- 20. Онкогены и опухолевые супрессоры регулируют: a) проницаемость митохондриальной мембраны Члены семейства Bcl2 выход из митохондрий цитохрома
- 21. t(14;18) IgH/ Bcl2 гиперэкспрессия Bcl2 (защитник) - В-клеточные лимфомы Мутации/инактивация Bax (эффектор) - Рак толстой кишки,
- 22. Ряд онкогенов и опухолевых супрессоров регулирует экспрессию белков семейства Bcl2 и/или проницаемость митохондриальной мембраны mTOR GSK-3β
- 23. нечувствительность к апоптогенному действию Fas-L (потеря экспрессии или мутации рецептора смерти Fas, гиперэкспрессия рецептора-ловушки DcR3); нарушения
- 24. сурвивин caspase 8 б) активность каспаз (онкогены HER2, Ras, Raf, PI3K, Akt, NFkB, опухолевые супрессоры PTEN,
- 25. t(14;18) IgH/ Bcl2 гиперэкспрессия Bcl2 - В-клеточные лимфомы Мутации/инактивация Bax - Рак толстой кишки, простаты Мутации/инактивация
- 26. Dying tumor cells can use the apoptotic process to generate potent growth-stimulating signals to stimulate the
- 27. Митотическая катастрофа - смерть клетки во время митоза или вследствие его неправильного завершения
- 28. Морфологические признаки митотической катастрофы Клетки хронического миелоидного лейкоза человека K562 (a) Нормальная интерфаза (b) Гигантские многоядерные
- 29. Механизмы индукции митотической катастрофы Mc Gee MM. Mediators Inflamm. 2015:146282 SAC - spindle assembly checkpoint
- 30. Гибель клетки при митотической катастрофе, как правило, происходит апоптоз-подобным путем Преждевременный вход в митоз и/или невозможность
- 31. Отличия некроза от апоптоза
- 32. Некроптоз Апоптоз + АТФ - АТФ Апоптогенные стимулы (TRAIL, TNFα и др.) этил- пируват Переключения апоптоз
- 33. W. Wu et al. Critical Reviews in Oncology/Hematology (2011) Ключевую роль в индукции некроптоза играют белки
- 34. Tait et al., J Cell Sci. 2014;127(Pt 10):2135-44 Механизм реализации некроптоза MLKL - псевдокиназа, фосфорилируется Rip3,
- 35. Аутофагия – программа лизосомальной деградации белков и других компонентов клетки. Индуцируется голоданием (недостатком глюкозы, факторов роста,
- 36. Аутофагия Умеренная (очищение клетки) Сильная Фактор выживания клетки Смерть клетки; выживание организма(?) В.Levine, Nature, 2007, 446:745-747
- 37. Регуляция аутофагии Синтез белка Инициация аутофагии
- 38. Регуляция аутофагии Аутофагия Beclin1 Vps34 Atg1, 13,17 Atg5, 10,12 и др. Lamp2
- 39. Белки семейства Bcl2 регулируют не только апоптоз, но и аутофагию Maiuri et al.., Nature Reviews in
- 40. В.Levine, Nature, 2007, 446:745-747 У мышей гетерозиготный нокаут гена Beclin1 (+/-) резко увеличивает вероятность развития лимфом,
- 41. Kimmelman, Genes & Development, 2011, 25:1999–2010
- 42. Взаимосвязь разных путей клеточной гибели Radogna et al., 2015. http://dx.doi.org/10.1016/j.bcp.2014.12.018
- 43. Лизозомная деградация в поглощенной клетке Поглощенная клетка не погибает и делится Overholtzer et al., Cell, 2007,
- 44. Энтоз индуцирует анеуплоидию Janssen, Medema. Nat Cell Biol. 2011;13(3):199-201
- 45. Роль энтоза в канцерогенезе Sun et al., Cell Res. 2014;24(11):1299-310 and 1288-98; Kroemer, Perfettini. Cell Res.
- 46. Нестабильность генома (мутации и эпигенетические изменения) Нечувствительность к рост-ингибирующим сигналам (приобретение бессмертия и др.) Постоянная инициация
- 48. Скачать презентацию
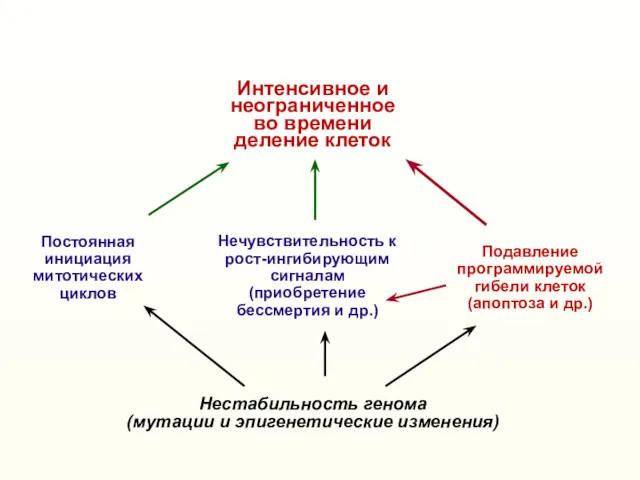
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
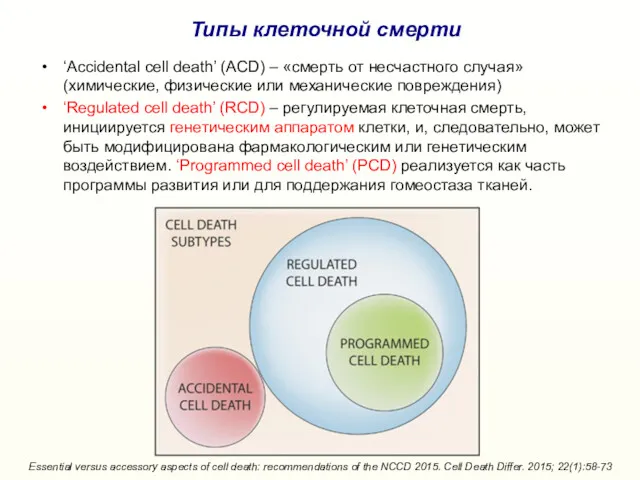
‘Accidental cell death’ (ACD) – «смерть от несчастного случая» (химические, физические
‘Accidental cell death’ (ACD) – «смерть от несчастного случая» (химические, физические

Способы программируемой гибели клеток:
Классификация Номенклатурного Комитета (2012)
Nomenclature Committee on Cell Death
Способы программируемой гибели клеток:
Классификация Номенклатурного Комитета (2012)
Nomenclature Committee on Cell Death

Для неопластических клеток характерно
подавление большинства
типов программируемой гибели клеток:
Апоптоз
Митотическая катастрофа
3.
Для неопластических клеток характерно
подавление большинства
типов программируемой гибели клеток:
Апоптоз
Митотическая катастрофа
3.
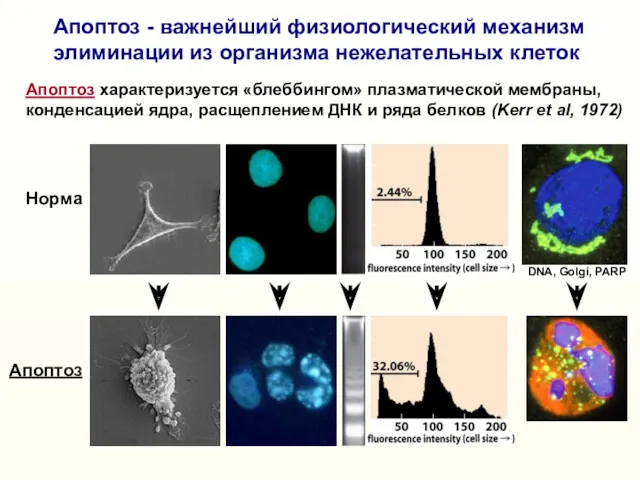
Апоптоз - важнейший физиологический механизм
элиминации из организма нежелательных клеток
Норма
Апоптоз
Апоптоз характеризуется
Апоптоз - важнейший физиологический механизм
элиминации из организма нежелательных клеток
Норма
Апоптоз
Апоптоз характеризуется
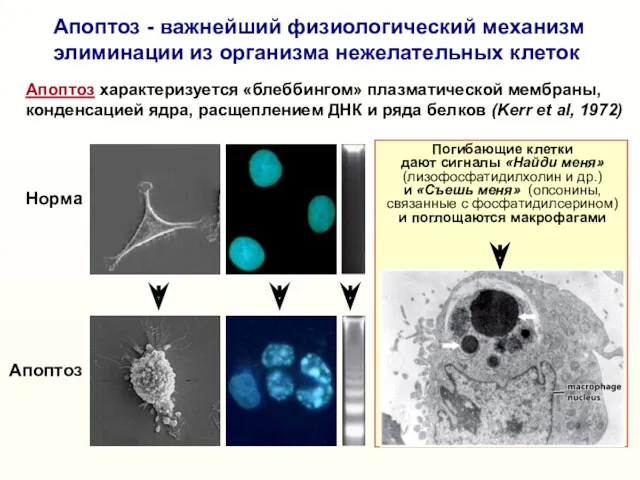
Погибающие клетки
дают сигналы «Найди меня»
(лизофосфатидилхолин и др.)
и «Съешь
Погибающие клетки
дают сигналы «Найди меня»
(лизофосфатидилхолин и др.)
и «Съешь

Существенные отличия апоптоза от некроза:
а) содержимое погибшей клетки не выходит
Существенные отличия апоптоза от некроза:
а) содержимое погибшей клетки не выходит
Апоптоз - важнейший физиологический механизм
элиминации из организма нежелательных клеток
Подавление апоптоза
Апоптоз - важнейший физиологический механизм
элиминации из организма нежелательных клеток
Подавление апоптоза
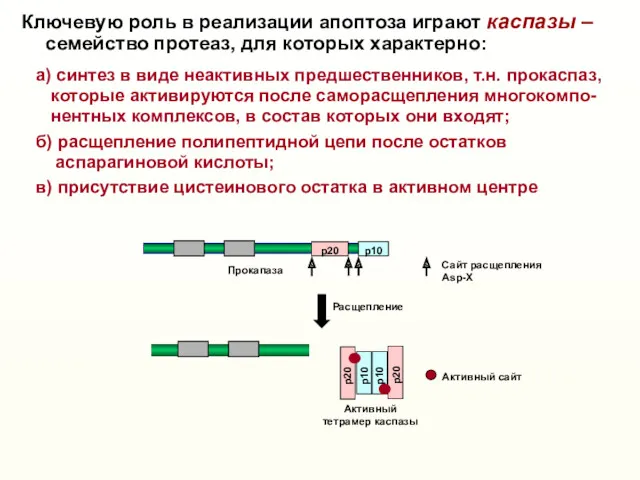
Активный
тетрамер каспазы
Сaйт расщепления Asp-X
Расщепление
Активный caйт
p20
p10
p20
p10
p10
p20
Ключевую роль в реализации апоптоза играют
Активный
тетрамер каспазы
Сaйт расщепления Asp-X
Расщепление
Активный caйт
p20
p10
p20
p10
p10
p20
Ключевую роль в реализации апоптоза играют
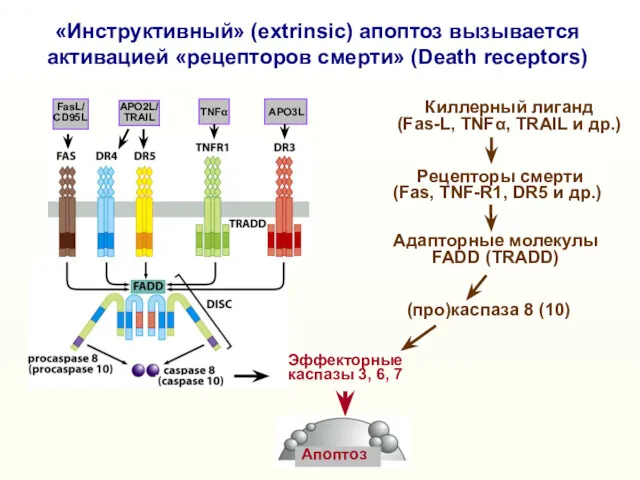
Эффекторные
каспазы 3, 6, 7
«Инструктивный» (extrinsic) апоптоз вызывается
активацией «рецепторов смерти»
Эффекторные
каспазы 3, 6, 7
«Инструктивный» (extrinsic) апоптоз вызывается
активацией «рецепторов смерти»
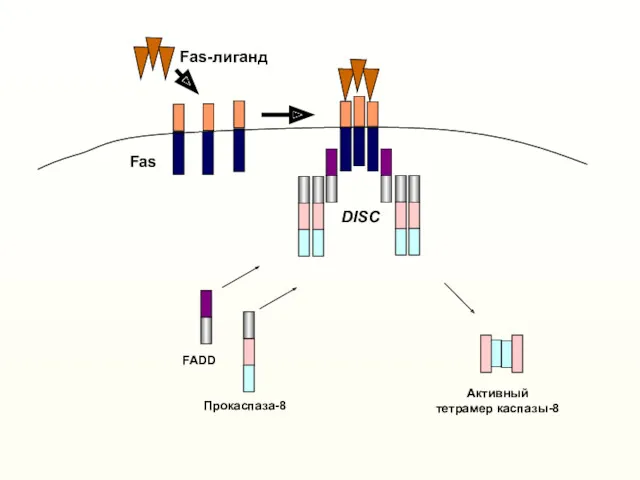
Прокаспаза-8
FADD
Fas
Прокаспаза-8
FADD
Fas
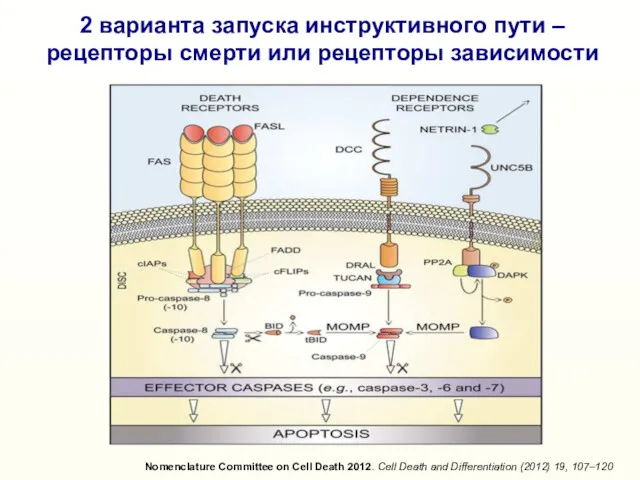
2 варианта запуска инструктивного пути –
рецепторы смерти или рецепторы зависимости
Nomenclature Committee
2 варианта запуска инструктивного пути –
рецепторы смерти или рецепторы зависимости
Nomenclature Committee
Эффекторные
каспазы 3,6,7
Киллерный лиганд
(Fas-L, TNFα, TRAIL и др.)
Рецепторы смерти
Эффекторные
каспазы 3,6,7
Киллерный лиганд
(Fas-L, TNFα, TRAIL и др.)
Рецепторы смерти
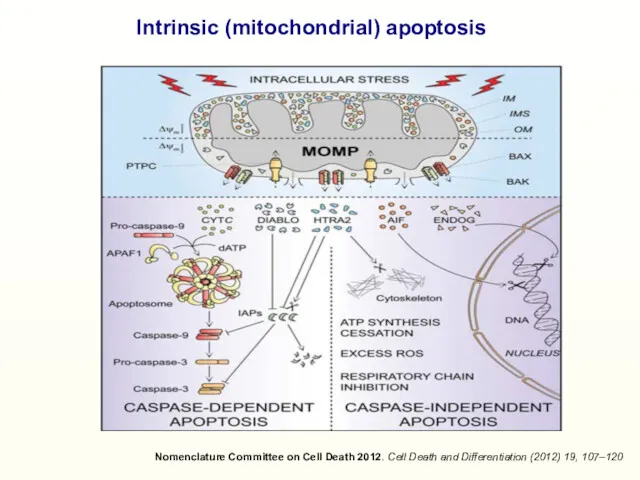
Nomenclature Committee on Cell Death 2012. Cell Death and Differentiation (2012)
Nomenclature Committee on Cell Death 2012. Cell Death and Differentiation (2012)

нечувствительность к апоптогенному действию Fas-L
(потеря экспрессии или мутации рецептора смерти
нечувствительность к апоптогенному действию Fas-L
(потеря экспрессии или мутации рецептора смерти
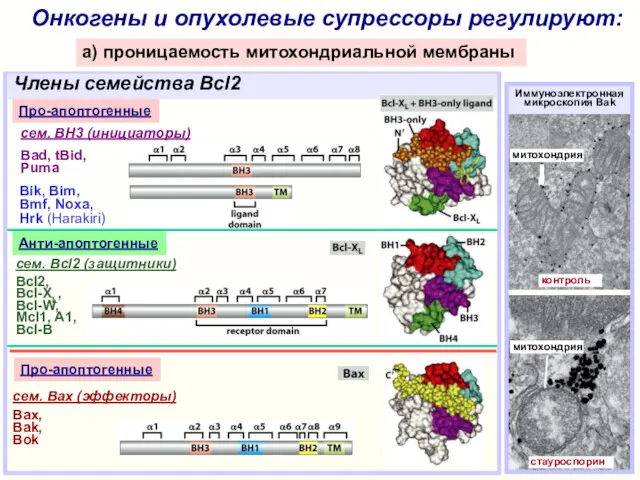
Онкогены и опухолевые супрессоры регулируют:
a) проницаемость митохондриальной мембраны
cем. Bcl2 (защитники)
Bcl2,
Bcl-XL,
Bcl-W,
Mcl1,
Онкогены и опухолевые супрессоры регулируют:
a) проницаемость митохондриальной мембраны
cем. Bcl2 (защитники)
Bcl2,
Bcl-XL,
Bcl-W,
Mcl1,
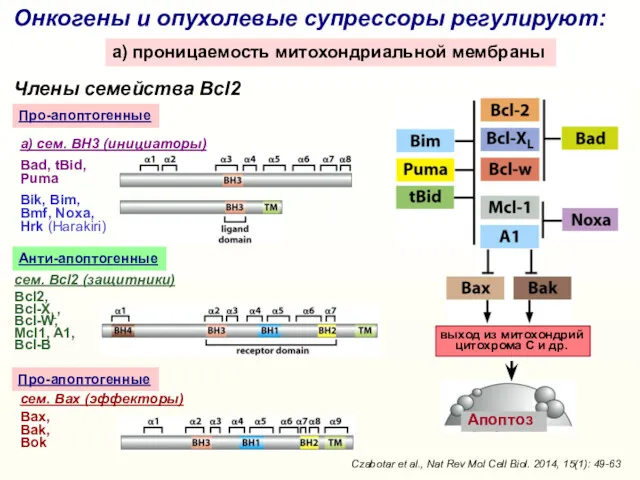
Онкогены и опухолевые супрессоры регулируют:
a) проницаемость митохондриальной мембраны
cем. Bcl2 (защитники)
Bcl2,
Bcl-XL,
Bcl-W,
Mcl1,
Онкогены и опухолевые супрессоры регулируют:
a) проницаемость митохондриальной мембраны
cем. Bcl2 (защитники)
Bcl2,
Bcl-XL,
Bcl-W,
Mcl1,
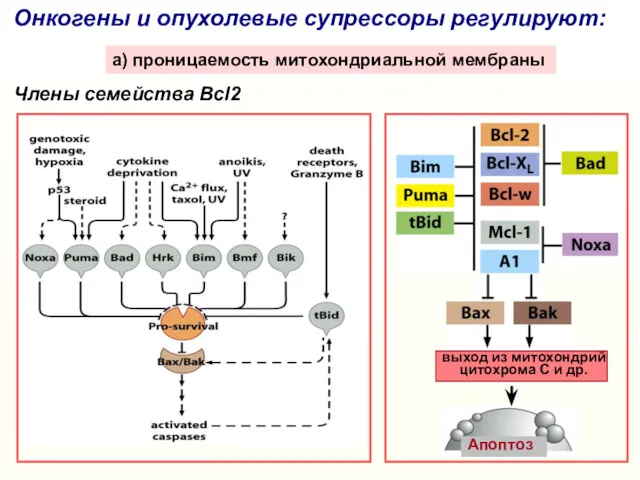
Онкогены и опухолевые супрессоры регулируют:
a) проницаемость митохондриальной мембраны
Члены семейства Bcl2
выход из
Онкогены и опухолевые супрессоры регулируют:
a) проницаемость митохондриальной мембраны
Члены семейства Bcl2
выход из
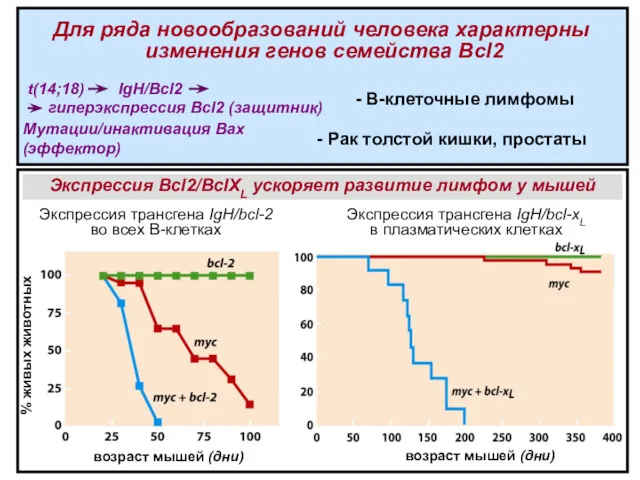
t(14;18)
IgH/
Bcl2
гиперэкспрессия Bcl2 (защитник)
- В-клеточные лимфомы
Мутации/инактивация Bax
(эффектор)
-
t(14;18)
IgH/
Bcl2
гиперэкспрессия Bcl2 (защитник)
- В-клеточные лимфомы
Мутации/инактивация Bax
(эффектор)
-
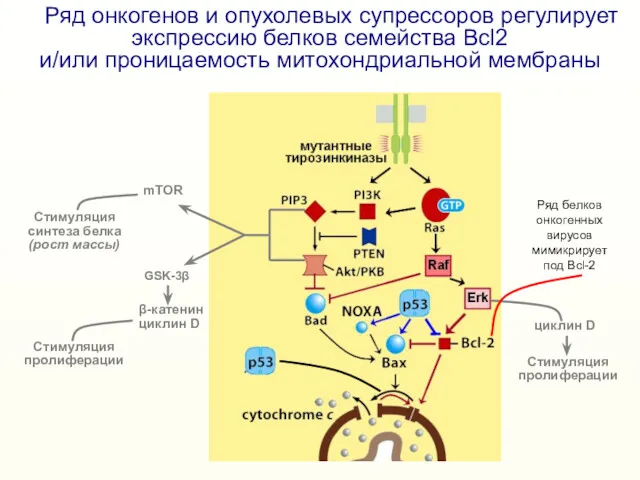
Ряд онкогенов и опухолевых супрессоров регулирует экспрессию белков семейства Bcl2
Ряд онкогенов и опухолевых супрессоров регулирует экспрессию белков семейства Bcl2

нечувствительность к апоптогенному действию Fas-L
(потеря экспрессии или мутации рецептора смерти
нечувствительность к апоптогенному действию Fas-L
(потеря экспрессии или мутации рецептора смерти
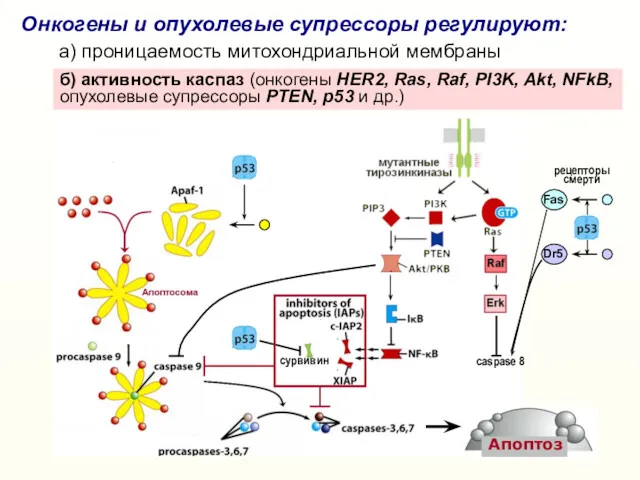
сурвивин
caspase 8
б) активность каспаз (онкогены HER2, Ras, Raf, PI3K, Akt, NFkB,
сурвивин
caspase 8
б) активность каспаз (онкогены HER2, Ras, Raf, PI3K, Akt, NFkB,

t(14;18)
IgH/
Bcl2
гиперэкспрессия Bcl2
- В-клеточные лимфомы
Мутации/инактивация Bax
- Рак толстой
t(14;18)
IgH/
Bcl2
гиперэкспрессия Bcl2
- В-клеточные лимфомы
Мутации/инактивация Bax
- Рак толстой
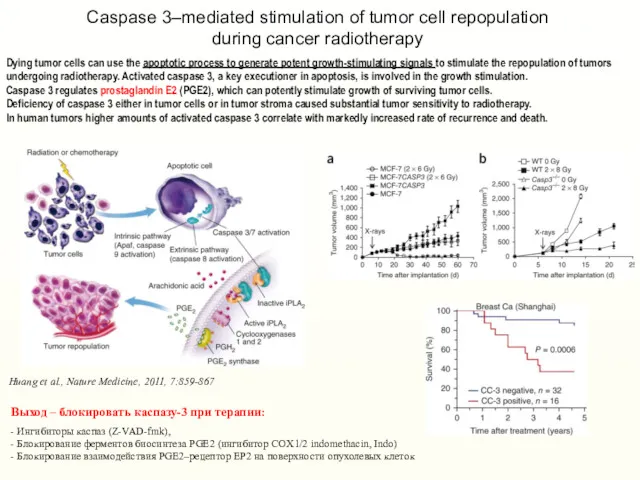
Dying tumor cells can use the apoptotic process to generate potent
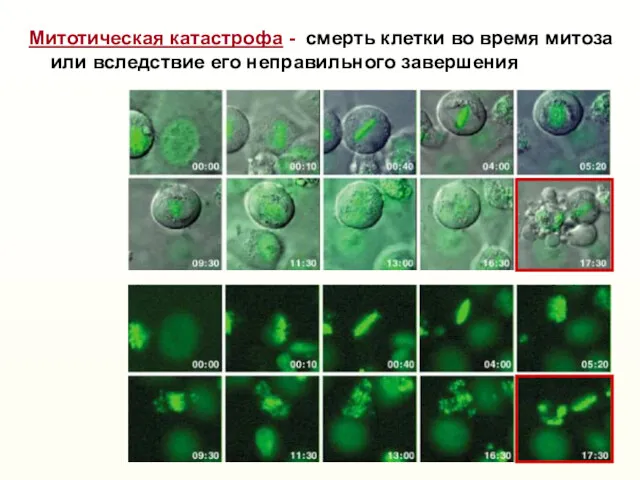
Митотическая катастрофа - смерть клетки во время митоза
или вследствие его
Митотическая катастрофа - смерть клетки во время митоза
или вследствие его
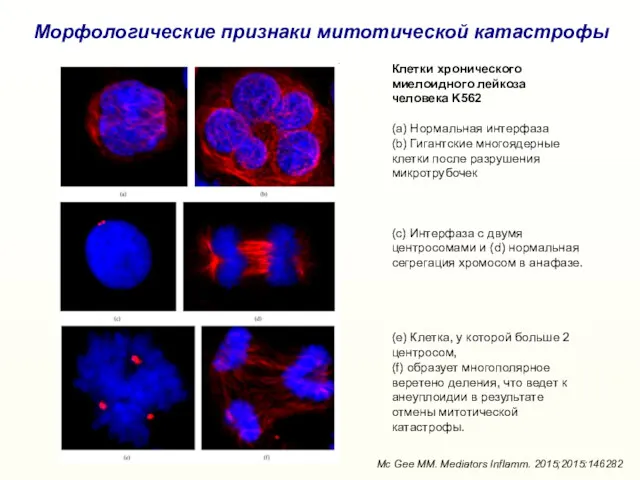
Морфологические признаки митотической катастрофы
Клетки хронического миелоидного лейкоза человека K562
(a) Нормальная
Морфологические признаки митотической катастрофы
Клетки хронического миелоидного лейкоза человека K562
(a) Нормальная
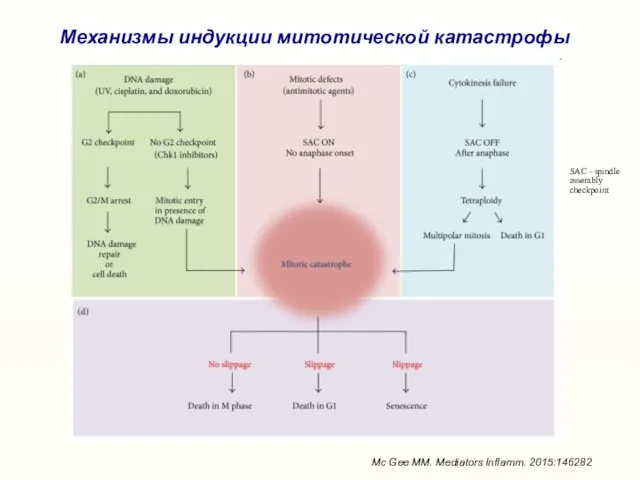
Механизмы индукции митотической катастрофы
Mc Gee MM. Mediators Inflamm. 2015:146282
SAC - spindle
Механизмы индукции митотической катастрофы
Mc Gee MM. Mediators Inflamm. 2015:146282
SAC - spindle
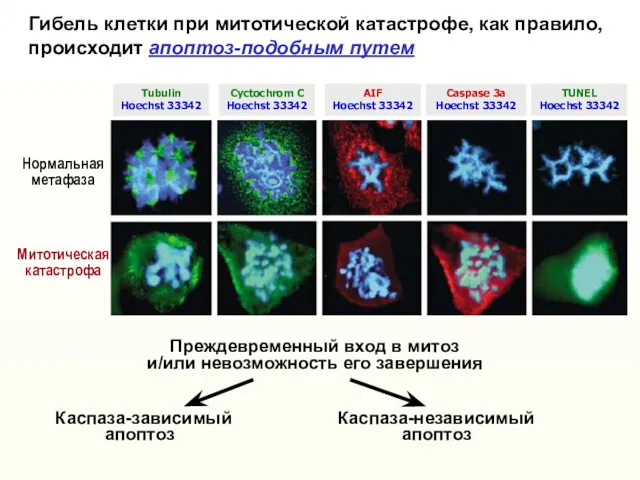
Гибель клетки при митотической катастрофе, как правило, происходит апоптоз-подобным путем
Преждевременный вход
Гибель клетки при митотической катастрофе, как правило, происходит апоптоз-подобным путем
Преждевременный вход

Отличия некроза от апоптоза
Отличия некроза от апоптоза
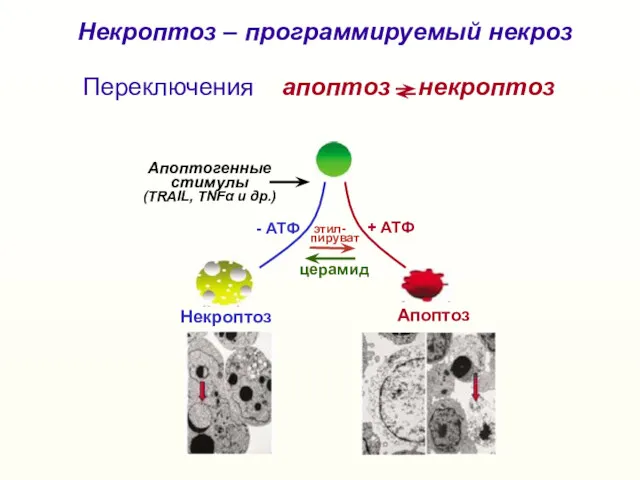
Некроптоз
Апоптоз
+ АТФ
- АТФ
Апоптогенные
стимулы
(TRAIL, TNFα и др.)
этил-
пируват
Переключения апоптоз некроптоз
церамид
Некроптоз – программируемый
Некроптоз
Апоптоз
+ АТФ
- АТФ
Апоптогенные
стимулы
(TRAIL, TNFα и др.)
этил-
пируват
Переключения апоптоз некроптоз
церамид
Некроптоз – программируемый
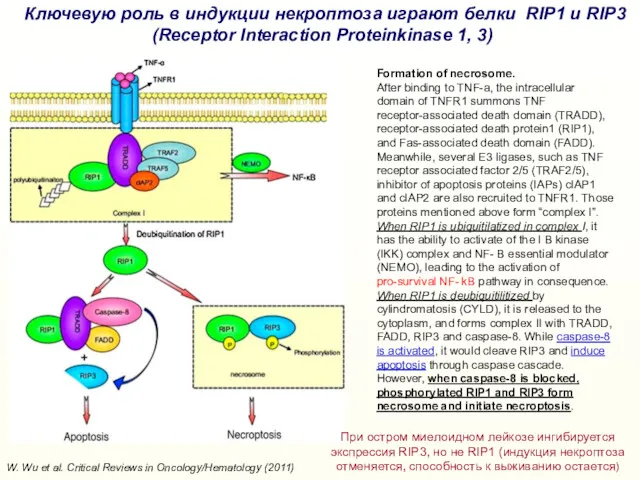
W. Wu et al. Critical Reviews in Oncology/Hematology (2011)
Ключевую роль
W. Wu et al. Critical Reviews in Oncology/Hematology (2011)
Ключевую роль
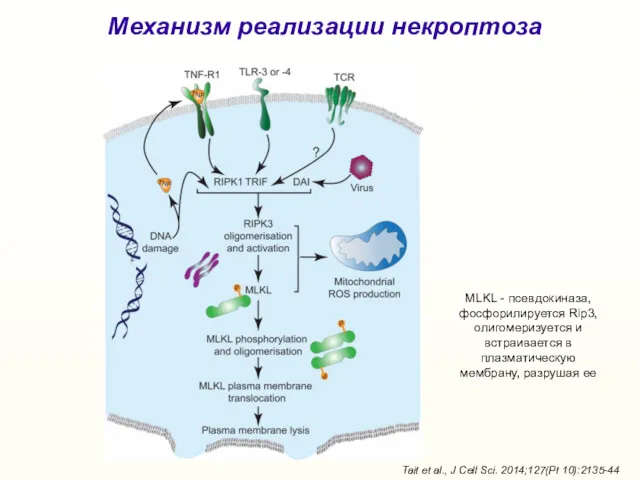
Tait et al., J Cell Sci. 2014;127(Pt 10):2135-44
Механизм реализации некроптоза
MLKL -
Tait et al., J Cell Sci. 2014;127(Pt 10):2135-44
Механизм реализации некроптоза
MLKL -
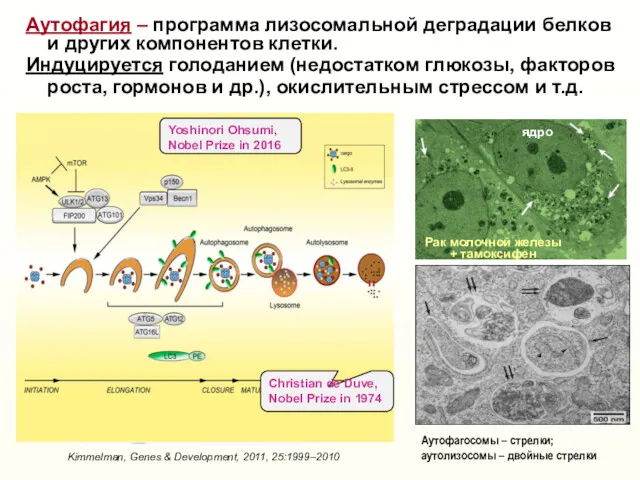
Аутофагия – программа лизосомальной деградации белков
и других компонентов клетки.
Индуцируется
Аутофагия – программа лизосомальной деградации белков
и других компонентов клетки.
Индуцируется
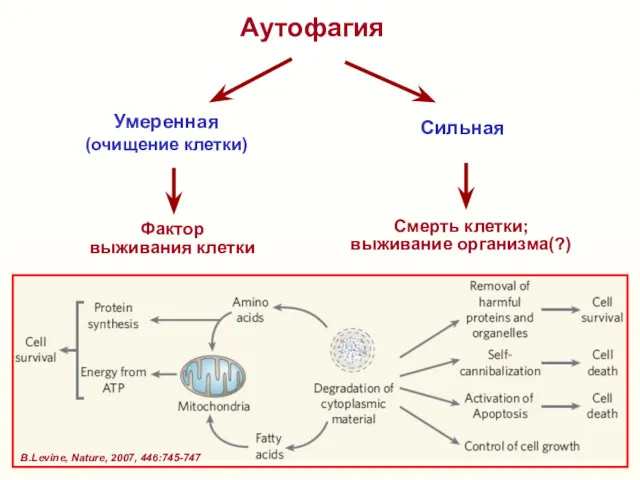
Аутофагия
Умеренная
(очищение клетки)
Сильная
Фактор
выживания клетки
Смерть клетки;
выживание организма(?)
В.Levine, Nature, 2007, 446:745-747
Аутофагия
Умеренная
(очищение клетки)
Сильная
Фактор
выживания клетки
Смерть клетки;
выживание организма(?)
В.Levine, Nature, 2007, 446:745-747
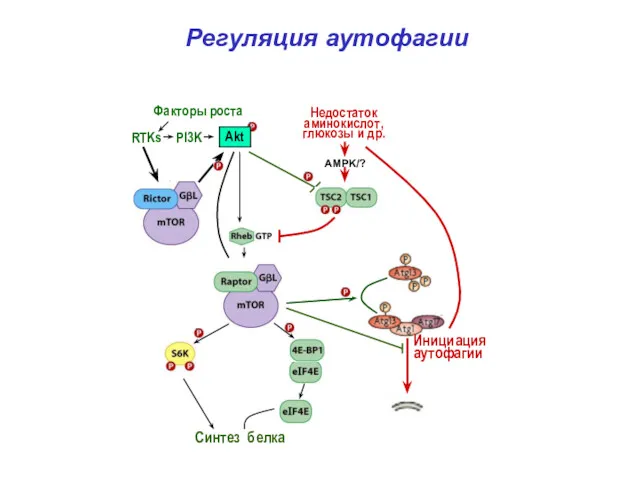
Регуляция аутофагии
Синтез белка
Инициация
аутофагии
Регуляция аутофагии
Синтез белка
Инициация
аутофагии
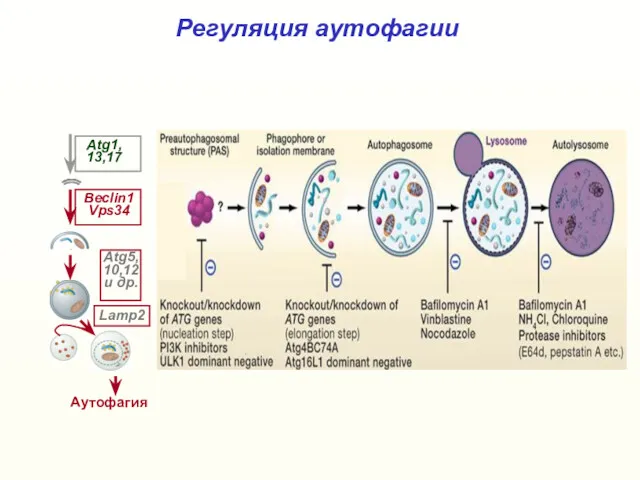
Регуляция аутофагии
Аутофагия
Beclin1
Vps34
Atg1,
13,17
Atg5,
10,12
и др.
Lamp2
Регуляция аутофагии
Аутофагия
Beclin1
Vps34
Atg1,
13,17
Atg5,
10,12
и др.
Lamp2
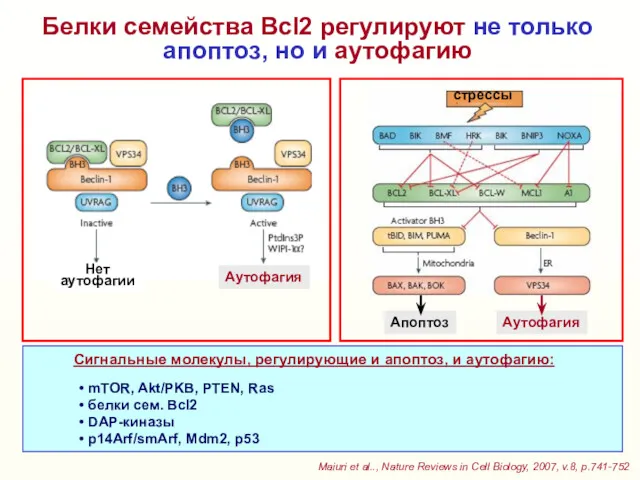
Белки семейства Bcl2 регулируют не только апоптоз, но и аутофагию
Maiuri et
Белки семейства Bcl2 регулируют не только апоптоз, но и аутофагию
Maiuri et
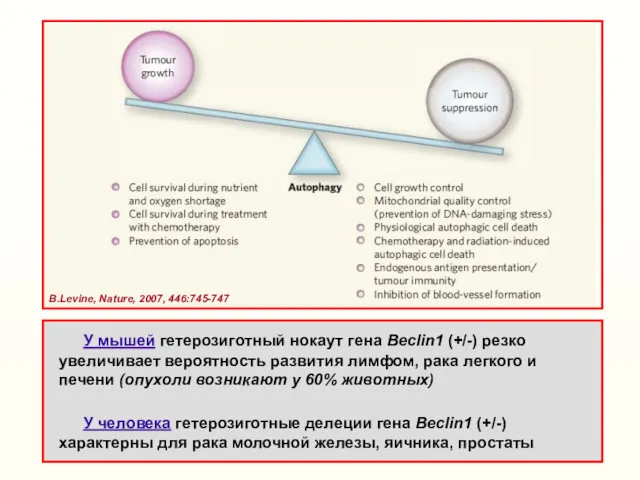
В.Levine, Nature, 2007, 446:745-747
У мышей гетерозиготный нокаут гена Beclin1 (+/-)
В.Levine, Nature, 2007, 446:745-747
У мышей гетерозиготный нокаут гена Beclin1 (+/-)
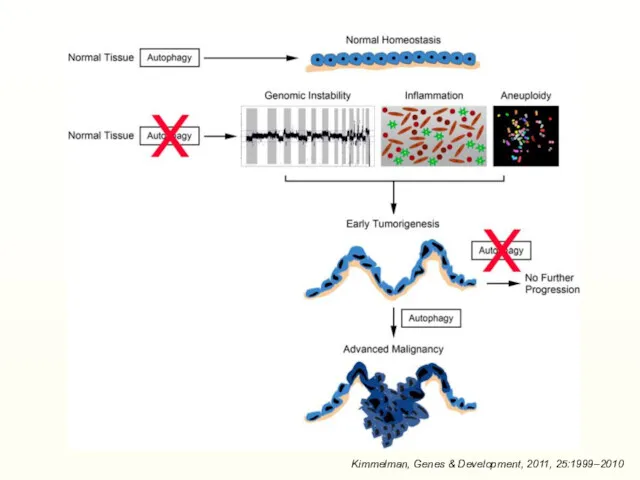
Kimmelman, Genes & Development, 2011, 25:1999–2010
Kimmelman, Genes & Development, 2011, 25:1999–2010
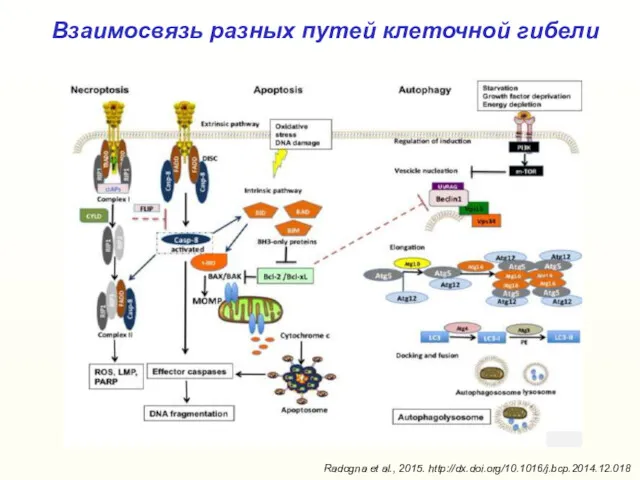
Взаимосвязь разных путей клеточной гибели
Radogna et al., 2015. http://dx.doi.org/10.1016/j.bcp.2014.12.018
Взаимосвязь разных путей клеточной гибели
Radogna et al., 2015. http://dx.doi.org/10.1016/j.bcp.2014.12.018
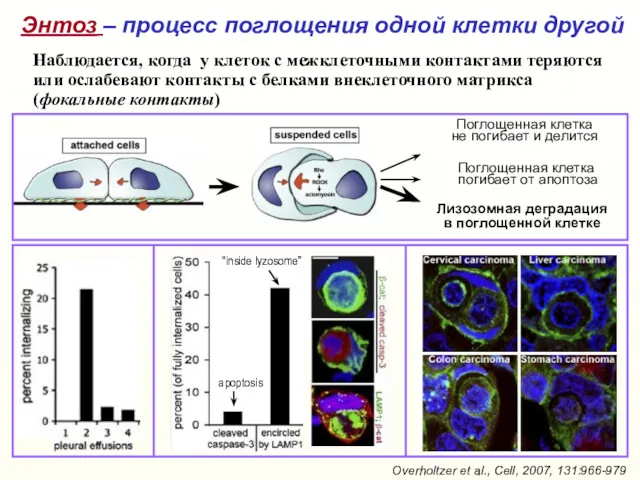
Лизозомная деградация
в поглощенной клетке
Поглощенная клетка
не погибает и делится
Overholtzer et al., Cell,
Лизозомная деградация
в поглощенной клетке
Поглощенная клетка
не погибает и делится
Overholtzer et al., Cell,
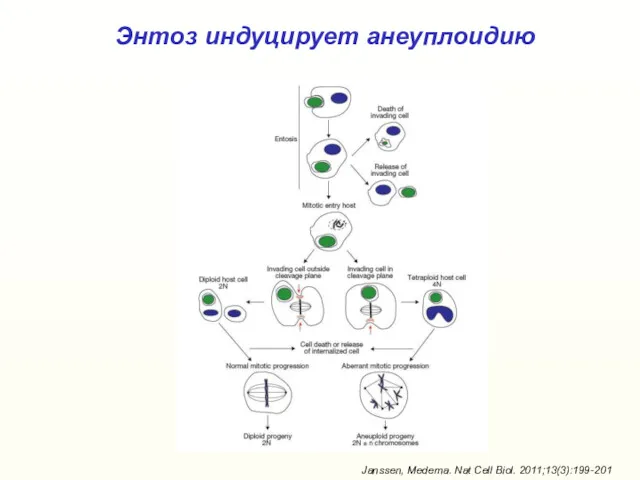
Энтоз индуцирует анеуплоидию
Janssen, Medema. Nat Cell Biol. 2011;13(3):199-201
Энтоз индуцирует анеуплоидию
Janssen, Medema. Nat Cell Biol. 2011;13(3):199-201

Роль энтоза в канцерогенезе
Sun et al., Cell Res. 2014;24(11):1299-310 and 1288-98;
Роль энтоза в канцерогенезе
Sun et al., Cell Res. 2014;24(11):1299-310 and 1288-98;
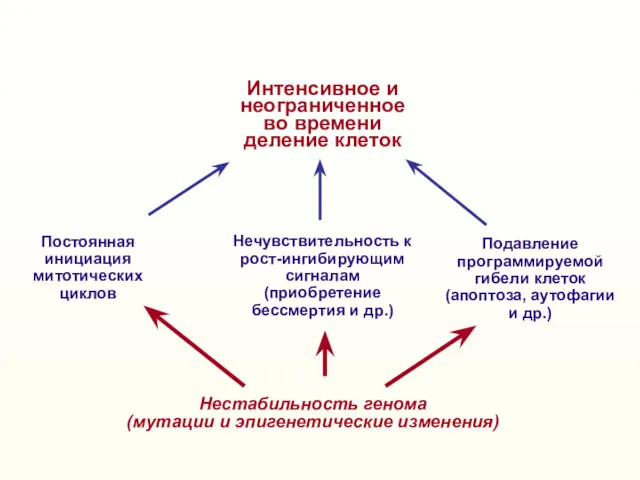
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
Нестабильность генома
(мутации и эпигенетические изменения)
Нечувствительность к
рост-ингибирующим сигналам
(приобретение бессмертия и др.)
Постоянная инициация
 Лучевая терапия: цели, методы, виды, осложнения
Лучевая терапия: цели, методы, виды, осложнения Фортранс. Рекомендации по приёму препарата фортранс
Фортранс. Рекомендации по приёму препарата фортранс Головная боль у детей. Классификация головной боли
Головная боль у детей. Классификация головной боли Балалардағы көмірсу алмасуының ерекшеліктері. Олардың бұзылыстарының семиотикасы
Балалардағы көмірсу алмасуының ерекшеліктері. Олардың бұзылыстарының семиотикасы Ультразвуковая диагностика заболеваний вен нижних конечностей
Ультразвуковая диагностика заболеваний вен нижних конечностей Сахарный диабет. Лечение
Сахарный диабет. Лечение Черепно-мозговая грыжа у детей
Черепно-мозговая грыжа у детей Сердечно-легочная реанимация
Сердечно-легочная реанимация Ақ қабықтың аурулары
Ақ қабықтың аурулары Натуральные соки
Натуральные соки Болезнь Виллебранда
Болезнь Виллебранда Медицинская демография. Динамика населения. Рождаемость. (Лекция 7)
Медицинская демография. Динамика населения. Рождаемость. (Лекция 7) Жедел аппендицит
Жедел аппендицит Почему важно грудное вскармливание
Почему важно грудное вскармливание Денсаулық сақтауға арналған ҚР бюджет жүйесі
Денсаулық сақтауға арналған ҚР бюджет жүйесі Prophylactic measure – the use of the tooth-brush
Prophylactic measure – the use of the tooth-brush Гемолитические анемии
Гемолитические анемии Санитарно-гигиеническое обследование промышленных предприятий
Санитарно-гигиеническое обследование промышленных предприятий Инвалидность и реабилитация как медико-социальная проблема
Инвалидность и реабилитация как медико-социальная проблема Hereditary Metabolic Disorder
Hereditary Metabolic Disorder Метод лактаційної аменореї
Метод лактаційної аменореї Ожирение - заболевание или эстетическая проблема
Ожирение - заболевание или эстетическая проблема Иммуноцитокины. Классификация
Иммуноцитокины. Классификация Фонетико-фонематическое недоразвитие
Фонетико-фонематическое недоразвитие Лекарственные средства, производные конденсированных гетероциклических систем. (Тема 5)
Лекарственные средства, производные конденсированных гетероциклических систем. (Тема 5) Введение в дерматологию. Кожа
Введение в дерматологию. Кожа Постреанимационная болезнь. Этиология. Патогенез
Постреанимационная болезнь. Этиология. Патогенез Көкірекаралық аурулар
Көкірекаралық аурулар