- Соматический гипермутагенез. Клональная селекция лимфоцитов

Содержание
- 2. Клональная селекция лимфоцитов Из клетки-предшественника развивается большое количество лимфоцитов, каждый - со своей "случайной" специфичностью Аутореактивные
- 3. Развитие эффекторных В лимфоцитов Костный мозг (эмбриональная печень) Селезенка Формирование репертуара специфичностей лимфоцитарных рецепторов происходит в
- 4. Постулаты теории клональной селекции Каждый лимфоцит несет рецептор единственного типа и уникальной специфичности. Высокоаффинное взаимодействие рецептора
- 5. Антигенраспознающие рецепторы лимфоцитов (структурная схема) Вариабельный домен (распознавание антигена) Константная часть (эффекторные функции) H H L
- 6. VL VL CL CL VH VH CH1 CH1 CH2 CH2 CH3 CH3 CH3 Vα Vβ Cα
- 7. Thy-1 (CD90) BCR TCR MHC-I MHC-II СУПЕРСЕМЕЙСТВО ИММУНОГЛОБУЛИНОВ включает в себя не только антитела CD3 CD2
- 8. VL CL VH CH1 CH2 CH2 CH3 CH3 CL VL CH1 VH 3D структура антитела (на
- 9. Механизмы возникновения мутаций в V-цепях BCR
- 10. При созревании В-лимфоцитов имеет место перегруппировка ДНК, приводящая к переносу одного из V-генов к одному из
- 11. Схема перестройки генов иммуноглобулинов Легкая цепь Тяжелая цепь
- 12. Разнообразие ИГ(И) достигается перегруппировкой в ДНК. В геноме человека находятся три независимых локуса генов ИГ. Каждый
- 13. Схема геномного устройства локусов иммуноглобулинов
- 14. Классы иммуноглобулинов Человек: IgM, IgD, IgG1, IgG2a, IgG2b, IgG3, IgE, IgA1, IgA2 Дисульфидные связи Углеводы Изотип
- 15. Вклад различных участков V-доменов Н- и L-цепей в построение антигенсвязывающего центра антител Размерность антигенсвязывающего участка: длина
- 16. Распознавание антигенов Т-клетками αβ Т-клетки узнают линейные пептидные эпитопы, которые могут находиться в любой части молекулы
- 17. ПОСЛЕДОВАТЕЛЬНОСТЬ ПЕРЕСТРОЙКИ V-ГЕНОВ, Перестройка V- гена на одной хромосоме Успешная перестройка (33%) Формирование «зрелого» V-гена. Неуспешная
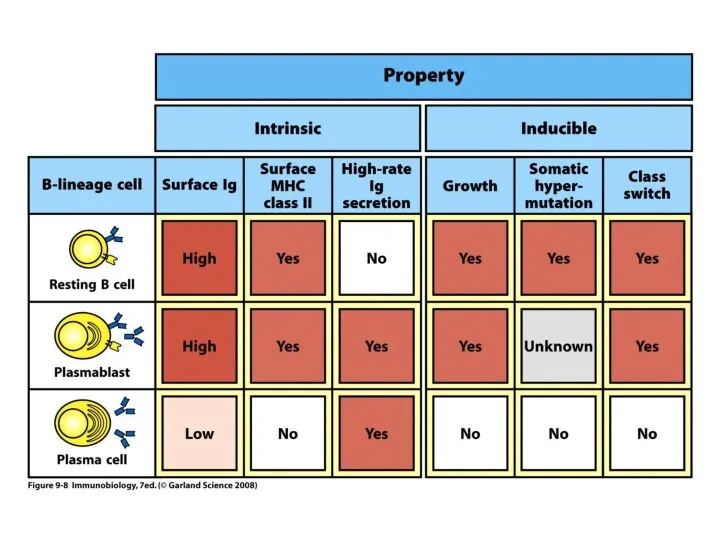
- 20. Центр размножения Особая область в лимфатических узлах или селезенке, в которой обнаруживается интенсивная пролиферация (деление) и
- 21. Рециркулирующие В – клетки встречаются со «ИСКОМЫМ» антигеном в лимфоузле
- 22. Зародышевые центры – ЗЦ (дифференцировка и селекция В-лимфоцитов) ТЕМНАЯ ЗОНА: В-л связавшие АГ и получившие дополнительный
- 23. роль Tfh (фолликулярных Т хелперов) На территории лимфоидного фолликула в ЛУ под контролем Tfh происходит :
- 24. Соматический гипермутагенез и переключение изотипов Герминальный центр (в селезенке или в лимфоузле)
- 25. Контроль аффинитета связи АГ с АТ Только эта клетка, чей рецептор в результате соматических гипермутаций приобрел
- 26. Отбор В-лимфоцитов с высокоаффинными рецепторами в лимфоузлах
- 27. Взаимодействие В-лимфоцитов с высокоаффинными рецепторами с фолликулярными Т- хелперами (Tfn) в лимфоузлах Долгоживущая плазматическая клетка памяти
- 28. Взаимодействие В-клеток с микроокружением Итог: ОТБОР КЛЕТОК АНТИТЕЛОПРОДУЦЕНТОВ 1. гибель клеток несущих низкоафинный рецептор ИГ или
- 29. Роль цитокинов в гуморальном иммунитете
- 30. Этапы иммунного ответа : остаются клетки памяти, их популяция не является однородной распознавание антигена клональная экспансия
- 32. Скачать презентацию
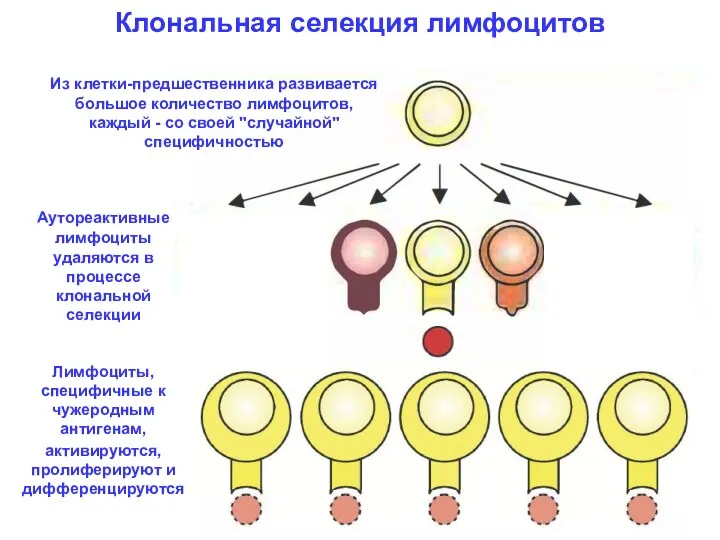
Клональная селекция лимфоцитов
Из клетки-предшественника развивается большое количество лимфоцитов, каждый - со
Клональная селекция лимфоцитов
Из клетки-предшественника развивается большое количество лимфоцитов, каждый - со
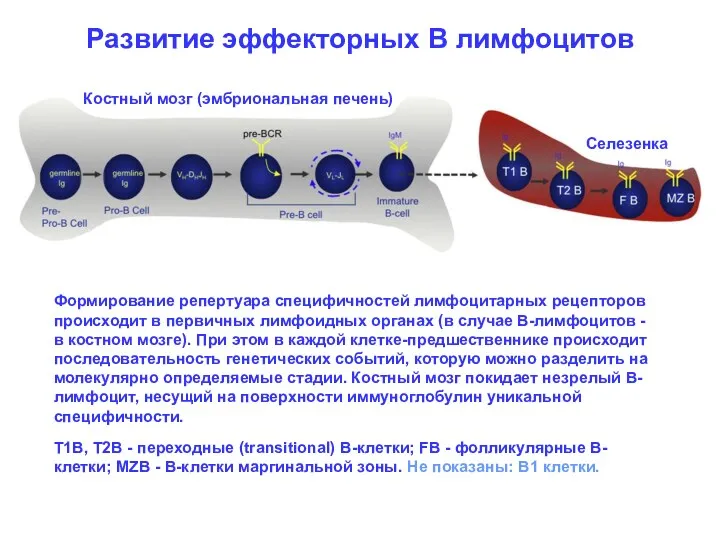
Развитие эффекторных В лимфоцитов
Костный мозг (эмбриональная печень)
Селезенка
Формирование репертуара специфичностей лимфоцитарных рецепторов
Развитие эффекторных В лимфоцитов
Костный мозг (эмбриональная печень)
Селезенка
Формирование репертуара специфичностей лимфоцитарных рецепторов

Постулаты теории клональной селекции
Каждый лимфоцит несет рецептор единственного типа и
Постулаты теории клональной селекции
Каждый лимфоцит несет рецептор единственного типа и
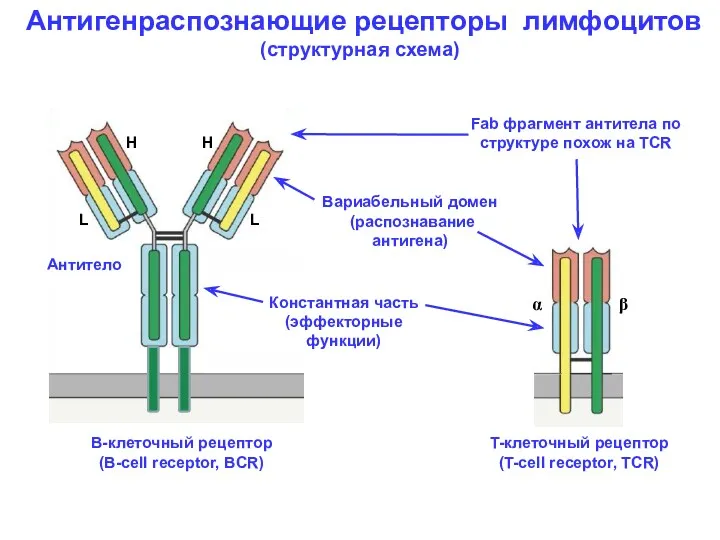
Антигенраспознающие рецепторы лимфоцитов
(структурная схема)
Вариабельный домен
(распознавание
антигена)
Константная часть
(эффекторные
функции)
H
H
L
L
В-клеточный
Антигенраспознающие рецепторы лимфоцитов
(структурная схема)
Вариабельный домен
(распознавание
антигена)
Константная часть
(эффекторные
функции)
H
H
L
L
В-клеточный
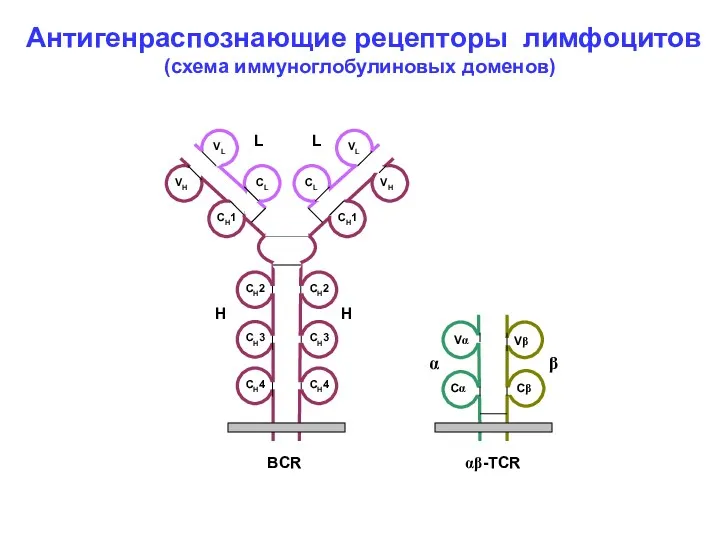
VL
VL
CL
CL
VH
VH
CH1
CH1
CH2
CH2
VL
VL
CL
CL
VH
VH
CH1
CH1
CH2
CH2
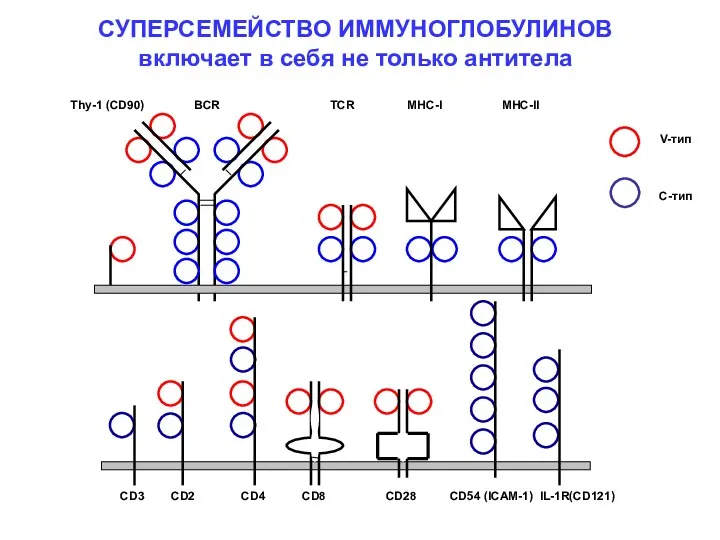
Thy-1 (CD90) BCR TCR MHC-I MHC-II
СУПЕРСЕМЕЙСТВО ИММУНОГЛОБУЛИНОВ
включает в себя не
Thy-1 (CD90) BCR TCR MHC-I MHC-II
СУПЕРСЕМЕЙСТВО ИММУНОГЛОБУЛИНОВ включает в себя не
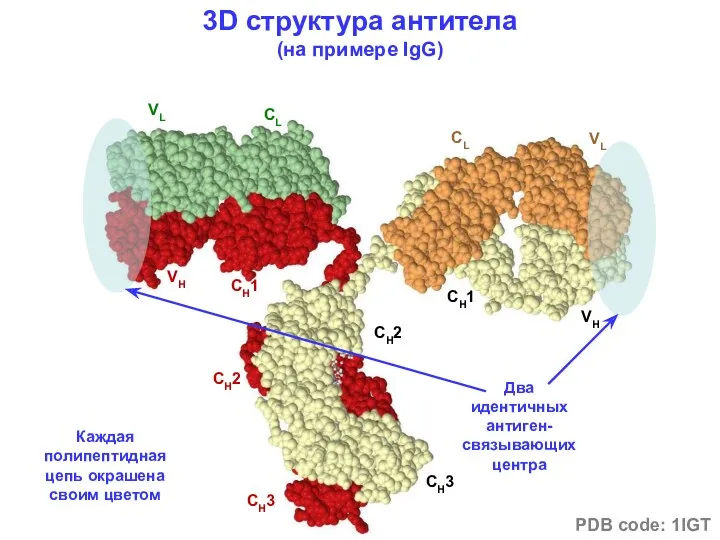
VL
CL
VH
CH1
CH2
CH2
CH3
CH3
CL
VL
CH1
VL
CL
VH
CH1
CH2
CH2
CH3
CH3
CL
VL
CH1

Механизмы возникновения мутаций в V-цепях BCR
Механизмы возникновения мутаций в V-цепях BCR
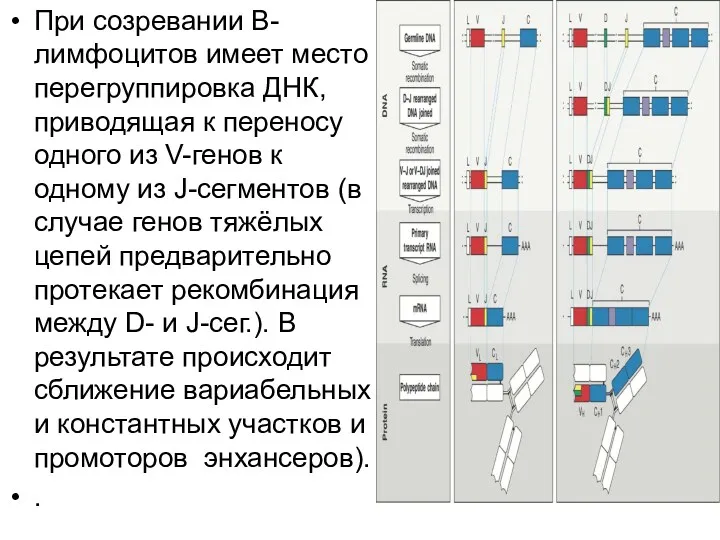
При созревании В-лимфоцитов имеет место перегруппировка ДНК, приводящая к переносу одного
При созревании В-лимфоцитов имеет место перегруппировка ДНК, приводящая к переносу одного
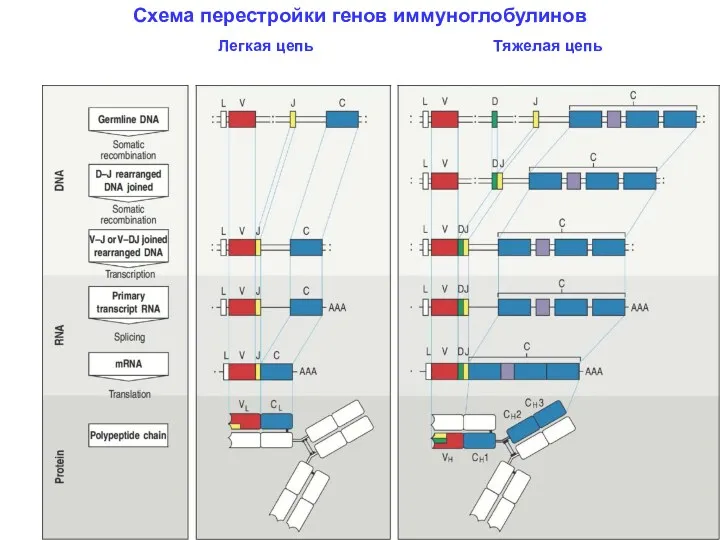
Схема перестройки генов иммуноглобулинов
Легкая цепь
Тяжелая цепь
Схема перестройки генов иммуноглобулинов
Легкая цепь
Тяжелая цепь

Разнообразие ИГ(И) достигается перегруппировкой в ДНК. В геноме человека находятся три
Разнообразие ИГ(И) достигается перегруппировкой в ДНК. В геноме человека находятся три
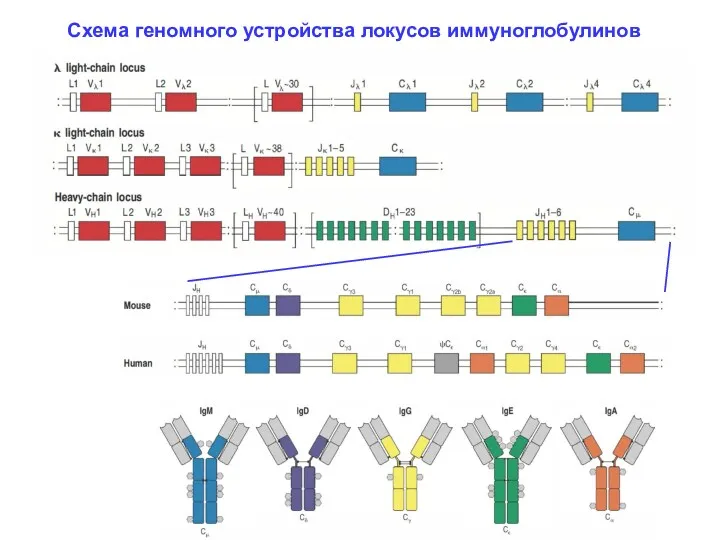
Схема геномного устройства локусов иммуноглобулинов
Схема геномного устройства локусов иммуноглобулинов
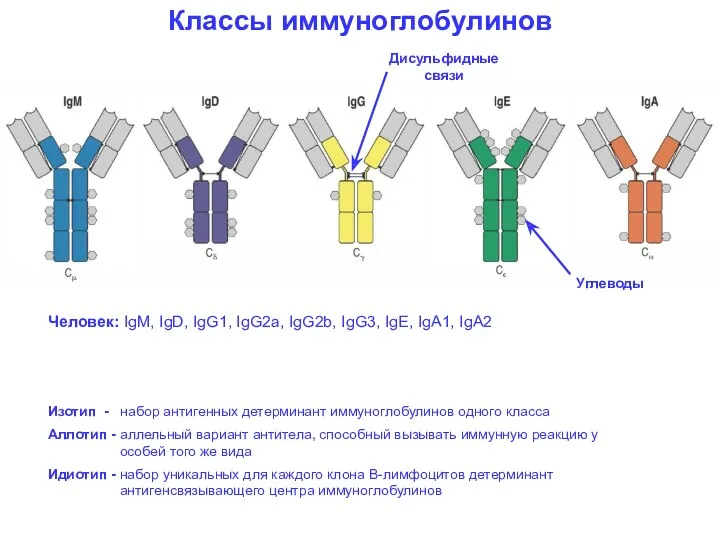
Классы иммуноглобулинов
Человек: IgM, IgD, IgG1, IgG2a, IgG2b, IgG3, IgE, IgA1,
Классы иммуноглобулинов
Человек: IgM, IgD, IgG1, IgG2a, IgG2b, IgG3, IgE, IgA1,
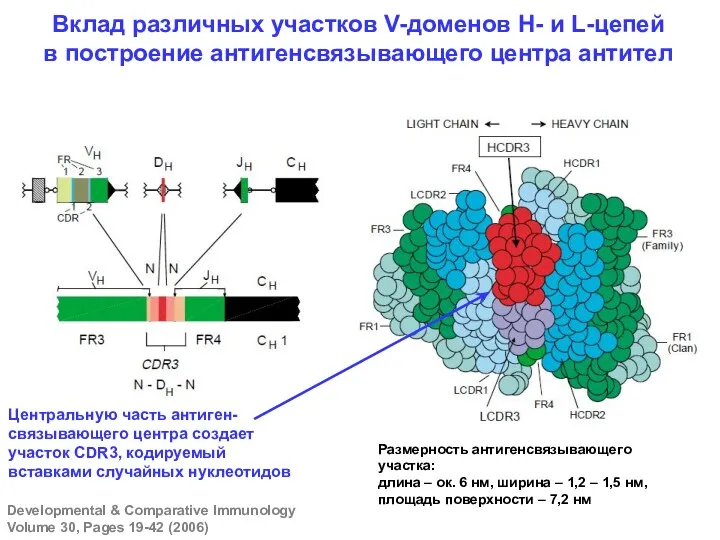
Вклад различных участков V-доменов Н- и L-цепей в построение антигенсвязывающего центра
Вклад различных участков V-доменов Н- и L-цепей в построение антигенсвязывающего центра
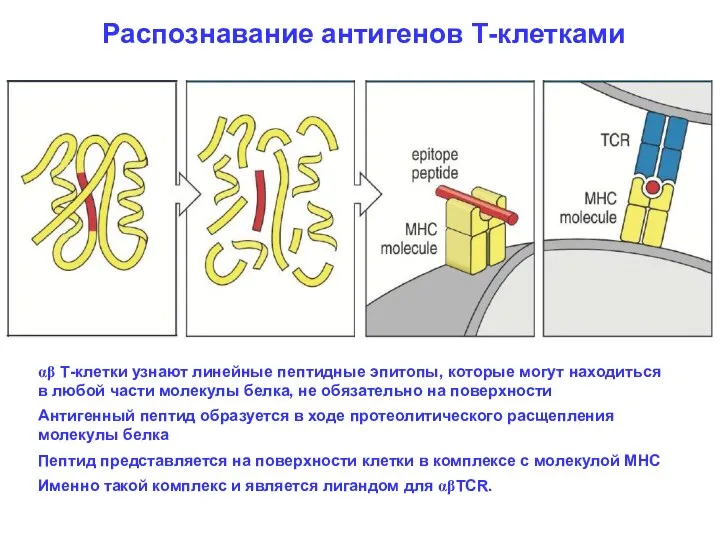
Распознавание антигенов Т-клетками
αβ Т-клетки узнают линейные пептидные эпитопы, которые могут
Распознавание антигенов Т-клетками
αβ Т-клетки узнают линейные пептидные эпитопы, которые могут
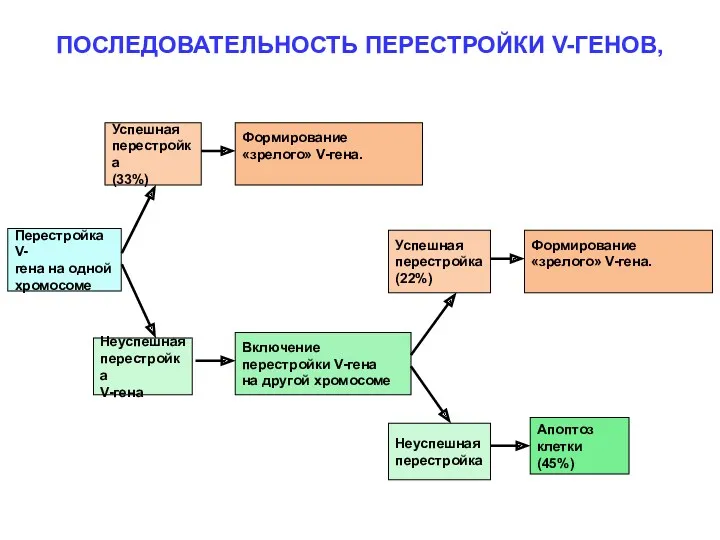
ПОСЛЕДОВАТЕЛЬНОСТЬ ПЕРЕСТРОЙКИ V-ГЕНОВ,
Перестройка V-
гена на одной
хромосоме
Успешная
перестройка
(33%)
Формирование
«зрелого» V-гена.
ПОСЛЕДОВАТЕЛЬНОСТЬ ПЕРЕСТРОЙКИ V-ГЕНОВ,
Перестройка V-
гена на одной
хромосоме
Успешная
перестройка
(33%)
Формирование
«зрелого» V-гена.


Центр размножения
Особая область в лимфатических узлах или селезенке, в которой обнаруживается
Центр размножения
Особая область в лимфатических узлах или селезенке, в которой обнаруживается
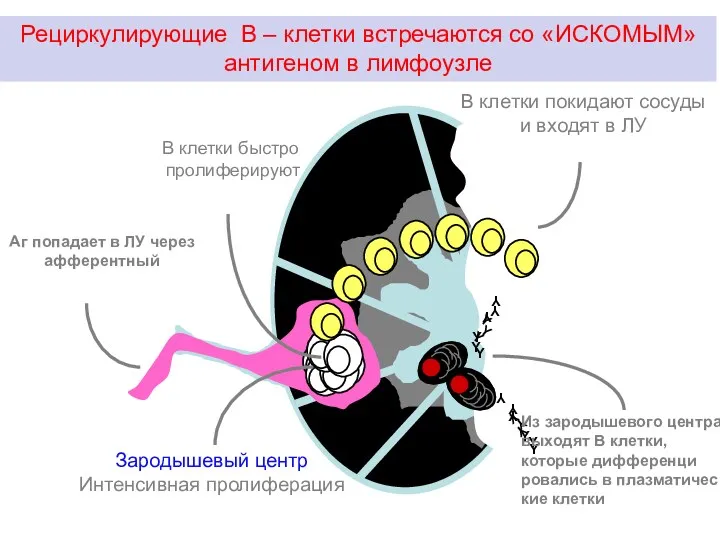
Рециркулирующие В – клетки встречаются со «ИСКОМЫМ» антигеном в лимфоузле
Рециркулирующие В – клетки встречаются со «ИСКОМЫМ» антигеном в лимфоузле
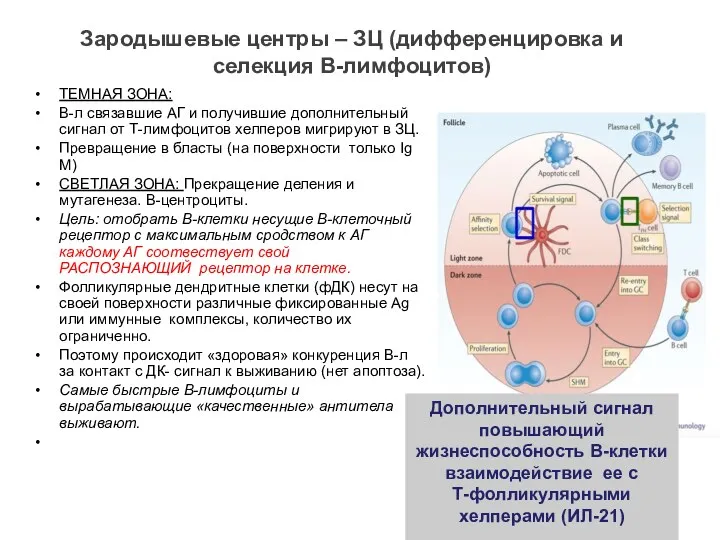
Зародышевые центры – ЗЦ (дифференцировка и селекция В-лимфоцитов)
ТЕМНАЯ ЗОНА:
В-л связавшие АГ
Зародышевые центры – ЗЦ (дифференцировка и селекция В-лимфоцитов)
ТЕМНАЯ ЗОНА:
В-л связавшие АГ
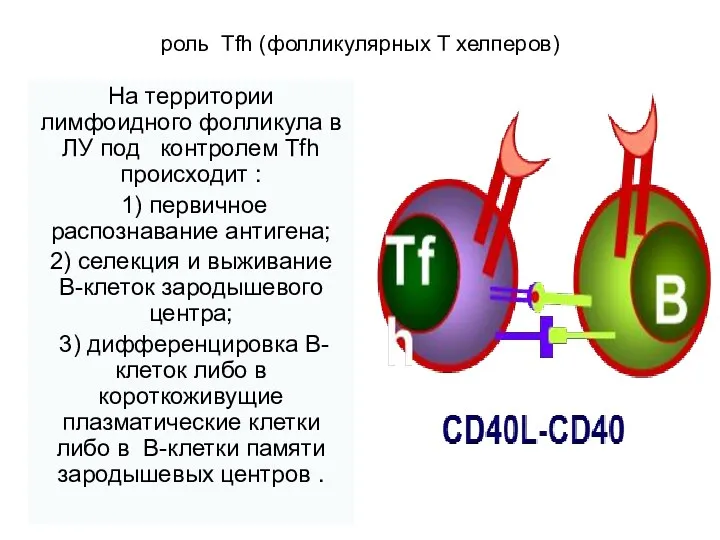
роль Tfh (фолликулярных Т хелперов)
На территории лимфоидного фолликула в
роль Tfh (фолликулярных Т хелперов)
На территории лимфоидного фолликула в

Соматический гипермутагенез и переключение изотипов
Герминальный центр
(в селезенке или в лимфоузле)
Соматический гипермутагенез и переключение изотипов
Герминальный центр
(в селезенке или в лимфоузле)
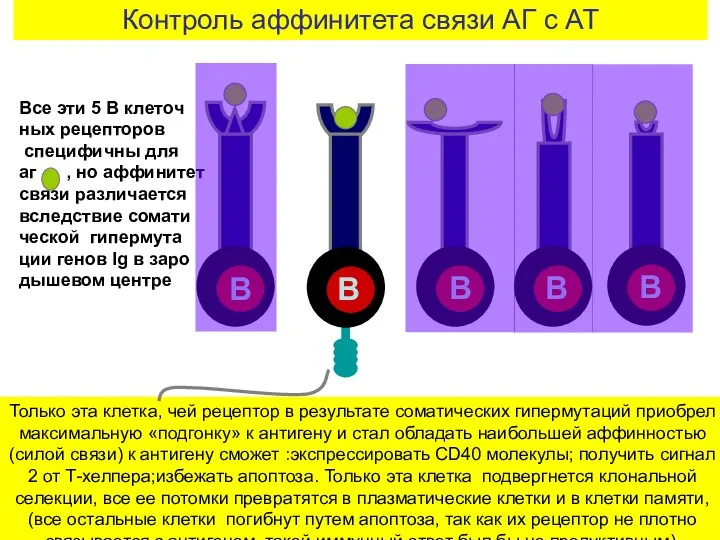
Контроль аффинитета связи АГ с АТ
Только эта клетка, чей рецептор в
Контроль аффинитета связи АГ с АТ
Только эта клетка, чей рецептор в
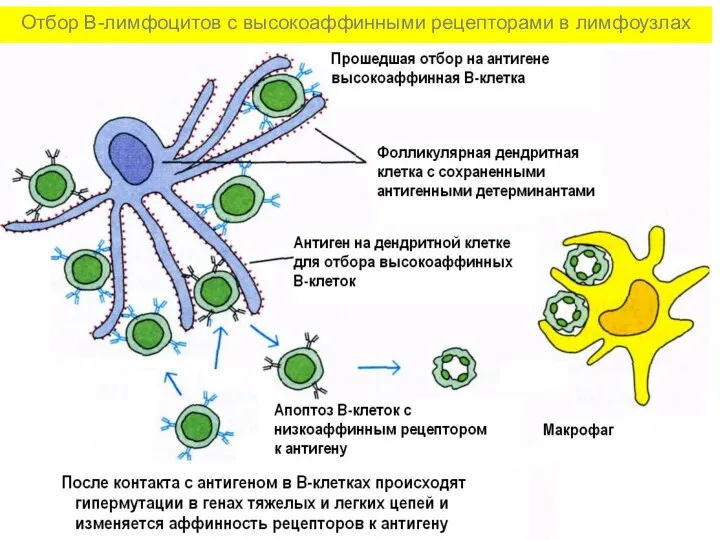
Отбор В-лимфоцитов с высокоаффинными рецепторами в лимфоузлах
Отбор В-лимфоцитов с высокоаффинными рецепторами в лимфоузлах
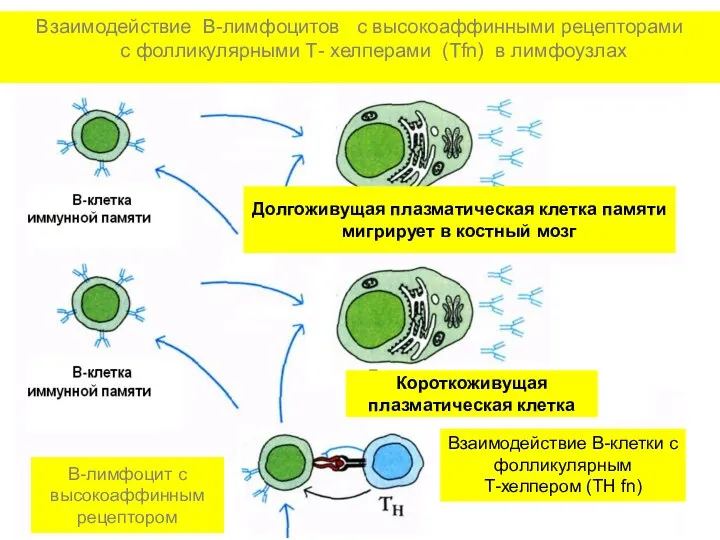
Взаимодействие В-лимфоцитов с высокоаффинными рецепторами с фолликулярными Т- хелперами (Tfn) в
Взаимодействие В-лимфоцитов с высокоаффинными рецепторами с фолликулярными Т- хелперами (Tfn) в
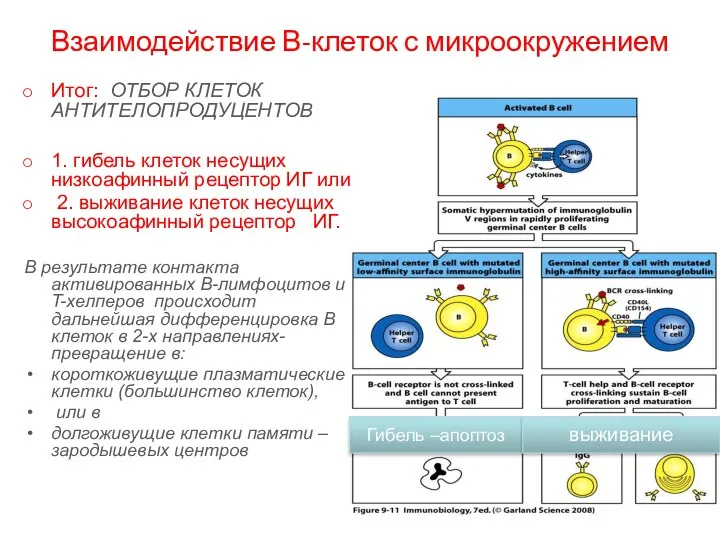
Взаимодействие В-клеток с микроокружением
Итог: ОТБОР КЛЕТОК АНТИТЕЛОПРОДУЦЕНТОВ
1. гибель клеток несущих низкоафинный
Взаимодействие В-клеток с микроокружением
Итог: ОТБОР КЛЕТОК АНТИТЕЛОПРОДУЦЕНТОВ
1. гибель клеток несущих низкоафинный
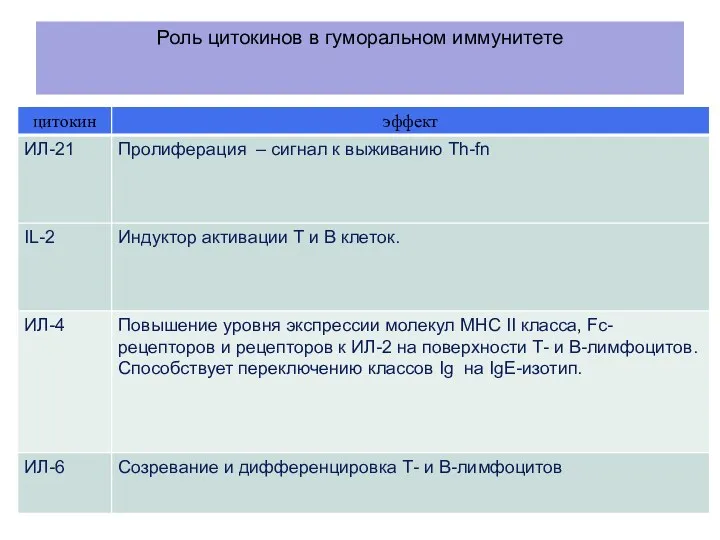
Роль цитокинов в гуморальном иммунитете
Роль цитокинов в гуморальном иммунитете
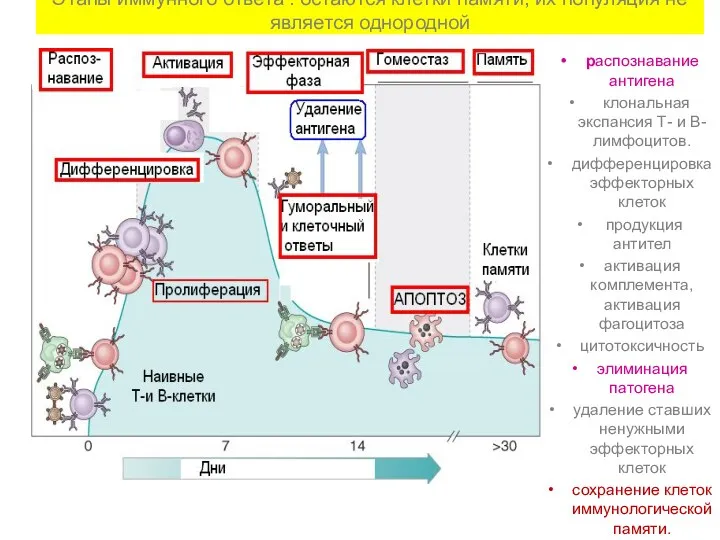
Этапы иммунного ответа : остаются клетки памяти, их популяция не является
Этапы иммунного ответа : остаются клетки памяти, их популяция не является
 Профилактика и лечение наследственных заболеваний
Профилактика и лечение наследственных заболеваний Наркомания. Кодировка различных состояний, вызываемых злоупотреблением ПАВ (МКБ-10, 4-й знак)
Наркомания. Кодировка различных состояний, вызываемых злоупотреблением ПАВ (МКБ-10, 4-й знак) Лечение открытых переломов
Лечение открытых переломов Полигенді аурулар
Полигенді аурулар Гломерулонефриты. Этиопатогенез, клиника, диагностика, лечение
Гломерулонефриты. Этиопатогенез, клиника, диагностика, лечение Медициналық құралдардың қауіпсіздігі мен сенімділігі
Медициналық құралдардың қауіпсіздігі мен сенімділігі Врожденные пороки развития центральной нервной системы
Врожденные пороки развития центральной нервной системы Андрогенодефицит и возможности заместительной гормональной терапии у мужчин
Андрогенодефицит и возможности заместительной гормональной терапии у мужчин Системные васкулиты
Системные васкулиты Особенности эндодонтической обработки временных и постоянных зубов с несформированным корнем
Особенности эндодонтической обработки временных и постоянных зубов с несформированным корнем Анатомия, гистология, физиология кожи. Методика обследования кожных больных
Анатомия, гистология, физиология кожи. Методика обследования кожных больных Неправильные положения плода
Неправильные положения плода Острый гломерулонефрит
Острый гломерулонефрит Кровообращение плода
Кровообращение плода Оказание первой помощи при жизнеугрожающих состояниях. СОЦМК
Оказание первой помощи при жизнеугрожающих состояниях. СОЦМК Травма грудной клетки
Травма грудной клетки Артериальная гипертензия при беременности
Артериальная гипертензия при беременности Радиационная гигиена
Радиационная гигиена Артрология. Соединения костей
Артрология. Соединения костей Ключевые моменты диализа. Стандарты компании Б Браун
Ключевые моменты диализа. Стандарты компании Б Браун Артроскопия крупных суставов
Артроскопия крупных суставов Ветеринарная фармакология. Общая рецептура
Ветеринарная фармакология. Общая рецептура Диагностика беременности
Диагностика беременности Невідкладна домедична допомога. Основи здоров`я. 9 клас
Невідкладна домедична допомога. Основи здоров`я. 9 клас Иммунодефицитные состояния. Аутоиммунные процессы
Иммунодефицитные состояния. Аутоиммунные процессы Проведение санитарно-противоэпидемических мероприятий при работе в отделениях хирургического профиля и операционного блока
Проведение санитарно-противоэпидемических мероприятий при работе в отделениях хирургического профиля и операционного блока Спинномозговые нервы
Спинномозговые нервы Холера. Этиология
Холера. Этиология