- Ревматология. Иммуногенетика

Содержание
- 2. Жоспары: 1. кіріспе 1.1 иммуногенетика түсінігі 1.2 Гистосәйкестіктің басты комплексі және қызметі
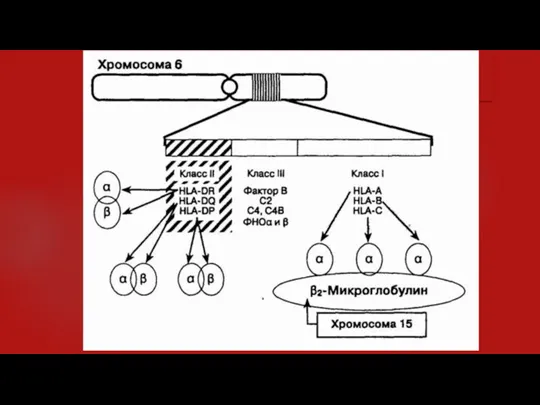
- 3. 2. Что такое главный комплекс гистосовместимости и какова его функция? ГКГ локализован в коротком плече 6-й
- 5. 3. Существуют ли отличия между комплексом гистосовместимости и человеческим лейкоцитарным антигеном (HLA)? В практических целях эти
- 6. 8. В чем смысл букв-окончаний (А, В...) при аббревиатуре HLA? В аббревиатурах HLA с последующим написанием
- 7. 10. Как буквы (области) А, В, С, D соотносятся с классом комплекса гистосовместимости? Молекулы класса I
- 8. 11. Как происходит связывание антигена? Все антигенсвязывающие участки имеют схожую конфигурацию. Существует углубление-борозда, стенки которой образуют
- 9. Антигенсвязывающий участок
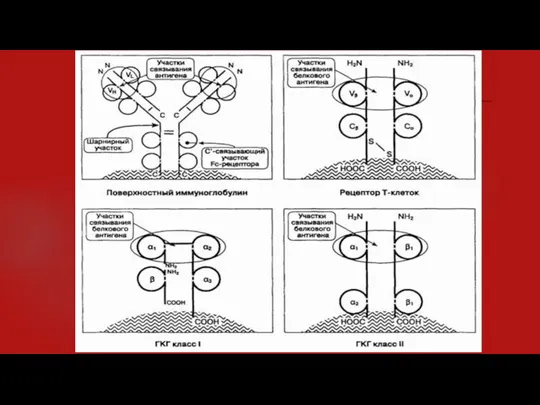
- 10. Иммуноглобулины, Т-клеточные рецепторы и молекулы ГКГ класса I и II являются трансмембранными, поверхностно-клеточными молекулами (см. рисунок).
- 14. Скачать презентацию

Жоспары:
1. кіріспе
1.1 иммуногенетика түсінігі
1.2 Гистосәйкестіктің басты комплексі және қызметі
Жоспары:
1. кіріспе
1.1 иммуногенетика түсінігі
1.2 Гистосәйкестіктің басты комплексі және қызметі

2. Что такое главный комплекс гистосовместимости и какова его функция?
ГКГ
2. Что такое главный комплекс гистосовместимости и какова его функция?
ГКГ

3. Существуют ли отличия между комплексом гистосовместимости и человеческим лейкоцитарным антигеном
3. Существуют ли отличия между комплексом гистосовместимости и человеческим лейкоцитарным антигеном

8. В чем смысл букв-окончаний (А, В...) при аббревиатуре HLA?
В
8. В чем смысл букв-окончаний (А, В...) при аббревиатуре HLA?
В

10. Как буквы (области) А, В, С, D соотносятся с классом

11. Как происходит связывание антигена?
Все антигенсвязывающие участки имеют схожую конфигурацию.
11. Как происходит связывание антигена?
Все антигенсвязывающие участки имеют схожую конфигурацию.
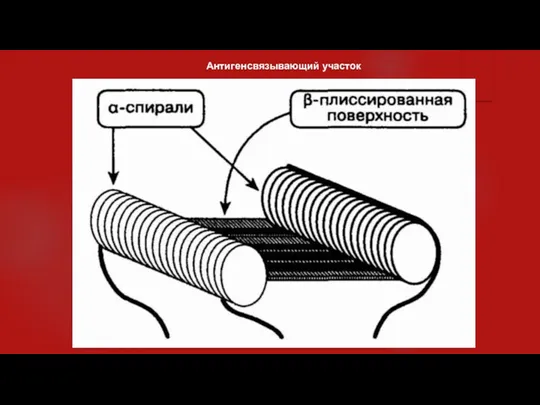
Антигенсвязывающий участок
Антигенсвязывающий участок


 Гигиенические требования к белью, комнатной одежде, одежды для прогулок и обуви детей от года до семи лет
Гигиенические требования к белью, комнатной одежде, одежды для прогулок и обуви детей от года до семи лет Гельминтозы собак
Гельминтозы собак Туберкулез костей и суставов
Туберкулез костей и суставов Нестероидные противовоспалительные средства (НПВС)
Нестероидные противовоспалительные средства (НПВС) Физическая реабилитация больных со сколиозом
Физическая реабилитация больных со сколиозом Типы и причины нарушения слуха
Типы и причины нарушения слуха Чесотка. Возбудители чесотки. Симптомы и лечение. Профилактика чесотки
Чесотка. Возбудители чесотки. Симптомы и лечение. Профилактика чесотки Деменции - клиника и этиология
Деменции - клиника и этиология Предмет иммунологии. История иммунологии
Предмет иммунологии. История иммунологии Отделение медицинской реабилитации взрослых для пациентов с соматическими заболеваниями
Отделение медицинской реабилитации взрослых для пациентов с соматическими заболеваниями Противобластомные (противоопухолевые)
Противобластомные (противоопухолевые) Оценка новорожденного ребенка сразу после рождения и принципы ухода за новорожденным
Оценка новорожденного ребенка сразу после рождения и принципы ухода за новорожденным Проект на тему: Моя будущая профессия – ветеринарный фельдшер
Проект на тему: Моя будущая профессия – ветеринарный фельдшер Болезнь Рейтера
Болезнь Рейтера Діріл. Пайда болатын аурулар. Дірілдің алдын алу шаралары
Діріл. Пайда болатын аурулар. Дірілдің алдын алу шаралары Лекарственные средства, влияющие на функцию органов дыхания
Лекарственные средства, влияющие на функцию органов дыхания Учение об инфекции патогенность и вирулентность микробов
Учение об инфекции патогенность и вирулентность микробов Клинико-психологическое сопровождение в рамках третичной профилактики пожилых с болезнью Альцгеймера на ранней стадии
Клинико-психологическое сопровождение в рамках третичной профилактики пожилых с болезнью Альцгеймера на ранней стадии Повреждения и неотложные хирургические заболевания органов брюшной полости. Часть 2
Повреждения и неотложные хирургические заболевания органов брюшной полости. Часть 2 Методы исследования детей с различными заболеваниями сердца
Методы исследования детей с различными заболеваниями сердца Норма и отклонение в физическом, психическом, интеллектуальном, речевом и сенсорном развитии ребенка
Норма и отклонение в физическом, психическом, интеллектуальном, речевом и сенсорном развитии ребенка Синдром после тотальной овариоэктоми. Посткастрационный синдром
Синдром после тотальной овариоэктоми. Посткастрационный синдром Режимы дня, труда и отдыха
Режимы дня, труда и отдыха Возможности ультразвуковой ангиологии в диагностике неврологических заболеваний
Возможности ультразвуковой ангиологии в диагностике неврологических заболеваний Заболевания желудка
Заболевания желудка Ауыздың кілегейлі қабығының гиперкератотикалық өзгерістері бар науқастарға көмекті жоспарлау
Ауыздың кілегейлі қабығының гиперкератотикалық өзгерістері бар науқастарға көмекті жоспарлау Зардап шеккендер мен науқастарды жедел көмек көлігімен тасымалдау
Зардап шеккендер мен науқастарды жедел көмек көлігімен тасымалдау Частичное вторичное отсутствие зубов на нижней челюсти
Частичное вторичное отсутствие зубов на нижней челюсти