- Секреты адипоцитов, эволюция представлений

Содержание
- 2. «Size, sites and cytes» Matthias Blucher, Stockholm, EASD 17.09.2015
- 3. Изменение представлений о жировой ткани в последние 20 лет До 80-х годов депо энергии органопротекция защита
- 4. Адипоциты
- 5. Физиология адипоцитов тогда и сейчас
- 6. Адипоцит
- 7. Признаками здоровой жировой ткани являются следующие: высокая плотность сосудов в пределах жирового депо; минимальная гипоксия и
- 8. Адипоцит
- 9. Адипоциты могут: Остаться неизменными, Увеличиться в размерах Уменьшиться Подвергнуться апоптозу или некрозу NB!!! Также они могут
- 10. Дедифференцировка и повторная ре-дифференцировка как часть жизненного цикла адипоцитов
- 11. Адипоциты
- 12. Адипоциты Белые адипоциты имеют крупные липидные капли, окруженные небольшим количеством цитоплазмы и смешенным к периферии ядром.
- 13. Расположение бурого жира у взрослого человека Гистология и ПЭТ Гистология Цервикальная область Надключичная Околопозвоночная Подмышечная Медиастинальная
- 14. Разница в содержании МХ между«белым», «бежевым» и «бурым» адипоцитами
- 15. В базальном состоянии адипоциты функционируют как адипоциты белой жировой ткани, но при адекватном раздражителе они могут
- 16. Белая жировая ткань
- 17. Бежевая жировая ткань
- 18. Бурая жировая ткань
- 19. Adipose Tissue Stem Cell
- 20. Бежевые адипоциты дифференцируются от клеток-предшественников эмбриональной мезодермы, которые экспрессируют Myf5 и Pax7 Две концепции происхождения адипоцитов.
- 21. Происхождение адипоцитов J.Dempersmier et al. 2015
- 22. Бурые адипоциты считаются метаболически более эффективными: всего лишь 63 г полностью активированных термогенных адипоцитов могут сжечь
- 23. Тем не менее существует мнение, что эктопическая экспрессия разобщающего белка 1 (UCP1) и протеина PR domain
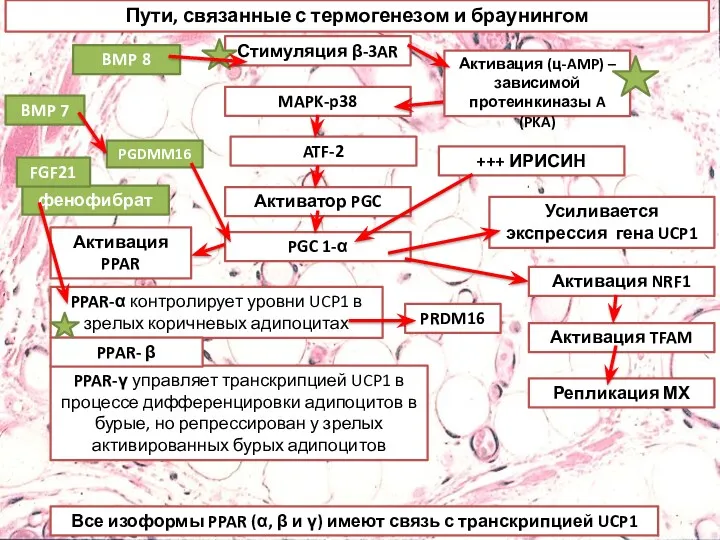
- 24. Пути, связанные с термогенезом и браунингом Стимуляция β-3AR MAPK-p38 ATF-2 Активатор PGC PGC 1-α Активация PPAR
- 25. Пути, связанные с термогенезом и браунингом
- 26. Пути, связанные с термогенезом и браунингом. Стимуляция бета-3-адренергическими рецепторами приводит к индукции PGC1, которая управляет активацией
- 27. Пути, связанные с термогенезом и браунингом Стимуляция β-3AR MAPK-p38 ATF-2 Активатор PGC PGC 1-α Активация PPAR
- 28. Фармакологическая индукция превращения белой жировой ткани в бурую Адренергическая стимуляция 5'АМФ-активируемая протеинкиназа-АМФК FGF21 Никотин Костные морфогенетические
- 29. Пути, связанные с термогенезом и браунингом Стимуляция β-3AR MAPK-p38 ATF-2 Активатор PGC PGC 1-α Активация PPAR
- 30. Пути, связанные с термогенезом и браунингом НУП
- 31. Адипоинсулярная ось обуславливает взаимодействие между инсулином и лептином для контроля аппетита и выработки глюкозы. Инсулин и
- 32. Алиментарная индукция превращения белой жировой ткани в бурую Капсаицин Ограничение потребления метионина Фукоксантин, каротиноид флавоноид лютеолин,
- 33. Пути, связанные с термогенезом и браунингом
- 34. Холодовая адаптация: рассеивание энергии, несократительный термогенез
- 35. Холодовая адаптация: рассеивание энергии, несократительный термогенез. Производство тепла начинается мгновенно с помощью дрожи - это прямая
- 36. Главное: –– Браунинг это приобретение буроподобного фенотипа адипоцитами белой жировой ткани, преимущественно адипоцитами подкожной жировой клетчатки
- 37. Главное: –– капсаицин, желчные кислоты, ресвератрол, ретиноевая кислота и некоторые липиды являются одними из наиболее изученных
- 38. Адипоцит как эндокринная клетка
- 39. Адипоцит как эндокринная клетка
- 40. NPY гипоталамуса Лептин приема пищи секреции инсулина - Лептин - NPY
- 41. Лептин - продукт (гормон) жировой ткани (ob ген) Концентрация прямо пропорциональна степени ожирения Связь с уровнем
- 42. Жировая ткань Л Е П Т И Н Ожирение Еда Инсулин Глюкокортикоиды Цитокины (TNF- α, IL-1)
- 43. Адипонектин универсальный антилипотоксичный агент имеет обратную связь с ожирением Оказывает противовоспалительное, инсулин-сенсибилизирующие и антиапоптотическое действие
- 44. Адипонектин Антиатерогенные эффекты Снижение экспрессии молекул адгезии Снижение адгезии моноцитов к эндотелию Снижение захвата окисленных LDL
- 45. Повышенный уровень церамидов ассоциируется с инсулинорезистентностью, воспалением и апоптозом Церамиды превращаются в сфингозины, что увеличивает клеточную
- 46. Гипотеза «экономного» генотипа Дополнительные калории и их отложение в виде триглицеридов способствовало выживанию, а значит, происходила
- 47. “Не настолько экономный генотип” Выживание Эффективная защита мышечной массы Снижение протеолиза (ЦНС) Инсулинорезистентность продолжительности жизни массы
- 48. Инсулинорезистентность Гиперинсулинемия Нарушения толерантности к глюкозе Гипертриглицеридемия Снижением ЛПВП и повышение ЛПНП Артериальная гипертензия Метаболический синдром
- 49. Андроидное ожирение Нарушение толерантности к глюкозе Гиперинсулинемия Артериальная гипертензия + гиперурикемия, ГЛЖ, гиперфибирногенемия, и др. «Смертельный
- 50. Инсулинорезистентность может быть определена как ситуация, в которой физиологическое количество инсулина не обеспечивает адекватного биологического эффекта
- 51. Insulin resistance is defined by the decreased ability of insulin to stimulate glucose disposal into target
- 52. Инсулинорезистентность: основная причина MetS и СД2 или защитный адаптивный ответ?
- 53. С.Nolan and Marc Prentki 2019 Молекулярная основа инсулин-индуцированного метаболического стресса
- 54. С.Nolan and Marc Prentki 2019
- 55. Новая концептуальная основа для рассмотрения резистентности к инсулину и метаболического синдрома (MetS) и связанных с ними
- 57. Скачать презентацию

«Size, sites and cytes»
Matthias Blucher, Stockholm, EASD 17.09.2015
«Size, sites and cytes»
Matthias Blucher, Stockholm, EASD 17.09.2015

Изменение представлений о жировой ткани в последние 20 лет
До 80-х годов
депо
Изменение представлений о жировой ткани в последние 20 лет
До 80-х годов
депо
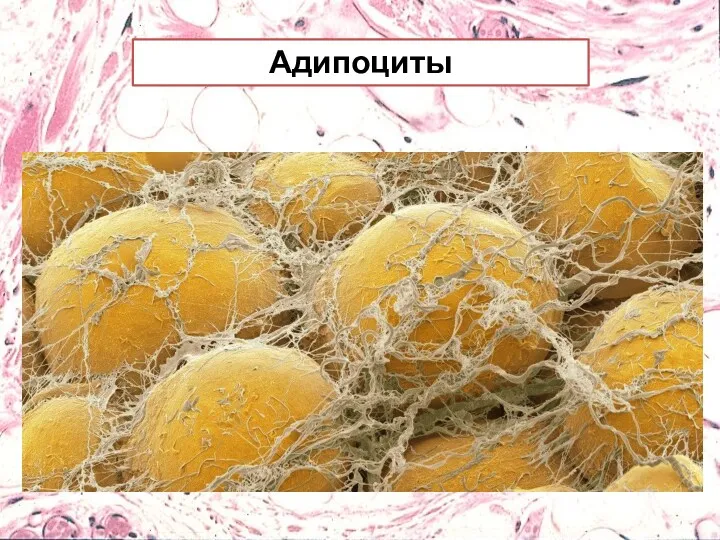
Адипоциты
Адипоциты
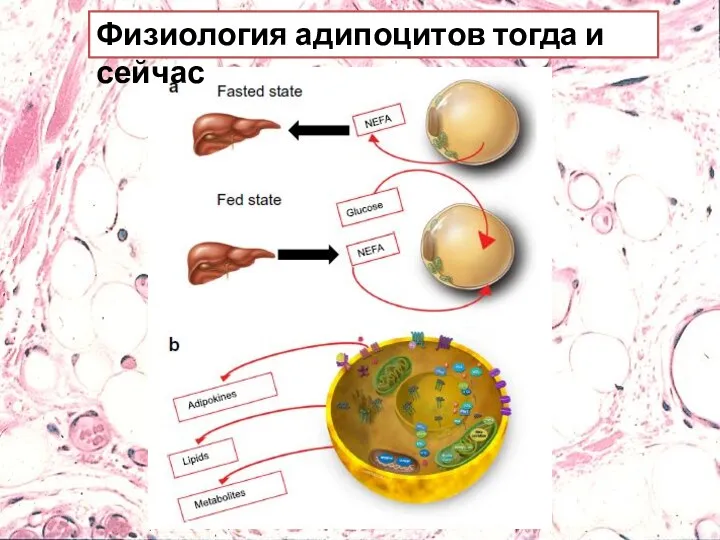
Физиология адипоцитов тогда и сейчас
Физиология адипоцитов тогда и сейчас
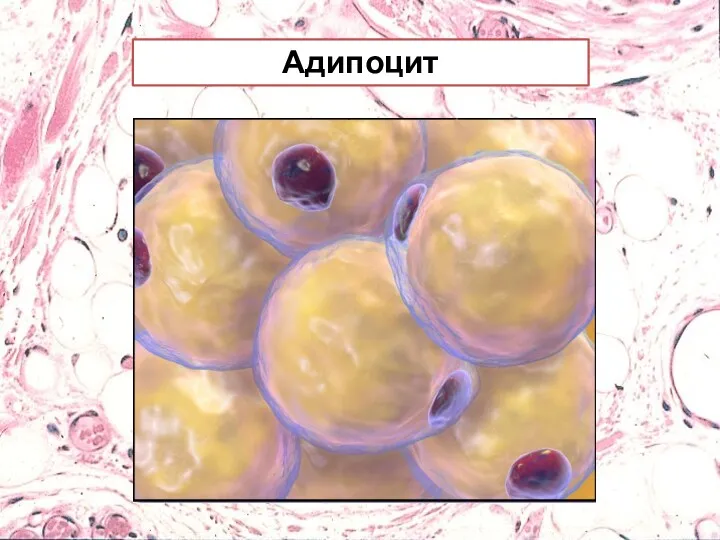
Адипоцит
Адипоцит

Признаками здоровой жировой ткани являются следующие:
высокая плотность сосудов в
Признаками здоровой жировой ткани являются следующие:
высокая плотность сосудов в
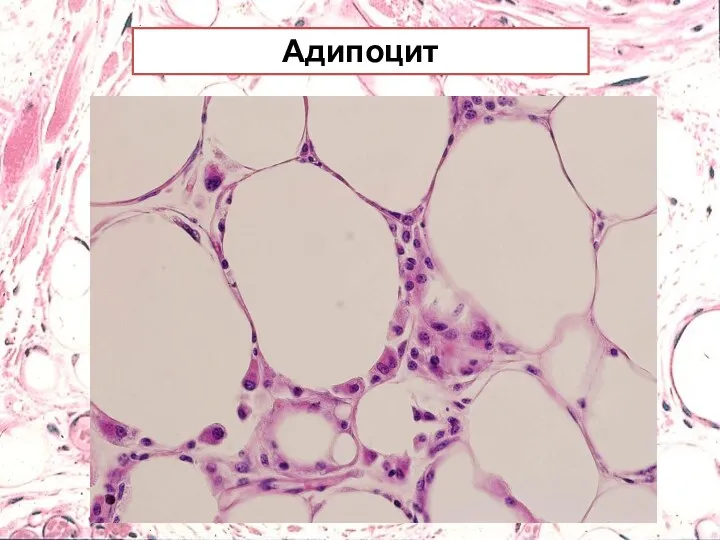
Адипоцит
Адипоцит

Адипоциты могут:
Остаться неизменными,
Увеличиться в размерах
Уменьшиться
Подвергнуться апоптозу или некрозу
NB!!! Также
Адипоциты могут:
Остаться неизменными,
Увеличиться в размерах
Уменьшиться
Подвергнуться апоптозу или некрозу
NB!!! Также
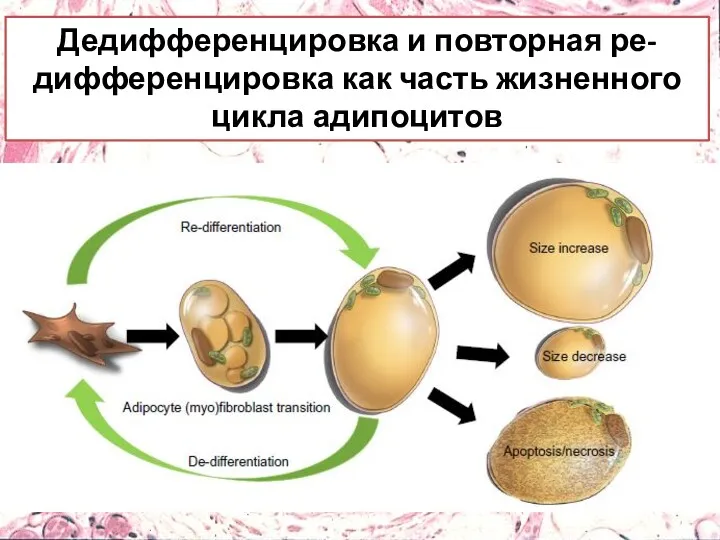
Дедифференцировка и повторная ре-дифференцировка как часть жизненного цикла адипоцитов
Дедифференцировка и повторная ре-дифференцировка как часть жизненного цикла адипоцитов
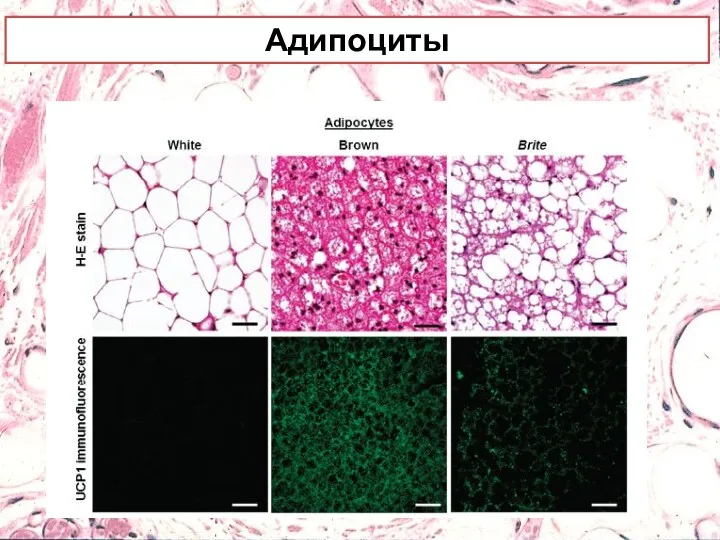
Адипоциты
Адипоциты

Адипоциты
Белые адипоциты имеют крупные липидные капли, окруженные небольшим количеством цитоплазмы
Адипоциты
Белые адипоциты имеют крупные липидные капли, окруженные небольшим количеством цитоплазмы
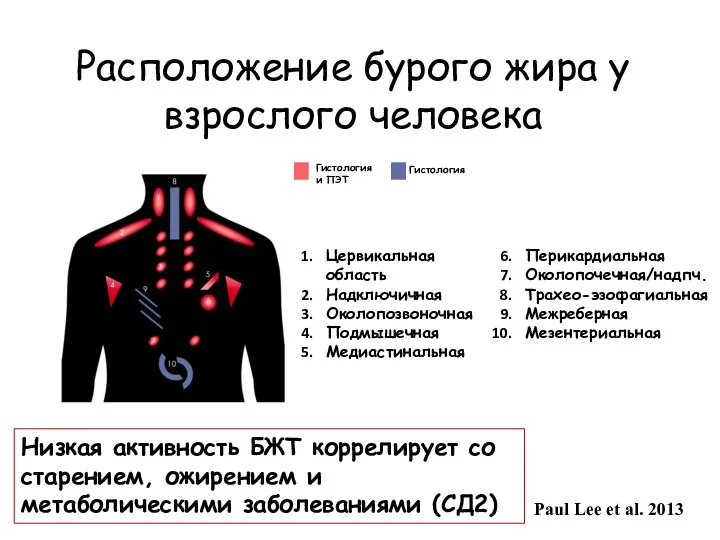
Расположение бурого жира у взрослого человека
Гистология и ПЭТ
Гистология
Цервикальная область
Надключичная
Околопозвоночная
Подмышечная
Медиастинальная
Перикардиальная
Околопочечная/надпч.
Трахео-эзофагиальная
Межреберная
Мезентериальная
Paul Lee
Расположение бурого жира у взрослого человека
Гистология и ПЭТ
Гистология
Цервикальная область
Надключичная
Околопозвоночная
Подмышечная
Медиастинальная
Перикардиальная
Околопочечная/надпч.
Трахео-эзофагиальная
Межреберная
Мезентериальная
Paul Lee
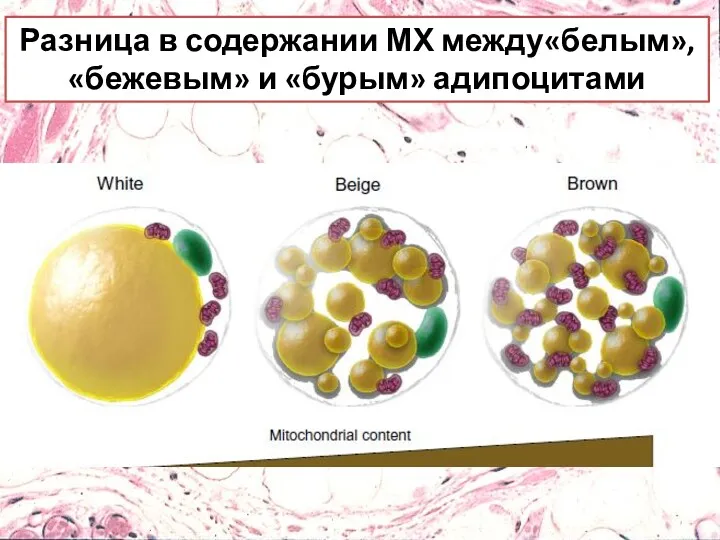
Разница в содержании МХ между«белым», «бежевым» и «бурым» адипоцитами
Разница в содержании МХ между«белым», «бежевым» и «бурым» адипоцитами

В базальном состоянии адипоциты функционируют как адипоциты белой жировой ткани, но
В базальном состоянии адипоциты функционируют как адипоциты белой жировой ткани, но
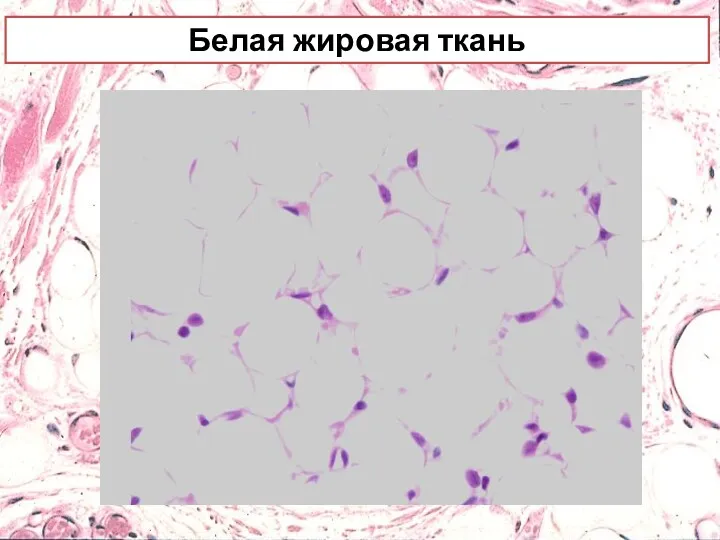
Белая жировая ткань
Белая жировая ткань
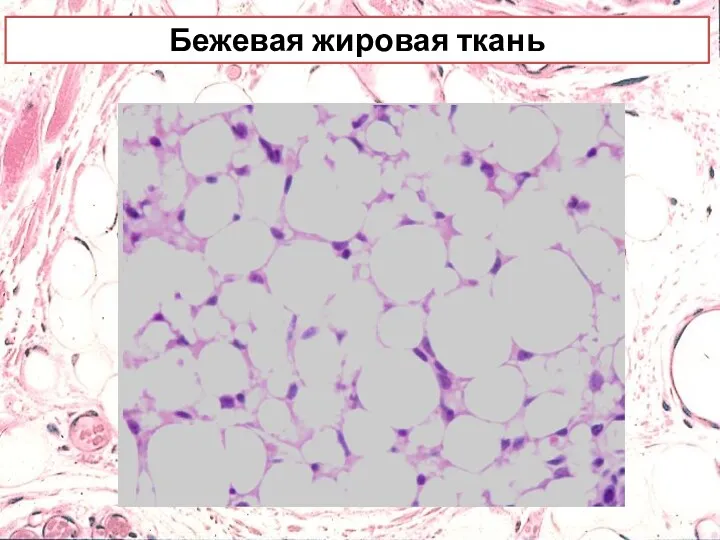
Бежевая жировая ткань
Бежевая жировая ткань
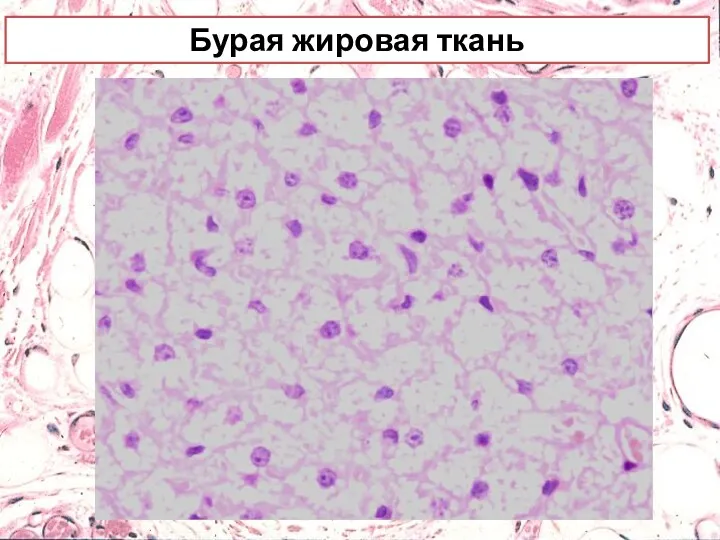
Бурая жировая ткань
Бурая жировая ткань
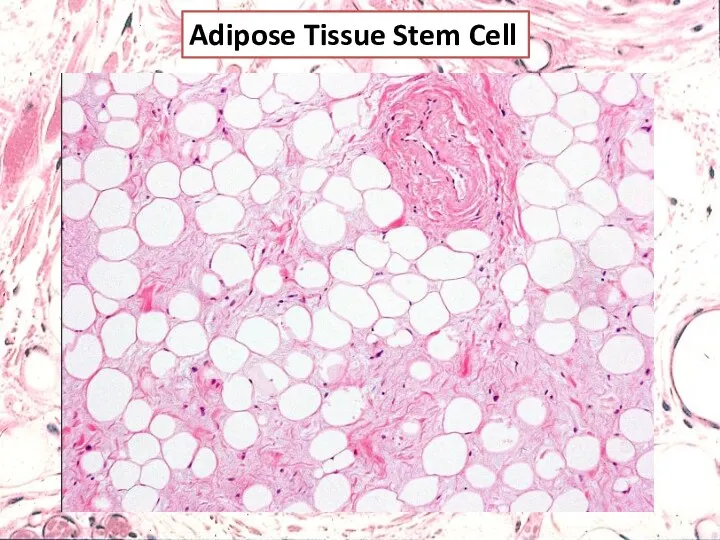
Adipose Tissue Stem Cell
Adipose Tissue Stem Cell

Бежевые адипоциты дифференцируются от клеток-предшественников эмбриональной мезодермы, которые экспрессируют Myf5 и
Бежевые адипоциты дифференцируются от клеток-предшественников эмбриональной мезодермы, которые экспрессируют Myf5 и
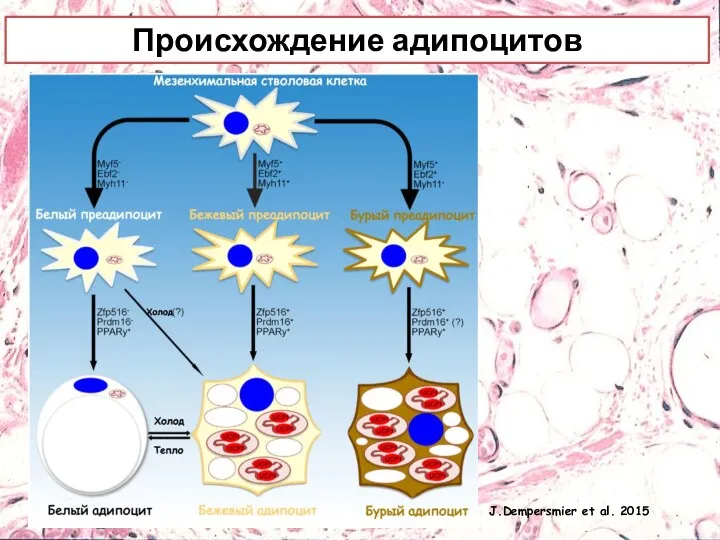
Происхождение адипоцитов
J.Dempersmier et al. 2015
Происхождение адипоцитов
J.Dempersmier et al. 2015

Бурые адипоциты считаются метаболически более эффективными: всего лишь 63 г полностью
Бурые адипоциты считаются метаболически более эффективными: всего лишь 63 г полностью

Тем не менее существует мнение, что эктопическая экспрессия разобщающего белка 1
Тем не менее существует мнение, что эктопическая экспрессия разобщающего белка 1
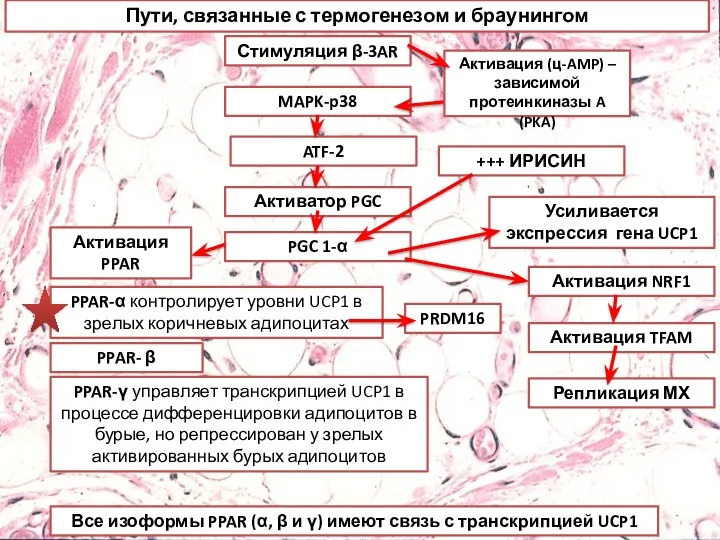
Пути, связанные с термогенезом и браунингом
Стимуляция β-3AR
MAPK-p38
ATF-2
Активатор PGC
PGC 1-α
Активация PPAR
Активация NRF1
Активация
Пути, связанные с термогенезом и браунингом
Стимуляция β-3AR
MAPK-p38
ATF-2
Активатор PGC
PGC 1-α
Активация PPAR
Активация NRF1
Активация
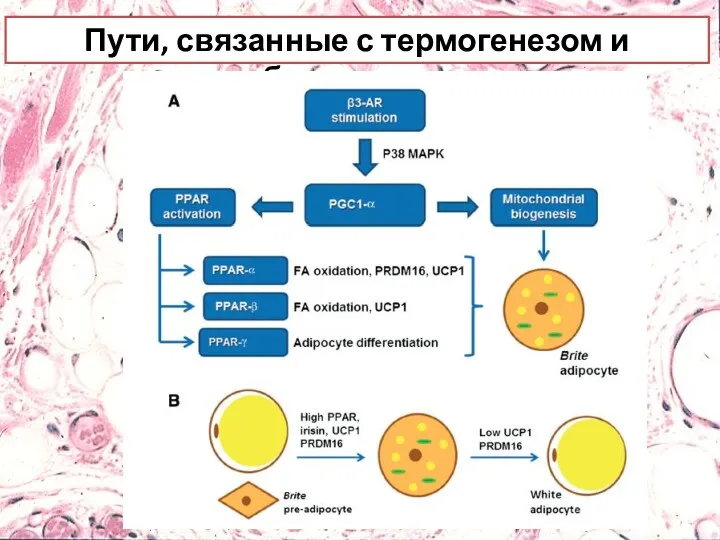
Пути, связанные с термогенезом и браунингом
Пути, связанные с термогенезом и браунингом

Пути, связанные с термогенезом и браунингом.
Стимуляция бета-3-адренергическими рецепторами приводит к индукции
Пути, связанные с термогенезом и браунингом.
Стимуляция бета-3-адренергическими рецепторами приводит к индукции
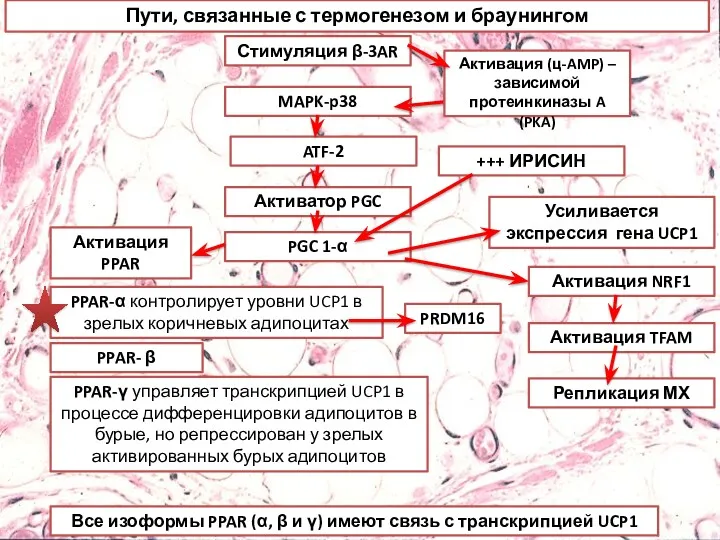
Пути, связанные с термогенезом и браунингом
Стимуляция β-3AR
MAPK-p38
ATF-2
Активатор PGC
PGC 1-α
Активация PPAR
Активация NRF1
Активация
Пути, связанные с термогенезом и браунингом
Стимуляция β-3AR
MAPK-p38
ATF-2
Активатор PGC
PGC 1-α
Активация PPAR
Активация NRF1
Активация

Фармакологическая индукция превращения белой жировой ткани в бурую
Адренергическая стимуляция
5'АМФ-активируемая протеинкиназа-АМФК
FGF21
Фармакологическая индукция превращения белой жировой ткани в бурую
Адренергическая стимуляция
5'АМФ-активируемая протеинкиназа-АМФК
FGF21
Пути, связанные с термогенезом и браунингом
Стимуляция β-3AR
MAPK-p38
ATF-2
Активатор PGC
PGC 1-α
Активация PPAR
Активация NRF1
Активация
Пути, связанные с термогенезом и браунингом
Стимуляция β-3AR
MAPK-p38
ATF-2
Активатор PGC
PGC 1-α
Активация PPAR
Активация NRF1
Активация
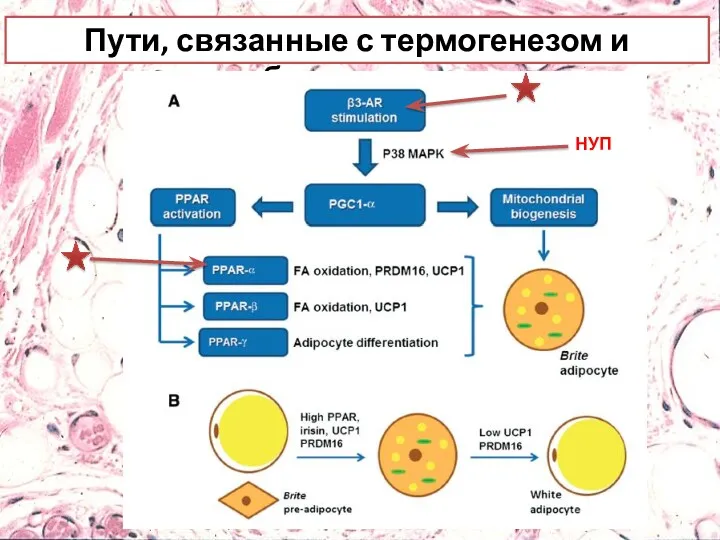
Пути, связанные с термогенезом и браунингом
НУП
Пути, связанные с термогенезом и браунингом
НУП
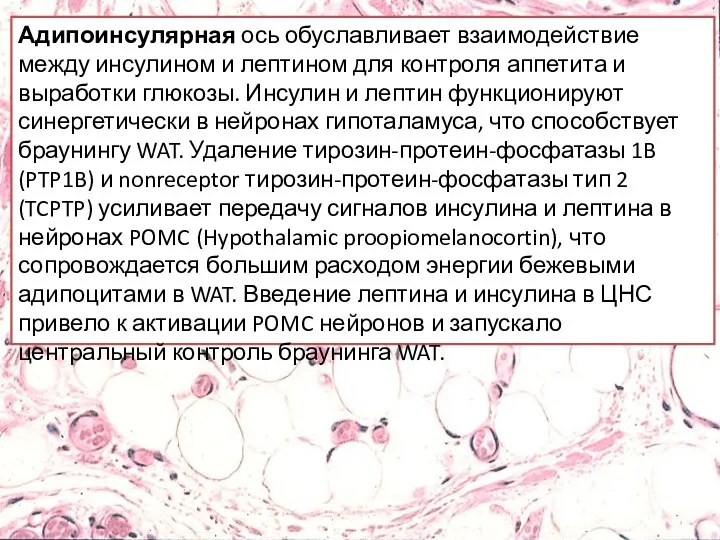
Адипоинсулярная ось обуславливает взаимодействие между инсулином и лептином для контроля аппетита
Адипоинсулярная ось обуславливает взаимодействие между инсулином и лептином для контроля аппетита

Алиментарная индукция превращения белой жировой ткани в бурую
Капсаицин
Ограничение потребления
Алиментарная индукция превращения белой жировой ткани в бурую
Капсаицин
Ограничение потребления
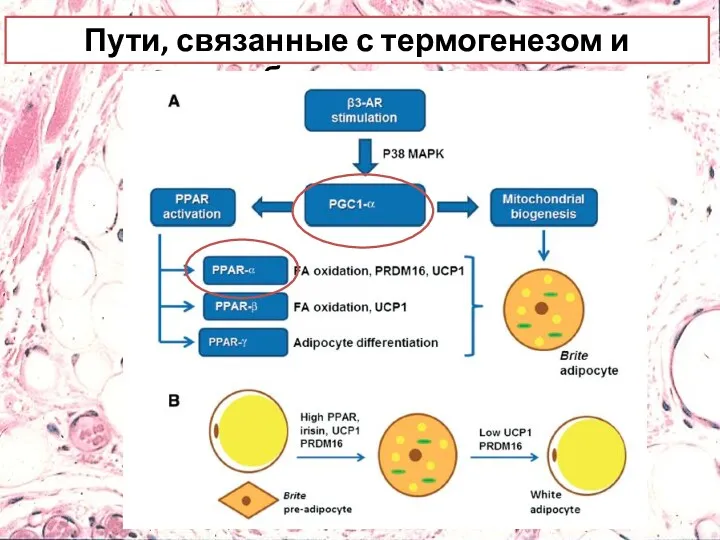
Пути, связанные с термогенезом и браунингом
Пути, связанные с термогенезом и браунингом
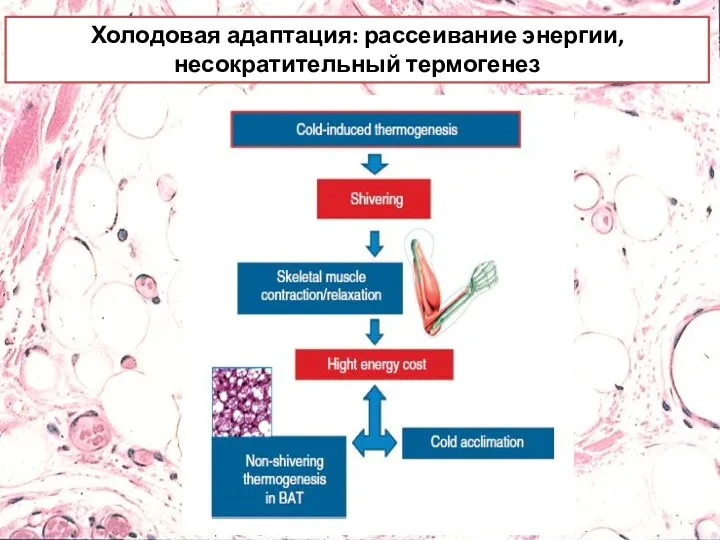
Холодовая адаптация: рассеивание энергии, несократительный термогенез
Холодовая адаптация: рассеивание энергии, несократительный термогенез

Холодовая адаптация: рассеивание энергии, несократительный термогенез.
Производство тепла начинается мгновенно с помощью
Холодовая адаптация: рассеивание энергии, несократительный термогенез.
Производство тепла начинается мгновенно с помощью

Главное:
–– Браунинг это приобретение буроподобного фенотипа адипоцитами белой жировой ткани,
Главное:
–– Браунинг это приобретение буроподобного фенотипа адипоцитами белой жировой ткани,

Главное:
–– капсаицин, желчные кислоты, ресвератрол, ретиноевая кислота и некоторые липиды
Главное:
–– капсаицин, желчные кислоты, ресвератрол, ретиноевая кислота и некоторые липиды
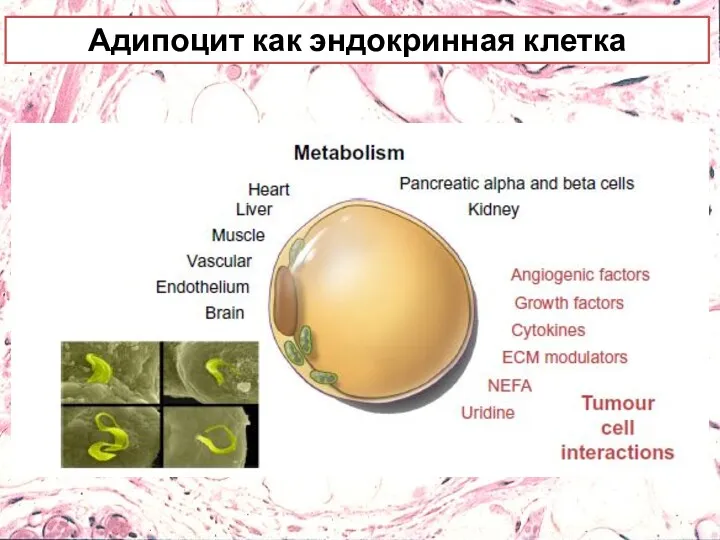
Адипоцит как эндокринная клетка
Адипоцит как эндокринная клетка
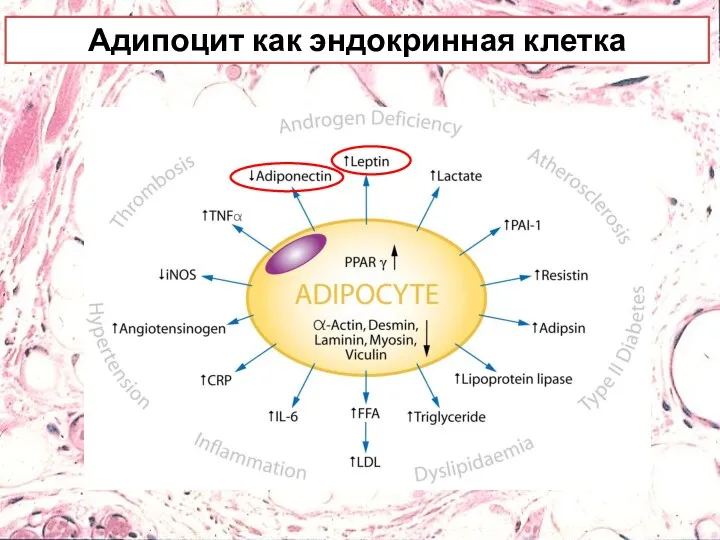
Адипоцит как эндокринная клетка
Адипоцит как эндокринная клетка
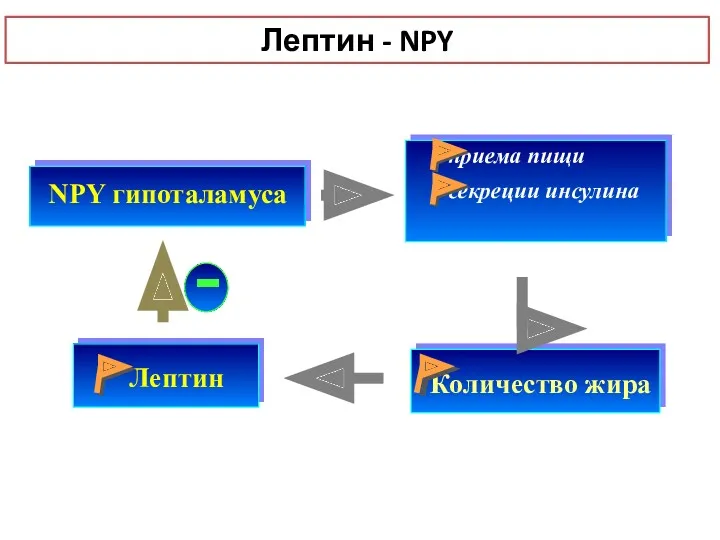
NPY гипоталамуса
Лептин
приема пищи
секреции инсулина
-
Лептин - NPY
NPY гипоталамуса
Лептин
приема пищи
секреции инсулина
-
Лептин - NPY

Лептин - продукт (гормон) жировой ткани (ob ген)
Концентрация прямо пропорциональна степени
Лептин - продукт (гормон) жировой ткани (ob ген)
Концентрация прямо пропорциональна степени
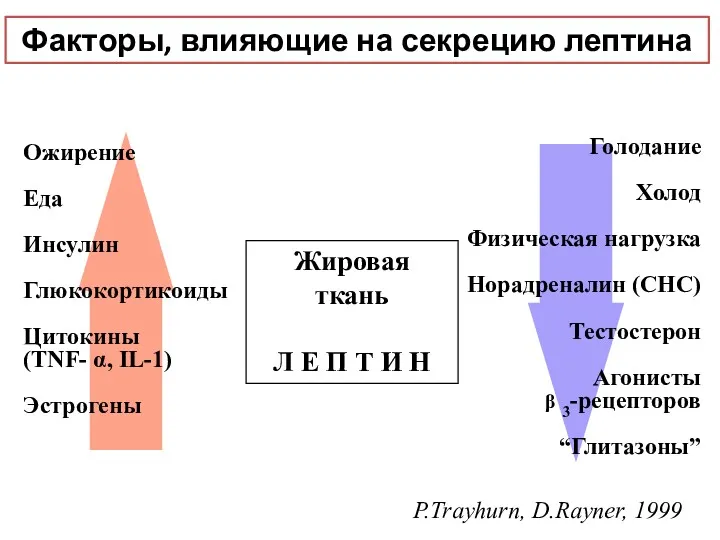
Жировая ткань
Л Е П Т И Н
Ожирение
Еда
Инсулин
Глюкокортикоиды
Цитокины
(TNF- α, IL-1)
Эстрогены
Голодание
Холод
Физическая нагрузка
Норадреналин
Жировая ткань
Л Е П Т И Н
Ожирение
Еда
Инсулин
Глюкокортикоиды
Цитокины
(TNF- α, IL-1)
Эстрогены
Голодание
Холод
Физическая нагрузка
Норадреналин

Адипонектин
универсальный антилипотоксичный агент
имеет обратную связь с ожирением
Оказывает противовоспалительное, инсулин-сенсибилизирующие
Адипонектин
универсальный антилипотоксичный агент
имеет обратную связь с ожирением
Оказывает противовоспалительное, инсулин-сенсибилизирующие
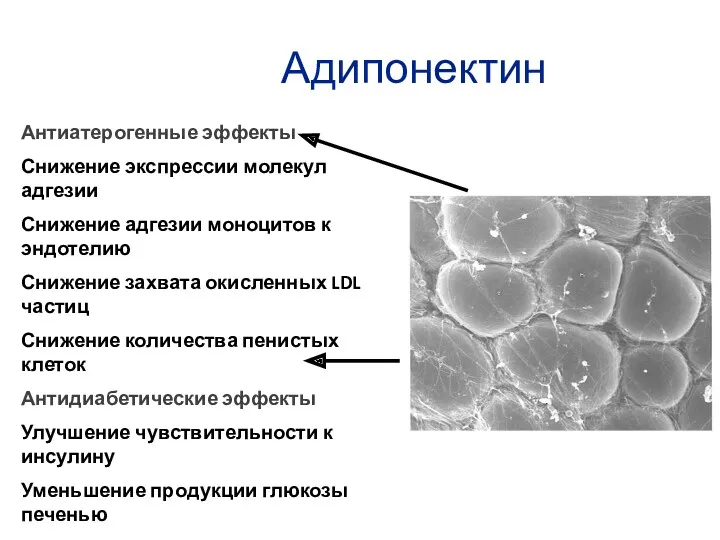
Адипонектин
Антиатерогенные эффекты
Снижение экспрессии молекул адгезии
Снижение адгезии моноцитов к эндотелию
Снижение захвата окисленных
Адипонектин
Антиатерогенные эффекты
Снижение экспрессии молекул адгезии
Снижение адгезии моноцитов к эндотелию
Снижение захвата окисленных

Повышенный уровень церамидов ассоциируется с инсулинорезистентностью, воспалением и апоптозом
Церамиды превращаются в
Повышенный уровень церамидов ассоциируется с инсулинорезистентностью, воспалением и апоптозом
Церамиды превращаются в
Гипотеза «экономного» генотипа
Дополнительные калории и их отложение в виде триглицеридов
Гипотеза «экономного» генотипа
Дополнительные калории и их отложение в виде триглицеридов
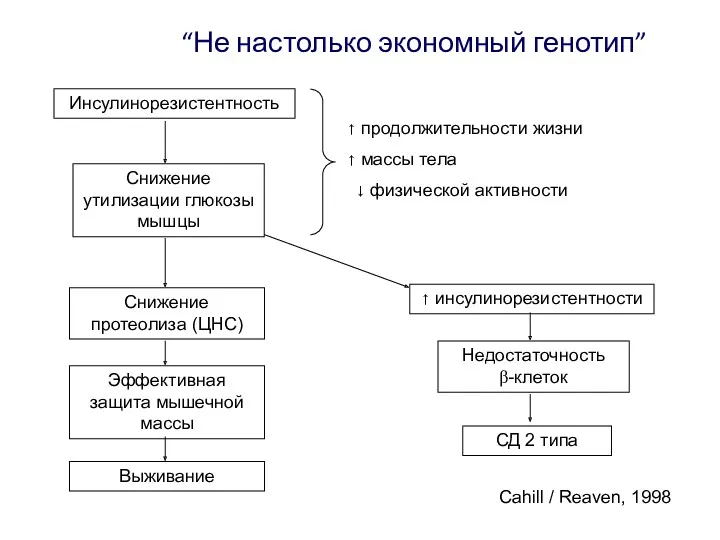
“Не настолько экономный генотип”
Выживание
Эффективная защита мышечной массы
Снижение протеолиза (ЦНС)
Инсулинорезистентность
продолжительности жизни
“Не настолько экономный генотип”
Выживание
Эффективная защита мышечной массы
Снижение протеолиза (ЦНС)
Инсулинорезистентность
продолжительности жизни

Инсулинорезистентность
Гиперинсулинемия
Нарушения толерантности к глюкозе
Гипертриглицеридемия
Снижением ЛПВП и повышение ЛПНП
Артериальная гипертензия
Метаболический синдром
Rеаven, 1988
Гиперинсулинемия
Нарушения толерантности к глюкозе
Гипертриглицеридемия
Снижением ЛПВП и повышение ЛПНП
Артериальная гипертензия
Метаболический синдром
Rеаven, 1988
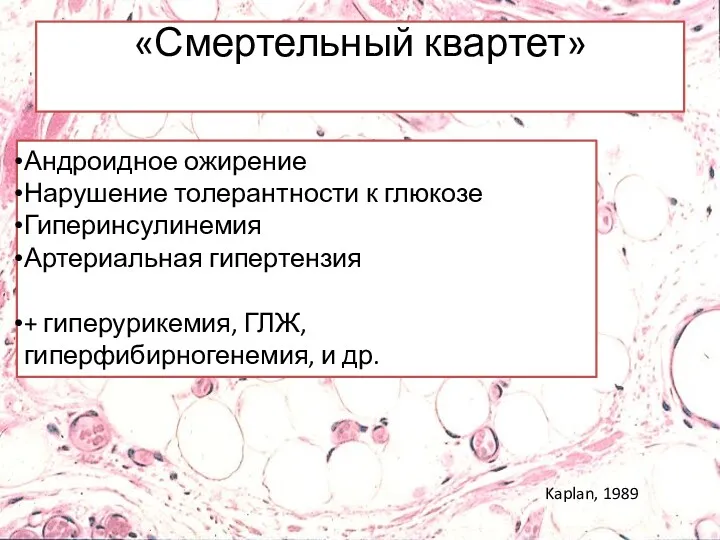
Андроидное ожирение
Нарушение толерантности к глюкозе
Гиперинсулинемия
Артериальная гипертензия
+ гиперурикемия, ГЛЖ, гиперфибирногенемия, и др.
«Смертельный
Андроидное ожирение
Нарушение толерантности к глюкозе
Гиперинсулинемия
Артериальная гипертензия
+ гиперурикемия, ГЛЖ, гиперфибирногенемия, и др.
«Смертельный
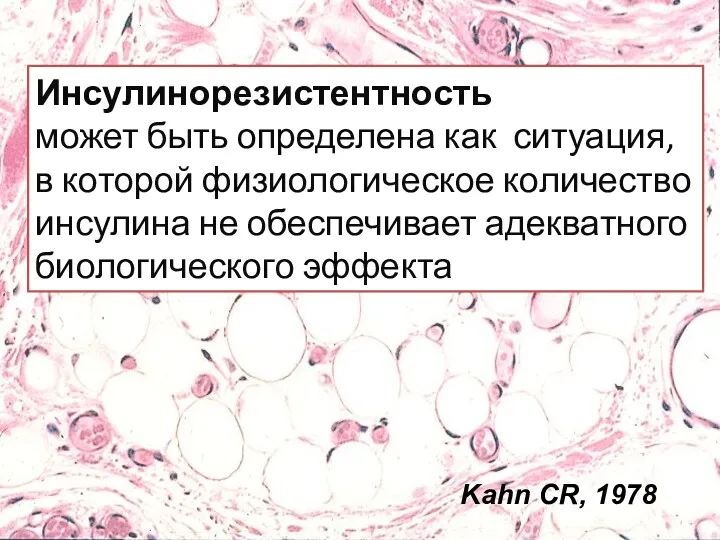
Инсулинорезистентность
может быть определена как ситуация, в которой физиологическое количество инсулина не
Инсулинорезистентность
может быть определена как ситуация, в которой физиологическое количество инсулина не
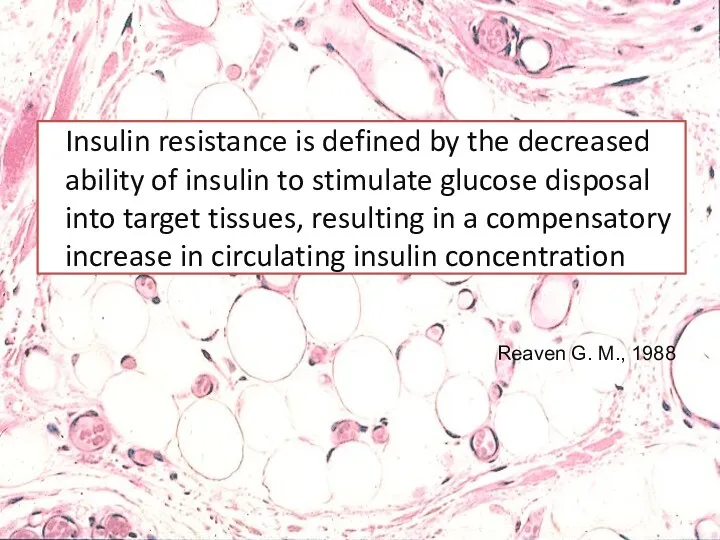
Insulin resistance is defined by the decreased ability of insulin to
Insulin resistance is defined by the decreased ability of insulin to
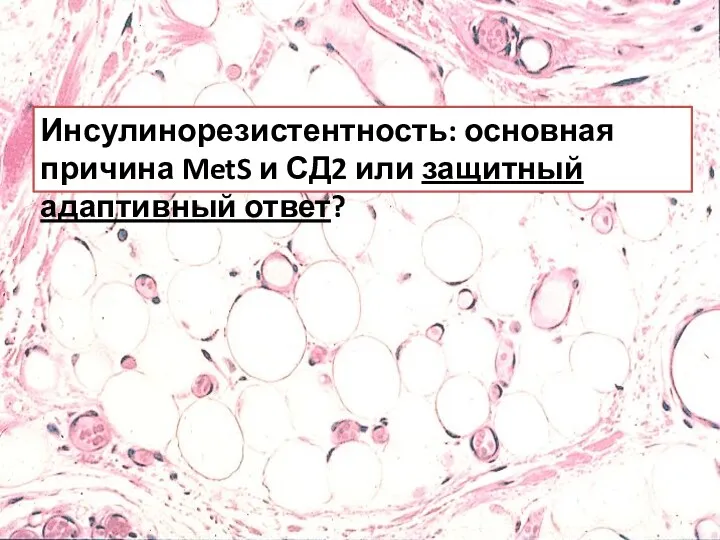
Инсулинорезистентность: основная причина MetS и СД2 или защитный адаптивный ответ?
Инсулинорезистентность: основная причина MetS и СД2 или защитный адаптивный ответ?
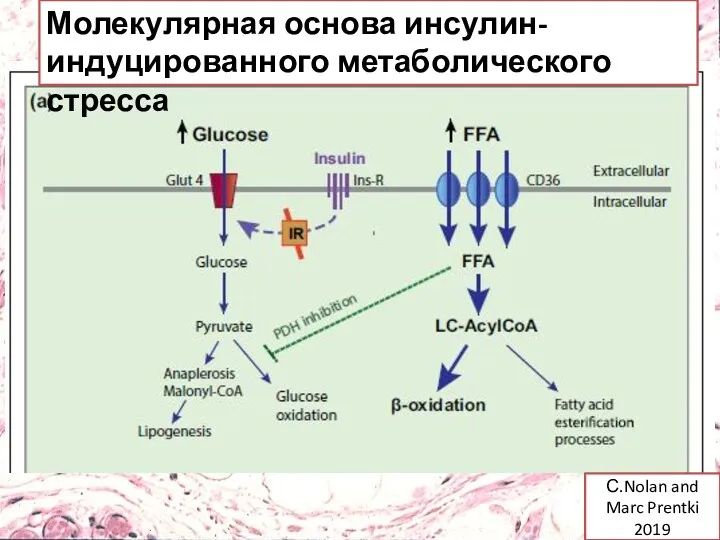
С.Nolan and Marc Prentki 2019
Молекулярная основа инсулин-индуцированного метаболического стресса
С.Nolan and Marc Prentki 2019
Молекулярная основа инсулин-индуцированного метаболического стресса
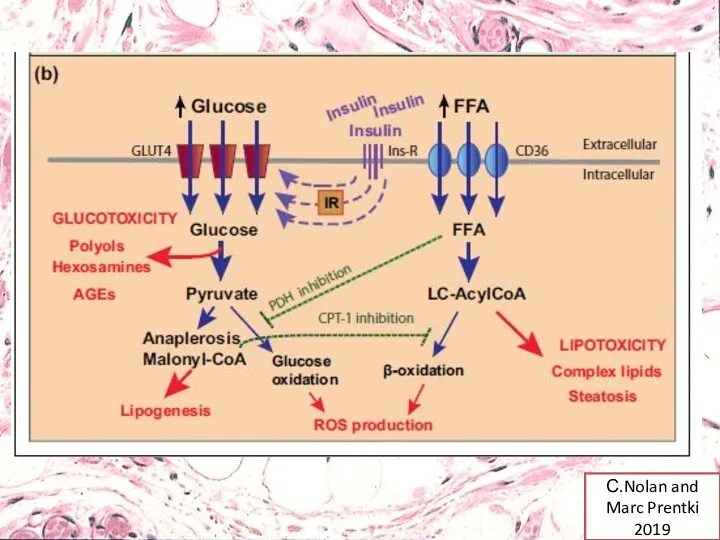
С.Nolan and Marc Prentki 2019
С.Nolan and Marc Prentki 2019
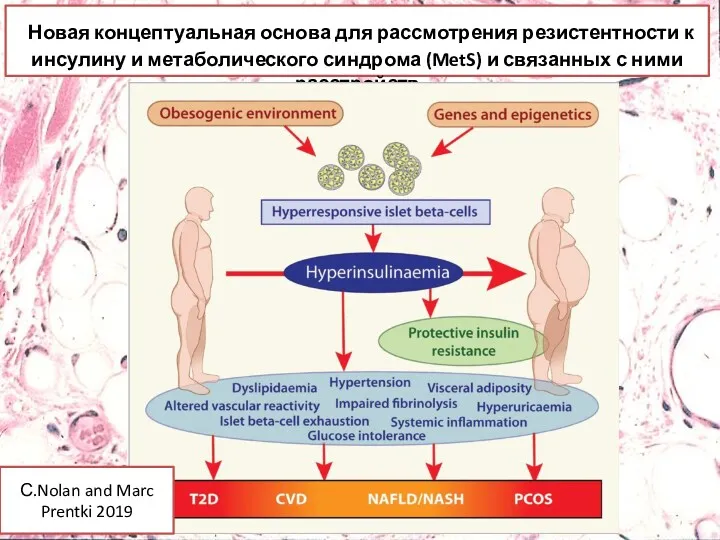
Новая концептуальная основа для рассмотрения резистентности к инсулину и метаболического
Новая концептуальная основа для рассмотрения резистентности к инсулину и метаболического
 Эпидемии и пандемии в Средневековой Европе
Эпидемии и пандемии в Средневековой Европе Мероприятия по снижению риска заражения и распространения новой коронавирусной инфекции 2019-nCoV
Мероприятия по снижению риска заражения и распространения новой коронавирусной инфекции 2019-nCoV Миогимнастика как метод профилактики и лечения деформаций зубочелюстной системы ребенка
Миогимнастика как метод профилактики и лечения деформаций зубочелюстной системы ребенка Клинический случай ХНН
Клинический случай ХНН Средства, действующие на ЦНС
Средства, действующие на ЦНС Виды повязок. Правила наложения бинтовых повязок
Виды повязок. Правила наложения бинтовых повязок Дефект межпредсердной перегородки (ДМПП)
Дефект межпредсердной перегородки (ДМПП) Травма головы, семинар. Оказание помощи пострадавшим с ЧМТ на догоспитальном этапе
Травма головы, семинар. Оказание помощи пострадавшим с ЧМТ на догоспитальном этапе Острые вялые параличи
Острые вялые параличи Теоретичні основи фізичної терапії та ерготерапії
Теоретичні основи фізичної терапії та ерготерапії Первая помощь при вывихах и переломах костей
Первая помощь при вывихах и переломах костей Фармакоекономічний аналіз використання лікарських засобів на прикладі Лікарняна каса Житомирської області
Фармакоекономічний аналіз використання лікарських засобів на прикладі Лікарняна каса Житомирської області Гормональная контрацепция. Взвешенный выбор
Гормональная контрацепция. Взвешенный выбор Эхокардиография, ультразвуковая кардиография
Эхокардиография, ультразвуковая кардиография Питание ребёнка первого года жизни
Питание ребёнка первого года жизни Эпилепсия. Классификация эпилепсии. Диагностические критерии. Неотложная помощь. Принципы лечения
Эпилепсия. Классификация эпилепсии. Диагностические критерии. Неотложная помощь. Принципы лечения Медичне страхування
Медичне страхування Неврологические осложнения остеохондроза позвоночника
Неврологические осложнения остеохондроза позвоночника Денсаулықпен байланысты өмір сүрудің сапасы
Денсаулықпен байланысты өмір сүрудің сапасы Задачи, организационная структура и органы управления Всероссийской службы медицины катастроф
Задачи, организационная структура и органы управления Всероссийской службы медицины катастроф Профилактика и лечение болезни Альцгеймера
Профилактика и лечение болезни Альцгеймера Дизентерия
Дизентерия Осторожно! Клещи
Осторожно! Клещи Современная система контроля и обеспечения качества лекарственных средств в Украине
Современная система контроля и обеспечения качества лекарственных средств в Украине Негізгі психопатологиялық синдромдар
Негізгі психопатологиялық синдромдар Болезни органов брюшной полости. Болезни органов ротовой полости
Болезни органов брюшной полости. Болезни органов ротовой полости Эндокринные заболевания и беременность
Эндокринные заболевания и беременность Рахит. Этиопатогенез рахита
Рахит. Этиопатогенез рахита