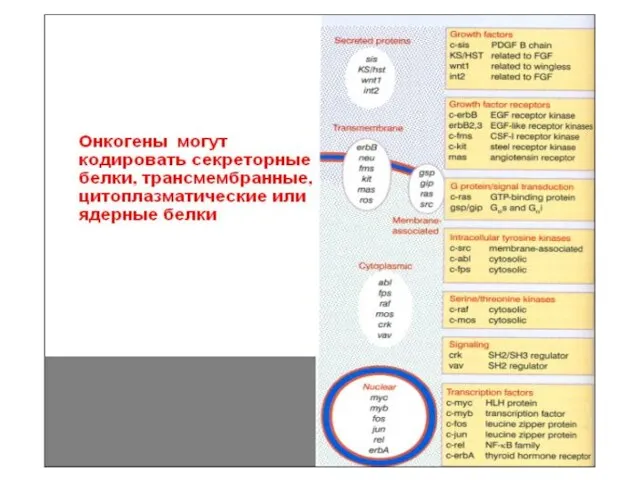
- Протоонкогены и передача клеточного сигнала

Содержание
- 2. 5.1. Онковирусы и онкогены
- 3. Различия между нормальными и трансформированными клетками
- 6. Общая модель возникновения рака
- 7. Гипотеза раковых стволовых клеток (РСК)
- 8. Две модели, объяснящие гетерогенность опухоли
- 9. Маркеры для обогащения РСК
- 10. Стратегия в лечении рака
- 11. Стратегия в лечении рака
- 12. Онковирусы
- 13. Два способа инфицирования вирусом
- 14. Трансформирующие вирусы, несущие онкогены
- 15. Трансформация вирусом
- 16. История вирусного канцерогенеза (1)
- 17. История вирусного канцерогенеза (2)
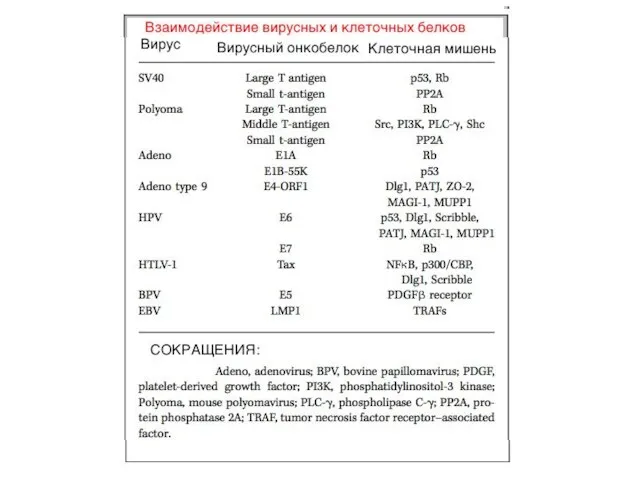
- 18. ALV/ASV, avian leukosis-sarcoma virus group; BLV, bovine leukemia virus; BKV, BK virus; BPV, bovine papillomavirus; FeLV/FeSV,
- 21. Вирус папилломы человека (HPVs) Причина рака шейки матки (90 % случаев)
- 22. Классификация HPVs Более 120 типов Серозные Кожные Около 30% онкогенных Высокий риск: HPV type 16, 18,
- 23. Организация эписомального и интегрированного HPV
- 24. Геном вируса HPV-16
- 25. Схематическая структура мини-генома HPV-16
- 26. Структура белка E7
- 27. Структура белка E7 (C-конец)
- 28. Функции вирусных белков
- 29. Жизненный цикл HPV
- 30. Структура регуляторного райтона (long control region: LCR)
- 31. Транскрипты HPV16
- 32. Механизм действия HPV белков Е6 и Е7
- 33. Роль NOTCH1 белка в канцерогенезе
- 34. Критические этапы HPVs -индуцированного канцерогенеза
- 36. Многостадийный механизм HPV-индуцированного канцерогенеза
- 37. Функциональные домены Т- антигена вируса SV 40
- 38. ДНК-вирусы, вызывающие рак у человека (1)
- 39. ДНК-вирусы, вызывающие рак у человека (2)
- 40. Разнообразные механизмы действия ДНК-онковирусов
- 41. Мишени для ДНК-онковирусов
- 42. Свойства вирусов человека
- 43. Новые онковирусы Новое исследование указывает на то, что белок синтезируемый вирусом, вызывающим рак, играет важнейшую роль
- 44. Основные принципы вирусного онкогенеза Вирусы могут вызывать рак у животных и человека Онковирусы часто вызывают продолжительную
- 45. Ретровирусы
- 46. Репликационный цикл ретровирусов
- 47. Генетическая организация некоторых ретровирусов А. «Недефективные» репликационно-компетентные вирусы . Показаны вирусы с простым или сложным геномом.
- 48. Генетическая организация некоторых ретровирусов В. Вирусы, несущие онкогены. Все дефективные, исключая RSV (вирус саркомы Рауша). MC-29
- 49. Ретровирусы, не несущие клеточные гены, но их активирующие
- 50. Ретровирусы, содержащие клеточные онкогены
- 51. Трансформирующий вирус несет копию гена хозяина
- 52. Трансформирующий ретровирус
- 53. Ретровирусная трансформация
- 54. Механизмы активации прото-онкогенов (ПО) Траснформирующая «сила» ПО может характеризоваться появлением фокусов. Тестируемая культура – 3Т3- мышиные
- 55. Вставки, транслокация или амплификация могут активировать ПО В случае вставки или транслокации существуют доказательства, что эти
- 56. Варианты активации c-myc
- 57. Другие ПО, активируемые вставкой ретровируса c-erbB, c-myb, c-mos, c-H-ras, c-raf
- 58. Результаты транслокации Часть белка ПО может быть активирована независимо от другой части (c-myc). Другой партнер в
- 59. Хромосомы пациента с острой промиелоцитной лейкемией (APL)
- 60. Филадельфийская хромосома (PH) Хроническая миелогенная лейкемия (CML) – 5000kb конца 9-й хр. (c-abl) b 5,8kb 22-й
- 61. Образование Филадельфийской хромосомы (CML)
- 62. Варианты Ph хромосомы
- 63. Лечение CML Стандартная терапия хронической миелоидной лейкемии – тирозин киназный ингибитор иматиниб (imatinib). Результат лечения -
- 64. Лимфома Беркита
- 65. Варианты транслокаций TCR - T-cell receptor (14q11)
- 66. Кариотип лимфомы Беркита
- 67. Лимфома Беркита
- 68. Классификация лимфом
- 69. Варианты транслокаций
- 70. 5.2. Передача клеточного сигнала (Сигнальная трансдукция)
- 71. 5.2.1. Факторы роста и их рецепторы
- 72. Прохождение сигнала
- 74. Способы прохождения сигнала
- 75. Роль заряда в прохождении сигнала
- 76. К-каналы
- 77. Структура К-канала
- 80. Факторы роста (GF) К настоящему времени установлено, что семейство факторов роста представлено 13 белками: EGF, transforming
- 81. Процессинг GF В некоторых случаях, как это показано для фактора NRG-1beta1, его расщепление происходит в аппарате
- 82. Механизм активации пролиганда протеазами и взаимодействие растворимого лиганда с рецептором.
- 83. Начало СТ
- 85. Характеристика RTKs У человека идентифицировано 58 RTKs, которые подразделяются на 20 подсемейств Все рецепторы имеют сходную
- 87. Семейство рецепторов RTKs
- 88. Внеклеточная часть некоторых рецепторов В основном, RTKs складываются в димеры, когда лиганд (красный) связывается с внеклеточным
- 89. Внутриклеточный домен RTKs E) Инсулин рецептор-подобные Insulin receptor-like (активация-петля-ингибирование, activation loop inhibition). В FGFR, инсулиновом рецепторе
- 91. Активация рецепторов сопровождается связыванием с нижестоящими молекулами
- 92. Активация PDGFR
- 93. Основные характеристики некоторых ФР и их рецепторов
- 94. Семейство Epidermal growth factor receptor (EGFR) EGF был изолирован в1962 Stanley Cohen. Его взаимодействие с EGFR
- 95. Общая структура EGFR
- 96. Структура EGFR белка (A), активация (B) и димеризация лигандом (C)
- 97. Лиганды для EGFR Идентифицировано 11 лигандов, которые разделены на 3 группы: 1.EGF, transforming growth factor-a, ампирегулин
- 98. Субстраты для EGFR Phospholipase Cg, CBL, GRB2, SHC и p85. Включая mitogen-activated protein kinase, phosphatidylinositol 3-kinase
- 99. Активируемые пути СТ
- 100. Активируемые пути СТ
- 101. Активируемые пути СТ
- 102. Мутации в раковых клетках мозга (глиобластома) и легких
- 103. Таргетная терапия EGFR-СТ
- 104. EGFR и рак
- 105. Down-регуляция EGFR
- 108. Cbl-белки в СТ
- 109. Cbl-направленная интернализация, мультиубиквитиляция и деградация активированных RTKs а). Присоединение GF индуцирует фос-ие RTK и мобилизацию Cbl
- 110. TKB домен имеет свободный доступ к рецептору (pTyr1045 EGF рецептор) и E3 лигазная функция Cbl может
- 111. Активация EGFR
- 113. Скачать презентацию

5.1. Онковирусы и онкогены
5.1. Онковирусы и онкогены
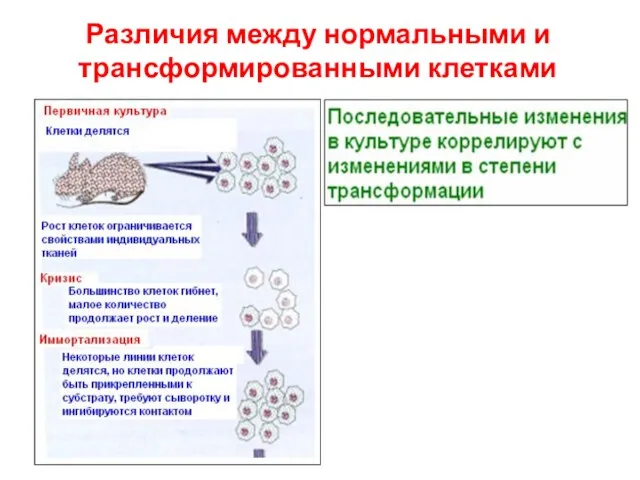
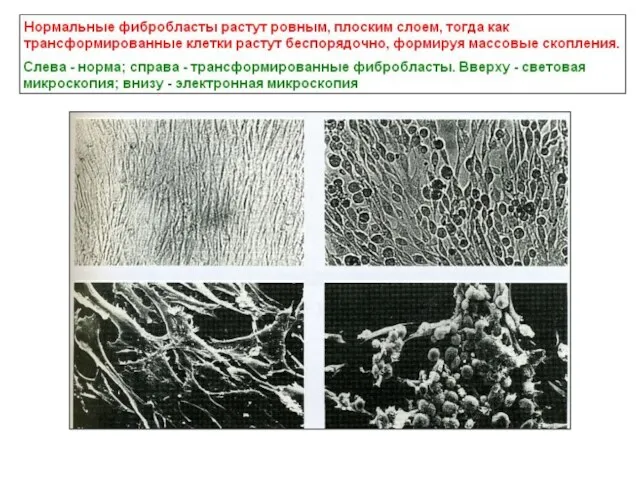
Различия между нормальными и трансформированными клетками
Различия между нормальными и трансформированными клетками
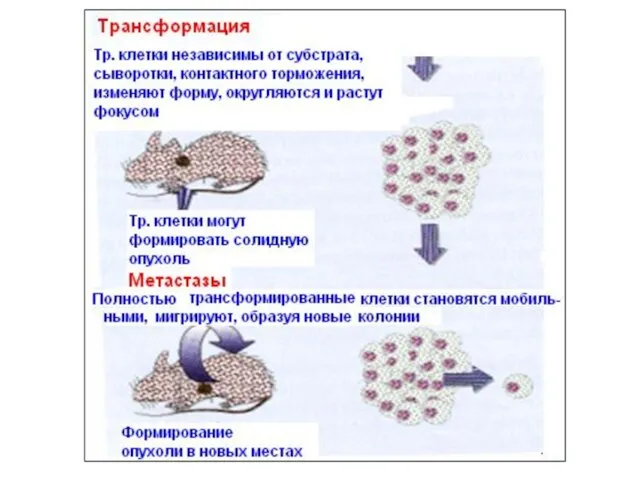
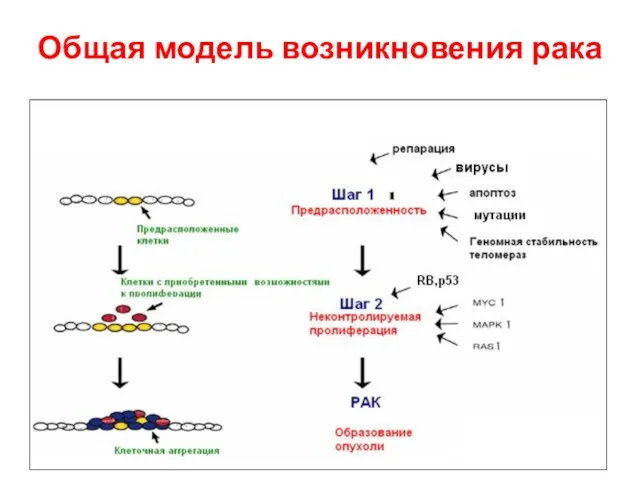
Общая модель возникновения рака
Общая модель возникновения рака

Гипотеза раковых стволовых клеток (РСК)
Гипотеза раковых стволовых клеток (РСК)
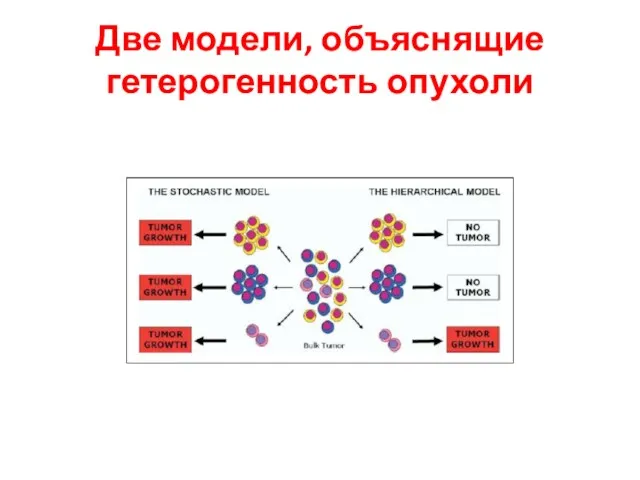
Две модели, объяснящие гетерогенность опухоли
Две модели, объяснящие гетерогенность опухоли

Маркеры для обогащения РСК
Маркеры для обогащения РСК
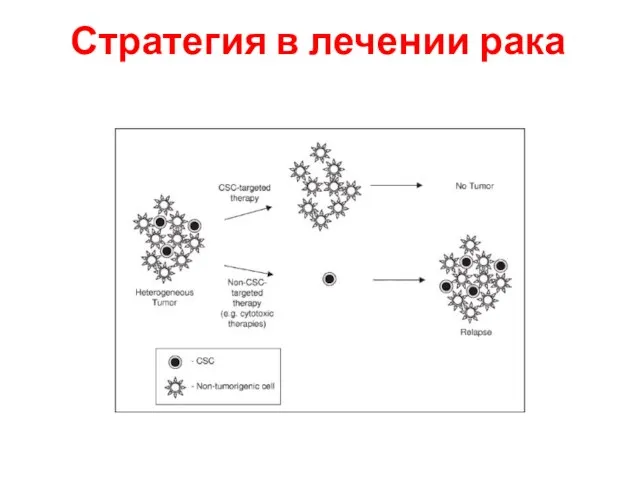
Стратегия в лечении рака
Стратегия в лечении рака
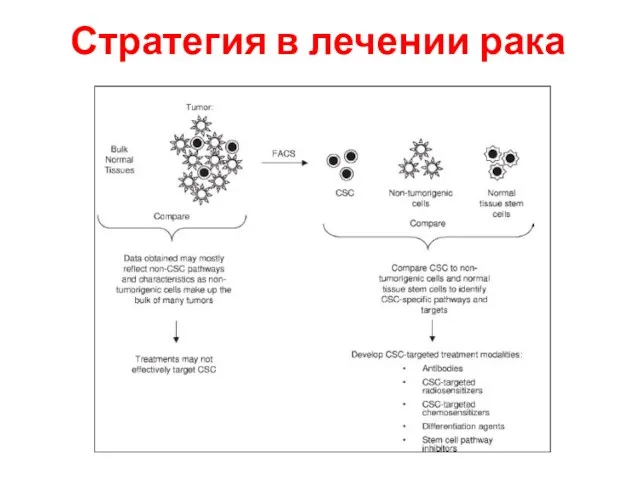
Стратегия в лечении рака
Стратегия в лечении рака

Онковирусы
Онковирусы
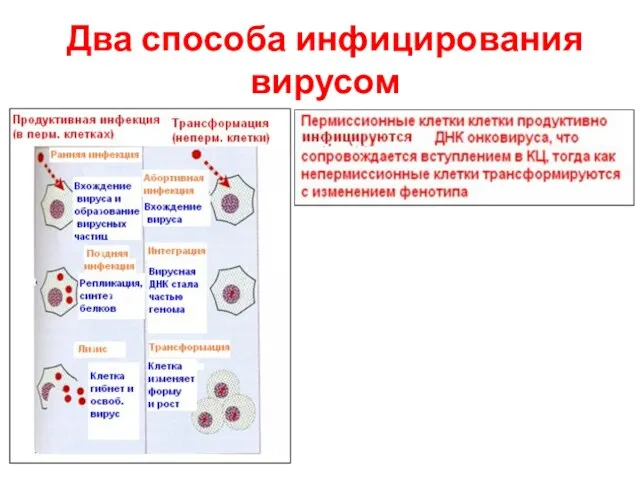
Два способа инфицирования вирусом
Два способа инфицирования вирусом
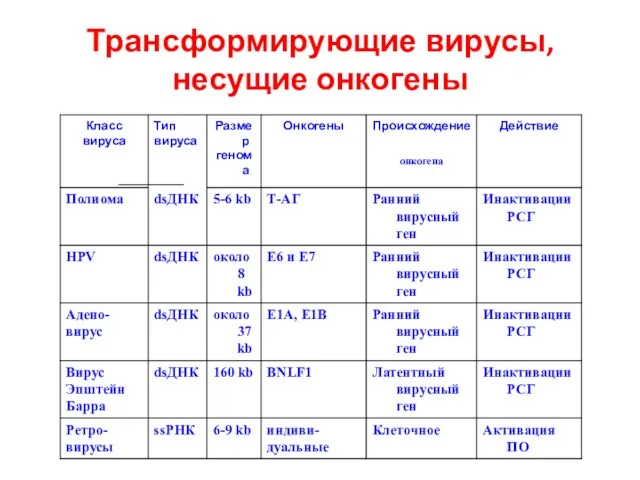
Трансформирующие вирусы, несущие онкогены
Трансформирующие вирусы, несущие онкогены
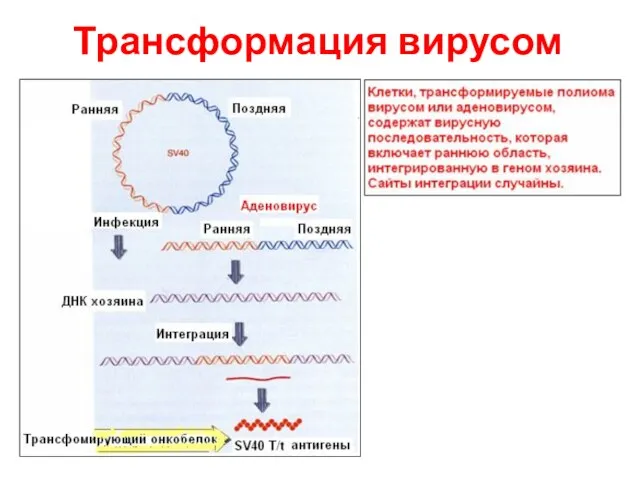
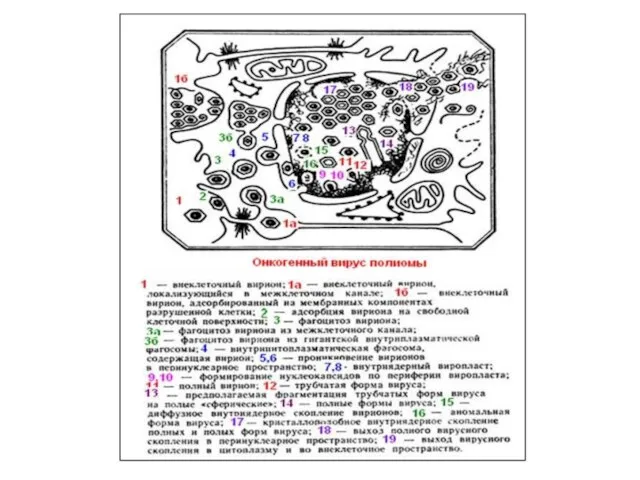
Трансформация вирусом
Трансформация вирусом

История вирусного канцерогенеза (1)
История вирусного канцерогенеза (1)
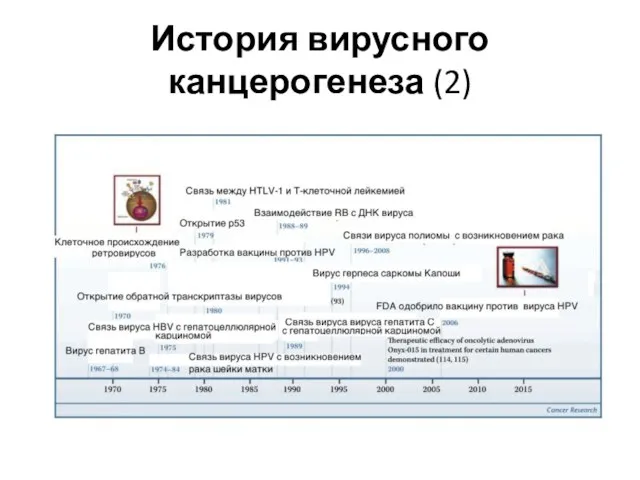
История вирусного канцерогенеза (2)
История вирусного канцерогенеза (2)
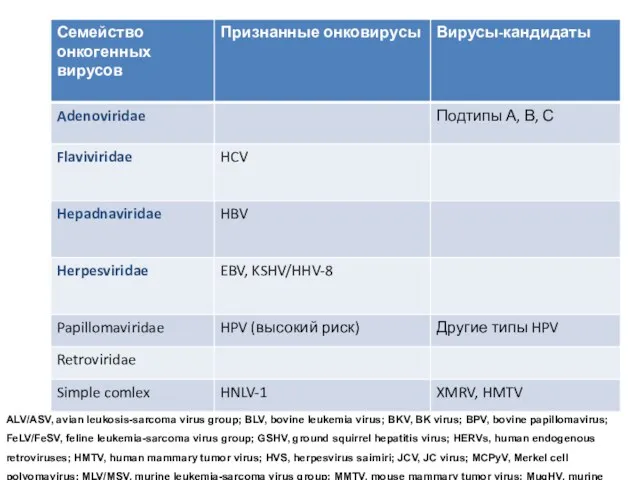
ALV/ASV, avian leukosis-sarcoma virus group; BLV, bovine leukemia virus; BKV, BK
ALV/ASV, avian leukosis-sarcoma virus group; BLV, bovine leukemia virus; BKV, BK

Вирус папилломы человека (HPVs)
Причина рака шейки матки (90 % случаев)
Вирус папилломы человека (HPVs)
Причина рака шейки матки (90 % случаев)
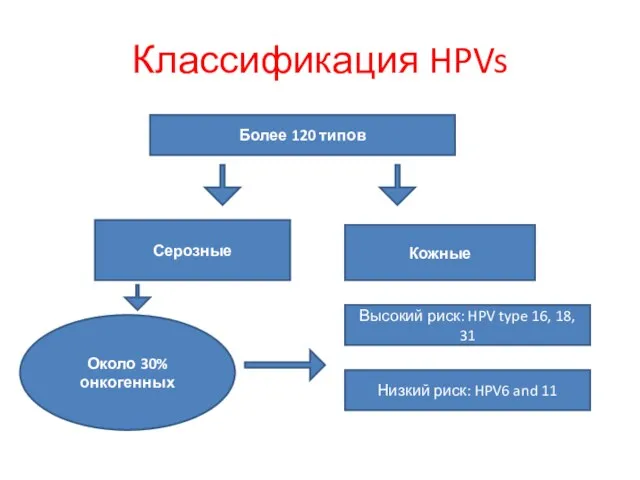
Классификация HPVs
Более 120 типов
Серозные
Кожные
Около 30% онкогенных
Высокий риск: HPV type 16, 18,
Классификация HPVs
Более 120 типов
Серозные
Кожные
Около 30% онкогенных
Высокий риск: HPV type 16, 18,
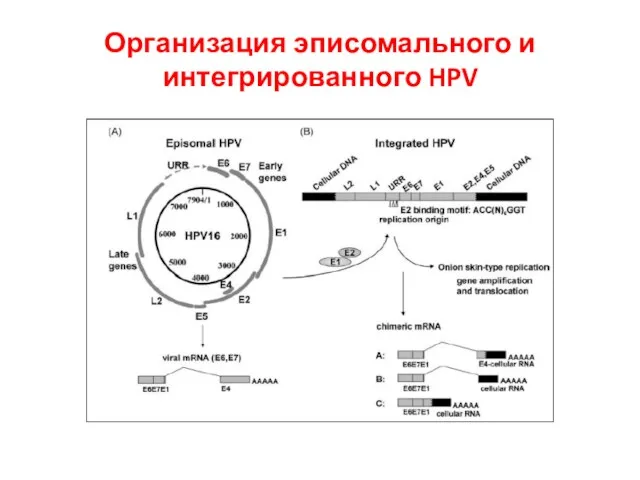
Организация эписомального и интегрированного HPV
Организация эписомального и интегрированного HPV
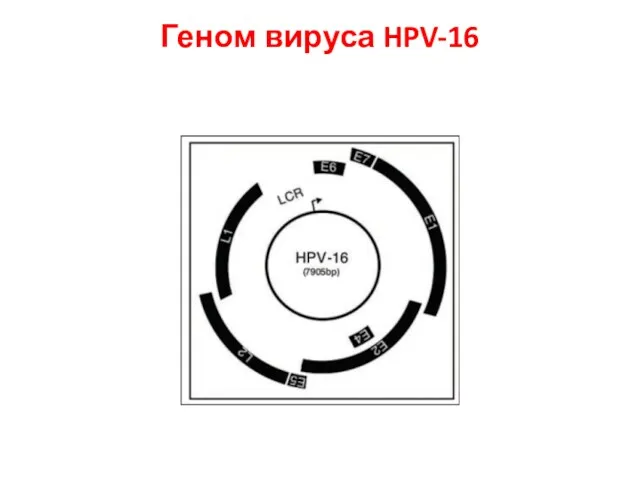
Геном вируса HPV-16
Геном вируса HPV-16
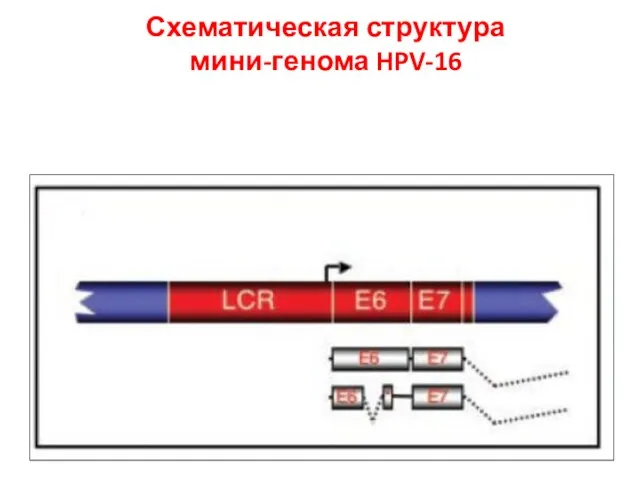
Схематическая структура
мини-генома HPV-16
Схематическая структура
мини-генома HPV-16
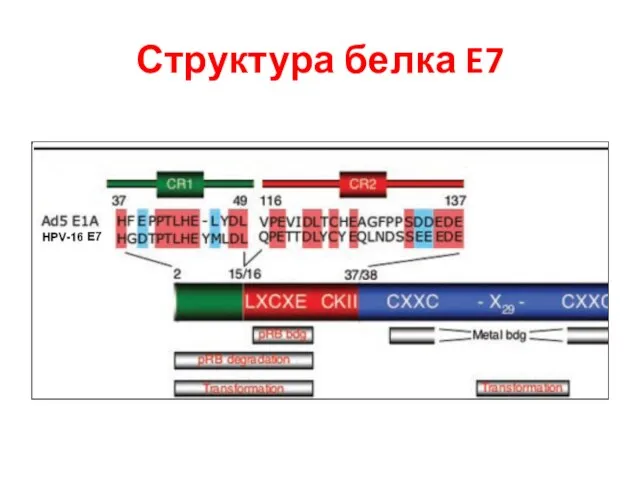
Структура белка E7
Структура белка E7
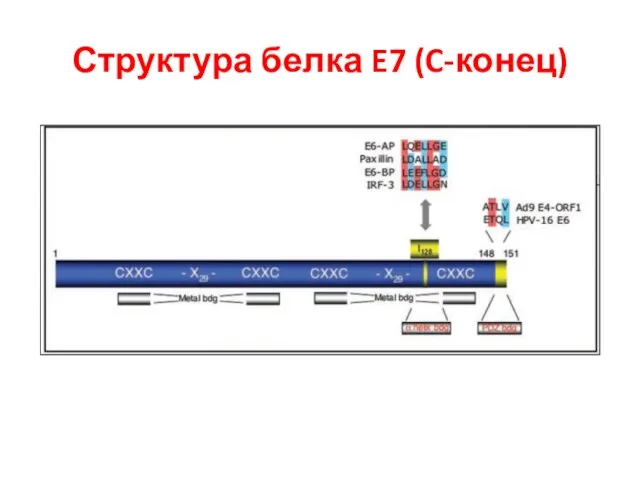
Структура белка E7 (C-конец)
Структура белка E7 (C-конец)
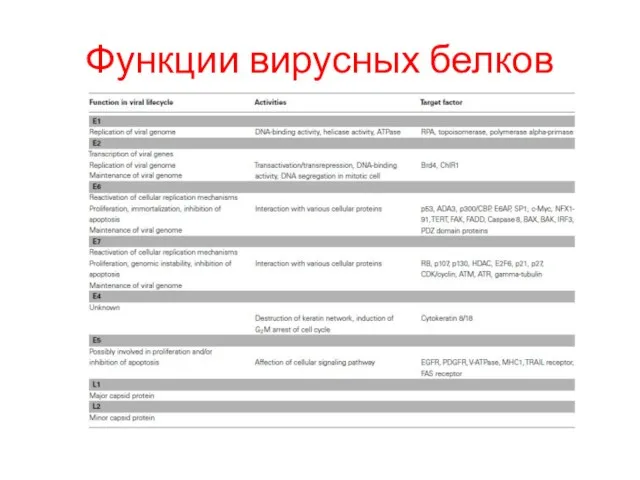
Функции вирусных белков
Функции вирусных белков
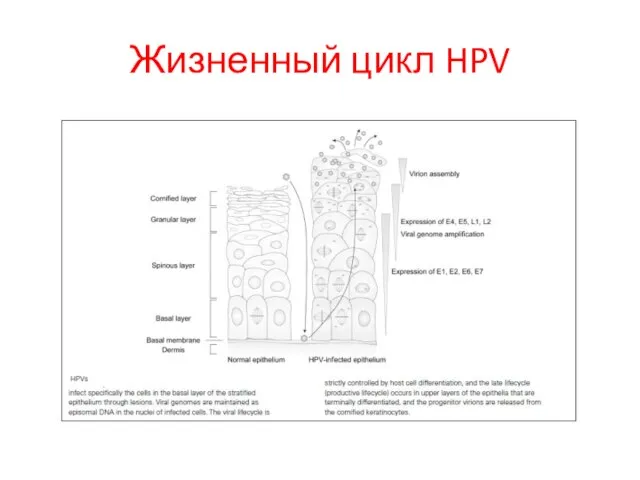
Жизненный цикл HPV
Жизненный цикл HPV
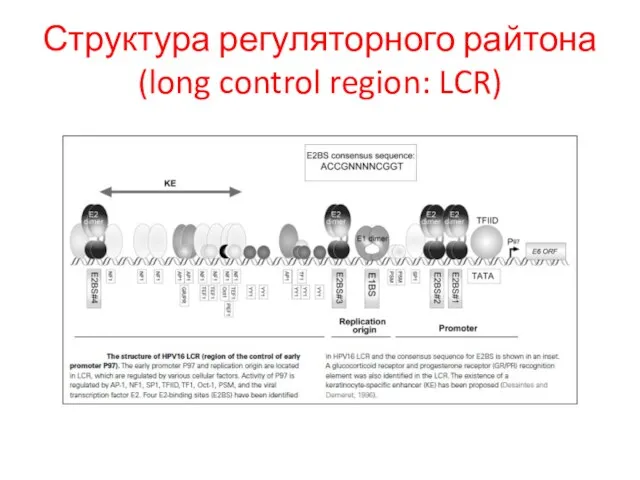
Структура регуляторного райтона (long control region: LCR)
Структура регуляторного райтона (long control region: LCR)
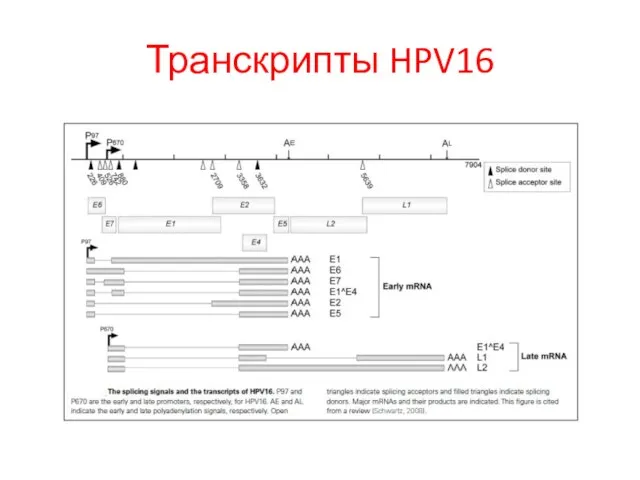
Транскрипты HPV16
Транскрипты HPV16
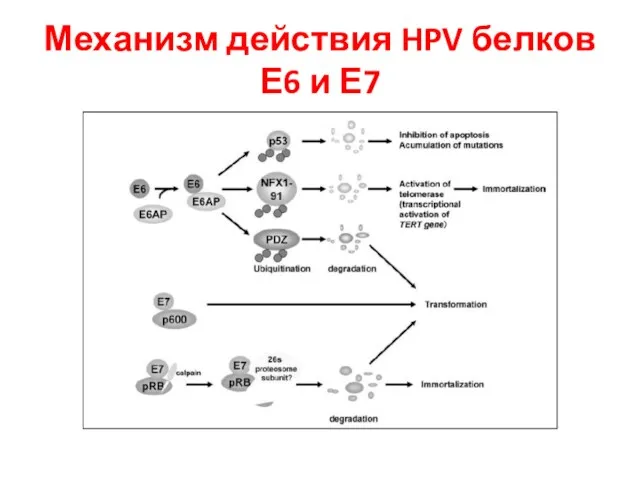
Механизм действия HPV белков Е6 и Е7
Механизм действия HPV белков Е6 и Е7

Роль NOTCH1 белка в канцерогенезе
Роль NOTCH1 белка в канцерогенезе
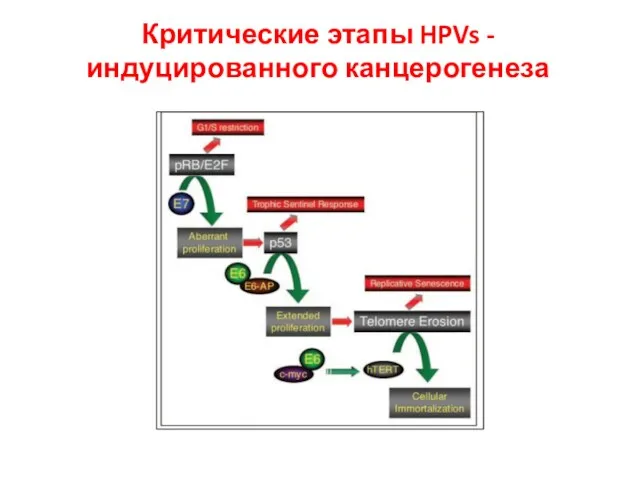
Критические этапы HPVs -индуцированного канцерогенеза
Критические этапы HPVs -индуцированного канцерогенеза
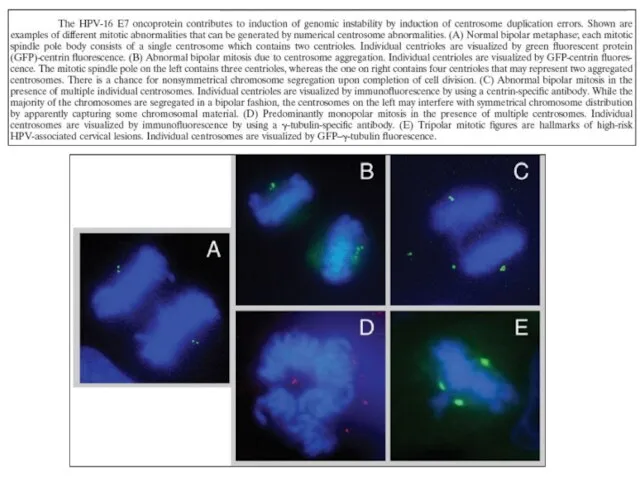
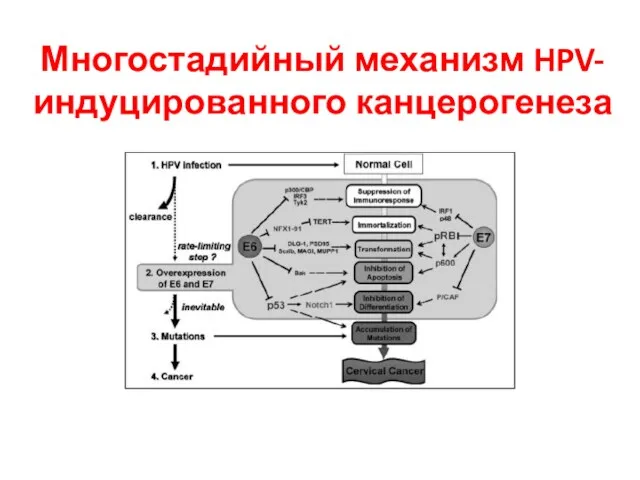
Многостадийный механизм HPV-индуцированного канцерогенеза
Многостадийный механизм HPV-индуцированного канцерогенеза
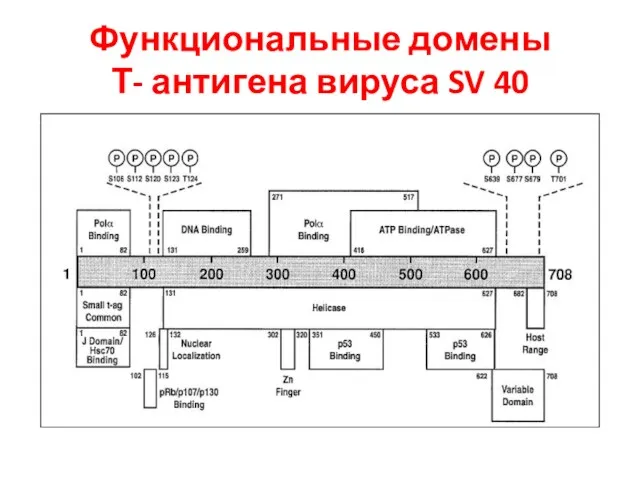
Функциональные домены
Т- антигена вируса SV 40
Функциональные домены
Т- антигена вируса SV 40
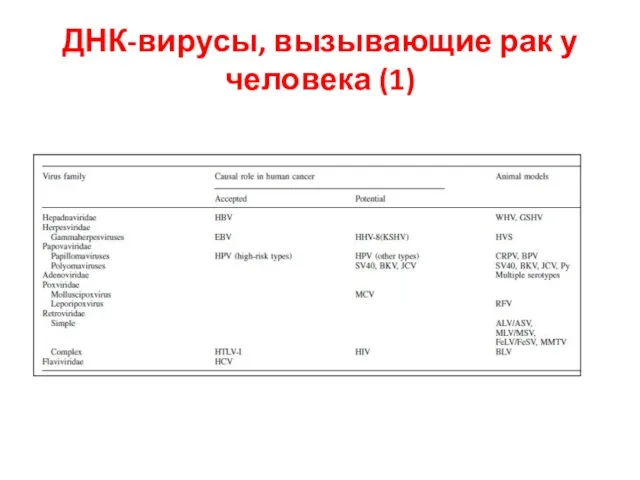
ДНК-вирусы, вызывающие рак у человека (1)
ДНК-вирусы, вызывающие рак у человека (1)
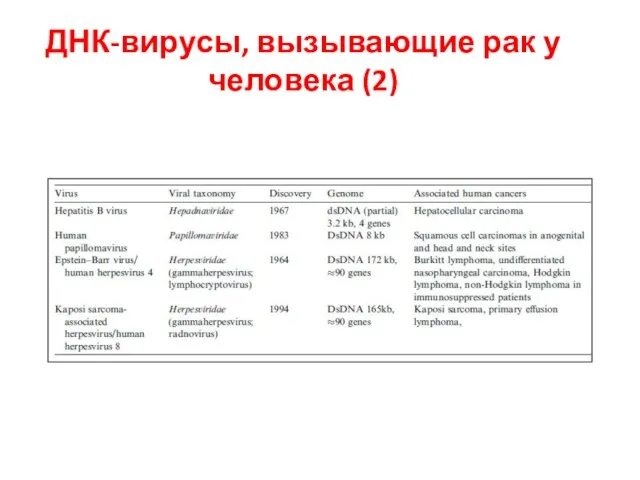
ДНК-вирусы, вызывающие рак у человека (2)
ДНК-вирусы, вызывающие рак у человека (2)
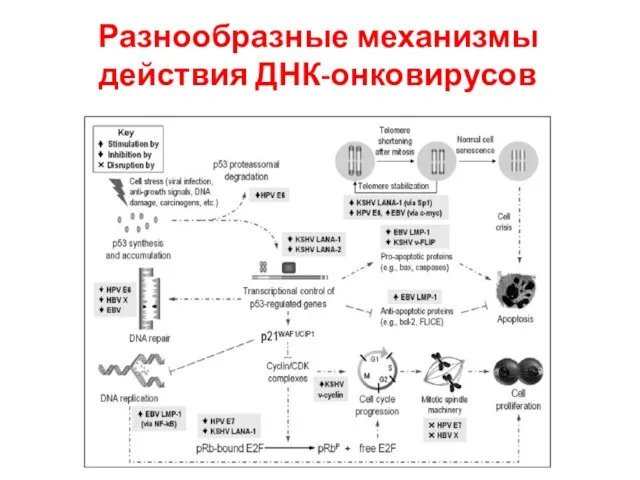
Разнообразные механизмы действия ДНК-онковирусов
Разнообразные механизмы действия ДНК-онковирусов

Мишени для
ДНК-онковирусов
Мишени для
ДНК-онковирусов

Свойства вирусов человека
Свойства вирусов человека

Новые онковирусы
Новое исследование указывает на то, что белок синтезируемый вирусом, вызывающим
Новые онковирусы
Новое исследование указывает на то, что белок синтезируемый вирусом, вызывающим

Основные принципы вирусного онкогенеза
Вирусы могут вызывать рак у животных и человека
Онковирусы
Основные принципы вирусного онкогенеза
Вирусы могут вызывать рак у животных и человека
Онковирусы

Ретровирусы
Ретровирусы
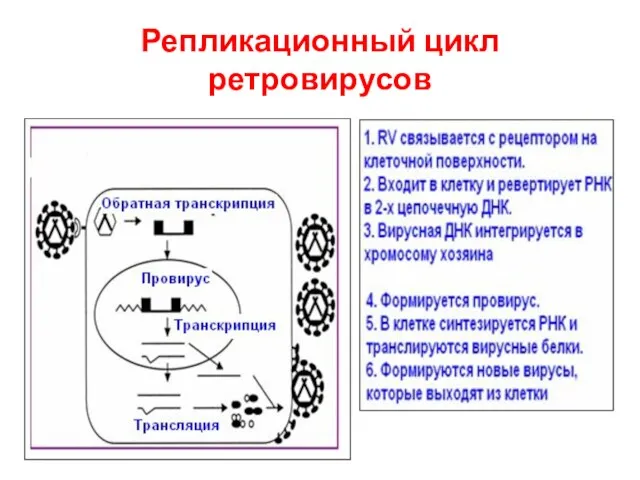
Репликационный цикл ретровирусов
Репликационный цикл ретровирусов

Генетическая организация некоторых ретровирусов
А. «Недефективные» репликационно-компетентные вирусы .
Показаны вирусы с
Генетическая организация некоторых ретровирусов
А. «Недефективные» репликационно-компетентные вирусы .
Показаны вирусы с
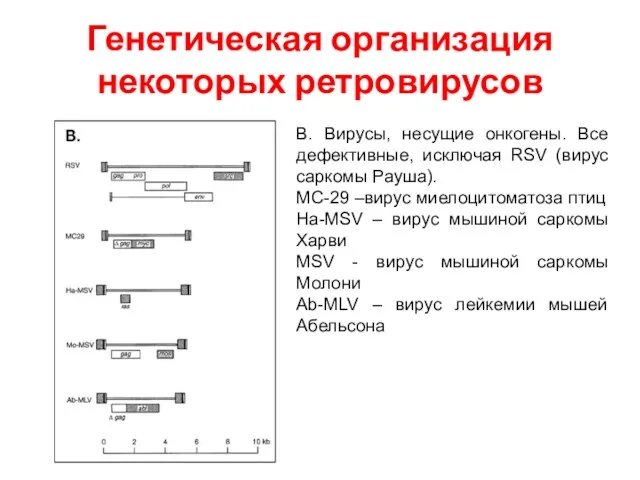
Генетическая организация некоторых ретровирусов
В. Вирусы, несущие онкогены. Все дефективные, исключая RSV
Генетическая организация некоторых ретровирусов
В. Вирусы, несущие онкогены. Все дефективные, исключая RSV
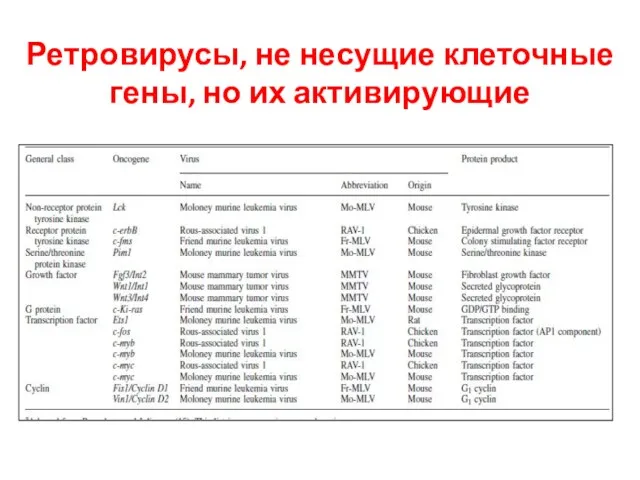
Ретровирусы, не несущие клеточные гены, но их активирующие
Ретровирусы, не несущие клеточные гены, но их активирующие
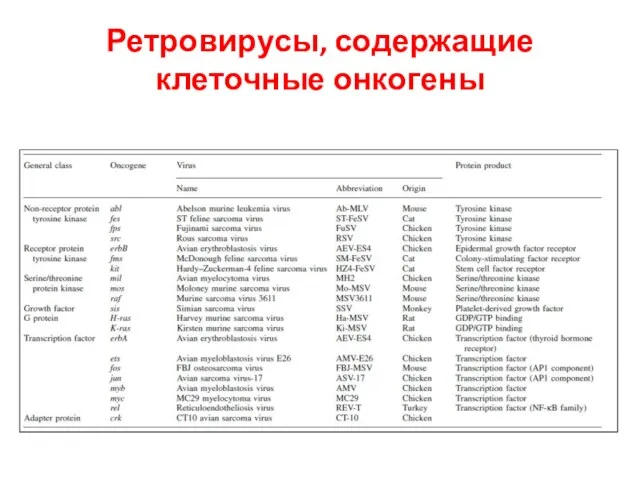
Ретровирусы, содержащие клеточные онкогены
Ретровирусы, содержащие клеточные онкогены

Трансформирующий вирус несет копию гена хозяина
Трансформирующий вирус несет копию гена хозяина
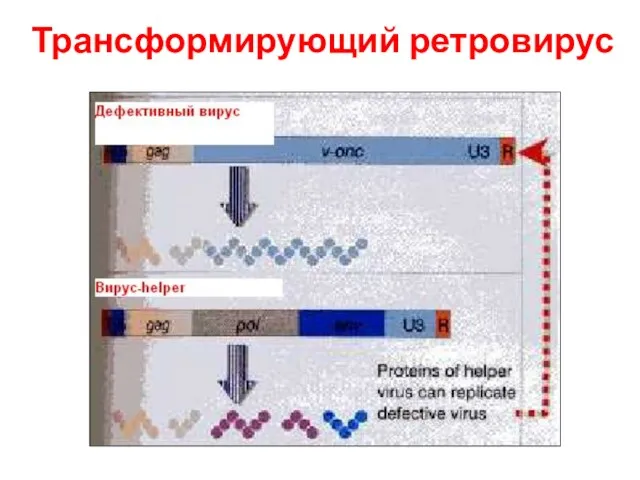
Трансформирующий ретровирус
Трансформирующий ретровирус
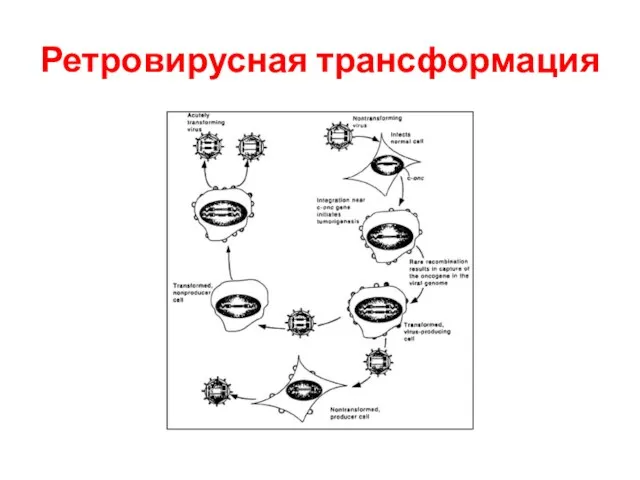
Ретровирусная трансформация
Ретровирусная трансформация

Механизмы активации
прото-онкогенов (ПО)
Траснформирующая «сила» ПО может характеризоваться появлением фокусов.
Тестируемая
Механизмы активации
прото-онкогенов (ПО)
Траснформирующая «сила» ПО может характеризоваться появлением фокусов.
Тестируемая

Вставки, транслокация или амплификация могут активировать ПО
В случае вставки или транслокации
Вставки, транслокация или амплификация могут активировать ПО
В случае вставки или транслокации
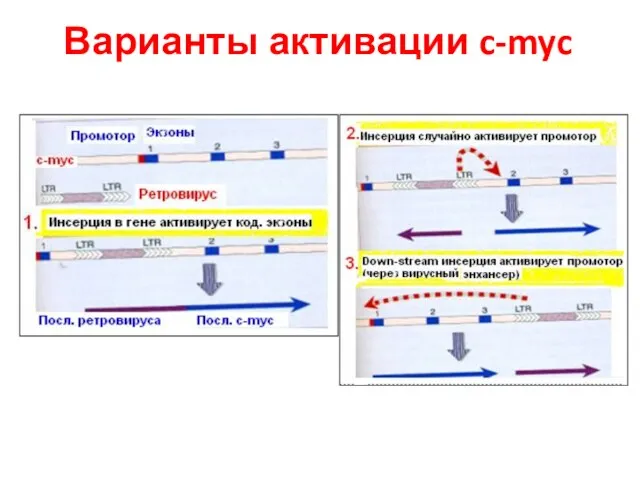
Варианты активации c-myc
Варианты активации c-myc

Другие ПО, активируемые вставкой ретровируса
c-erbB,
c-myb,
c-mos,
c-H-ras,
c-raf
Другие ПО, активируемые вставкой ретровируса
c-erbB,
c-myb,
c-mos,
c-H-ras,
c-raf

Результаты транслокации
Часть белка ПО может быть активирована независимо от другой части
Результаты транслокации
Часть белка ПО может быть активирована независимо от другой части
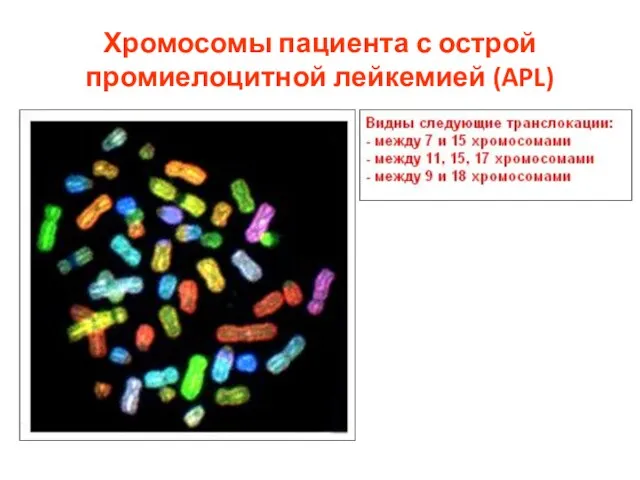
Хромосомы пациента с острой промиелоцитной лейкемией (APL)
Хромосомы пациента с острой промиелоцитной лейкемией (APL)

Филадельфийская хромосома (PH)
Хроническая миелогенная лейкемия (CML) – 5000kb конца 9-й хр.
Филадельфийская хромосома (PH)
Хроническая миелогенная лейкемия (CML) – 5000kb конца 9-й хр.
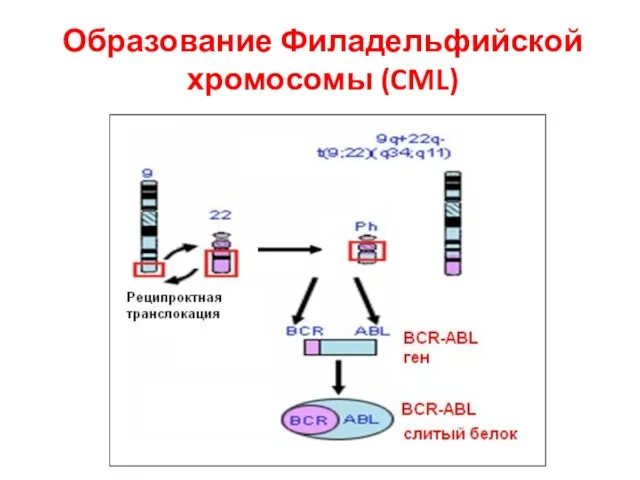
Образование Филадельфийской хромосомы (CML)
Образование Филадельфийской хромосомы (CML)
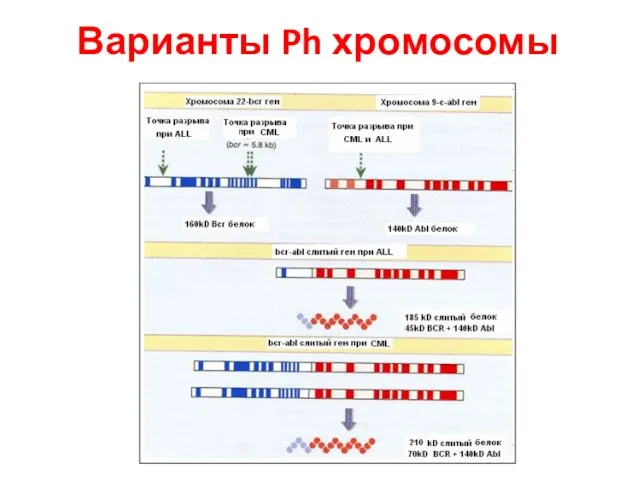
Варианты Ph хромосомы
Варианты Ph хромосомы

Лечение CML
Стандартная терапия хронической миелоидной лейкемии – тирозин киназный ингибитор иматиниб
Лечение CML
Стандартная терапия хронической миелоидной лейкемии – тирозин киназный ингибитор иматиниб
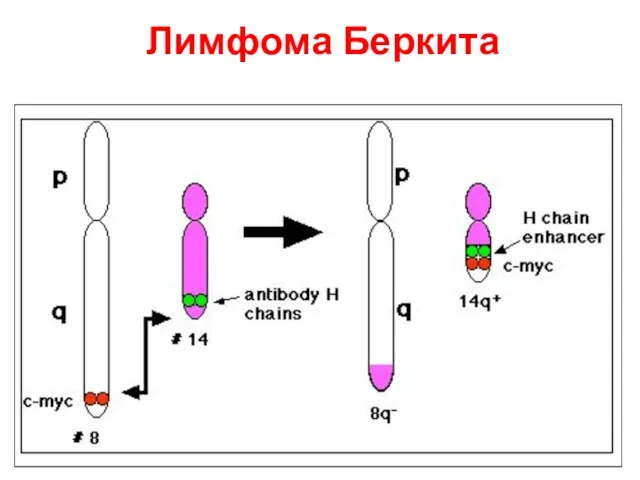
Лимфома Беркита
Лимфома Беркита
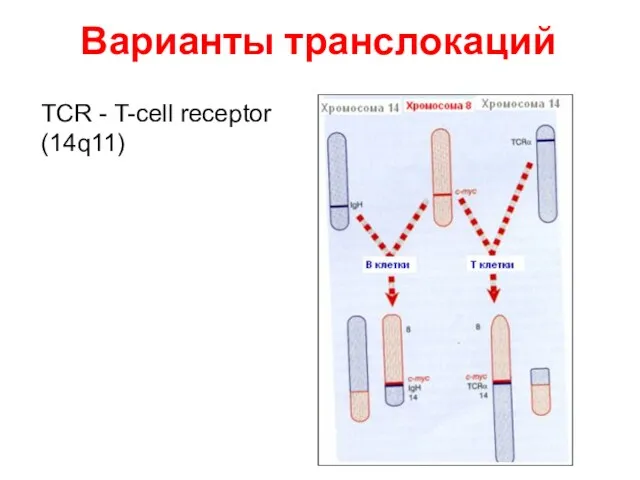
Варианты транслокаций
TCR - T-cell receptor (14q11)
Варианты транслокаций
TCR - T-cell receptor (14q11)
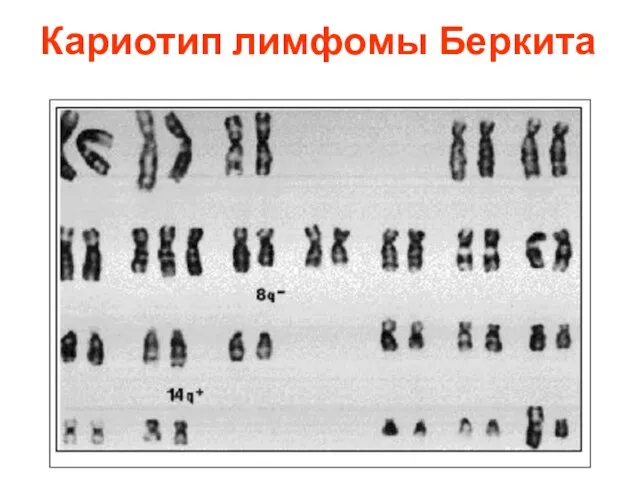
Кариотип лимфомы Беркита
Кариотип лимфомы Беркита

Лимфома Беркита
Лимфома Беркита

Классификация лимфом
Классификация лимфом
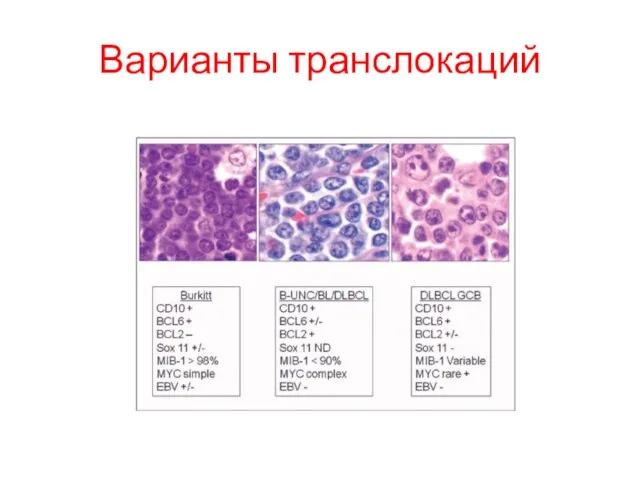
Варианты транслокаций
Варианты транслокаций

5.2.
Передача клеточного сигнала (Сигнальная трансдукция)
5.2.
Передача клеточного сигнала (Сигнальная трансдукция)

5.2.1.
Факторы роста и их рецепторы
5.2.1.
Факторы роста и их рецепторы
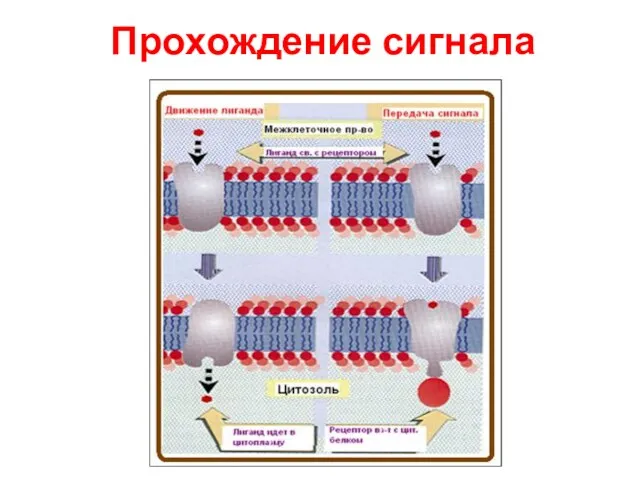
Прохождение сигнала
Прохождение сигнала
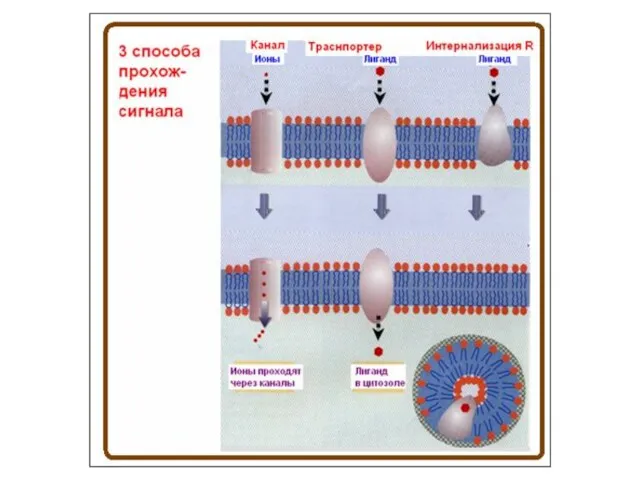
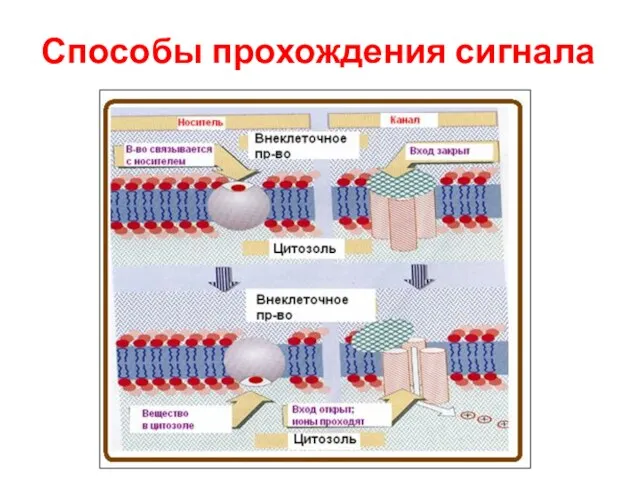
Способы прохождения сигнала
Способы прохождения сигнала
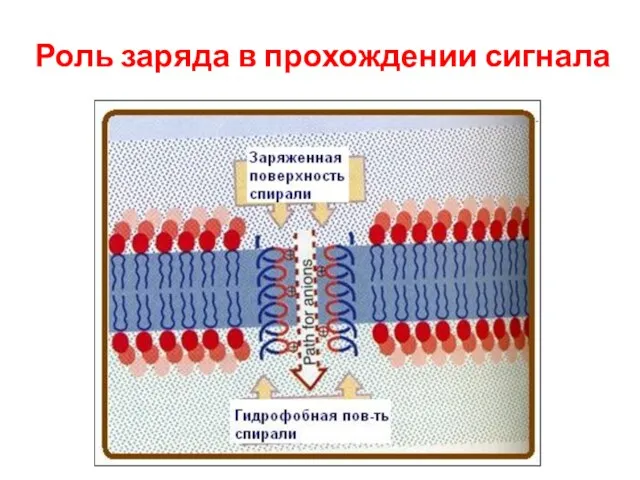
Роль заряда в прохождении сигнала
Роль заряда в прохождении сигнала
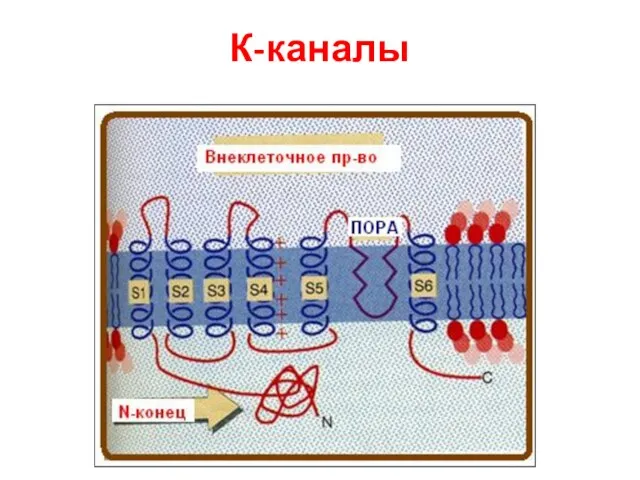
К-каналы
К-каналы
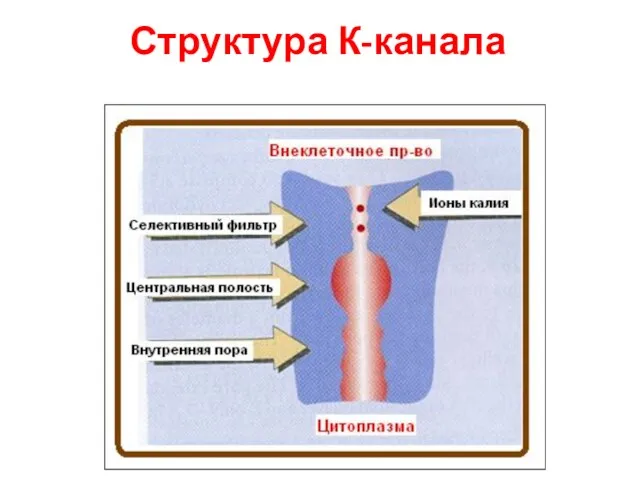
Структура К-канала
Структура К-канала
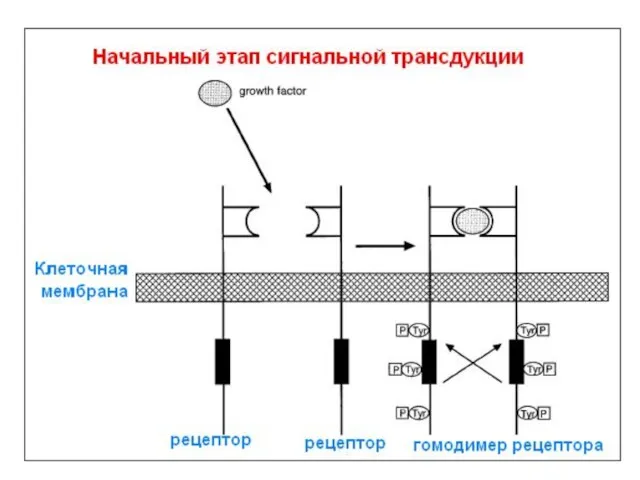

Факторы роста (GF)
К настоящему времени установлено, что семейство факторов роста представлено
Факторы роста (GF)
К настоящему времени установлено, что семейство факторов роста представлено

Процессинг GF
В некоторых случаях, как это показано для фактора NRG-1beta1, его
Процессинг GF
В некоторых случаях, как это показано для фактора NRG-1beta1, его
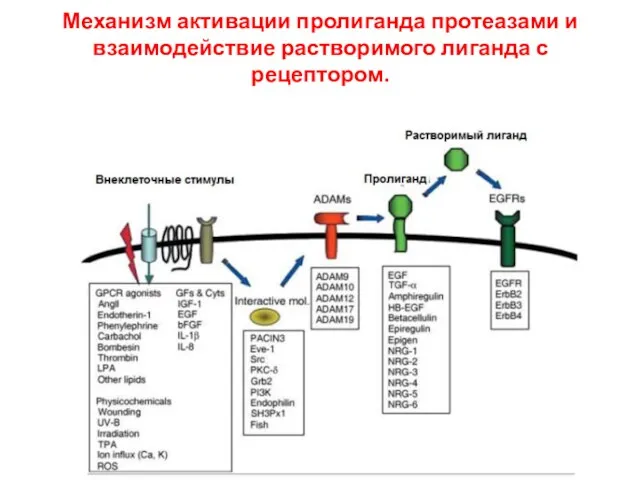
Механизм активации пролиганда протеазами и взаимодействие растворимого лиганда с рецептором.
Механизм активации пролиганда протеазами и взаимодействие растворимого лиганда с рецептором.
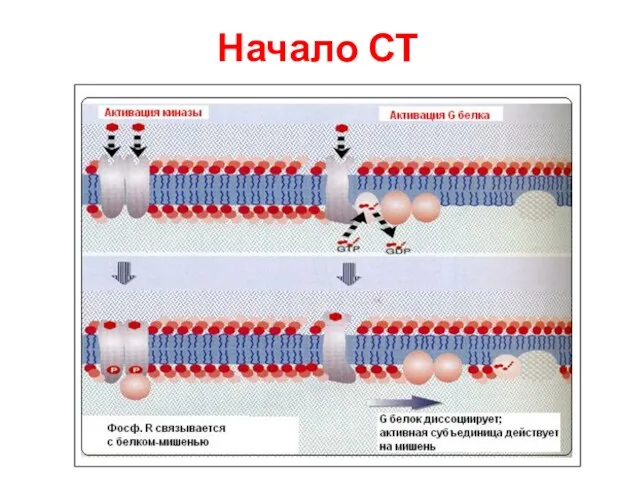
Начало СТ
Начало СТ
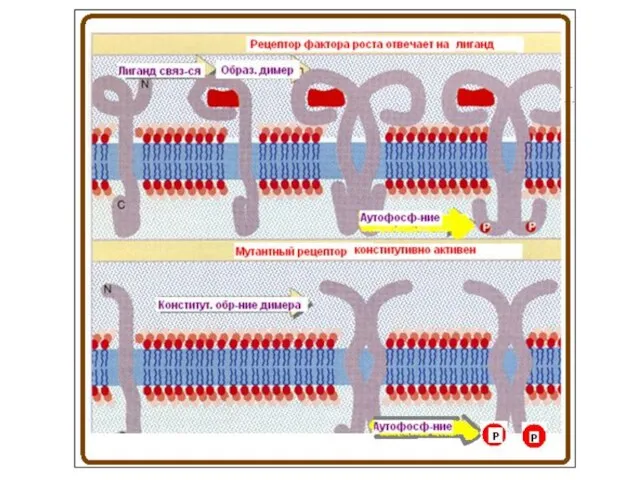

Характеристика RTKs
У человека идентифицировано 58 RTKs, которые подразделяются на 20 подсемейств
Все
Характеристика RTKs
У человека идентифицировано 58 RTKs, которые подразделяются на 20 подсемейств
Все
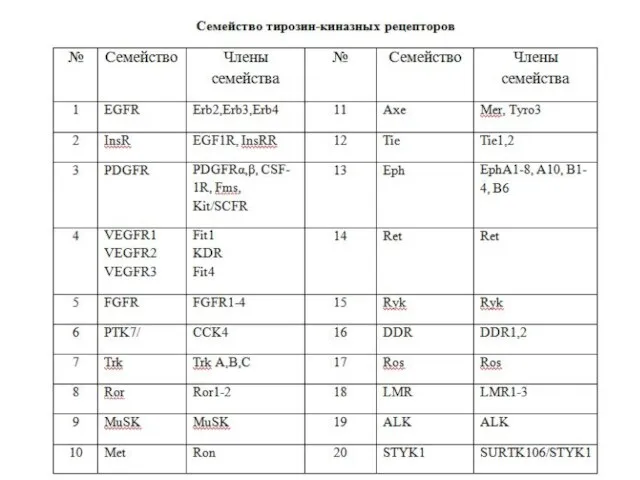
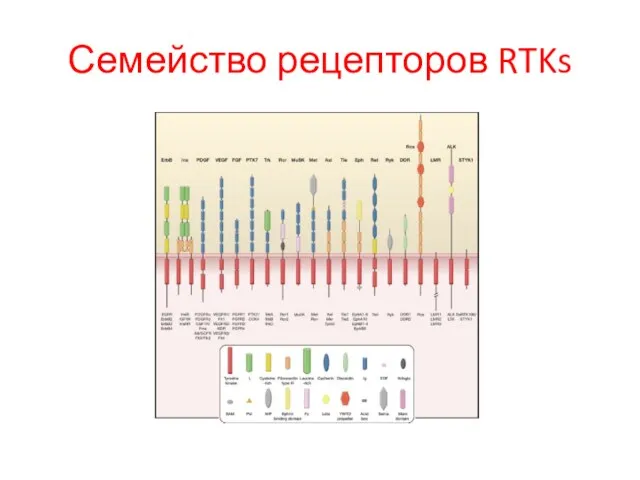
Семейство рецепторов RTKs
Семейство рецепторов RTKs
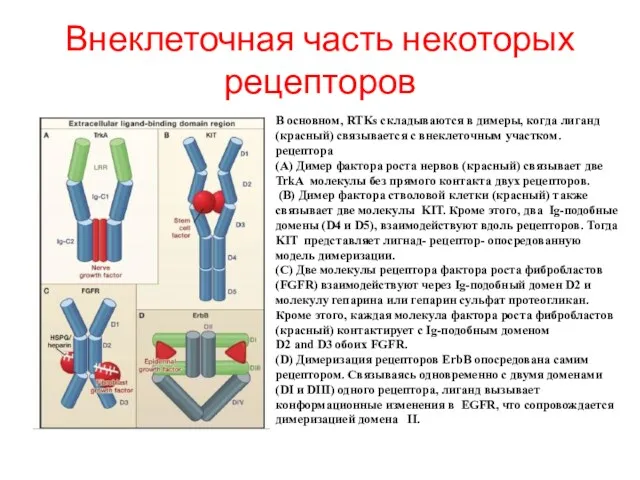
Внеклеточная часть некоторых рецепторов
В основном, RTKs складываются в димеры, когда лиганд
Внеклеточная часть некоторых рецепторов
В основном, RTKs складываются в димеры, когда лиганд

Внутриклеточный домен RTKs
E) Инсулин рецептор-подобные Insulin receptor-like (активация-петля-ингибирование, activation loop inhibition).
Внутриклеточный домен RTKs
E) Инсулин рецептор-подобные Insulin receptor-like (активация-петля-ингибирование, activation loop inhibition).
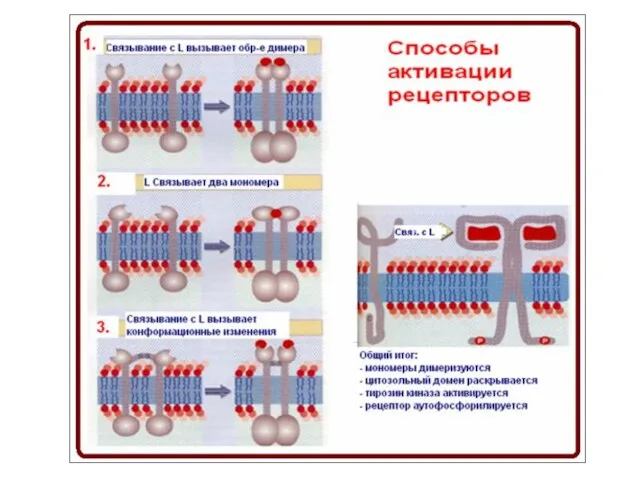
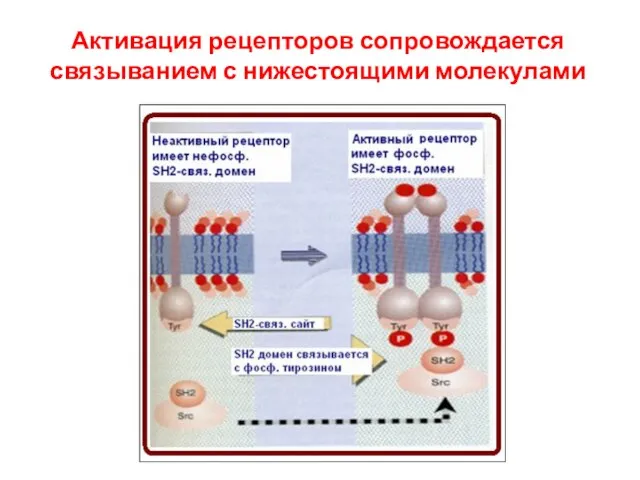
Активация рецепторов сопровождается связыванием с нижестоящими молекулами
Активация рецепторов сопровождается связыванием с нижестоящими молекулами
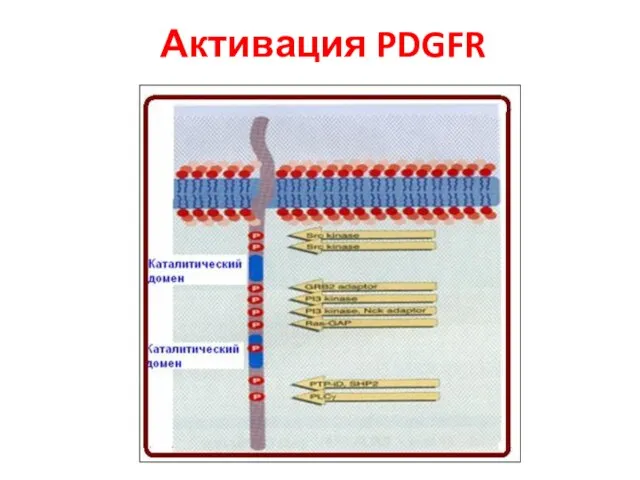
Активация PDGFR
Активация PDGFR
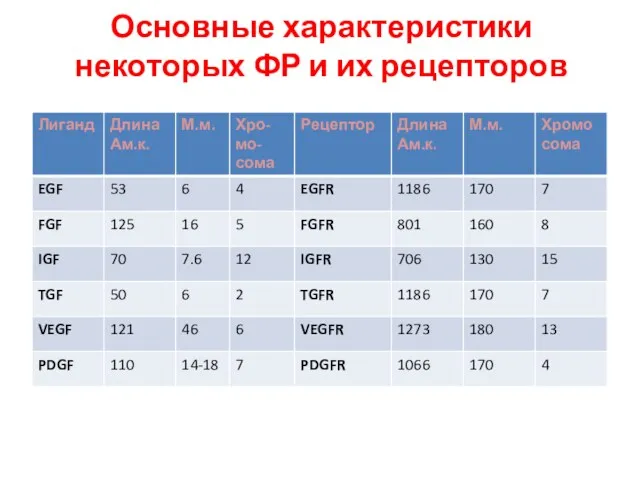
Основные характеристики некоторых ФР и их рецепторов
Основные характеристики некоторых ФР и их рецепторов

Семейство Epidermal growth factor receptor (EGFR)
EGF был изолирован в1962 Stanley Cohen.
Его
Семейство Epidermal growth factor receptor (EGFR)
EGF был изолирован в1962 Stanley Cohen.
Его
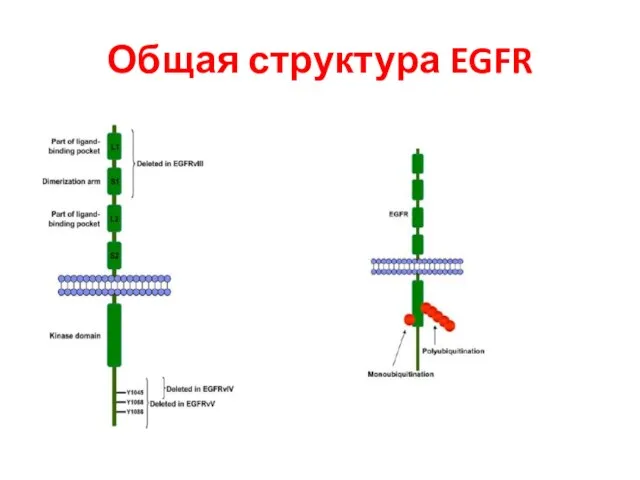
Общая структура EGFR
Общая структура EGFR
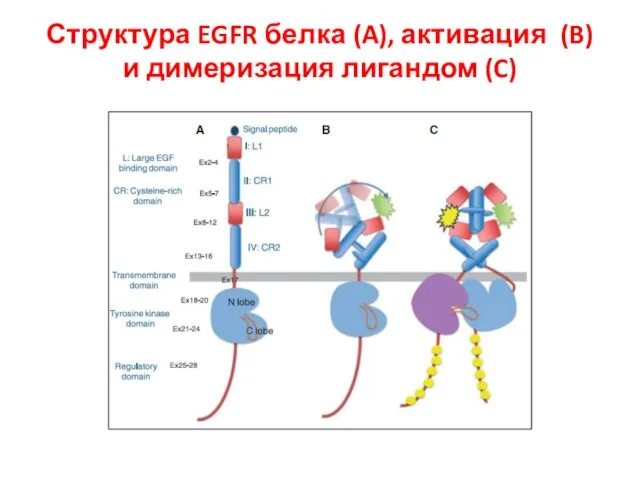
Структура EGFR белка (A), активация (B) и димеризация лигандом (C)
Структура EGFR белка (A), активация (B) и димеризация лигандом (C)

Лиганды для EGFR
Идентифицировано 11 лигандов, которые разделены на 3 группы:
1.EGF,
Лиганды для EGFR
Идентифицировано 11 лигандов, которые разделены на 3 группы:
1.EGF,

Субстраты для EGFR
Phospholipase Cg, CBL, GRB2, SHC и p85.
Включая mitogen-activated
protein
Субстраты для EGFR
Phospholipase Cg, CBL, GRB2, SHC и p85.
Включая mitogen-activated
protein
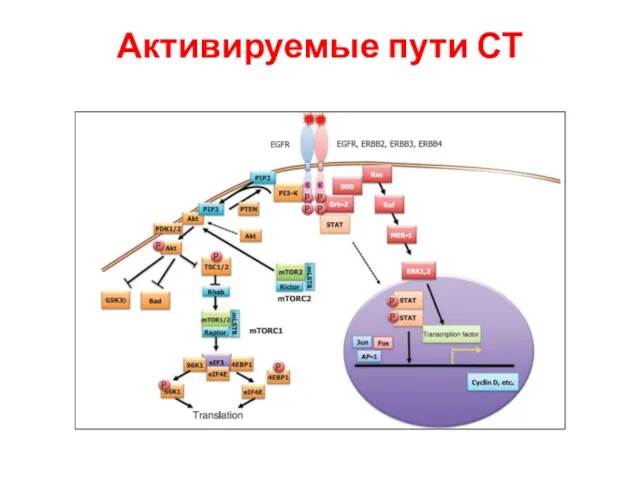
Активируемые пути СТ
Активируемые пути СТ
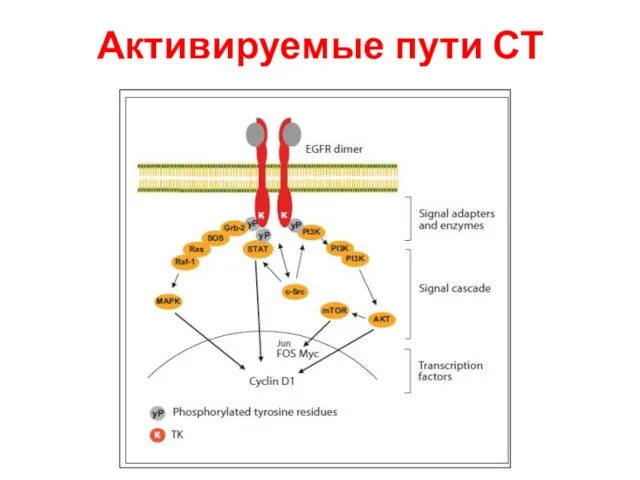
Активируемые пути СТ
Активируемые пути СТ
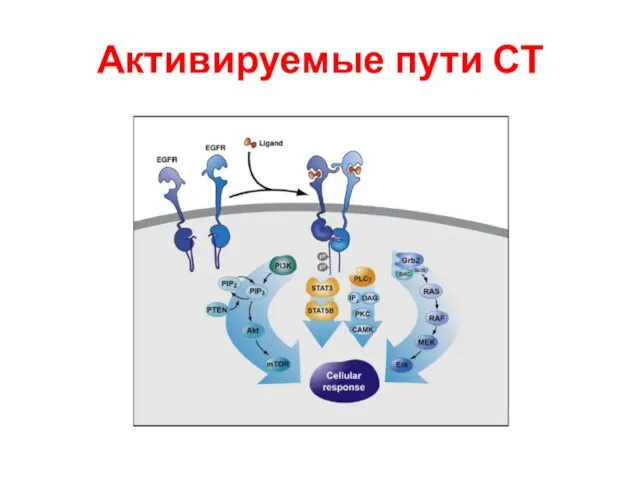
Активируемые пути СТ
Активируемые пути СТ
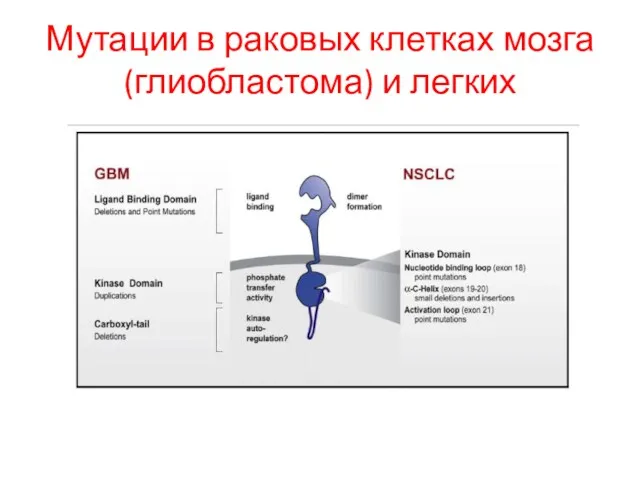
Мутации в раковых клетках мозга (глиобластома) и легких
Мутации в раковых клетках мозга (глиобластома) и легких
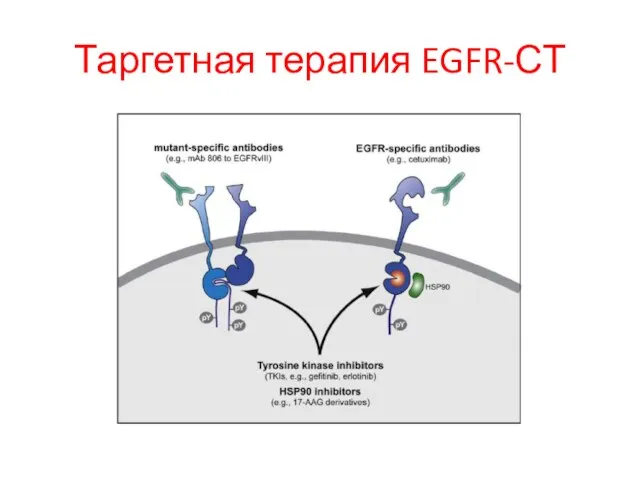
Таргетная терапия EGFR-СТ
Таргетная терапия EGFR-СТ

EGFR и рак
EGFR и рак

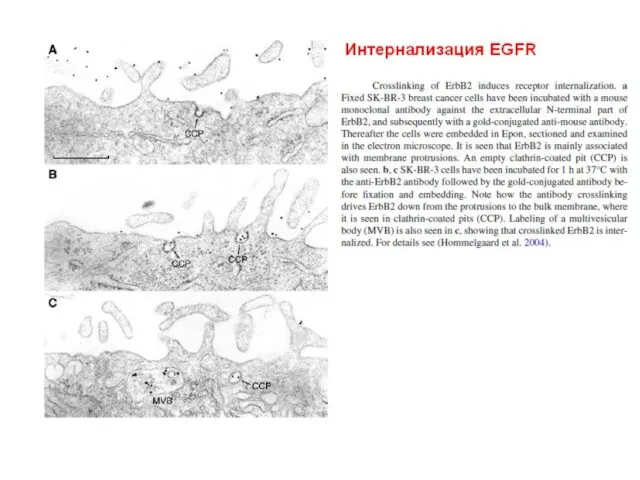
Down-регуляция EGFR
Down-регуляция EGFR
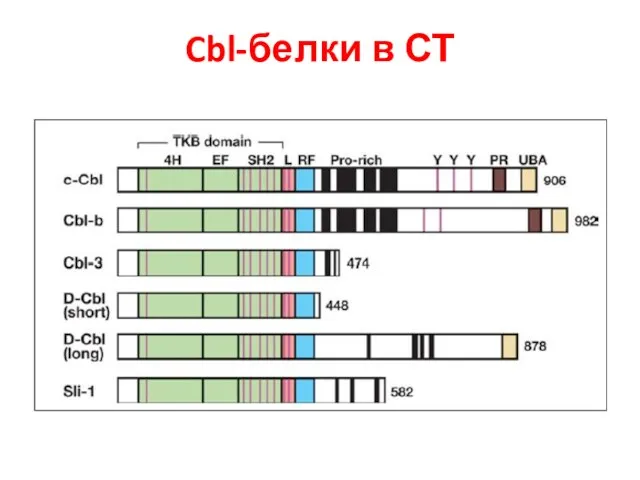
Cbl-белки в СТ
Cbl-белки в СТ
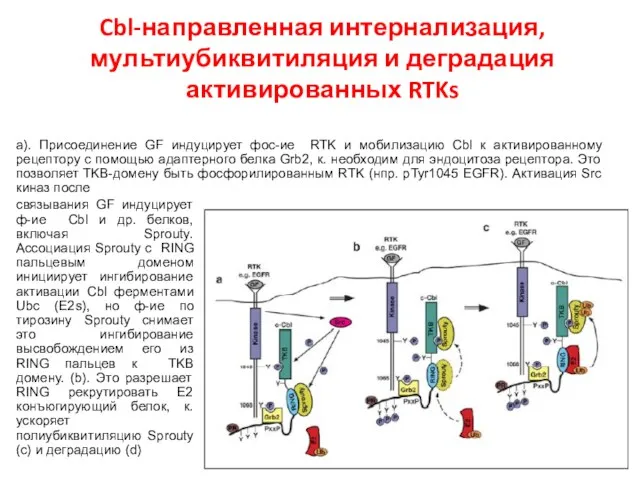
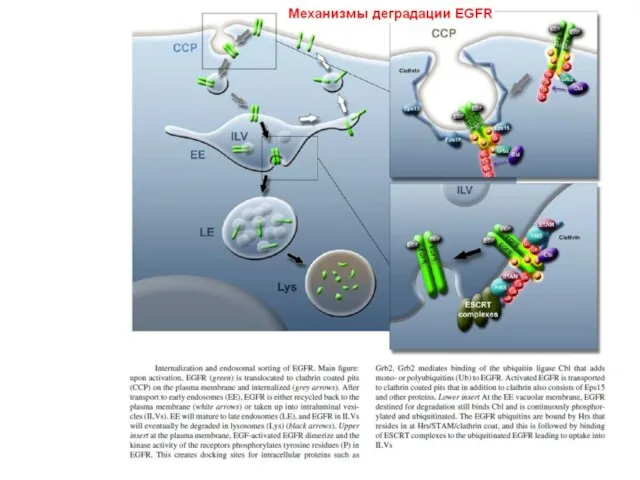
Cbl-направленная интернализация, мультиубиквитиляция и деградация активированных RTKs
а). Присоединение GF индуцирует фос-ие
Cbl-направленная интернализация, мультиубиквитиляция и деградация активированных RTKs
а). Присоединение GF индуцирует фос-ие
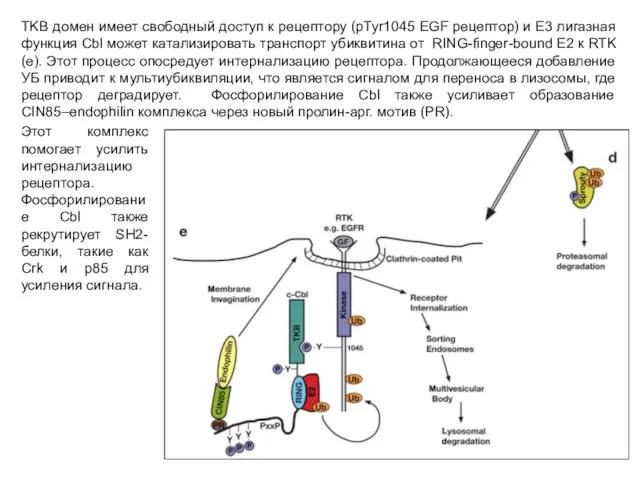
TKB домен имеет свободный доступ к рецептору (pTyr1045 EGF рецептор) и
TKB домен имеет свободный доступ к рецептору (pTyr1045 EGF рецептор) и
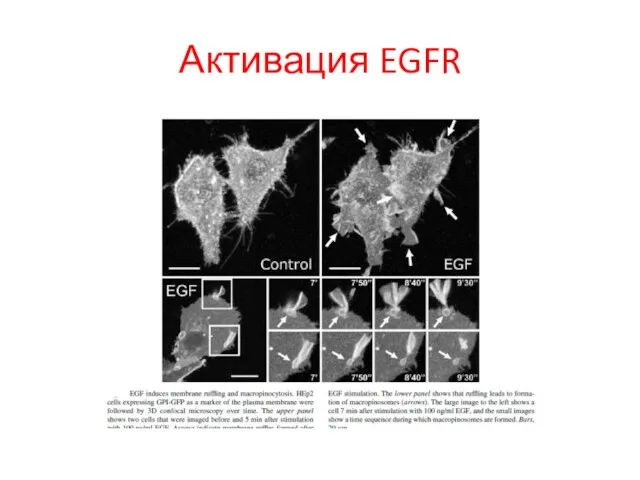
Активация EGFR
Активация EGFR
 Түбір өзектерін толтыруға арналған пломбалық жадығаттар.Жіктелуі, қолдану көрсеткіштері
Түбір өзектерін толтыруға арналған пломбалық жадығаттар.Жіктелуі, қолдану көрсеткіштері Патофизиология пищеварения
Патофизиология пищеварения Классификация эндокринных желез
Классификация эндокринных желез Регуляция менструального цикла
Регуляция менструального цикла Оказание первой помощи при спортивных травмах
Оказание первой помощи при спортивных травмах Оздоровительные базы, санатории (часть 2)
Оздоровительные базы, санатории (часть 2) Гигиенические требования к инфекционным стационарам
Гигиенические требования к инфекционным стационарам Angina Pectoris
Angina Pectoris Вирусные кишечные инфекции
Вирусные кишечные инфекции Лекция по питанию
Лекция по питанию Патронаж здорового новорожденного на дому с оформлением учебной амбулаторной карты
Патронаж здорового новорожденного на дому с оформлением учебной амбулаторной карты Антибиотиктермен емдеудің резистенттілігінің мәселелері және оның шешу жолдары
Антибиотиктермен емдеудің резистенттілігінің мәселелері және оның шешу жолдары Лабораторные методы диагностики вирусных болезней
Лабораторные методы диагностики вирусных болезней Асфиксии новорождённых
Асфиксии новорождённых Показания и противопоказания к лечебному массажу
Показания и противопоказания к лечебному массажу Рак предстательной железы
Рак предстательной железы Клиническая фармакология лекарственных средств, влияющих на гемостаз
Клиническая фармакология лекарственных средств, влияющих на гемостаз Көкрек және оның ағзаларының жарақаттары Кеуденің ауырып қалуы
Көкрек және оның ағзаларының жарақаттары Кеуденің ауырып қалуы Система здравоохранения в России
Система здравоохранения в России Балаларда периодонтиттерді емдеу ерекшеліктері
Балаларда периодонтиттерді емдеу ерекшеліктері Лейкозы у детей
Лейкозы у детей Ишемический колит
Ишемический колит Основы черпно-челюстнолицевой хирургии. Будущее специальности. Врожденные и приобретенные дефекты ЧЛО
Основы черпно-челюстнолицевой хирургии. Будущее специальности. Врожденные и приобретенные дефекты ЧЛО Очаговый туберкулез
Очаговый туберкулез Атрофические заболевания головного мозга. Болезнь альцгеймера. Болезнь пика
Атрофические заболевания головного мозга. Болезнь альцгеймера. Болезнь пика Ана және бала өлімін төмендету аспектісіндегі халық денсаулығын сақтау және нығайтудағы халықаралық тәжірибе
Ана және бала өлімін төмендету аспектісіндегі халық денсаулығын сақтау және нығайтудағы халықаралық тәжірибе Қоршаған орта мен тұрғындар денсаулығы үшін Қазақстан аймақтарындағы көп жылдық ядролық сынақтар салдары
Қоршаған орта мен тұрғындар денсаулығы үшін Қазақстан аймақтарындағы көп жылдық ядролық сынақтар салдары Инфекции мочевыводящих путей в амбулаторной прaктике на примере Республики Карелия: диагностика и лечение
Инфекции мочевыводящих путей в амбулаторной прaктике на примере Республики Карелия: диагностика и лечение